Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
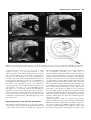
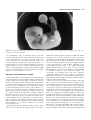
Human Reproduction Update 2000, Vol. 6 No. 3 pp. 268–278 © European Society of Human Reproduction and Embryology Fluid compartments of the embryonic environment Eric Jauniaux1,* and Beatrice Gulbis2 1 Academic Departments of Obstetrics and Gynaecology, Royal Free and University College London Medical School, UCL campus, London, UK and 2Academic Department of Clinical Chemistry, Academic Hospital Erasme, Université Libre de Bruxelles, Brussels, Belgium Received on September 28, 1999; accepted on February 15, 2000 The exocoelomic cavity was probably the last remaining physiological body fluid cavity to be explored in the human embryo. Its unique anatomical position has enabled us to study the protein metabolism of the early placenta and secondary yolk sac and to explore materno–embryonic transfer pathways. The exocoelomic cavity forms inside the extraembryonic mesoderm alongside the placental chorionic plate and is now believed to be an important transfer interface and a reservoir of nutrients for the embryo. Maternal or placental proteins filtered in the extraembryonic coelomic cavity are probably absorbed by the secondary yolk sac which is directly connected with the primitive digestive system throughout embryonic development. Protein electrophoresis has shown that the coelomic fluid results from an ultrafiltrate of maternal serum with the addition of specific placental and secondary yolk sac bioproducts demonstrating that the exocoelomic cavity is a physiological liquid extension of the early placenta. The selective sampling of fluid from the exocoelomic cavity has also offered a novel approach to the study of drug and toxin transfer across the early human placenta and as a unique tool to explore embryonic physiology in vivo. Further investigation should include a comparison between the coelomic fluid values of a molecule and its quantifiable presence in decidual, placental and fetal tissues. Key words: amniotic fluid/coelomic fluid/first trimester of pregnancy/placenta/yolk sac TABLE OF CONTENTS Introduction Embryology of the human placenta and adnexae Embryonic physiology Biology of the extraembryonic coelom Biology of the early amniotic cavity Prospects in the investigation of the biology of embryonic fluids References 268 269 270 271 275 276 277 Introduction With the development of amniocentesis for genetic investigation, the amniotic fluid (AF) became the first fluid available to study the fetal environment in utero. AF analyses have been performed from the third month of gestation onwards and have demonstrated important variation in gas tension, acid–base status and biochemical composition with gestational age and differences compared with maternal blood. During the 1970s, first trimester AF biochemistry was investigated (Sinha and Carlton, 1970; Johnell and Nilsson, 1971). Due to technical difficulties in recognizing accurately the different anatomical structures of the early gestational sac in utero, it is likely that most of the few samples obtained in these studies, at <12 weeks gestation, were a mixture of coelomic fluid (CF) and AF. In fact the original anatomical finding that the extraembryonic coelom is a fluid cavity, which surrounds the embryo and fetus during most of the first trimester (Boyd and Hamilton, 1970), was completely ignored by most authors in the 1960s and 70s. Furthermore, some authors believed that it was a thin virtual space containing a gelatinous substance that could not be aspirated (MacCarthy and Saunders, 1978). The advent of high resolution transvaginal ultrasound transducers at the end of the 1980s has enabled a more detailed morphological assessment of the early gestational sac in utero. In particular, the membrane separating the exocoelomic and amniotic cavities can now be clearly identified and CF can be selectively aspirated from 5 weeks gestation (Figure 1). In 1991, two independent teams, based at King’s College Hospital Medical School (Jauniaux et al., 1991) and St Bartholomew’s Hospital Medical College (Wathen et al., 1991), reported what they believed were the first data on the biochemistry of the extraembryonic coelom. However, in 1958, McKay et al. had already published a study on the protein content of the coelomic and amniotic fluids of five normal first-trimester pregnancies, obtained during hysterotomy (McKay et al., 1958). Although the number of samples studied was extremely small, the total protein concentration found by these authors in chorionic or CF was very similar to that found in our studies, >30 years later (Jauniaux et al., 1991, 1993; Gulbis et al., 1992). *To whom correspondence should be addressed at The Academic Department of Obstetrics and Gynaecology, University College London, 86–96 Chenies Mews, London WC1E 6HX, UK. Phone: +44 207 2096057; Fax: +44 207 3837429; E-mail: [email protected] Embryonic fluid compartments 269 Figure 1. Coelomic fluid aspiration procedure. (A) The exocoelomic fluid cavity (EEC) is located by means of transvaginal ultrasound. (B–D) The needle (20-gauge) is inserted between the placenta (P) and the amniotic cavity (AC); the procedure takes between 30 s and 1 min. YS = yolk sac; D = decidua; M = maternal tissue. Coelocentesis has a success rate of >95% at 6–11 weeks gestation. In theory, it is the ideal alternative to early amniocentesis and chorion villous sampling (CVS), because the risk of directly injuring the growing embryo or damaging its placenta is very low (Jurkovic et al., 1993). Furthermore, the procedure is easy to learn, induces only minimal discomfort to the mother and is associated with a very low rate of contamination of the sample by maternal cells (Jurkovic et al., 1995). However, the only study, so far, evaluating the safety of CF aspiration in ongoing pregnancies, has shown that the risk of miscarriage after coelocentesis is ~25% (Ross et al., 1997). This finding and the high failure rate of cell growth from CF currently limits the application of coelocentesis to exploring the biology of materno– embryonic exchanges at a time of gestation when fetal blood can not be obtained. The main findings of these studies and their contribution to our understanding of embryonic physiology are the basis of this review. Embryology of the human placenta and adnexae The formation of the placenta begins 13–15 days after ovulation, corresponding to stage 6 of embryonic development and to the end of the fourth week after the last menstrual period (Boyd and Hamilton, 1970). The primary villi are composed of a central mass of cytotrophoblast surrounded by a thick layer of syncytiotrophoblast. During the following week of gestation, they acquire a central mesenchymal core from the extraembryonic mesoderm and become branched, forming the secondary villi. The appearance of embryonic blood vessels within the mesenchymal core transforms the secondary villi into tertiary villi. At the end of the fifth gestational week, all three primitive types of placental villi can be found but tertiary villi progressively predominate. Up to the 9–10th week post-menstruation, which corresponds to the last week of the embryonic period (stages 19–23), villi cover the entire surface of the chorionic sac (Figure 2). As the gestational sac grows during fetal life, the villi associated with the decidua capsularis (surrounding the amniotic sac) degenerate to form the chorion laeve, whereas the villi associated with the decidua basalis proliferate, forming the chorion frondosum or definitive placenta (Jauniaux et al., 1992). The extraembryonic coelom or exocoelomic cavity develops during the fourth week after the last menstrual period (Boyd and Hamilton, 1970). It surrounds the blastocyst which is composed of two cavities separated by the bilaminar embryonic disk, i.e. the amniotic cavity and the primary yolk sac (Figure 3). At the end of the fourth week of gestation, the developing exocoelomic cavity 270 E.Jauniaux and B.Gulbis Figure 2. Diagrams showing the different anatomical barriers inside the first trimester gestational sacs. U = uterus; P = placenta; UC = umbilical cord; ECC = exocoelomic cavity; SYS = secondary yolk sac; AC = amniotic cavity; AM = amniotic membrane. placental chorionic plate at the end of the first trimester (Boyd and Hamilton, 1970). The amniotic cavity is smaller than the exocoelomic cavity up to 9 weeks gestation. Thus, during the second and third month of pregnancy, the embryo and subsequently the fetus are surrounded by the amniotic cavity which is, in turn, surrounded by the exocoelomic cavity containing the secondary yolk sac. The secondary yolk sac is an independent organ floating inside the exocoelomic cavity (Figure 3). It forms at the beginning of the fifth week post-menstruation and develops rapidly so that by the 37th menstrual day it is larger than the amniotic cavity (Boyd and Hamilton, 1970). From the sixth week of gestation it appears as a spherical and cystic structure covered by numerous superficial small vessels merging at the basis of the vitelline duct. This connects the yolk sac to the ventral part of the embryo, the gut and main blood circulation (Figure 4). The wall of the secondary yolk sac is formed by an external mesothelial layer facing the extraembryonic coelom, a vascular mesenchyme and an endodermal layer facing the yolk sac cavity. The extraembryonic human circulation is first established within the vitelline duct artery via the dorsal aorta (Jones and Jauniaux, 1995). During the 10th week of gestation the yolk sac starts to degenerate and rapidly ceases to function (Jones and Jauniaux, 1995). Embryonic physiology Figure 3. Schematic representations of human pregnancies at the beginning (A) and at the end (B) of the 4th menstrual week and during the fifth (C) and the sixth (D) menstrual week. splits the extraembryonic mesoderm into two layers, the somatic mesoderm, lining the trophoblast and the splanchnic mesoderm covering the secondary yolk sac and the embryo (Figure 3). There is no anatomical barrier between the mesenchyme of the placental fetal plate and the exocoelomic or chorionic cavity (Jones and Jauniaux, 1995). At ~31 days menstrual age, the gestational sac is 2–3 mm in diameter and can be detected by means of transvaginal ultrasound imaging. The amniotic cavity develops during the third week of pregnancy from the inner cell mass of the implanted blastocyst and grows inside the extraembryonic coelom fusing with the Human embryonic physiology and developmental biology are treated only incidentally in classical embryology textbooks and have been overshadowed by the extraordinary profusion of anatomical descriptions of embryos and early fetuses (Jauniaux and Gulbis, 1997). Over the last decade, developmental biology has become one of the leading fields of fundamental research. However, the developmental physiology of most embryonic organs remains largely unknown because of the limited access to these organs for in-vivo and in-vitro experimentation. Since the placenta and its membranes are larger than the fetus up to midpregnancy and, therefore, more accessible for research, it is not surprising that most of our knowledge on the physiology of the embryo and early fetus is essentially that of its adnexae which provide the habitat in which the embryo and its functions develop. Human placentation is theoretically haemochorial and mainly characterized by diffuse infiltration by extravillous trophoblastic cells of the uterine endometrium and superficial myometrium. Classically, immediately after implantation, a number of endometrial vessels are opened by the phagocytic activity of the extravillous trophoblastic cells and the maternal circulation starts in the intervillous space (Ramsey and Donner, 1980). This dogma has been challenged by the data of Hustin et al. which show that during the first trimester of pregnancy, the intervillous space of the definitive placenta is separated from the uterine circulation by trophoblastic plugs obliterating the tip of the uteroplacental arteries (Hustin and Schaaps, 1987; Hustin et al., 1988). At the end of the first trimester, these plugs are progressively dislocated allowing maternal blood to flow freely and continuously into the intervillous space. In-vivo measurements of oxygen concentrations in early human pregnancy have shown that the placental oxygen pressure is 2–3 times lower at 8–10 weeks than after 12 weeks (Rodesch et al., Embryonic fluid compartments 271 Figure 4. Photograph of an embryo and its yolk sac at 8 + 4 weeks gestation. The yolk sac shows a honeycomb pattern and is covered by numerous small vessels merging at the basis of the vitelline duct. 1992). Furthermore, there is mounting evidence that early placental trophoblast function and development is influenced by oxygen tension (Genbacev et al., 1996, 1997). The results of our recent experiments on trophoblast antioxidant defence are consistent with the proposal that the placenta develops in a physiologically low oxygen environment during the early part of gestation (Watson et al., 1997, 1998a,b). The investigation of the effect of oxygen metabolites on embryonic development is likely to become an important field of research during the next decade (Umaoka et al., 1992; Parman et al., 1999). Biology of the extraembryonic coelom Coelomic fluid (CF) has a lower pH, base excess and bicarbonate value than maternal venous blood and has a higher carbon dioxide pressure, higher lactate and phosphate concentrations and lower protein concentrations than the maternal serum. These findings are consistent with a metabolic anaerobic acidosis status which is mainly due to the accumulation of acidic byproducts from the placental metabolism in the exocoelomic cavity (Jauniaux et al., 1994a). As a consequence of respiratory alkalosis of pregnancy, maternal renal excretion of bicarbonate secondarily increases and the overall maternal blood pH remains relatively unchanged (Blackburn and Loper, 1992). Except for the total protein and lactate concentrations, there is no significant variation in the coelomic fluid biochemical composition at 7–11 weeks gestation and no correlation of the different variables between coelomic fluid and maternal serum, suggesting that the coelomic fluid acid– base regulation is not directly influenced by maternal physiological changes. For the duration of the first trimester, the CF is yellow coloured and more viscous than the AF, which is always clear. This is mainly due to a higher protein concentration in CF than in AF. We found that at 6–12 weeks gestation, the mean total protein concentration is 18 times lower in the CF than in maternal serum but 54 times higher in CF than in AF (Jauniaux et al., 1993). In fact, the concentration of almost every protein is higher in CF than in AF, ranging between 2 and 50 times depending on the corresponding molecular weight of the protein investigated (Table I). Small molecules such as urea can easily cross most plasma membrane or tissue and it is, therefore, not surprising that no significant difference was observed between urea concentration in maternal serum and both embryonic compartments (Table I). However, a gradient is observed for slightly larger molecules, e.g. creatinine. These findings indicate that transfer through the amniotic membrane separating the two embryonic fluid cavities is limited. There is an increase of most protein concentrations observed in the exocoelomic cavity during the first trimester (Figure 5). This can be explained by the slow turnover of the coelomic fluid and/or the increased production of these proteins by fetal organs. In contrast, there is a physiological decrease in total maternal serum protein which occurs mainly during the first 3 months of gestation, ranging from 10 to 14% of non-pregnant values (Blackburn and Loper, 1992). During the first trimester, albumin concentration in maternal serum demonstrates a relative decrease due to increased maternal blood volume and haemodilution, while globulin and fibrinogen concentrations demonstrate both absolute and relative increases. During the same period, the amounts of total protein and pre-albumin, increase in CF whereas the amounts of albumin do not change significantly (Jauniaux et al., 1994b). There is no difference in crown–rump length (CRL), yolk sac volume and the concentration of protein in the CF between mothers with low serum pre-albumin concentrations and mothers with high serum 272 E.Jauniaux and B.Gulbis Table I. Concentration of various proteins and other molecules in embryonic fluids and maternal serum according to their main site of production or origin during the first trimester of pregnancy Molecules Maternal serum Coelomic fluid Amniotic fluid Total protein (g/l)a 71.3 3.5 0.2 Creatinine (µmol/l)a 50.1 43.6 27.7 Urea (mmol/l)a 7.2 8.3 7.2 Albumin (g/l)b 45.5 1.7 ND Pre-albumin (g/l)b 1.14 0.04 ND Tyroxine (nmol/l)c 180 0.9 0.02 Relaxin (ng/l)d 1000 122 9 Immunoglobulin G (mg/dl)e 907 32 3 e ND Mother 122 1 Complement factors 3 (mg/dl)e 114 ND ND Complement factors 4 (mg/dl)e 21 ND ND Iron (µmol/l) f 21 4.8 1.8 Glucose (mmol/l)g 3.4 2.7 2.8 IGF-I (µg/l)h 233 41 38 1057 Immunoglobulin A (mg/dl) Villous tissue Intact HCG (mIU/ml)i 80193 105605 (mIU/ml)i 70 11200 169 Free β-HCG (mIU/ml)i 45 1478 20 hPL (ng/ml)j 210 80 30 Progesterone (pg/ml)k 17 240 8 Oestradiol (pg/ml)k 917 8469 1898 Activin A (ng/ml)l 0.68 0.98 0.09 Inhibin B (pg/ml)l 5.9 24.3 6.3 β2-microglobulin (mg/l)m 0.9 4.7 N/D 0.3 0.6 0.9 687 199 40 Vitamin B12 (ng/l)n 405 3680 987 Prolactin (mIU/l) 709 371 40 Placental protein 14 (µ g/l)j 642 4416 77 Interleukin-6 (ng/ml)p 40 88 17 IGFBP-1 (µg/l)h 76 150 16 123 167 49 AFP (kIU/l)k 1.4 21816 27096 Erythropoietin (mIU/ml)q 15.4 15.5 5.0 τ-glutamyltransferase (IU/l)m 9 2 25 Ferritin (µg/l)f 49 287 2.0 Cancer antigen 125 (IU/ml)r 35 35 496 Free α-HCG Lactate (mmol/l) g IGF-II (µg/l)h Decidua IGFBP-2 (µg/l) h Secondary yolk sac Embryo/fetus ND = not detectable; IGF = insulin-like growth factors; IGFBP = insulin-like growth factor binding proteins; HCG = human chorionic gonadotrophin; AFP = α-fetoprotein. a Mean value (Jauniaux et al., 1991); bmean value (Jauniaux et al., 1994b); cmean value (Contempre et al., 1993); dmedian value (Johnson et al., 1994); emedian value (Jauniaux et al., 1995a); fmedian value (Gulbis et al., 1994); gmean value (Jauniaux et al., 1994a); hmean value (Miell et al., 1997); imean value (Jauniaux et al., 1995b); jmedian value (Wathen et al., 1992); kmean value (Jauniaux et al., 1993); lmedian value (Luisi et al., 1998); mmedian value (Gulbis et al., 1996); nmedian value (Campbell et al., 1992a); omedian value (Wathen et al., 1993); pmedian value (Jauniaux et al., 1996a); qmedian value (Campbell et al. , 1992b); rmedian value (Campbell et al., 1992c). Embryonic fluid compartments Figure 5. Diagram showing the changes of total protein concentration with gestational age in coelomic fluid (squares) and amniotic fluid (triangles). Note the linear increase in coelomic fluid concentration at 8–12 weeks and the exponential increase in amniotic fluid concentration after 12 weeks. pre-albumin concentrations. These results suggest that the total protein concentration in CF is not directly influenced by changes in maternal serum protein concentrations during the first trimester. Fetal growth and development are closely dependent on the availability of a constant supply of amino acids from the mother for protein synthesis (Blackburn and Loper, 1992). The trophoblast functions of nutrient transport and protein synthesis generate high concentrations of amino acids in the placenta and in fetal blood during the second half of pregnancy (Sibley and Boyd, 1992). Significant positive relationships between maternal serum and placental tissue are found for several amino acids indicating that active amino acid transport and accumulation by the human syncytiotrophoblast occurs as early as 7 weeks gestation (Jauniaux et al., 1998a). The transplacental flux of most amino acid transport from the maternal blood to the exocoelomic cavity is against a concentration gradient (Jauniaux et al., 1994c). The concentration distribution of individual amino acids in coelomic and amniotic fluid are related indicating a passive transfer through the amniotic membrane for these small molecules. A coelomic–maternal gradient is observed for most amino acids measured and positive correlations are found between maternal serum and CF for concentration of α-aminobutyric acid, tyrosine and histidine suggesting that these amino acids are only partially retained and/ or are transferred more rapidly by the early placenta. Molecules such as thyroid hormones, immunoglobulins (Ig), complement factors, relaxin or iron are not synthesized by the feto–placental unit during the first trimester but play an essential role in fetal development. These molecules are detectable in CF indicating materno–embryonic transfer, probably from when the tertiary placental villi are formed. In particular, thyroxine (T4) and 3′,5′,3-triiodothyronine (rT3) have been found in CF samples suggesting that maternal thyroid hormones are potentially available to the embryo as early as 5 weeks gestation (Contempre et al., 1993). Another example are the immunoglobulins; trace values of IgG and IgM have only been found in cultures of human fetal liver and spleen from the end of the third trimester and IgA synthesis has not been demonstrated in vitro until after 30 weeks gestation (Gitlin and Biasucci, 1969). Only very low amounts of IgG and IgM can be detected in the plasma of 12–14 week fetuses (Gitlin, 1984). The concentration of IgG, including IgG specific 273 against Toxoplasma gondii, cytomegalovirus and rubella virus and IgA values are measurable in CF samples from 6 weeks gestation whereas IgM is not (Jauniaux et al., 1995a). This suggests that the placental transfer of IgG and IgA begins very early in pregnancy. IgG and IgA molecules in CF could play a role in the initial antigenic challenge of blood cell precursors to early congenital infection and in limiting the potentially devastating effects of congenital infections. Using the same methodology, placental iron transfer has been demonstrated from 7 weeks gestation and iron was found to accumulate in the exocoelomic cavity (Gulbis et al., 1994). The distribution of iron and iron-binding proteins between the maternal and embryo–placental compartments in the first trimester is comparable with that found later in gestation (Figure 6). The trophoblast produces a variety of specific proteins such as human chorionic gonadotrophin (HCG), human placental lactogen (hPL), activin A or inhibin which are excreted in both maternal and embryonic fluid compartments (Table I). The higher concentrations of these molecules found in CF compared with maternal serum can be explained by the close anatomical relationship existing between the exocoelomic cavity and the trophoblast as both structures are only separated by the loose mesenchymal tissue of the chorionic plate (Jones and Jauniaux, 1995). An exception to this principle is pregnancy-associated protein A (PAPP-A) which is theoretically synthesized by the villous tissue but found in higher concentration in maternal serum than in CF (Iles et al., 1994). Free α-HCG and free β-HCG concentrations are 185 and 33 times higher respectively in the CF of normal pregnancy than in the corresponding maternal serum samples (Jauniaux et al., 1995b). This finding supports the hypothesis that, in the first trimester, there is an excess of α- over β-subunit secretion by the villous trophoblast (Nagy et al., 1994) and confirms that the HCG clearance rate is slower in the exocoelomic cavity than in maternal circulation. It is likely that maternal serum HCG concentrations are influenced by both villous and extravillous trophoblastic synthesis whereas (the exocoelomic cavity being completely surrounded by villous tissue), coelomic HCG concentrations are only influenced by villous trophoblastic secretion. In view of the large amount of free α-HCG that is present in CF and the observation that free αHCG can stimulate decidual prolactin secretion in vitro (Blithe and Iles, 1995), it is likely that this high concentration of placental protein in CF has a regulatory effect on the endocrine function of the materno–placental interface. In contrast to maternal serum, CF concentrations of intact HCG and free α-HCG decrease progressively at 8–12 weeks gestation and free β-HCG values do not vary (Jauniaux et al., 1995b). Oestradiol and progesterone concentrations also decrease in coelomic fluid between the second and the third month of gestation whereas the opposite was true in maternal serum. These changes in CF hormonal concentrations are probably secondary to a decrease in the exchange surface, as two thirds of the primitive placental ring start to degenerate during the third month of gestation (Jauniaux et al., 1993). The decrease in intact HCG and free α-HCG coelomic values with advancing gestation may also be secondary to a simultaneous decline in the number of differentiating cytotrophoblastic cells and/or to the disappearance of two thirds of the original placental tissue which takes place during the same period. 274 E.Jauniaux and B.Gulbis Figure 6. Distribution of iron and iron-binding proteins between the maternal and embryo–placental compartments during the first trimester of pregnancy (modified from Gulbis et al., 1994). MS = maternal serum; ECF = extracoelomic fluid; AF = amniotic fluid. T bars indicate SD. Molecules, e.g. vitamin B12, prolactin and placental protein 14 (PP14), are known to be mainly produced by the uterine decidua (Table I). They are often found at higher concentrations in CF than in maternal serum, suggesting preferential pathways between the decidual tissue and the embryonic fluid cavities via the villous trophoblast. This mechanism may be a pivotal in providing the developing embryo with sufficient nutrient before the intervillous circulation becomes established. Molecules such as insulin-like growth factors (IGFs) and their binding proteins (IGFBPs) are also important in fetal growth and are produced by various maternal and fetal tissues. IGF-I and II concentrations are highest in maternal serum and low in CF and AF (Miell et al., 1997). IGFBP-1 concentrations are higher in CF than either maternal serum or AF and show a significant correlation to gestational age. Analysis of IGFBP-1 phosphoforms show clear differences in phosphorylation of IGFBP-1 between compartments with maternal serum containing predominantly the phosphorylated forms and CF almost exclusively the non-phosphorylated forms. These findings suggest that the high IGF-II concentrations and lack of inhibitory phosphoforms of IGFBP-1 in CF could potentially enhance mitogenic activity in the early human gestational sac (Miell et al., 1997). The endodermal layer of the secondary yolk sac is known to synthesize several serum proteins in common with the fetal liver, e.g. α-fetoprotein (AFP), α1-antitrypsin, albumin, pre-albumin and transferrin (Gitlin and Perricelli, 1970; Shi et al., 1985). With rare exceptions, the synthesis of most of these proteins is confined to embryonic compartments and the contribution of the yolk sac to the maternal protein pool is limited. This can explain why their concentration is always higher in embryonic fluids than in maternal serum (Tables I and II). AFP is also produced by the embryonic liver from 6 weeks until delivery, has a high molecular Table II. Embryonic fluids and maternal blood gases, acid–base values and electrolyte concentration (modified from Jauniaux et al., 1994) Variable Maternal blood Coelomic fluid Amniotic fluid pH 7.38 7.18 7.42 CO2 pressure (mmHg) 43 56 53 Base excess (mmol/l) –2.6 –7.8 10.2 Bicarbonate (mmol/l) 22 18 38 Chloride (mmol/l) 105 105 92 Potassium (mmol/l) 4.0 3.9 3.6 Sodium (mmol/l) 134 134 131 Phosphate (mmol/l) 1.01 2.1 0.71 weight (±70 kDa) and conversely to HCG was found in similar amounts on both sides of the amniotic membrane (Table I). Analysis of concanavalin A affinity molecular variants of AFP have demonstrated that both exocoelomic fluid and amniotic fluid AFP molecules originated mainly from albumin the yolk sac while maternal serum AFP molecules came mainly from the fetal liver (Jauniaux et al., 1993). These results suggest that the human secondary yolk sac also has an excretory function and secretes AFP into the embryonic and extraembryonic compartments (Figure 7). The potential absorptive role of the yolk sac membrane has been recently evaluated by examining the distribution of proteins and enzymes between CF and yolk sac fluid and by comparing the synthesising capacity of secondary yolk sac, fetal liver and placenta for HCG and AFP (Gulbis et al., 1998). The distribution Embryonic fluid compartments Figure 7. Percentage of Con A non-reactive α-fetoprotein (AFP) [% Con A(–)] from exocoelomic fluid (ECF), amniotic fluid (AF), maternal serum (MS), fetal liver, and yolk sac collected during the first trimester of gestation (From Jauniaux et al., 1993). of the placental-specific protein, HCG, in yolk sac fluid and CF and the absence of HCG mRNA expression in yolk sac tissue has provided the first biological evidence of its absorptive function (Figure 8). Similarities in the composition of the yolk sac and CF fluid suggest that there is a free transfer for most molecules between the two compartments through the layers of the dividing wall. The yolk sac lumen contains digestive enzymes which were not found inside CF but are present (in increasing concentrations as pregnancy advances) in the amniotic cavity. These findings suggest that the yolk sac membrane is an important zone of transfer between the extraembryonic and embryonic compartments and that the main flux of molecules occurs from outside the yolk sac, i.e. from the exocoelomic in the direction to the lumen and subsequently to the embryonic gut and circulation. When, after 10 weeks gestation, the cellular components of the wall of the secondary yolk start to degenerate, this route of transfer is no longer functional and most exchanges between the exocoelomic cavity and the fetal circulation must then take place at the level of the chorionic plate. Biology of the early amniotic cavity During the first trimester, the amniotic membrane floats freely between the embryonic cavities. Despite its apparent simplicity (Jones and Jauniaux, 1995), direct transfer from the exocoelomic to the amniotic cavity via the amniotic membrane is limited and the AF contains very low protein concentrations (Gulbis et al., 1992, Jauniaux et al., 1993). The total AF protein concentration is 50 and 900 times lower than in CF and maternal serum (Table I) respectively. Almost all individual proteins, except AFP, are present at very low concentrations in the AF. The vitelline duct has the same cellular constitution as the secondary yolk sac (Jones and Jauniaux, 1995). AFP and other yolk sac proteins found in the early AF could be excreted at the level where the duct fuses with the primitive umbilical cord. Yolk sac AFP could also be moved in AF via the vitelline duct and from 10 weeks post-menstruation when the anal membranes break down, intestinal AFP is also found in AF (Boyd and Hamilton, 1970). During the second and 275 Figure 8. Expression of β-human chorionic gonadotrophin (β-HCG), α-fetoprotein (αFP) and β-actin mRNAs in four series of placenta (P), yolk sac (Y), and fetal liver (L) matched tissue samples. (1 = 8 weeks gestation; 2 = 9 ωεεκσ gestation; 3 = 7 weeks gestation; 4 = 12 weeks gestation) (from Gulbis et al., 1998). third trimester, the pool of AF is subject to a constant turnover, with the accumulation of fetal lung fluid and urine and removal by fetal swallowing (MacCarthy and Saunders, 1978). When the definitive placenta has formed, movement of water and electrolytes also takes place across the free placental membranes. AF samples, collected before 11 weeks gestation, have a higher pH and base excess, higher concentrations of lactate and bicarbonate and lower concentrations of total protein, phosphate, chloride, sodium and potassium than the CF collected during the same period of gestation (Jauniaux et al., 1994a). Of these biological parameters, only the pH and the bicarbonate levels varied at 7–11 weeks gestation (Table II). The metabolic alkalosis of first-trimester AF probably results from the accumulation of bicarbonate and the increased consumption of organic anions such as lactate by the embryonic tissue. The embryonic skin, which only becomes keratinized during the second trimester, is probably the major source of AF in early pregnancy (MacCarthy and Saunders, 1978). These results indicate that in contrast to the CF composition which is mainly influenced by placental and yolk sac bioproducts, the first trimester AF composition is mainly influenced by fetal bioproducts which may diffuse through the fetal skin or through the oropharyngeal and cloacal membranes (Jauniaux et al., 1994a; Gulbis et al., 1996). The latter rupture around the end of the fifth and seventh week of gestation respectively, allowing free circulation between fetal digestive and respiratory tracts and the amniotic cavity (O’Rahilly and Muller 1992). Fetal urine is a major source of AF in the latter half of pregnancy. AF electrolyte composition, protein patterns and acid– base balance change rapidly at the end of the first trimester (Gulbis et al., 1992, 1996; Jauniaux et al., 1993, 1994a). Various mechanisms should be considered in order to explain these changes, in particular, the metabolic activity of the definitive kidneys, the lungs and the digestive tract. The development of nephrons starts around the beginning of the third month of 276 E.Jauniaux and B.Gulbis Table III. Mean concentration of various drugs in maternal serum and embryonic fluids 5–25 min after a single i.v. bolus (Diazepam 0.1 mg/kg, Fentanyl 1.5 µg/kg, Propofol 3 mg/kg, Inulin 5 mg/kg) or after chronic intake (Cotinine) Drugs Reference Maternal serum Coelomic fluid Amniotic fluid Diazepam (ng/ml) Jauniaux et al., 1996b 189 6.9 7.4 Fentanyl (ng/ml) Shannon et al., 1998 1.3 ND 1.1 Propofol (µg/ml) Jauniaux et al., 1998b 1.96 ND ND Inulin (mg/ml) Jauniaux et al., 1997b 6.9 5.1 3.0 Cotinine (ng/ml) Jauniaux et al., 1999b 72 99 108 ND = not detectable. gestation and, theoretically, the metanephros or definitive kidney produces urine from 10 weeks onwards (O’Rahilly and Muller 1992). At ~11 weeks, we have observed an abrupt increase in β2-microglobulin and τ-glutamyl transferase (τGT) AF concentrations and it is possible that this reflects the maturation of the fetal renal glomerular function (Gulbis et al., 1996). In particular, β2-microglobulin concentrations found in the amniotic cavity during the second trimester are linked with the establishment of glomerular filtration in the definitive fetal kidneys at a time when the tubular function is still immature. The AF β2-microglobulin concentration decreases during the third trimester as a consequence of the increasing reabsorption capacity of proximal tubular cells. The very low activity of τGT in CF suggests that the placental villi are not a main source of this enzyme. The changes observed in AF composition at the end of the first trimester are characterized by a decrease in pH, base excess and bicarbonate values and an increase in carbon dioxide pressure and chloride concentrations. It is of interest that, except for the total protein values which remained lower, the mean value of the other amniotic biological parameters obtained at >11 weeks gestation were similar to those found in CF at <11 weeks (Jauniaux et al. 1994a). Similar changes occur during the second and third trimesters (Sinha and Carlton, 1970) and probably reflect the increasing contribution of the different fetal organs to AF composition. Prospects in the investigation of the biology of embryonic fluids The importance of the materno–embryonic exchange system in humans has previously been difficult to assess, due to the difficulties involved in accessing embryonic fluid or blood. Investigation of the various biochemical constituents in embryonic fluids have demonstrated that the exocoelomic cavity is not a virtual space, and that it is the site of important molecular exchanges between the mother and the embryo. Thus the study of the different molecular concentrations in embryonic fluid has provided useful information for understanding the transfer pathways between the mother and the embryo (Table I). However, these studies were purely descriptive and more studies comparing the CF concentrations of a molecule with its quantifiable presence in decidual, placental and fetal tissues (Gulbis et al., 1994, 1998; Jauniaux et al., 1996a; 1998b; Riley, 1999) are needed. CF can be collected in cases of early pregnancy failure. The CF concentration of total protein is extremely low in all missed abortions with advanced trophoblastic necrosis, whereas the HCG concentration is low only when there are no embryonic remnants on ultrasound (Jauniaux et al., 1995c). Normal or high maternal serum AFP values and AFP molecules (predominantly of yolk sac origin in the CF of pregnancies with an empty gestational sac on ultrasound) provide further evidence that the most likely explanation for this feature is the early death of the embryo with persistence of the placental tissue (Jauniaux et al., 1995c). Thus, the vast majority of pregnancies traditionally classified as anembryonic gestation, in fact result from early embryonic demise. The embryo having developed for at least 14 days after ovulation corresponds to the stage of embryonic life when the secondary yolk sac starts to form. Similar studies are needed to better understand the pathophysiology of early pregnancy failure. Placental physiology is notoriously difficult to study in vivo. In-vitro models have been extensively used and most information has been obtained from the placenta at term. The presence of the exocoelomic cavity between the placenta and the amniotic sac containing the fetus, the disappearance of two thirds of the placental tissue mass and the major changes in both maternal and fetal circulations at the end of the first trimester, make comparisons between first and third trimester placental physiology almost impossible. The unique anatomical position of the exocoelomic cavity, which in primates is in direct contact with the mesenchyme of the placental villi, opens new possibilities to perform in vivo, physiological experiments traditionally performed in vitro. Catheters (Ward et al., 1998) and sensors (Jauniaux et al., 1999a) can be inserted, under ultrasound guidance, and used to gain some insight into in-vivo first-trimester placental physiology. Placental transfers have been mainly studied in experimental animals such as guinea pigs and monkeys because they have haemochorial placentas similar to those of humans (Burton, 1992). In lower vertebrates, the yolk sac serves as the principal membrane for placental exchanges. In particular, rodents have a subsidiary yolk sac placenta which completely envelops the fetus throughout the whole gestation and serves as the principal site for the acquisition of protein by the fetus. Due to the complexity of the experimental situation in the intact animal and the Embryonic fluid compartments considerable embryological differences between human and some animal species, the precise mechanism of transfer of many substances across the human placenta has not been elucidated. However, the selective sampling of fluid from the exocoelomic cavity has also offered a novel approach to the study of drug transfer across the early human placenta (Table III). So far, these studies have demonstrated that the permeability of the placenta is greater in early pregnancy than at term but also because of the slow turnover of CF substances to which the mother is chronically exposed are likely to accumulate inside the exocoelomic cavity (Jauniaux and Gulbis, 1998). This prolonged exposure to toxins such as tobacco carcinogens (Jauniaux et al., 1999b,c) has important teratological implications and should be further explored. References Blackburn, S.T. and Loper, D.L. (1992) Maternal, Fetal and Neonatal Physiology: A Clinical Perspective. Saunders, Philadelphia, USA. Blithe, D.L. and Iles, R.K. (1995) The role of glycosylation in regulating the glycoprotein hormone free alpha-subunit and free beta-subunit combination in the extraembryonic coelomic fluid of early pregnancy. Endocrinology, 136, 903–910. Boyd, J.D. and Hamilton, W.J. (1970) The Human Placenta. Heffer, Cambridge, UK. Burton, G.J. (1992) Human and animal models: Limitations and comparisons. In Barnea, E., Hustin, J. and Jauniaux, E. (eds), The First Twelve Weeks of Gestation. Springer-Verlag, Heidelberg, Germany, pp. 469–485. Campbell, J., Wathen, N., Perry, G. et al. (1992a) The coelomic cavity: an important site of materno–fetal nutrient exchange in the first trimester of pregnancy. Br. J. Obstet. Gynaecol., 100, 765–767. Campbell, J., Wathen, N., Lewis, M. et al. (1992b) Erythropoietin levels in amniotic fluid and extraembryonic coelomic fluid in the first trimester of pregnancy. Br. J. Obstet. Gynaecol., 9, 974–976. Campbell, J., Kitau, M., Cass, P. et al. (1992c) CA 125 in matched samples of amniotic fluid, extra-embryonic coelomic fluid and maternal serum in the first trimester of pregnancy. J. Obstet. Gynaecol., 100, 563–565. Contempre, B., Jauniaux, E., Calvo, R. et al. (1993) Detection of thyroid hormones in human embryonic cavities during the first trimester of pregnancy. J. Clin. Endocrinol. Metab., 77, 1719–1722. Genbacev, O., Joslin, R., Damsky, C.H. et al. (1996) Hypoxia alters early gestation human cytotrophoblast differenciation/invasion in vitro and models the placental defects that occur in preeclampsia. J. Clin. Invest., 97, 540–550. Genbacev, O., Zhou, Y., Ludlow, J.W. and Fisher, S.J. (1997) Regulation of human placental development by oxygen tension. Science, 277, 1669– 1672. Gitlin, D. (1984) Protein transport across the placenta and protein turnover between amniotic fluid, maternal and fetal circulation. In Moghissi, K.S. and Hafez, E.S.E. (eds), The Placenta. Charles C.Thomas, Springfield, USA, pp. 151–72. Gitlin, D. and Biasucci, A. (1969) Development of τG, τA, τM, β1C/β1a, C′1 esterase inhibitor, ceruloplasmin, haptoglobin, fibrinogen, plasminogen, α1-antitrypsin, orosomucoid, β-lipoprotein, α2-macroglobulin and prealbumin in the human conceptus. J. Clin. Invest., 48, 1433–1446. Gitlin, D. and Perricelli, A. (1970) Synthesis of serum albumin, prealbumin, alphafetoprotein, alpha1-antitrypsin and transferrin by the human yolk sac. Nature, 228, 995–997. Gulbis, B., Jauniaux, E., Jurkovic, D. et al. (1992) Determination of protein pattern in embryonic cavities of early human pregnancies: A model to understand materno–embryonic exchanges. Hum. Reprod., 7, 886–889. Gulbis, B., Jauniaux, E., Decuyper, J. et al. (1994) Distribution of iron and iron-binding proteins in first trimester human pregnancies. Obstet. Gynecol., 84, 289–293. Gulbis, B., Jauniaux, E., Gervy, C. et al. (1996) Biochemical investigation of kidney functional maturation in early human pregnancy. Pediatr. Res., 39, 1–5. Gulbis, B., Jauniaux, E., Cotton, F. and Stordeur, P. (1998) Protein and enzyme patterns in the fluid cavities of the first trimester gestational sac: 277 relevance to the absorptive role of secondary yolk sac. Mol. Hum. Reprod., 4, 857–862. Hustin, J, and Schaaps, J.P. (1987) Echographic and anatomic studies of the maternotrophoblastic border during the first trimester of pregnancy. Am. J. Obstet. Gynecol., 157, 162–168. Hustin, J., Schaaps, J.P., and Lambotte, R. (1988) Anatomical studies of the utero–placental vascularization in the first trimester of pregnancy. Trophoblast Res., 3, 49–60. Iles, R.K., Wathen, N.C., Sharma, K.B. et al. (1994) Pregnancy-associated plasma protein A levels in maternal serum, extraembryonic coelomic and amniotic fluids in the first trimester. Placenta, 15, 693–699. Jauniaux, E. and Gulbis, B. (1997) Embryonal physiology. In Jauniaux, E., Barnea, R., Edwards, R. (eds) Embryonic Medicine and Therapy. Oxford University Press, Oxford, UK, pp. 223–243. Jauniaux, E. and Gulbis, B. (1998) In-vivo study of placental drug transfer during the first trimester of human pregnancy. Trophoblast Res., 12, 257– 264. Jauniaux, E., Jurkovic, D., Gulbis, B. et al. (1991) Biochemical composition of coelomic fluid in early human pregnancy. Obstet. Gynecol., 78, 1124– 1128. Jauniaux, E., Burton, G.J. and Jones, C.P.J. (1992) Early human placental morphology. In Barnea, E., Hustin, J. and Jauniaux, E. (eds), The First Twelve Weeks of Gestation. Springer-Verlag, Heidelberg, Germany, pp. 45–64. Jauniaux, E., Gulbis, B., Jurkovic, D. et al. (1993) Protein and steroid levels in embryonic cavities of early human pregnancy. Hum. Reprod., 8, 782–787. Jauniaux, E., Jurkovic, D., Gulbis, B. et al. (1994a) Investigation of the acid– base balance of coelomic and amniotic fluids in early human pregnancy. Am. J. Obstet. Gynecol., 170, 1359–1365. Jauniaux, E., Gulbis, B., Jurkovic, D. et al. (1994b) Relationship between protein levels in embryological fluids and maternal serum and yolk sac size during early human pregnancy. Hum. Reprod., 9, 161–166. Jauniaux, E., Sherwood, R., Jurkovic, D. et al. (1994c) Amino acids concentration in human embryological fluids. Hum. Reprod., 9, 1175– 1179. Jauniaux, E., Jurkovic, D., Gulbis, B. et al. (1995a) Materno–fetal immunoglobulin transfer and passive immunity during the first trimester of human pregnancy. Hum. Reprod., 10, 3297–3300. Jauniaux, E., Gulbis, B., Nagy, A.M. et al. (1995b) Coelomic fluid chorionic gonadotropin and protein levels in normal and complicated first trimester human pregnancies. Hum. Reprod., 10, 214–220. Jauniaux, E., Gulbis, B., Jurkovic, D. et al. (1995c) The origin of alphafetoprotein in first trimester anembryonic pregnancies. Am. J. Obstet. Gynecol., 173, 1749–1753. Jauniaux, E., Gulbis, B., Schandene, L. et al. (1996a) Distribution of interleukin-6 in maternal and embryonic tissues during the first trimester. Mol. Hum. Reprod., 2, 239–243. Jauniaux, E., Jurkovic, D., Lees, C. et al. (1996b) In vivo study of diazepam transfer across the first trimester human placenta. Hum. Reprod., 11, 889– 892. Jauniaux, E., Gulbis, B., Gerlo, E. and Rodeck, C.H. (1998a) Free amino acid distribution in the first trimester human gestational sac. Early Hum. Dev., 51, 159–169. Jauniaux, E., Gulbis, B., Shannon, C. et al. (1998b) Placental propofol transfer and fetal sedation during maternal general anaesthesia in early pregnancy. Lancet, 352, 290–291. Jauniaux, E., Watson, A., Ozturk, O. et al. (1999a) In vivo measurement of intrauterine gases and acid–base values in early human pregnancy. Hum. Reprod., 14, 2901–2904. Jauniaux, E., Gulbis, B., Acharya, G. et al. (1999b) Maternal tobacco exposure and cotinine levels in fetal fluids in the first half of pregnancy. Obstet. Gynecol., 93, 25–29. Jauniaux, E., Gulbis, B., Acharya, G. and Thiry, P. (1999c) Fetal amino acid and enzymes levels with maternal smoking. Obstet. Gynecol., 93, 680– 683. Jones, C.P.J. and Jauniaux, E. (1995) Ultrastucture of the materno–embryonic interface in the first trimester of pregnancy. Micron, 2, 145–173. Jurkovic, D., Jauniaux, E., Campbell, S. et al. (1993) Coelocentesis: a new technique for early prenatal diagnosis. Lancet, 341, 1623–1624. Jurkovic, D., Jauniaux, E., Campbell, S. et al. (1995) Detection of sickle gene by coelocentesis in early pregnancy: A new approach to prenatal diagnosis of single gene disorders. Hum. Reprod., 10, 1287–1289. Johnson, M.R., Jauniaux, E., Jurkovic, D. et al. (1994) Relaxin levels in embryonic fluids during the first trimester. Hum. Reprod., 9, 1561–1562. 278 E.Jauniaux and B.Gulbis Johnell, H.E. and Nilsson, B.A. (1971) Oxygen tension, acid–base status and electrolytes in human amniotic fluid. Acta Obstet. Gynecol. Scand., 50, 183–192. Luisi, S., Battaglia, C., Florio, P. et al. (1998) Activin A and inhibin B in extraembryonic coelomic and amniotic fluids, and maternal serum in early pregnancy. Placenta, 19, 435–438. MacCarthy, T. and Saunders, P. (1978) The origin and circulation of the amniotic fluid. In Fairweather, D.V.I. and Eskes, T.K.A.B. (eds), Amniotic Fluid: Research and Clinical Application. 2nd edn. Excerpta Medica, Amsterdam, The Netherlands, pp. 1–18. McKay, D.J., Richardson, M.V. and Hertig, A.T. (1958) Studies of the function of early human trophoblast: III. A study of the protein structure of mole fluid, chorionic and amniotic fluids by paper electrophoresis. Am. J. Obstet. Gynecol., 75, 699–707. Miell, J.P., Jauniaux, E., Langford, K. et al. (1997) Insulin growth factor binding proteins concentration and post translational modification in embryological fluid. Mol. Hum. Reprod., 3, 343–349. Nagy, A.M., Jauniaux, E., Jurkovic, D. and Meuris, S. (1994) Placental overproduction of human chorionic gonadotropin α-subunit in early pregnancy as evidenced in exocoelomic fluid. J. Endocrinol., 142, 511– 516. O’Rahilly, R. and Muller, F. (1992) Human Embryology and Teratology. Wiley-Liss, New York, USA. Parman, T., Wiley, M.J. and Wells, P.G. (1999) Free radical-mediated oxidative DNA damage in the mechanism of thalidomide teratogenicity. Nature Med., 5, 582–585. Ramsey, E.M. and Donner, N.W. (1980) Placental Vasculature and Circulation. Georg Thieme, Stuttgart, Germany, 101 pp. Riley, S.C., Leask, R., Chard, T. et al. (1999) Secretion of matrix metalloproteinase-2, matrix metalloproteinase-9 and tissue inhibitor of metalloproteinases into the intrauterine compartments during early pregnancy. Mol. Hum. Reprod., 5, 376–381. Rodesch, F., Simon, P., Donner, C. and Jauniaux, E. (1992) Oxygen measurements in the maternotrophoblastic border during early pregnancy. Obstet. Gynecol., 80, 283–285. Ross, J., Jurkovic, D. and Nicolaides, K.H. (1997) Coelocentesis: a study of short-term safety. Prenat. Diagn., 17, 913–917. Shannon, C., Jauniaux, E., Gulbis, B. et al. (1998) Placental transfer of fentanyl in early human pregnancy. Hum. Reprod., 13, 2317–2320. Shi, W.K., Hopkins, B., Thompson, S. et al. (1985) Synthesis of apolipoproteins, alphfoetoprotein, albumin and transferrin by the human foetal yolk sac and other foetal organs. J. Embryol. Exp. Morphol., 85, 191–206. Sibley, C.P. and Boyd, R.D.H. (1992) Mechanisms of transfer across the human placenta. In Polin, R.A. and Fox, W.W. (eds), Fetal and Neonatal Physiology. Vol. 1. Saunders, Philadelphia, USA, pp. 62–74. Sinha, R. and Carlton, M. (1970) The volume and composition of amniotic fluid in early pregnancy. J. Obstet. Gynaecol. Br. Commonw., 77, 211– 214. Umaoka, Y., Noda, Y., Narimoto, K. and Mori, T. (1992) Effects of oxygen toxicity on early development of mouse embryo. Mol. Reprod. Dev., 31, 28–33. Ward, S., Jauniaux, E., Shannon, C. et al. (1998) Electrical potential difference between exocoelomic fluid and maternal blood in early pregnancy. Am. J. Physiol., 274, R1492–R1495. Wathen, N.C., Cass, P.L., Kitau, M.J. and Chard, T. (1991) Human chorionic gonadotrophin and alpha-fetoprotein levels in matched samples of amniotic fluid, extraembryonic exocoelomic fluid, and maternal serum in the first trimester of pregnancy. Prenat. Diagn., 11, 145–151. Wathen, N.C., Cass, P.L., Campbell, J. et al. (1992) Levels of placental protein 14, human placental lactogen and unconjugated oestriol in extraembryonic coelomic fluid. [Letter.] Placenta, 13, 195–197. Wathen, N.C., Campbell, D.J., Patel, B. et al. (1993) Dynamics of prolactin in amniotic fluid and extraembryonic coelomic fluid in early human pregnancy. Early Hum. Dev., 35, 167–172. Watson, A.L., Palmer, M.E., Jauniaux, E. and Burton, G.J. (1997) Variation in expression of copper/zinc superoxide dismutase in villous trophoblast of the human placenta with gestational age. Placenta, 18, 295–199. Watson, A.L., Skepper, J.N., Jauniaux, E. and Burton, G.J. (1998a) Changes in the concentration, localisation and activity of catalase within the human placenta during early gestation. Placenta, 19, 27–34. Watson, A.L., Palmer, M.E., Skepper, J.N. et al. (1998b) Susceptibility of human placental syncytiotrophoblast mitochondria to oxygen-mediated damage in relation to gestational age. J. Clin. Endocrinol. Metab., 83, 1697–1705. Received on September 28, 1999; accepted on February 15, 2000