Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
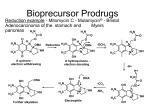
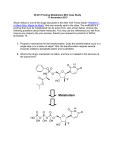
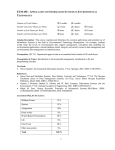
Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Molecular Cancer Therapeutics Small Molecule Therapeutics Bisphosphonamidate Clodronate Prodrug Exhibits Selective Cytotoxic Activity against Melanoma Cell Lines Marie R. Webster1,4, Chandrashekhar Kamat2, Nick Connis2, Ming Zhao2,3, Ashani T. Weeraratna4, Michelle A. Rudek2,3, Christine L. Hann2, and Caren L. Freel Meyers1 Abstract Bisphosphonates are used clinically to treat disorders of calcium metabolism and malignant bone disease and are known to inhibit cancer cell growth, adhesion, and invasion. However, clinical use of these agents for the treatment of extraskeletal disease is limited because of low cell permeability. We recently described a bisphosphonamidate prodrug strategy for efficient intracellular release of bisphosphonates, including clodronate (CLO), in non–small cell lung cancer cells. To evaluate anticancer activity of this prodrug class across many cancer cell types, the bisphosphonamidate clodronate prodrug (CLO prodrug) was screened against the NCI-60 cell line panel, and was found to exhibit selectivity toward melanoma cell lines. Here, we confirm efficient cellular uptake and intracellular activation of this prodrug class in melanoma cells. We further demonstrate inhibition of melanoma cell proliferation, induction of apoptosis, and an antitumor effect of CLO prodrug in a xenograft model. These data suggest a novel therapeutic application for the CLO prodrug and potential to selectively target melanoma cells. Mol Cancer Ther; 13(2); 297–306. 2013 AACR. Introduction Bisphosphonates are stable analogs of pyrophosphate bearing a P-C-P linkage, and are used clinically to treat disorders of calcium metabolism (1, 2). The two classes of bisphosphonates, non-nitrogen containing bisphosphonates (NNBP) and nitrogen-containing bisphosphonates (NBP), differ structurally in the substitution pattern at the bridging carbon, and display distinct mechanisms of action. The NBP class, bearing nitrogen-containing substituents at the bridging carbon, acts through inhibition of isoprenoid biosynthesis enzymes, including farnesyl pyrophosphate synthase (FPPS; refs. 3 and 4), to reduce FPP levels and prevent downstream protein prenylation events essential for normal protein function. The NNBP class, which lacks a nitrogen-containing substituent at the bridging carbon, is known to undergo conversion to nonhydrolyzable ATP analogs (i.e., AppCCl2p). These metabolites are thought to inhibit ATP-dependent processes including the ATP/ADP translocase at the mito- Authors' Affiliations: Departments of 1Pharmacology and Molecular Sciences and 2Oncology; 3Analytical Pharmacology Core of the Sidney Kimmel Comprehensive Cancer Center, Johns Hopkins University School of Medicine, Baltimore, Maryland; and 4The Wistar Institute, Philadelphia, Pennsylvania Note: Supplementary data for this article are available at Molecular Cancer Therapeutics Online (http://mct.aacrjournals.org/). Current address for M.R. Webster: The Wistar Institute, Philadelphia, PA. Corresponding Authors: Caren L. Freel Meyers, WBSB 307B, 725 N. Wolfe St., Baltimore, MD 21205. Phone: 410-502-4807; Fax: 410-9553023; E-mail: [email protected] or Christine Hann, [email protected] doi: 10.1158/1535-7163.MCT-13-0315 2013 American Association for Cancer Research. chondrial membrane, leading to apoptosis in osteoclasts (5, 6). Osteolytic lesions develop in a majority of patients with multiple myeloma, and bone metastases are common in patients with breast, prostate, and lung cancers (7). Bisphosphonates with antiresorptive activity are used in adjuvant therapies to effectively treat malignant bone disease from advanced cancers. The ability of NBPs to inhibit cancer cell invasion and adhesion, and exert growth inhibitory activity in the bone microenvironment is well documented (7–10). Consequently, there is increasing interest to evaluate clinically used bisphosphonates as potential therapeutic agents for extraskeletal cancers (1). The most potent NBP, zoledronate, displays variable anticancer activity against several cancer cell lines (IC50s ranging from 14 to >100 mmol/L; refs. 11–15). Inhibition of FPPS by NBPs leads to a reduction in prenylation of small G proteins, which are essential for migration, adhesion, invasion, and proliferation. Furthermore, accumulation of isopentenyl pyrophosphate (IPP) as a result of FPPS inhibition leads to immune cell activation (10, 16) and production of cytotoxic ATP analogs, which induce apoptosis (10, 17). In vitro studies in human melanoma cell lines showed that the NBPs zoledronate and pamidronate inhibit proliferation and induce apoptosis (15, 18); however, zoledronate, which was the more potent of the two, displayed a modest IC50 100 mmol/L in the cell lines tested. Contributing to these activities in melanoma is the finding that NBPs inhibit several matrix metalloproteinases (MMP), which are important for tumor invasion and metastasis (19, 20). Relevant to this study, the NNBP clodronate is minimally active against breast, ovarian, and thyroid cancer cell lines (high mmol/L to low mmol/L IC50s; refs. 12, 21, www.aacrjournals.org Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. 297 Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Webster et al. and 22), and is inactive against melanoma (15), although it is reported to inhibit MMPs important for tumor invasion in melanoma cells (23, 24). In contrast to the limited preclinical activity observed, results from clinical trials suggest potential benefits of adjuvant therapy with clodronate for breast cancer in older patients. A recent clinical trial suggests clodronate increases bone metastasis–free and non-bone metastasis–free intervals, and recurrence-free interval in postmenopausal women (25). These results are consistent with two earlier studies showing increases in bone metastasis–free interval and an overall survival benefit in postmenopausal women (26), and a disease-free benefit in older patients (27). A study by Saarto and colleagues (28) suggested no benefit of adjuvant clodronate; however, this study was carried out in a predominantly premenopausal population. A primary limitation of the use of bisphosphonates in treating extraskeletal tumors is poor cell permeability, a consequence of the highly polar, polyanionic structure of bisphosphonates under physiological conditions. To address poor cellular uptake of bisphosphonates, we have developed a bisphosphonamidate prodrug strategy for efficient intracellular delivery of CLO and methylene bisphosphonate (MBP) in non–small cell lung cancer (NSCLC) cells (29). In each case, the neutral prodrug incorporates two biodegradable nitroaryl groups and two halobutyl amine masking groups (Fig. 1). After cellular uptake, the nitroaryl delivery groups undergo intracellular activation via an enzymatically driven nitroreduction, resulting in release of a halobutyl phosphonamidate anion. This phosphonamidate anion is chemically labile, undergoing a series of spontaneous cyclization and P-N hydrolysis reactions to release the fully unmasked bisphosphonate (Fig. 1). The CLO and MBP prodrugs exhibited significantly improved anticancer activity against NSCLC cells (low mmol/L IC50s), compared with the parent bisphosphonates (no activity up to low mmol/L concentrations), with the CLO prodrug showing the most potent activity (29). The CLO prodrug was submitted for analysis against the NCI-60 panel to evaluate its activity across multiple tumors types. Here we report the activity of the CLO prodrug against 9 different cancer cell types, and further characterization of our CLO prodrug in 2 melanoma cell lines, SK-Mel-5 and UACC-62. Our results support efficient intracellular activation of bisphosphonamidate prodrugs in melanoma cells, and demonstrate growth inhibition (GI) and induction of apoptosis by the CLO prodrug. We further demonstrate that intratumoral injection of the CLO prodrug in subcutaneous SK-Mel5 xenografts causes tumor growth inhibition. Materials and Methods Cell culture and LC/MS-MS UACC-62 cells were acquired from the National Cancer Institute DTP catalog. SK-Mel-5 melanoma cells were acquired from American Type Culture Collection. Both cell lines were immediately expanded and multiple ali- 298 Mol Cancer Ther; 13(2) February 2014 Figure 1. Bisphosphonamidate prodrugs of CLO and MBP. quots were cryopreserved. The cell lines were used within 6 months of resuscitation. Peripheral blood mononuclear cells were a generous gift from Dr. J. Conejo-Garcia (The Wistar Institute, Philadelphia, PA) and were obtained by leukapheresis/elutriation from healthy de-identified donors, under institutional review board’s approval (EX2100250-2). Keratinocytes were maintained in keratinocyte-SRM (1) media (Gibco; 17005-042). Keratinocytes were isolated from neonatal foreskins after routine circumcision (EX21212264-1a; The Wistar Institute). UACC62 cells were maintained in RPMI 1640 with 10% FBS, 1% pen/strep, 1% glutamate, and 1% sodium pyruvate. SKMel-5 cells were maintained in Eagle’s Minimum Essential Media (EMEM) with 10% FBS, 1% pen/strep, and 1% glutamine. Liquid chromatography/tandem mass spectrometry (LC/MS-MS) experiments were conducted using a Quadrapole ABI5500 mass spectrometric detector (Applied Biosystems). The instrument was equipped with an electrospray interface in positive ion mode, and controlled using the Analyst version 1.2 software (Applied Biosystems). Molecular Cancer Therapeutics Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Clodronate Prodrug Is Cytotoxic against Melanoma NCI-60 cell line screen The effect of clodronate prodrug on the NCI-60 cell lines was determined using an assay to detect total protein. After 48 hours, the assay was quenched with the addition of 50 mL of cold TCA, and incubation at 4 C for 60 minutes. The supernatant was removed, the plates were washed 5 times with tap water, and then air dried. Sulforhodamine B (SRB, 100 mL) solution [0.4% (w/v) in 1% acetic acids, 100 mL] was added to each well. The plates were incubated at room temperature (RT) for 10 minutes, unbound stain was removed by washing 5 times with 1% acetic acid, and the plates were allowed to air dry. Bound stain was solubilized with 10 mmol/L trizma base, and the absorbance at 515 nm was read using a plate reader (described on the DTP-NCI website: dtp.nci.nih.gov). that caused a 50% decrease in number of cells compared with control. Cell-cycle analysis Cell-cycle distribution was determined using a BD FACSCalibur flow cytometer. SKMel-5 and UACC-62 melanoma cells were plated at 6.7 104 cells per well in flat bottom 6-well plates. Cells were dosed as described earlier. At 24, 48, or 72 hours following drug treatment, the cells were analyzed as previously described (29). DNA content was analyzed using a BD FACSCalibur cytometer with 10,000 propidium iodide (PI)-positive gated events recorded per sample using CellQuest Pro software. Cell-proliferation assay Cell proliferation was determined using the Promega CellTiter 96 Aqueous One Solution Cell Proliferation Assay MTS assay. SK-Mel-5 and UACC-62 cells were plated at 6.0 103 and 2.5 103 cells per well, respectively, in flat bottom 96-well plates in 99 mL of media and allowed to adhere overnight. The drugs were serially diluted in 100% dimethyl sulfoxide (DMSO). For each drug treatment group, 1 mL of a 100 stock solution was added to each well for a final DMSO concentration of 1% (v/v). Cells were treated for 24, 48, or 72 hours, and analyzed as previously described (29). Measurement of caspase-3 activity SK-Mel-5 and UACC-62 melanoma cells were plated at 6.7 104 cells per well in flat bottom 6-well plates. Cells were dosed as described earlier. At 24 or 48 hours following drug treatment, the media was collected and the cells were washed with 1 mL PBS. Each well was trypsinized with 600 mL trypsin for 3 to 5 minutes. The trypsin reaction was quenched with an equal volume of media. All supernatants and washes were combined and centrifuged at 1,100 rpm for 5 minutes. The supernatant was removed, the cells were washed with 1 mL FBS with 1% PBS. The cells were centrifuged at 1,100 rpm for 5 minutes. Cells were resuspended in 250 mL Cytofix/Cytoperm (Pharmingen Kit #2075KK) and incubated at room temperature for 20 minutes. After 20 minutes, cells were centrifuged at 1,100 rpm for 5 minutes. Cells were washed 2 with Perm/wash solution, centrifuging, and decanting after each wash. Cells were resuspended in 50 mL Perm/ wash solution, 20 mL FITC Rabbit antiactive caspase-3 antibody (BD Pharmigen; 559341) was added, and samples were incubated at RT away from light. Cells were spun at 1,100 rpm for 5 minutes. Cells were washed with 1 mL Perm/Wash, spun at 1,100 rpm for 5 minutes, and resuspend in 300 mL Perm/Wash solution. The cells were analyzed by flow cytometry and compared with control. Cells not treated with antibody were used as a negative control, cells treated with 500 nmol/L velcade (bortezomib) were used as a positive control. Quadrants were set using the positive control, negative control and 0 mmol/L clodronate prodrug samples. The percentage of cells in the upper and lower left was determined by Cell Quest Pro software. Trypan blue cell viability assay SK-Mel-5 and UACC-62 melanoma cells were plated at 7.1 104 and 3 104 cells per well, respectively, in flat bottom 12-well plates. Cells were dosed as described earlier. At 24, 48, or 72 hours following drug treatment, the cells were collected and analyzed as previously described (29). The absolute number of cells was determined at each drug concentration. The cell number for each concentration was converted to percentage of control for each time point, and plotted using GraphPad Prism 4.0. The EC50 was calculated as the concentration of drug In Vivo Efficacy Study Female, athymic nude mice (3–4 weeks old, purchased from Taconic) were injected subcutaneously with 4.5 106 of SK-Mel-5 cells suspended in 100 mL of 1:1 PBS/matrigel. After tumors reached a volume of 200 mm3, mice were randomized into treatment groups as follows: treatment group 1 (4 mice): vehicle twice per week; treatment group 2 (5 mice): drug twice per week. Prodrug was dissolved in corn oil and administered intratumorally at a final dose of 200 mg/kg. Mice were weighed and tumor volume was measured twice per week and monitored according to Detection of intracellular MBP prodrug and methylenebisphosphonate SK-Mel-5 and UACC-62 melanoma cells were plated at 1.3 105 cells per well in flat bottom 6-well plates. Cells were dosed as described earlier. At 2, 4, or 8 hours following drug treatment, the media was removed, and the cells were washed with 1 mL PBS. To each well, 600 mL of trypsin was added and incubated for 3 to 5 minutes. The trypsin reaction was quenched with an equal volume of media. The cells were transferred to a 15 mL conical and centrifuged at 1,100 rpm for 5 minutes. Supernatant was removed, and the cells were resuspended in 200 mL of media. The cells were then treated and analyzed using LC/MS-MS for intracellular levels of MBP prodrug and derivatized MBP as previously described (29). www.aacrjournals.org Mol Cancer Ther; 13(2) February 2014 Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. 299 Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Webster et al. ACUC protocols. Tumor length (mm) and tumor width (mm) were measured and used to calculate tumor volumes: [tumor length (mm) (tumor width tumor width) (mm)2]/2. A 2-tailed paired t test was used to determine significant difference (P < 0.05) between vehicle- and drug-treated groups. Results NCI-60 cell line panel screening reveals selective toxicity of the CLO prodrug against melanoma cell lines The CLO prodrug was submitted to the National Cancer Institute Developmental Therapeutics Program for screening against the NCI-60 cell line panel. The results from the NCI-60 5-dose screen were analyzed for 3 different endpoints, including growth inhibition, total growth inhibition (TGI), and lethal concentration (LC). The CLO prodrug exhibits general growth inhibitory activity against all the cell lines tested (Supplementary Table S1). A TGI concentration, where growth was inhibited 100%, was reported for 18 of 60 cell lines (Table 1), and the CLO prodrug was shown to be cytotoxic (LC50) against 4 cell lines. A concentration of the CLO prodrug where 50% of the cells died (LC50) could be measured for 4 cell lines (Table 1); 3 of these were melanoma cell lines (LOX IMVI, SK-MEL-5, and UACC-62), suggesting that the CLO Table 1. Results from NCI-60 screen of the CLO prodrug Leukemia NSCLC Colon cancer CNS cancer Melanoma Renal cancer Breast cancer Cell line TGI (mmol/L)a LC50 (mmol/L)b RMPI-8826 NCI-H522 COLO 205c HT29 SF-295 SNB-75 LOX IMVIc MDA-MB-4 35 SK-MEL-2 SK-MEL-5c UACC257 UACC-62c A498 RXF 393 MCF7 HS 578T BT-549 MDA-MB-468 9.12 7.76 7.94 22.2 16.98 6.76 4.64 22.6 6.57 4.24 7.89 5.03 6.82 4.13 1.52 9.21 9.31 8.59 >100 >100 34.4 >100 >100 >100 54.4 >100 >100 8.88 >100 78.1 >100 >100 >100 >100 >100 >100 a TGI is the minimum concentration of the CLO prodrug required for total growth inhibition. b LC50 is the concentration of the CLO prodrug required to cause 50% cell death compared with 0 hour. c Cell lines where an LC50 could be measured. 300 Mol Cancer Ther; 13(2) February 2014 prodrug may exhibit selective cytotoxicity against melanoma. We selected 2 of these cell lines, SK-Mel-5 and UACC-62, to further evaluate the cellular uptake, intracellular activation, and anticancer activity of the CLO prodrug. Bisphosphonamidate prodrugs show enhanced cell permeability and undergo intracellular activation in SK-Mel-5 and UACC-62 cells The activity of the CLO prodrug in melanoma cells is consistent with increased cell permeability of the bisphosphonamidate prodrug. In a previous study, we demonstrated the chemistry of prodrug activation as well as intracellular prodrug activation through a combination of 31P NMR and LC/MS-MS experiments carried out with the structurally related methylenebisphosphonate (MBP) prodrug as a model for this prodrug class (29). Likewise, we have evaluated cellular uptake and intracellular activation of the same model MBP prodrug in SK-Mel-5 and UACC-62 melanoma cells. Briefly, intracellular levels of bisphosphonate prodrug were measured in SK-Mel-5 and UACC-62 melanoma cells using the LC/MS-MS method previously reported by our group (29). Methylenebisphosphonate released following intracellular activation of the MBP prodrug (Fig. 2 and Supplementary Fig. S1A) is itself difficult to detect by mass spectrometry; however, the corresponding TBS ester (TBS-MBP), generated by derivatization of MBP with N-t-butyldimethylsilyl-N-methyltrifluoroacetamide (MTBSTFA), is easily detected by LC/ MS-MS. Thus, SK-Mel-5 and UACC-62 melanoma cell lines were treated with the MBP prodrug at varying concentrations (0, 10, or 30 mmol/L) for 2, 4, and 8 hours (Fig. 2 and Supplementary Fig. S1). LC/MS-MS analysis of cell lysates indicated that intracellular prodrug levels are achieved in a concentration-dependent manner; at 8 hours, MBP prodrug levels of 175.6 and 408.3 ng/104 cells were observed in SK-Mel-5 cells treated with 10 and 30 mmol/L MBP prodrug, respectively (Fig. 2B). A similar observation was made in UACC-62 cells treated with the MBP prodrug at 8 hours (Supplementary Fig. S1B). The presence of intracellular bisphosphonate, in SK-Mel-5 and UACC-62 cells treated with 10 or 30 mmol/L of the MBP prodrug or 30 mmol/L bisphosphonate, was determined by detection of the corresponding TBS ester (TBS-MBP) formed following treatment of cell lysate with MTBSTFA (Fig. 2C and Supplementary Fig. S1C). Intracellular bisphosphonate is detected as early as 2 hours in SKMel-5 and UACC-62 cells treated with 10 and 30 mmol/L MBP prodrug (Fig. 2A and Supplementary Fig. S1A). CLO prodrug is cytotoxic to SK-Mel-5 and UACC-62 melanoma cell lines Clodronate has shown little or no activity against multiple cancer cell lines, including melanoma (18); however, we have previously demonstrated that the cell-permeable bisphosphonamidate prodrug of CLO displays dramatically increased activity against the A549 (NSCLC) cell line (29). Results from the NCI-60 screen suggests that the CLO Molecular Cancer Therapeutics Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Clodronate Prodrug Is Cytotoxic against Melanoma Figure 2. The bisphosphonamidate prodrug class is cell permeable and undergoes intracellular activation in SK-Mel-5 cells. A, MBP prodrug permeability and intracellular activation confirmed by LC/MSMS analysis. B, ion chromatograms showing absence of MBP prodrug in untreated cells (a), and detection of MBP prodrug in cells treated with 10 mmol/L 4 (175.6 ng/10 cells) (b) and 30 4 mmol/L (408.3 ng/10 cells) (c) MBP prodrug. C, ion chromatograms showing absence of tetra-silyl bisphosphonate following MTBSTFA reaction of lysate from untreated cells (a), and detection of TBS-MBP in cells treated with 30 mmol/L MBP (b), cells treated with 10 mmol/L MBP prodrug (c), and cells treated with 30 mmol/L MBP prodrug (d). Ion chromatograms are offset in the y-dimension for clarity. prodrug exhibits a more pronounced affect against SKMel-5 and UACC-62 melanoma cell lines compared with A549 cells. Growth inhibition was measured in the A549 cells with a GI50 of 4.25 mmol/L (50% less growth compared with control), but TGI and cell death were not observed. In contrast, TGI and cell death were observed www.aacrjournals.org in SK-MEL-5 and UACC-62 cells treated with the CLO prodrug (Table 1). To further characterize this activity, we evaluated the effect of the CLO prodrug on proliferation and viability of SK-Mel-5 and UACC-62 melanoma cells, using the MTS and cell number assays. Dose–response curves were generated using drug concentrations ranging Mol Cancer Ther; 13(2) February 2014 Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. 301 Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Webster et al. Figure 3. CLO prodrug is cytotoxic to SK-Mel-5 and UACC-62 melanoma cell lines. A and B, MTS assay of SK-Mel-5 (A) or UACC-62 (B) melanoma cells treated with clodronate for 72 hours (*), or CLO prodrug for 24 hours (&), 48 hours (~), and 72 hours (^); C and D, cell number, determined using trypan blue, of SK-Mel-5 (C) or UACC-62 (D) treated with CLO prodrug at 24 hours (&), 48 hours (~), and 72 hours (^). from 0.3 to 300 mmol/L, and cell proliferation was measured at 24, 48, and 72 hours after drug treatment (Fig. 3A and B and Supplementary Table S2), using the MTS assay. As expected, the CLO prodrug displays a remarkable enhancement in activity compared with clodronate in both cell lines. The IC50s of CLO prodrug against melanoma cell lines, SK-Mel-5 (7 1 mmol/L) and UACC-62 (11.7 3 mmol/L) at 72 hours, are comparable with the IC50 values previously determined in A549 cells (4.4 2 mmol/L), suggesting comparable uptake across these cell lines. In both melanoma cell lines, there is a full antiproliferative effect by 48 hours, whereas in A549 cells a full effect is not observed until 72 hours. To correlate the growth inhibition of melanoma cells with viability, absolute cell number at varying drug concentration was determined using a trypan blue exclusion assay. The number of viable cells at each dose was measured at 0, 24, 48, and 72 hours following drug treatment and plotted as a percentage of control (Fig. 3C and D). The EC50 of the CLO prodrug in SK-Mel-5 cells is 6.3 3 mmol/L, which is similar to its IC50 of 6.1 1 mmol/L (determined at 48 hours by the MTS assay; Supplementary Table S2). Similarly, the EC50 of 12.7 5 mmol/L for the CLO prodrug in UACC-62 cells is comparable with its IC50 of 8 4 mmol/L at 48 hours; however, in both cell lines there seems to be a greater antiproliferative effect at 24 hours compared with the effect on cell viability at the same drug treatment time. This difference at 24 hours could reflect the time required for prodrug activation and conversion of released bisphosphonate to the active metabolite. As a measure of general toxicity, we assessed the effect of CLO prodrug on normal cells and determined that CLO prodrug does not affect the viability of PBMCs or primary keratinocytes (Supplementary Fig. S2). 302 Mol Cancer Ther; 13(2) February 2014 CLO prodrug induces apoptosis in melanoma cell lines To further characterize the mechanism of cytoxicity induced by the CLO prodrug in SK-Mel-5 and UACC62 cells, we evaluated the effect of drug exposure on cellcycle distribution and induction of apoptosis (Fig. 4A–C). The cell-cycle profile of SK-Mel-5 and UACC-62 cells treated with increasing concentrations of CLO prodrug was analyzed at 24, 48, and 72 hours. A pronounced subG1 peak was observed in cells treated with 10 mmol/L CLO prodrug after 48 hours, characteristic of caspase-activated DNase (CAD) activity (Fig. 4C). CAD is downstream of caspase-3, which catalyzes cleavage of ICAD to CAD (30); thus, the increased percentage of cells in sub-G0–G1 phase suggests the activation of caspase-3 (Fig. 4D and E). Strikingly, the percentage of cells with active CAD nearly doubles at each time point in SK-Mel-5 cells treated with 10 mmol/L CLO prodrug, and in UACC-62 cells treated with >30 mmol/L CLO prodrug. To confirm this, SK-Mel-5 and UACC-62 cells were treated with increasing concentrations of the CLO prodrug and assayed for activated caspase-3 at 24 and 48 hours by flow cytometry using an antibody specific for cleaved caspase-3 (Fig. 4F and G). Velcade (bortezomib) was used as a positive control for activated caspase-3–mediated apoptosis. In both cell lines, significant caspase-3 activity was measured at >3 mmol/L CLO prodrug, providing support for a mechanism involving induction of apoptosis by CLO prodrug. Intratumoral delivery of CLO prodrug causes tumor growth inhibition On the basis of the promising cytotoxicity of CLO prodrug in vitro, we sought to determine its activity in a mouse xenograft model. As a proof of concept that CLO Molecular Cancer Therapeutics Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Clodronate Prodrug Is Cytotoxic against Melanoma Figure 4. CLO prodrug induces apoptosis in melanoma cell lines. A, cell-cycle analysis of SK-Mel-5 cells after 48 hours of treatment with 0 mmol/L (A), 3 mmol/L (B), and 10 mmol/L (C) CLO prodrug. D and E, quantification of percent SKMel-5 (D) or UACC-62 (E) cells in sub-G0–G1 at 24, 48, and 72 hours when treated with 0, 1, 3, 10, 30, and 100 mmol/L CLO prodrug. F and G, percentage of SK-Mel-5 (F) or UACC-62 (G) cells positive for activated caspase-3 at 24 and 48 hours posttreatment with CLO prodrug. Velcade (500 nmol/L) was used as a positive control at 24 and 48 hours. Cells (–) drug and (–) antibody was used as a negative control at 24 and 48 hours, to determine background cellular fluorescence. prodrug exhibits antitumor effects in vivo, CLO prodrug was delivered by intratumoral injection into SK-Mel-5 subcutaneous xenografts. Tumor growth was compared with vehicle control as fold change in tumor size (Fig. 5A). After 17-day treatment, as compared with a vehicle control group, statistically significant (P < 0.05) decrease in tumor growth was observed with twice weekly injection of the CLO prodrug, compared with control. This difference further increased after the 19th day (P < 0.001), suggesting that if good bioavailability in tumor tissues is achieved, an antitumor effect will be observed. Mouse weight was measured twice weekly as a measure of general toxicity (Fig. 5B). In contrast, IT administration of a similar dose of clodronate did not have a significant effect on tumor growth of SK-Mel-5 xenografts (Supplementary Fig. S3). Discussion Poor cellular uptake has limited the use of bisphosphonates in extraskeletal diseases. Previously, we described a www.aacrjournals.org prodrug strategy for efficient intracellular delivery of bisphosphonates to overcome this limitation. Here, we report the results from evaluation of a bisphosphonamidate clodronate prodrug (CLO prodrug) against the NCI60 cell line panel, a widely used resource, which has been well-characterized molecularly and pharmacologically. Results from the screen indicate that the CLO prodrug seems selectively cytotoxic against melanoma cells lines, displaying LC50 values in the micromolar range for only 4 cell lines, 3 of which are melanoma lines. Further investigation of CLO prodrug activity against 2 susceptible cell lines, SK-Mel-5 and UACC-62, confirms remarkably enhanced activity of the CLO prodrug compared with clodronate, which presumably results from enhanced cell permeability and consequently increased intracellular clodronate concentration. This notion is further supported by the LC/MS-MS detection of high intracellular levels of the structurally related model MBP prodrug and released MBP in both cell lines; however, cytotoxic effects of the Mol Cancer Ther; 13(2) February 2014 Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. 303 Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Webster et al. Figure 5. Intratumoral delivery of CLO prodrug causes tumor growth inhibition of SK-Mel-5 xenografts. A, tumor size of CLO prodrug-treated mice compared with control was followed for 4 weeks. There was a significant TGI of CLO prodrug (&) compared with control (*) in mice dosed twice per week. B, weights of nude mice bearing xenografts treated with IT CLO prodrug (&) were compared with control (*). A statistically significant (P < 0.05) decrease in tumor growth was observed with the CLO prodrug, compared with control. , this difference further increased after the 19th day (P < 0.001). prodrug itself cannot be ruled out. Interestingly, the CLO prodrug seems to exert its anticancer effects by a distinct mechanism in melanoma cells. Our previous investigation of the CLO prodrug in A549 cells revealed a somewhat less potent effect on cell viability compared with growth inhibition, and these effects generally correlated with a mechanism involving G1-phase cell-cycle arrest. In contrast, the low micromolar activity of the CLO prodrug in growth inhibition and cell viability assays in melanoma cells seems to correlate with induction of apoptosis in both cell lines, as evidenced by the appearance of a sub-G0–G1 peak in cell-cycle analysis experiments and increased activation of caspase-3 in both cell lines. Induction of apoptosis is consistent with a mechanism involving CLO prodrug activation to release CLO and subsequent conversion of CLO to the nonhydrolyzable ATP analog (AppCl2p), a known inhibitor of mitochondrial ADP/ ATP translocase (5, 6). Interestingly, although clodronate exhibits minimal activity against cancer cell lines, studies by Yang and colleagues highlight induction of apoptosis in human thyroid cancer cell lines correlating with calcium release from mitochondrial and endoplasmic reticulum that leads to membrane depolarization of mitochondria (21, 22). Nevertheless, CLO exhibits only weak growth inhibitory activity in these cells lines, presumably as a result of poor cellular uptake. It is tempting to speculate that a similar effect on intracellular calcium signaling may underlie the potent, selective cytotoxicity of the CLO prodrug against melanoma cells, as calcium signaling is known to be crucial in melanoma progression (31–33); however, further investigation is required to determine the molecular mechanism of action of the CLO prodrug in melanoma cells. The surprising selective cytotoxicity of the CLO prodrug against SK-Mel-5 and UACC-62 cells suggests a new application of bisphosphonates in melanoma therapy and offers a solution to the problem of poor cellular uptake of this class. Indeed, preliminary evaluation of the in vivo antitumor effect of the prodrug is promising, as intratu- 304 Mol Cancer Ther; 13(2) February 2014 moral injection of SK-Mel-5 xenografts causes significant tumor growth inhibition. Advances in molecular targeting and immunotherapy have added to the current treatment options for patients with metastatic melanoma. BRAF, a key member of the MAPK signaling cascade, is mutated at the V600 residue in 45% of metastatic melanoma cases and is predictive for responsiveness to the tyrosine kinase inhibitor, vemurafenib (34). Notably, the cell lines on the NCI-60 panel, most sensitive to the CLO prodrug (Supplementary Table S1) all harbor a BRAFV600E mutation, which could suggest that the CLO prodrug may have an effect on RAF function; however, we note that other cell lines harboring the BRAFV600E mutation (Supplementary Table S1) are not susceptible to the cell killing effects of CLO prodrug. A second newly approved therapy, Ipilimumab, is a monoclonal antibody that targets the immune checkpoint, CTLA-4, and was shown to significantly improve overall survival; a favorable response was observed in 20% of patients (35, 36). In 2011, both vemurafenib and ipilimumab received approval of the U.S. Food and Drug Administration for the treatment of patients with metastatic melanoma. Although each represents an improvement for treatment of patients with melanoma, their use is restricted to specific subsets of patients with metastatic melanoma, and neither agent is curative. Thus, for patients who are not candidates for either therapy, or for those who progress while on therapy, treatment with DNA damaging agents, such as dacarbazine and temozolomide, remain the only therapeutic options (37, 38). Response rates to these agents are low, and long-term benefit is rare; thus, there continues to be great need for the development of melanoma-specific treatments. Disclosure of Potential Conflicts of Interest No potential conflicts of interest were disclosed. Authors' Contributions Conception and design: M.R. Webster, C.L. Hann, C.L. Freel Meyers Development of methodology: M.R. Webster, C. Kamat, M. Zhao, M.A. Rudek, C.L. Hann Molecular Cancer Therapeutics Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Clodronate Prodrug Is Cytotoxic against Melanoma Acquisition of data (provided animals, acquired and managed patients, provided facilities, etc.): M.R. Webster, C. Kamat, N. Connis, A.T. Weeraratna, M.A. Rudek, C.L. Hann Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computational analysis): M.R Webster, C. Kamat, M. Zhao, C.L. Hann, C.L. Freel Meyers Writing, review, and/or revision of the manuscript: M.R. Webster, C. Kamat, N. Connis, A.T. Weeraratna, M.A. Rudek, C.L. Hann, C.L. Freel Meyers Administrative, technical, or material support (i.e., reporting or organizing data, constructing databases): M.R. Webster Study supervision: C. Kamat, C.L. Hann, C.L. Freel Meyers Acknowledgments The authors thank Dr. M. Herlyn (The Wistar Institute) for supplying keratinocytes and Dr. J. Conejo-Garcia (The Wistar Institute) for supplying PBMCs for general toxicity studies and I. Dobromilskaya (Johns Hopkins University) for technical help with in vivo studies. Grant Support C.L. Hann received an award from the Flight Attendants Medical Research Institute. M.A. Rudek was supported by the Analytical Pharmacology Core of the Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins University (NIH grants P30 CA006973 and UL1 RR025005), the Shared Instrument Grant (1S10RR026824-01) and an award from the Johns Hopkins FAMRI Center of Excellence. The project described was supported in part by grant number UL1 RR025005 from the National Center for Research Resources (NCRR), a component of the National Institutes of Health (NIH) and NIH Roadmap for Medical Research. C.L. Freel Meyers was supported by NIH grant AI099704 and an award from the Johns Hopkins FAMRI Center of Excellence. M.R. Webster was supported by an award from the Johns Hopkins FAMRI Center of Excellence and training grant (2 T32 CA 9171-36) at The Wistar Institute. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Received April 26, 2013; revised October 14, 2013; accepted November 17, 2013; published OnlineFirst December 5, 2013. References 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. Morgan G, Lipton A. Antitumor effects and anticancer applications of bisphosphonates. Semin Oncol 2010;37:S30–40. Gnant M. Bisphosphonates in the prevention of disease recurrence: current results and ongoing trials. Curr Cancer Drug Targets 2009;9: 824–33. Dunford JE, Kwaasi AA, Rogers MJ, Barnett BL, Ebetino FH, Russell GG, et al. Structure-activity relationships among the nitrogen containing bisphosphonates in clinical use and other analogues: time-dependent inhibition of human farnesyl pyrophosphate synthase. J Med Chem 2008;51:2187–95. €ikko € nen J, Taskinen M, Dunford JE, Mo € nkko € nen H, Auriola S, Ra € nkko € nen J. Correlation between time-dependent inhibition of Mo human farnesyl pyrophosphate synthase and blockade of mevalonate pathway by nitrogen-containing bisphosphonates in cultured cells. Biochem. Biophys Res Commun 2011;407:663–7 € nkko € nen J. IdentificaAuriola S, Frith J, Rogers MJ, Kiovuniemi A, Mo tion of adenine nucleotide-containing metabolites of bisphosphonate drugs using ion-pair liquid chromatography-electrospray mass spectrometry. J Chromat B 1997;704:187–95. €pa €nkangas JP, Ylitalo KV, Mo € nkko € nen Lehenkari PP, Kellinsalmi M, Na J, Rogers MJ, et al. Further insight into mechanism of action of clodronate: inhibition of mitochondrial ADP/ATP translocase by a nonhydrolyzable, adenine-containing metabolite. Mol Pharmacol 2002;61:1255–62. Coleman RE. Skeletal complications of malignancy. Cancer 1997;80: 1588–94. Boissier S, Magnetto S, Frappart L, Cuzin B, Ebetino FH, Delmas PD, et al. Bisphosphonates inhibit prostate and breast carcinoma cell adhesion to unmineralized and mineralized bone extracellular matrices. Cancer Res 1997;57:3890–4. Boissier S, Ferreras M, Peyruchaud O, Magnetto S, Ebetino FH, Colombel M, et al. Bisphosphonates inhibit breast and prostate carcinoma cell invasion, an early event in the formation of bone metastases. Cancer Res 2000;60:2949–54. zardin P. Bisphosphonates' antitumor activity: an unraveled side of Cle a multifaceted drug class. Bone 2011;48:71–9. Shipman CM, Rogers MJ, Apperley JP, Russell RG, Croucher PI. Bisphosphonates induce apoptosis in human myeloma cell lines: a novel anti-tumor activity. Br J Haematol 1997;98:665–72. Lipton A. Emerging role of bisphosphonates in the clinic-antitumor activity and prevention of metatasis to bone. Cancer Treatment Rev. 2008;34:525–30. Knight LA, Conroy M, Fernando A, Polak M, Kurbacher CM, Cree IA. Pilot studies of the effect of zoledronic acid (Zometa) on tumor-derived cells ex vivo in the ATP-based tumor chemosensitivity assay. Anticancer Drugs 2005;16:969–76. Matsumoto S, Kimura S, Segawa H, Kuroda J, Yuasa T, Sato K, et al. Efficacy of the third-generation bisphosphonate zoledronic acid alone www.aacrjournals.org 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. and combined with anti-cancer agents against small cell lung cancer cell lines. Lung Cancer 2005;47:31–9. €ller C, Riebeling C, Orfanos CE, Geilen CC. NitrogenForsea AM, Mu containing bisphosphonates inhibit cell cycle progression in human melanoma cells. Br J Cancer 2004;91:803–10. € nkko € nen H, Rogers MJ, Mo € nkko € nen J, Roelofs AJ, Jauhiainen M, Mo Thompson K. Peripheral blood monocytes are responsible for gdT cell activation induced by zoledronic acid through accumulation of IPP/ DMAPP. Br J Haematol 2009;144:245–50. zardin P. The molecular basis of bisphosphonate activity: a Green J, Cle preclinical perspective. Semin Oncol 2010;37:S3–11. Riebeling C, Forsea A-M, Raisova M, Orfanos CE, Geilen CC. The bisphosphonate pamidronate induces apoptosis in human melanoma cells in vitro. Br J Cancer 2002;87:366–71. Heikkila P, Teronen O, Moianen M, Konttinen YT, Hanemaaijer R, Laitinen M, et al. Bisphosphonates inhibit stromelysin-1 (MMP-3), matrix metalloelastase (MMP-12), collagenase-2 (MMP-13) and enamelysin (MMP-20), but not urokinase-type plasminogen activator, and diminish invasion and migration of human malignant endothelial cells. Anticancer Drugs 2002;13:245–54. Hofmann UB, Westphal JF, van Muijen GNP, Ruiter DJ. Matrix metalloproteinases in human melanoma. J Invest Dermatol 2000;115: 337–44. Yang DM, Chi CW, Chang HM, Wu LH, Lee TK, Lin JD, et al. Effects of clodronate on cancer growth and Ca2þ signaling of human thyroid carcinoma cell lines. Anticancer Res 2004;24:1617–24. Yang DM, Teng HC, Chen KH, Tsai ML, Lee TK, Chou YC, et al. Clodronate-induced cell apoptosis in human thyroid carcinoma is mediated via the P2 receptor signaling pathway. J Pharmacol Exp Ther 2009;330:613–23. Teronen O, Konttinen YT, Lindgvist C, Salo T, Ingman T, Lauhio A, et al. Human neutrophil collagenase MMP-8 in peri-implant sulcus fluid and its inhibition by clodronate. J Dent Res 1997;76:1529–37. Teronen O, Konttinen YT, Lindgvist C, Salo T, Ingman T, Lauhio A, et al. Inhibition of matrix metalloproteinase-1 by dichloromethylene bisphosphonate (clodronate). Calcif Tissue Int 1997;61:59–61. Paterson AHG, Anderson SJ, Lembersky BC, Fehrenbacher L, Falkson CI, King KM, et al. Oral clodronate for adjuvant treatment of operable breast cancer (National Surgical Adjuvant Breast and Bowel Project protocol B-34): a multicentre placebo-controlled, randomized trial. Lancet Oncol 2012;13:734–42. Powles T, Paterson A, McCloskey E, Schein P, Scheffler B, Tidy A , et al. Reduction in bone relapse and improved survival with oral clodronate for adjuvant treatment of operable breast cancer [ISRCTN83688026]. Breast Cancer Res 2006;8:R13. Diel IJ, Solomayer EF, Costa SD, Gollan C, Goerner R, Wallwiener D , et al. Reduction in new metastases in breast cancer with adjuvant clodronate treatment. N Engl J Med 1998;339:357–63. Mol Cancer Ther; 13(2) February 2014 Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. 305 Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Webster et al. 28. Saarto T, Vehmanen L, Virkkunen P, Blomgvist C. Ten-year follow-up of a randomized controlled trial of adjuvant clodronate treatment in node-positive breast cancer patients. Acta Oncol 2004;43:650–6. 29. Webster MR, Zhao M, Rudek MA, Hann CL, Freel Meyers CL. Bisphosphonamidate clodronate prodrug exhibits potent anticancer activity in non-small-cell lung cancer cells. J Med Chem 2011;54:6647–56. 30. Sakahira H, Enari M, Nageta S. Cleavage of CAD inhibitor in CAD activation and DNA degradation during apoptosis. Nature 1998;39: 96–9. 31. Weeraratna AT, Jiang Y, Hostetter G, Rosenblatt K, Duray P, Bittner M, et al. Wnt5a signaling directly affects cell motility and invasion of metastatic melanoma. Cancer Cell 2002;1:279–88. 32. Dissanayake SK, Wade M, Johnson CE, O'Connell MP, Leotlela PD, French AD, et al. The Wnt5A/protein kinase C pathway mediates motility in melanoma cells via the inhibition of metastasis supressors and initiation of an epithelial to mesenchymal transition. J Biol Chem 2007;28:17259–71. 306 Mol Cancer Ther; 13(2) February 2014 33. Dissanayake SK, Weeraratna AT. Detecting PKC phosphorylation as part of the Wnt/calcium pathway in cutaneous melanoma. Method Mol Biol 2008;466:157–72. 34. Chapman PB, Hauschild A, Robert C, Haanen JB, Ascierto P, Larkin J, et al. BRIM-3 Study Group. Improved survival with vemurafenib in melanoma with BRAF V600E mutation. N Engl J Med 2011;364: 2507–16. 35. Hodi FS, O'Day SJ, McDermott DF, Weber RW, Sosman JA, Haanen JB, et al. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med 2010;363:711–23. 36. Robert C, Thomas L, Bondarenko I, O'Day S, M D JW, Garbe C, et al. Ipilimumab plus dacarbazine for previously untreated metastatic melanoma. N Engl J Med 2011;364:2517–26. 37. Atallah E, Flaherty L. Treatment of metastatic malignant melanoma. Curr Treat Options Oncol 2005;6:185–93. 38. Sun W, Schuchter LM. Metastatic melanoma. Curr Treat Options Oncol 2001;2:193–202. Molecular Cancer Therapeutics Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research. Published OnlineFirst December 5, 2013; DOI: 10.1158/1535-7163.MCT-13-0315 Bisphosphonamidate Clodronate Prodrug Exhibits Selective Cytotoxic Activity against Melanoma Cell Lines Marie R. Webster, Chandrashekhar Kamat, Nick Connis, et al. Mol Cancer Ther 2014;13:297-306. Published OnlineFirst December 5, 2013. Updated version Supplementary Material Access the most recent version of this article at: doi:10.1158/1535-7163.MCT-13-0315 Access the most recent supplemental material at: http://mct.aacrjournals.org/content/suppl/2013/12/05/1535-7163.MCT-13-0315.DC1 Cited articles This article cites 38 articles, 5 of which you can access for free at: http://mct.aacrjournals.org/content/13/2/297.full.html#ref-list-1 E-mail alerts Sign up to receive free email-alerts related to this article or journal. Reprints and Subscriptions Permissions To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at [email protected]. To request permission to re-use all or part of this article, contact the AACR Publications Department at [email protected]. Downloaded from mct.aacrjournals.org on June 18, 2017. © 2014 American Association for Cancer Research.