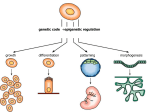
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Physiol Rev 87: 1343–1375, 2007; doi:10.1152/physrev.00054.2006. Hedgehog Signaling in Development and Homeostasis of the Gastrointestinal Tract GIJS R. VAN DEN BRINK Center for Experimental and Molecular Medicine, Academic Medical Center, Amsterdam; Department of Gastroenterology and Hepatology, Leiden University Medical Center, Leiden, The Netherlands Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 I. Introduction II. The Hedgehog Pathway A. Hedgehog secretion B. Hedgehog reception C. Hedgehog intracellular signaling D. Identifying Hedgehog target cells E. Hedgehog pathway antibodies, a cautionary note III. Hedgehog Signaling in the Developing Gut A. Patterning of the developing gut B. Early development of the gut tube and left-right axis formation C. Anterior-posterior axis patterning of the gut tube D. Endodermal cytodifferentiation and radial patterning of the gut tube IV. Hedgehog Signaling and Homeostasis of the Adult Gut A. Morphogenesis versus morphostasis B. Stomach C. Small intestine D. Colon V. Hedgehog Signaling and Gastrointestinal Carcinogenesis A. Evidence for a role for Hedgehog signaling in initiation of gastrointestinal carcinogenesis? B. Hedgehog signaling in cancers of the proximal gastrointestinal tract C. Hedgehog signaling in colorectal cancer D. Therapeutic targeting of the Hedgehog pathway VI. Conclusion 1343 1345 1345 1345 1346 1347 1348 1348 1348 1348 1350 1355 1362 1362 1363 1364 1364 1367 1367 1367 1368 1369 1369 van den Brink GR. Hedgehog Signaling in Development and Homeostasis of the Gastrointestinal Tract. Physiol Rev 87: 1343–1375, 2007; doi:10.1152/physrev.00054.2006.—The Hedgehog family of secreted morphogenetic proteins acts through a complex evolutionary conserved signaling pathway to regulate patterning events during development and in the adult organism. In this review I discuss the role of Hedgehog signaling in the development, postnatal maintenance, and carcinogenesis of the gastrointestinal tract. Three mammalian hedgehog genes, sonic hedgehog (Shh), indian hedgehog (Ihh), and desert hedgehog (Dhh) have been identified. Shh and Ihh are important endodermal signals in the endodermal-mesodermal cross-talk that patterns the developing gut tube along different axes. Mutations in Shh, Ihh, and downstream signaling molecules lead to a variety of gross malformations of the murine gastrointestinal tract including esophageal atresia, tracheoesophageal fistula, annular pancreas, midgut malrotation, and duodenal and anal atresia. These congenital malformations are also found in varying constellations in humans, suggesting a possible role for defective Hedgehog signaling in these patients. In the adult, Hedgehog signaling regulates homeostasis in several endoderm-derived epithelia, for example, the stomach, intestine, and pancreas. Finally, growth of carcinomas of the proximal gastrointestinal tract such as esophageal, gastric, biliary duct, and pancreatic cancers may depend on Hedgehog signaling offering a potential avenue for novel therapy for these aggressive cancers. I. INTRODUCTION One of the central problems of biology is the emergence of complexity. How did complex multicellular orwww.prv.org ganisms evolve, and how is genetic information translated to a spatially organized body plan? The hallmark of a developing multicellular organism is the fact that decisions regarding cell fate are not taken at the level of the 0031-9333/07 $18.00 Copyright © 2007 the American Physiological Society 1343 1344 GIJS R. VAN DEN BRINK FIG. 1. The Drosophila Hedgehog mutant. A: a schematic representation of the Drosophila larva. The larva is patterned into different segments. The posterior half of each segment or naked cuticle is smooth, whereas the anterior half is covered with bristles, the so-called denticles. B: wild-type fruitfly. C: the Hedgehog mutant. Note the absence of most of the naked cuticle in each segment. The resulting prominence of denticles gave the larve the aspect of a hedgehog. [B and C from Nusslein-Volhard and Wieschaus (143), with permission from Nature Publishing Group.] Physiol Rev • VOL FIG. 2. The French flag model of the mechanism of action of a morphogen. In this model, undifferentiated cells (left) can choose three different cell fates (blue, white, or red). This cell fate is specified in a cell position-dependent manner by localized production of a morphogen (middle). Cells respond in a distinct phenotypic manner to different thresholds of the concentration of the morphogen that depend on their distance from the source. It is important to realize that in reality many morphogens and their antagonists generate much more complex patterns. Also, morphogens such as those of the Hedgehog and Wnt pathway are not soluble molecules that spread to a tissue through simple diffusion but are lipid modified and very poorly soluble. Therefore, gradients probably form by active only partially resolved mechanisms. [Modeled after Wolpert (208).] ingly, of the 15 loci identified in the Nüsslein-Volhard and Wieschaus screen, 4 play a role in Hedgehog signaling. The screen identified the Drosophila hedgehog gene, Hedgehog receptor patched (Ptc), downstream transcription factor ci, and fused, a kinase involved in Hedgehog signaling (see below). The experiments by NüssleinVolhard and Wieschaus laid the foundation of our current understanding of the molecular mechanisms of tissue patterning. Many of the genes that were identified play a role in morphogenetic pathways. The existence of morphogens had been inferred at the beginning of the 20th century from experiments in developmental biology. The concept was most clearly voiced by Wolpert in the 1960s (208), who developed the so-called French flag model (Fig. 2). Morphogens are substances that originate from a localized source and form a concentration or activity gradient through a tissue. A morphogen receiving cell has one or more concentration thresholds that result in the expression of a distinct set of target genes. Thus morphogen concentration gradients confer positional information by specifying distinct cellular phenotypes in a field of receiving cells depending on their distance from the source of the signal (Fig. 2). In accordance with the limited number of loci identified by Nüsslein-Volhard and Wieschaus, a very limited number of highly conserved morphogenetic signaling pathways have now been identified. Hogan (66) coined the term morphogenetic code for these pathways that are used repeatedly in varying constellations to shape different organs. The four most important pathways that are conserved from fruitfly to human are the Hedgehog, Wnt, transforming growth factor (TGF)-, and receptor tyrosine kinase pathways. The ligands for receptor tyrosine 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 individual cell. To allow the building of complex organs with intricate patterns of cellular specialization, such decisions are taken at the population level in a cell nonautonomous manner. A breakthrough in the elucidation of the genetic control of such intercellular communication in developmental patterning events was made by Nüsslein-Volhard and Wieschaus (143). They performed a saturating mutagenesis screen in Drosophila (the fruitfly) and studied the effect of mutations on the segmented Drosophila embryo. Surprisingly, their mutagenesis screen in which they were estimated to have mutated about half the genes in the Drosophila genome only resulted in a small number of mutations (15 different loci) that affected Drosophila segmentation. In embryos of wild-type Drosophila, a band of bristles called denticles is present across the anterior half of each segment, whereas the posterior half is smooth (the so-called naked cuticle). In their screen, Nüsslein-Volhard and Wieschaus (143) identified a group of mutants that affected the patterning within the segments but at the same time left the number of segments unaltered. In one of these so-called segment polarity mutants, the posterior half of each segment failed to develop, resulting in a larva that is entirely covered by denticles (Fig. 1). This phenotype gave the larva the aspect of a hedgehog, which inspired the name. Interest- HEDGEHOG SIGNALING 1345 kinase receptors include, for example, the fibroblast growth factors (FGF), epidermal growth factor (EGF), and platelet-derived growth factor (PDGF). Below I discuss some of the most important aspects of Hedgehog secretion, reception, and signal transduction. As this review aims primarily to provide an overview of the role of Hedgehog signaling in the gastrointestinal tract, many important aspects of Hedgehog biology are not discussed. Several excellent reviews have been written for those interested in a more in-depth discussion of various aspects of the Hedgehog pathway (71, 77, 119, 147, 201). A. Hedgehog Secretion After the identification of the Hedgehog mutant in 1980, the Drosophila hedgehog gene was published in 1992 by three independent groups (112, 137, 183). Three murine hedgehog homologs were described a year later (40). They were named sonic hedgehog (Shh), after a popular Sega computer game character, desert hedgehog (Dhh), after an Egyptian species of hedgehog (Hemiechinus auritus), and indian hedgehog (Ihh), a hedgehog species endemic in Pakistan (Hemiechinus micropus). These three hedgehog genes are highly conserved between mouse and humans (130). Unfortunately, very little is known about the factors that control the transcriptional regulation of the three different hedgehog genes, particularly in the gut. Hedgehog proteins are extensively processed posttranslationally. Hedgehogs are produced as a ⬃45-kDa precursor protein that is cleaved autocatalytically, yielding a 19-kDa NH2-terminal fragment that contains all the signaling functions and a 26-kDa COOH-terminal fragment that catalyzes the cleavage and acts as a cholesterol transferase (Fig. 3) (16, 111, 156, 157). Although morphogens act in a graded manner through tissues, this is mostly not through simple diffusion of a soluble protein (127). In fact, an important feature of Hedgehog proteins is that the mature NH2-terminal fragments are dually lipid modified and therefore poorly soluble. The signaling protein is covalently linked to a palmitate and a cholesterol group (127). These hydrophobic modifications integrate Hedgehog protein in the cell membrane and play an important role in the regulation of the range of Hedgehog signaling in a tissue. Conflicting reports have been published on the role of the cholesterol moiety in Hedgehog signaling (113, 115). The palmitoylation is essential to both the activity of the Hedgehog protein and the signaling range (25). Work from several laboratories has shown that long-range signaling occurs by multimers of NH2-terminal Hedgehog protein that require both cholesterol and palmitate modPhysiol Rev • VOL FIG. 3. Posttranslational Hedgehog processing. The 45-kDa hedgehog precursor protein is processed to a 19-kDa NH2-terminal signaling protein by the COOH-terminal half, which catalyzes this reaction and acts as a cholesterol transferase. The cholesterol-modified NH2-terminal protein is further lipid modified at its NH2 terminus by palmitoylation, a reaction that is catalyzed by Skinny Hedgehog. ification for their formation (25, 212). Release of lipidmodified Hedgehog protein from the Hedgehog-producing cell requires Dispatched (Disp), a 12-pass transmembrane protein with a sterol-sensing domain. Disp1⫺/⫺ mutant mice have a phenotype similar to mice that lack the Hedgehog signaling receptor Smoothened (Smo), suggesting that Disp1 is required for all Hedgehog signaling activity (20, 94, 121). A study by Panáková et al. (149) in Drosophila suggested that Hedgehog multimers may form by association of the lipid moieties with the outer phospholipid layer of lipoprotein particles and that this association is required for Hedgehog signaling activity. This is a very interesting finding that may have important implications for vertebrate Hedgehog signaling (see, for example, a recent comment on the possible implications for atherosclerotic plaques, Ref. 11). Thus Hedgehog proteins are not soluble signals but dually lipid modified, and their spread through tissues is only partially understood. B. Hedgehog Reception The Hedgehog signal is transmitted by a seven-span transmembrane receptor Smoothened (Smo). Interestingly, Hedgehogs do not bind to Smo but control the activity of Smo indirectly by binding to a second receptor, Patched (Ptc). Two Ptc genes exist in vertebrates, Ptc1 (51) and Ptc2 (139). The Ptc genes encode for 12-span 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 II. THE HEDGEHOG PATHWAY 1346 GIJS R. VAN DEN BRINK In Drosophila, Smo is localized mainly in the cytoplasm in its inactive form and accumulates at the cell surface upon binding of Hedgehog to Ptc (33, 216). In vertebrate systems, inactive Smo is present in the cell membrane and intracellular vesicles (when overexpressed in cell lines) (23, 30, 76). The most notable change in Smo cellular distribution upon pathway activation in vertebrates is rapid accumulation on the primary cilium (Fig. 5). Corbit et al. (30) have shown that Smo is enriched in cilia in Hedgehog receiving cells in nonoverexpressing cells in vivo, that constitutively active Smo is enriched in the cilium in vitro, and that point mutations in the motif necessary for translocation to the cilium interfere with Hedgehog pathway activation in vitro. Interestingly, mutations in intraflagellar transport proteins, necessary for cilia formation, interfere with Hedgehog signaling (59, 72, 118, 132; for review, see Ref. 173). Together these findings suggested a functional role of the ciliary enrichment of activated Smo. In addition to Ptc, several other proteins have been identified that also bind Hedgehog and modulate signaling. Hedgehog-interacting protein (Hhip) is a transcriptional target that acts as a negative regulator of Hedgehog signaling by binding Hedgehog on receiving cells (26). Recently, a novel class of Hedgehog binding proteins has been identified in Drosophila that have two vertebrate homologs, Cdo and Boc. These Hedgehog binding proteins facilitate binding and positively regulate signaling (189, 210, 214). Boc acts as a Hedgehog receptor on commissural axons. Given the established role for Cdo and Boc in myogenic differentiation (84, 85), it is likely that they act as receptors in the regulation of myogenesis by Hedgehog signaling (see Ref. 41 for review). C. Hedgehog Intracellular Signaling FIG. 4. The current model of the mode of action of the Hedgehog receptor system. A: in the absence of ligand, Ptc pumps a Smo inhibitory molecule from one cellular compartment to another where it inhibits Smo. Upon binding of Hedgehog to Ptc, this pump function is abrogated and the inhibitory action on Smo released. Physiol Rev • VOL The analysis of Hedgehog signaling downstream from Smo (Fig. 5) is considerably complicated by the relative lack of conservation between Drosophila and vertebrates (71, 147). Although the intracellular transduction of the Hedgehog signal is relatively well understood in Drosophila, understanding of vertebrate signal transduction is far from complete; extensive discussion of the mechanism of Hedgehog signal transduction is therefore beyond the scope of this review. In Drosophila, the intracellular transduction of the Hedgehog signal depends on the processing of downstream transcription factor Ci. Ci exists in a full-length activating form and truncated repressor form. In the absence of Hedgehog, Ci is processed to its repressor form by a complex of four kinases [protein kinase A (PKA), casein kinase 1 (CK1), glycogen synthase kinase 3 (GSK3), and Fused kinase (Fu)] held together by Cos2. In the presence of Hedgehog, Cos2 is recruited by Smo and Ci processing terminated resulting 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 transmembrane receptors with two large hydrophilic extracellular loops that mediate Hedgehog binding. In the absence of Hedgehog, Ptc inhibits signaling by Smo (Fig. 4). Ptc1 is a transcriptional target of Hedgehog signaling and acts in a negative-feedback loop to restrict the range of Hedgehog signaling in a tissue. The mechanism of the inhibitory action of Ptc on Smo is only partially understood. Ptc acts catalytically (184) and seems to control the localization of a secondary Smo-inhibiting molecule. The nature of this inhibitory molecule is a matter of active investigation (12, 39). Even though Ptc may act as a pump that controls the localization of a secondary Smo antagonist, it has been shown that Ptc most likely acts in a cell autonomous fashion (on the cell expressing Ptc and not on surrounding cells) at least in Drosophila and the chick neural tube (14). This suggests that Ptc may pump this molecule from the outside to the inside of the cell or from one cellular compartment to another. The inhibitory action of Ptc is relieved upon binding of Hedgehog to Ptc. HEDGEHOG SIGNALING 1347 in the stabilization of full-length Ci and transcriptional activation of Hedgehog target genes (for review, see for example Refs. 67, 71, 83, 147). This function of Cos2 as an “on/off” switch does not seem to be conserved in vertebrates. Convincing evidence for a vertebrate Cos2 homolog is currently not available (200). Important evidence has been presented that shows that Suppressor of fused Su(Fu) may function as a central intracellular on/off switch in vertebrates (200). Indeed, the Hedgehog pathway is completely on in Su(Fu)⫺/⫺ mice (28, 182), which strongly resembled the Ptc1 null phenotype (182). Although Drosophila does have a Su(Fu) gene, it does not seem to play a similar role as Su(Fu) mutant Drosophila are healthy and fertile (159). The Hedgehog transcription factors are conserved end points in intracellular Hedgehog signal transduction. All aspects of Hedgehog signaling are mediated via the vertebrate homologs of Drosophila Ci, the Glioblastoma (Gli) transcription factors Gli1, Gli2, and Gli3 (74). As in Drosophila, the activity of these transcription factors is regulated in a ligand-dependent manner. Gli2 and Gli3 are the major Glis to transduce the Hedgehog signal in the gut (see below). Both Gli2 and Gli3 can also act as a repressor of Hedgehog target expression in vivo (116, 148, 203). This is related to the fact that Gli2 and Gli3 (and not Gli1) can be processed to a repressor form similar to the Drosophila homolog Ci. However, processing of Gli3 to its repressor form is much more efficient than that of Gli2 (148). As with Ci, the Gli3 repressor form is formed by a partial proteolytic processing reaction that requires priming phosphorylation by PKA and subsequent phosphorylation by CK1 and GSK3 (188, 203). Tempe et al. (188) have shown that this phosphorylation can result in ubiquitination and partial proteosomal degradation that generates a truncated version of Gli3 that lacks the transcripPhysiol Rev • VOL tional activation domain and acts as a dominant negative Gli (188). This negative regulatory action of PKA, CK1, and GSK3 (Drosophila homolog Shaggy) on Hedgehog signaling through the generation of a suppressor form of Ci/Gli protein is conserved from Drosophila to vertebrates. Gli2 processing and degradation and Gli1 degradation are regulated in a similar manner as that of Gli3 (9, 129, 148, 188). D. Identifying Hedgehog Target Cells In the analysis of the role of Hedgehog signaling in any organ, it is essential to identify Hedgehog receiving cells. Although a number of different target genes have been identified, their regulation often differs in time or per organ. Ptc1 and Gli1 are two target genes that seem to have been conserved particularly well throughout vertebrate evolution, and their expression pattern is the best reflection of Hedgehog signaling activity in most if not all situations in vertebrates. A difference may exist for the sensitivity of the expression of Ptc1 and Gli1 for the range of the Hedgehog signal in the developing vertebrate gut. In the developing stomach and colon, both Shh and Ihh are expressed in the epithelium at E18.5. Ptc1 is expressed at high levels in a small zone very close to the Hedgehog expressing cells, whereas expression of Gli1 is also very intense in the smooth muscle layer at much greater distance (164). It is, therefore, probably safest to use both Ptc1 and Gli1 as readouts for Hedgehog pathway activity and realize that these targets may have some level of independent regulation. As Ptc1 is the receptor for Hedgehog and required to initiate Hedgehog signaling, it is to be expected that low levels of Ptc1 may be expressed in tissues in a Hedgehog-independent manner. 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 FIG. 5. A model of Hedgehog signaling in the vertebrate. A: in the absence of Hedgehog signaling, Smo localizes mainly to the cell membrane and intracellular vesicles. GLI2 and GLI3 are processed by the proteasome upon phosphorylation by protein kinase A (PKA), glycogen synthase kinase 3 (GSK3), and casein kinase 1 (CK1). For GLI3, degradation is partial, resulting in a GLI3 protein that lacks the transcriptional activation domain and represses expression of hedgehog target genes. Degradation of GLI2 is complete. The vertebrate COS2 homolog (possibly KIF7) and SUFU further repress unprocessed cytoplasmic GLI via an unresolved mechanism. B: in the presence of Hedgehog, repression of SMO by PTC is relieved, and SMO translocates to the primary cilium where it activates GLI proteins. 1348 GIJS R. VAN DEN BRINK E. Hedgehog Pathway Antibodies, a Cautionary Note immunohistochemistry with those obtained by in situ hybridization until better antibodies become available. The analysis of the role of Hedgehog signaling in development and the adult critically depends on the reliable detection of components of the Hedgehog pathway such as Shh, Ihh, Dhh, Smo, Ptc, and transcription factors Gli1–3. A large variety of commercial antibodies is currently available (mainly from Santa Cruz). With many if not most of these antibodies, we have not been able to obtain credible results with western blot or immunohistochemistry, especially when compared with in situ hybridization. In our hands the specificity of the available commercial antibodies may even vary between batches. Some of the antibodies that gave staining patterns that compared well with in situ hybridization in our hands still give variable results in the literature. For example, using the I-19 antibody against Ihh from Santa Cruz, Varnat et al. (202) observed staining at the base of the crypts, whereas Jones et al. (80) describe Ihh expression mainly at the tips of the villi using the same antibody. Since Ihh mRNA is mainly expressed at the crypt villus junction with a diminishing gradient towards the top of the villus, the staining pattern observed by Jones et al. (80) probably best reflects the expression of Ihh protein. It is furthermore important to realize that many anti-Hedgehog antibodies are produced against the extremely conserved NH2-terminal 19-kDa protein. This results in easy cross-reactivity of antibodies for different Hedgehog proteins. This may explain, for example, why Jones et al. (80) detect abundant staining on villi of the small intestine with an anti-Shh antibody, whereas Shh expression is very low and restricted to a few cells in the small intestinal crypts as detected by in situ hybridization (6, 198), making it very unlikely that abundant Shh protein can be detected in the enterocytes on the villi. Most likely this is explained by cross-reactivity of the anti-Shh antibody for Ihh protein. Additionally, using the Santa Cruz C-20 antibody against Ptc1, we have previously reported epithelial staining in the colon of mouse, rat, and human (197). We have now performed in situ hybridizations for Ptc1 in the mouse colon and found that although Ptc1 mRNA is expressed in the epithelium of the anorectum, no Ptc1 mRNA can be detected in the epithelium of the mouse colon using two different Ptc1 probes. Instead, Ptc1 localizes to the mesenchyme just underlying the differentiated colonic enterocytes (unpublished observations). Hopefully more reliable anti-Hedgehog pathway antibodies will be available soon. Of course, it should be realized that Hedgehogs are proteins that can travel great distances through tissues and that a perfect correlation between Hedgehog mRNA and protein may not always be present. However, given the poor performance of many Hedgehog pathway antibodies, it is important to compare results obtained with III. HEDGEHOG SIGNALING IN THE DEVELOPING GUT The gastrointestinal tract forms from two germ layers, the endoderm and mesoderm, and is innervated by cells derived from the third germ layer, the ectoderm. During development the gut evolves from a simple tube that is morphologically homogeneous to a highly complex organ that has distinct functional domains along the anterior-posterior and vertical (cross-sectional) axis and develops multiple accessory organs. Three different regions are normally recognized. The foregut forms the esophagus, stomach, proximal duodenum, thymus, thyroid, airways, pancreas, and liver. The midgut will form most of the intestine distal from the common bile duct that derives its blood supply from the superior mesenteric artery: the distal duodenum, the jejunum, ileum, cecum, and the ascending and proximal transverse colon. The hindgut forms the distal transverse colon, descending colon, sigmoid, and anorectum that develop around the inferior mesentery artery. The patterning mechanisms along the vertical, horizontal, and left-right axes and induction of growth of accessory organs all involve cross-talk between cells of the different germ layers. Such interactions between different groups of cells in which a signal is emitted from one group of cells to change the nature or behavior of the receiving cells are termed inductive signaling. Two types of inductive signaling are generally recognized. In instructive interaction, the signal is necessary to specify the fate of the receiving cell. In permissive interaction, the signal simply allows the expression of a phenotype that is already intrinsic to the receiving cell. Much of this instructive communication between germ layers is through the action of morphogens among which are members of the Hedgehog signaling pathway. Below I first review some of our understanding of the development of the gut and the role of germ layer interactions, focusing on mouse development. I then discuss the available data that show a role for Hedgehog signaling in gut development. B. Early Development of the Gut Tube and Left-Right Axis Formation In all animals the primitive gut or archenteron is formed during gastrulation. The term gastrulation is derived from the Greek word “gaster” and means “to form a stomach.” At this early point in development, the mouse embryo is a cup-shaped epithelial layer termed epiblast 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 Physiol Rev • VOL A. Patterning of the Developing Gut HEDGEHOG SIGNALING that is enveloped by the visceral endoderm (Fig. 6). The epiblast will form three germ layers during gastrulation: definitive (gut) endoderm, mesoderm, and ectoderm. The definitive endoderm gives rise to the epithelium of the gastrointestinal tract, thymus, thyroid, and respiratory tract; the mesoderm will form the cardiovascular system, muscles, blood, and bone; the ectoderm forms the epithelium of the skin and the central nervous system. Gastrulation starts at embryonic day (E) 6.25 with the formation of the primitive streak, a thickening on the posterior margin of the epiblast (and future posterior end of the embryo) that forms by epithelial reorganization and is elongated anteriorly over the epiblast along the midline of the embryo (176). Epiblast cells undergo a process of epithelial to mesenchymal transition (EMT) at the primitive streak and ingress through the primitive streak and node (Fig. 6). The embryonic endodermal cells that have undergone EMT are inserted in the visceral endoderm, gradually displacing it. Other cells that have undergone EMT migrate in between the endoderm and epiblast (now Physiol Rev • VOL the primitive ectoderm) forming the mesoderm (176). At the anterior end of the primitive streak, a small group of cells forms a notch that is termed the node. Cells of the node regulate left-right axis formation of the embryo and are therefore essential for left-right axis formation of the gut and its derived organs such as looping of the gut tube and correct positioning of the pancreas and liver (65). The node has a central depression termed the pit, which is populated by ciliated cells that generate a leftward flow of extraembryonic fluid. This leftward flow is the earliest recognized left-right symmetry breaking event during murine embryogenesis (65) and is essential for the establishment of asymmetric mesodermal expression of genes involved in a program that specifies the left side of the mouse embryo. Nodal flow may act by causing asymmetric distribution of a soluble signal to the left of the developing embryo (142, 144). A “left-side program” is subsequently activated in the left lateral plate mesoderm, which contributes to the mesoderm of the gut and has been shown to play an essential role in gut looping in zebrafish (68). By E7.5, the mouse embryo has developed “insideout” with an internal layer of ectoderm that is surrounded by mesoderm and endoderm (206). Formation of the gut tube is initiated by the formation of the anterior intestinal portal (AIP; Fig. 7). The AIP is an endodermal invagination at the anterior tip of the embryo at the level of the cranial neural fold of the ectoderm (91). Formation of the AIP is closely followed by the formation of a second endodermal invagination at the caudal end of the embryo, the caudal intestinal portal (CIP) (91). At E8.5, a complex turning process of the embryo is initiated that folds the endoderm to the inside of the embryo and elongates the AIP and CIP while bringing their openings together around the yolk sac stalk upon completion of the turning process at E9.5 (Fig. 7) (91, 206). 1. Expression of hedgehogs during gastrulation Shh expression is first detected at E7.25 in the midline mesoderm of the head process (40). At E7.75, Shh expression is initiated in the node, the notochord, and the definitive endoderm (40, 215). Endodermal expression of Shh is initiated in the anterior endoderm, and its expression expands to the posterior endoderm (40). Ihh is expressed in the posterior node and the visceral endoderm at E7.75 (46, 215). Ihh expression in the definitive endoderm is initiated at later stages of development (13). 2. Hedgehog signaling and left-right axis formation Hedgehog signaling is dispensable for early gastrulation in the mouse as all three germ layers are normally formed in mice that lack Smo and therefore lack all Hedgehog signaling (215). At a later stage of gastrulation, Hedgehog signaling is involved in gut tube formation as 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 FIG. 6. Germ layer formation. A: at E5.5, the mouse embryo consists of a single cell layer, the epiblast, enveloped in visceral endoderm. B: during gastrulation, the posterior end of the epiblast forms a thickened region, the primitive streak. Epiblast cells undergo epithelial to mesenchymal transition as they pass through the primitive streak and form the mesenchyme and endoderm. The remaining epiblast cells form the ectoderm. The endodermal cells migrate anteriorly (arrow) while displacing the visceral endoderm. The embryo is now still inside-out and will get its correct orientation during a complex turning movement. 1349 1350 GIJS R. VAN DEN BRINK FIG. 7. Turning of the embryo. A: at E8, the ventral side of the cupped embryo is oriented towards the outside. B: after a complex turning movement, the ventral side ends up at the inside of the cupped embryo bringing the anterior intestinal portal (AIP) and caudal intestinal portal (CIP) in close apposition. Physiol Rev • VOL (0.3–5 m) membrane particles (that they termed “nodal vesicular parcels” or NVPs) in an FGF signaling-dependent manner. The NVPs were transported to the left by nodal flow and seemed to contain Hedgehog protein. Recombinant Shh could reverse inhibition of asymmetric Ca2⫹ signaling by an FGF inhibitor (185). The symmetrical expression pattern of Hedgehog receptor and transcriptional target Ptc1 in the node seems to contradict the possibility that Hedgehog containing NVPs induces asymmetric Hedgehog signaling in the node. However, the authors of the node work suggest that Ptc1 may be a short-range Hedgehog signaling target (at least in the fruitfly imaginal disks) and that short-range and longrange Hedgehog signaling may have alternative mechanisms of action (65). The authors suggest that the NVPs may be important in the long-range signaling of Hedgehogs in the node and may be important for the asymmetric induction of Ca2⫹ signaling. Although this remains a possibility, this suggestion is highly speculative and awaits further experimental evidence. In conclusion, Hedgehog signaling plays an important role in left-right axis specification of the gut and other organs in the body. To date, the most important evidence is for a role for Hedgehog signaling in midline specification, which functions as a barrier to restrict diffusion of asymmetric signals to one side of the developing embryo. An alternative more direct role for Hedgehog signaling remains a possibility but awaits further evidence. C. Anterior-posterior Axis Patterning of the Gut Tube 1. Instructive signals from the node and mesoderm establish the anterior-posterior axis early in development After gastrulation, the endodermal layer is still a histologically uniform pseudostratified layer of cuboidal epithelial cells that is surrounded by a thin layer of me- 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 both Smo and Shh/Ihh compound mutant mice (215) fail to close the midgut. This defect may be related to that fact that they do not undergo the embryonic turning process, something that will impair the normal endodermal folding movement that results in gut tube closure. Several different mice with mutations in the Hedgehog pathway display defective left-right axis formation. Smo mutant and Shh/ Ihh compound mutant mice do not initiate cardiac looping (lung or gut phenotype cannot be examined in Smo mutant or Shh/Ihh compound mutant mice as they do not survive beyond E9.5) (215). Shh mutant mice have a less severe cardiac phenotype with delayed and incomplete looping and fail to develop asymmetrically lobed lungs (193, 215). Both Ihh and Shh mutant mice display gut malrotation (164). These data indicate that Shh and Ihh play redundant roles in left-right axis formation, consistent with the expression of both Shh and Ihh in the node. Smo and Shh/Ihh mutant mice fail to activate the genetic program necessary for the specification of the left side of the body. Both mutants lack expression of nodal in the left lateral plate mesoderm, a critical component of the left-side specific gene program (215). As the Hedgehog target gene Ptc1 is symmetrically expressed in the node, it is most likely that the effect of Hedgehog signaling on left-right axis formation is indirect. For example, Hedgehog signaling is important in the specification of the midline, which has been proposed to play an important role in the stabilization of left-right specific gene expression by functioning as a barrier that prevents the bilateral diffusion of long-range asymmetric signals (for review, see Capdevila et al., Ref. 18). A possible more direct role for Hedgehog signaling in left-right axis formation has been proposed by Tanaka et al. (185). It has been shown that nodal flow initiates asymmetric Ca2⫹ signaling at the left border of the node (133). Tanaka and colleagues found that this Ca2⫹ signal depends on FGF signaling. When the authors labeled membrane lipids with a fluorescent dye, they made the interesting observation that nodal cells released small HEDGEHOG SIGNALING Physiol Rev • VOL been examined if Hedgehog signaling is involved in these early AP patterning events. 2. Instructive signals from the endoderm pattern the gut along the AP axis during later stages of development The endodermal layer holds the information necessary for its differentiation along the AP axis at later stages in development in both mice (177) and rats (38). This indicates that the mesenchyme mainly has a permissive role in endodermal development from this point onward. For example, when undifferentiated E14 rat intestinal endoderm is recombined with mesenchyme from different regions of the fetal small intestine, the endoderm seems to determine the region-specific expression of brush-border enzymes, and when E14 rat gastric or lung endoderm was recombined with small intestinal mesenchyme, the endoderm developed according to its place of origin (38). The only exception to the endodermal autonomic region-specific development was in the prospective colonic endoderm. The colonic endoderm expressed small intestinal enzymes when combined with small intestinal mesenchyme, indicating that cell fate of the colonic endoderm remains somewhat plastic at E14 (38). It was even shown that normal intestinal endodermal development is possible when E14 rat endoderm is recombined with fetal skin fibroblasts, with the fibroblasts developing into a smooth muscle layer (instead of skeletal muscle) that was ␣-smooth muscle positive (38, 95). This indicates that the mesenchyme not only plays mainly a permissive role at later stages of endoderm development but that signals from the endoderm play an instructive role in the development of the mesoderm at this stage. Because Hedgehogs are expressed in the endoderm and signal to the mesenchyme, this may implicate the pathway in the instructive signaling to the mesenchyme at this stage. 3. Endodermal appendage formation One of the most notable aspects of AP axis formation of the gut is the formation of endodermal appendages (Fig. 8). During and right after the turning of the embryo between E8.5 and E9.5, a series of appendages originate from the gut tube by a process of budding and elongation. Buds are formed that generate the thymus (E9.5–E10.5) (126), thyroid (E8.5) (31), airways (E9.0 –E9.5) (117), liver (E8.5) (82), and pancreas (E9.5) (100). The specification of the endodermal zone destined for appendage formation and the subsequent morphogenetic process of budding and elongation seems dependent on instructive mesodermal signals. For example, at E8.25–E8.5 signals derived from the cardiac mesoderm play an essential role in the development of the liver and airways from a region of the ventral foregut endoderm that lies in close apposition to the cardiac mesoderm (56, 81, 174). The notochord, a 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 sodermal cells. Despite its morphological uniformity, the endodermal layer is already patterned along the anteriorposterior (AP) axis at this point in development (53, 206). Lawson and Pedersen (108) microinjected single endodermal cells of E6.7 early primitive streak stage mouse embryos with horseradish peroxidase and traced them for up to 48 h in culture. Their experiment showed that the first endodermal cells to exit the primitive streak formed the anterior-most endoderm, whereas cells that were still in transit through the primitive streak formed more posterior endoderm (108). The earliest AP patterning of endodermal cells may be related to this timing of the exit of the primitive streak. For example, the endodermal cells that were first to exit the primitive streak and have moved most anteriorly express the homeobox gene Hhex (or Hex) (192), whereas later endodermal cells express FoxA2 (or Hnf3), and the last endodermal cells to exit the primitive streak and thus form the posterior endoderm express Cdx2 (7). Hereafter further AP patterning occurs through instructive and permissive interactions between the endoderm and mesoderm. The major instructive interactions in early intestinal development seem to be from the mesoderm to the endoderm. Unfortunately, few studies exist that examined the role of endodermal mesodermal interactions in the mouse during early gut development. In a recent study that examined the role of germ layer interactions in patterning of the early (E7.5–E7.75) mouse endoderm, the endoderm survived for at least 2 days when it was cultured in vitro in the absence of mesoderm and maintained the regionspecific expression of two early posterior endodermal markers the anterior marker -cardiac actin (bCa) and the posterior marker intestinal fatty acid binding protein (iFabp) gene (205). In this elegant study, the expression of several regulators and markers of endodermal differentiation was only induced when the endoderm was cultured with the other two germ layers. The effect was mediated by soluble factors as it also occurred in the presence of a membrane that separated the endoderm from the mesoderm/ectoderm. These results might still simply indicate that endoderm arrests in development when cultured without trophic factors derived from the other germ layers and that the effect would therefore be an example of permissive induction. However, evidence for an instructive role for the mesoderm/ectoderm was provided in further experiments. When anterior endoderm was cultured with posterior mesoderm/ectoderm, the endoderm expressed the posterior marker iFabp. When posterior endoderm was cultured with anterior mesoderm/ectoderm it expressed anterior marker bCa. These data indicate that AP patterning of the early mouse endoderm occurs through both timing of the exit of the primitive streak and instructive action of soluble factors derived from the other two germ layers. It has not 1351 1352 GIJS R. VAN DEN BRINK mesodermal structure that is formed during gastrulation and ensures structural support along the axis of the growing embryo, is in close contact with the dorsal endoderm until E9.0 and specifies the dorsal pancreatic domain in the chick embryo (61, 102). Inductive signals from endothelial cells are involved in the specification of the thyroid and both pancreatic buds (see below). 4. Hedgehog signaling and endodermal appendage formation A) AIRWAYS. After the formation of a central ventral tracheal bud and two adjacent ventrolateral lung buds at E9.5 (19), the trachea and lungs separate from the esophagus by a process of septation and elongation. Shh is expressed at high levels in the tracheal bud (117). Signaling is from the endoderm to the mesoderm as the Gli transcription factors are exclusively expressed in the foregut mesoderm (74). One of the roles of Hedgehog signaling to the mesoderm is in the correct specification of the mesodermal septum that separates the esophagus and trachea. In Shh⫺/⫺ mice, tracheal budding is delayed and the elongating trachea is hypoplastic and fails to separate correctly from the gut tube. The proximal esophagus and trachea are therefore poorly separated (117, 153). The lungs of Shh⫺/⫺ mice are severely hypoplastic and do not lobulate, and the airways fail to branch and recruit proliferating mesenchymal cells, suggesting a role for Hedgehog signaling in branching morphogenesis (117, 153). Gli transcription factors play a redundant role in airway budding, elongation, and branching. Gli1 mutant mice are healthy and do not seem to have a phenotype (150). Gli2⫺/⫺ mice recapitulate many of the aspects of the Physiol Rev • VOL 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 FIG. 8. A budding phase between E8.5 and E9.5 generates the endodermal appendages. Schematic depiction of the E9.5 embryo. Red font indicates exclusion of Shh expression, and green font indicates high Shh expression. Thb, thyroid bud; tb, tracheal bud; lb, lung bud; hb, hepatic bud; pb, pancreatic bud. phenotype of the Shh⫺/⫺ mice with hypoplastic trachea and lungs and failure to induce mesenchymal proliferation (138). The Gli3⫺/⫺ mouse shows normal airway development but loss of one copy of the Gli3 gene in Gli2⫺/⫺ Gli3⫹/⫺ mice aggravates the airway phenotype and is indiscernible from the Shh⫺/⫺ mouse. The phenotype of mice completely deficient for both Gli2 and Gli3 is more severe than that of Shh⫺/⫺ mice as in these mice there is no development of the trachea and lungs at all (138). This may suggest a role for another Hedgehog in airway budding and outgrowth. Expression of Ihh in the foregut has not been reported, but low expression levels may have been missed. A major target and effector of Hedgehog signaling in airway development is transcription factor Foxf1, which is expressed in the mesenchyme (124). Expression of Foxf1can be induced by ectopic expression of Shh, and Shh⫺/⫺ mice lack Foxf1 expression in the foregut mesenchyme (124). Foxf1⫺/⫺ mice degenerate before endodermal budding begins (125), but Foxf1⫹/⫺ mice do have an airway phenotype with a failure of the elongating trachea to separate completely from the esophagus and reduced branching morphogenesis similar to the Hedgehog pathway mutant mice (124). Thus Hedgehog signaling from the endoderm to the mesoderm is essential for the formation of the tracheal bud and subsequent branching morphogenesis of the airways and plays a critical role in the stimulation of growth of the recruited mesenchyme. B) THYROID. The adult thyroid gland is located in the cervical region anterior to the trachea in humans and mice. The bulk of the gland is made up of follicles consisting of thyroid follicular cells that store and release thyroid hormone. C cells produce calcitonin and are found in the interfollicular space. Both cell types have a different embryonic origin as the follicular cells are endoderm derived whereas C cells are derived from the neural crest (31). Thyroid morphogenesis is initiated at E8.5 when the thyroid primordium buds of the ventral wall of the pharyngeal foregut endoderm that lies in close contact with the aortic sac endothelium (2, 43). At E9.5, the thyroid bud separates from the gut tube while it remains connected by a thin thyroglossal duct and remains in contact with the aortic arch. The thyroglossal duct disappears and the thyroid becomes completely separated from the gut by E11.5. At E13.5, the thyroid is at its final position in the midline at the ventral side of the trachea. It now bifurcates to form two lobes that develop in close contact with the carotid arteries (2). At E15.5, the thyroid organizes into a follicular structure, and genes involved in thyroid hormone production are induced (31). Similar to the specification of the pancreatic buds (see below) and in contrast to specification of the airways, expression of Shh is excluded from the thyroid primordium (44). In the absence of Hedgehog signaling, the thyroid bud is therefore correctly specified. Although no HEDGEHOG SIGNALING Physiol Rev • VOL FIG. 9. A pancreatic and hepatic endodermal domain are specified in the ventral endoderm. A population of endodermal cells at the lip of the ventral endoderm are specified as Pdx1-positive, albumin-negative ventral pancreatic precursor cells, whereas cells of the more cranial endoderm that lies in close apposition to the cardiac mesenchyme are specified as Pdx1-negative albumin-positive hepatic precursor cells. 102). The ventral pancreatic domain is specified at the lip of the mouse ventral foregut endoderm (Fig. 9). This endodermal region adopts a pancreatic cell fate around the six- to eight-somite stage (E8) just before the turning of the embryo (34). The endoderm that lies slightly more cranial is closely apposed to the cardiac mesoderm which secretes FGFs to suppress a pancreatic and induce a hepatic cell fate in this endodermal region (34, 81). The ventral and dorsal pancreatic buds develop between E8.5 and E9.5 exactly where the endoderm interacts with the aorta dorsally and the two vitelline veins ventrally (106, 107). It was shown by a variety of different experimental approaches that this endodermal contact with the endothelium is both necessary and sufficient to determine the expression domains of Pdx1 and insulin (106). Similar to thyroid development (discussed above), pancreatic development needs exclusion of Hedgehog expression from both the ventral and dorsal pancreatic primordia (4, 34, 61, 102). At E9.5–E10.5, Shh and Ihh are expressed uniformly throughout the endoderm of the gut tube with the exception of Pdx1 positive pancreatic precursor cells (4, 114). Forced expression of Shh in the pancreatic primordia using a Pdx1 promoter resulted in abnormal development of the pancreatic endoderm and mesoderm. The pancreatic mesoderm of these transgenic animals contained ␣-actin positive smooth muscle cells and c-kit positive cells of Cajal. Both cell types are normally exclusively found in intestinal mesenchyme. The pancreatic buds in Pdx1-Shh mice did contain endocrine and acinar cells, but these failed to cluster into islets and acinar structures and produced mucins that are typical for intestinal epithelial cells (4). Thus suppression of Shh 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 Shh is expressed in the thyroid at any point in development, the thyroid does not separate into two distinct lobes in Shh⫺/⫺ mice but becomes a single unilateral mass, the same size of a single thyroid lobe in control mice (44). The single thyroid gland of Shh null mice formed normal follicles that contained apparently normally differentiating thyrocytes. In an elegant study by Alt et al. (2), it was shown that the thyroid phenotype is indirect, in accordance with the lack of Shh expression in the thyroid. The authors showed that thyroid growth is directed towards endothelial cells in the zebrafish. Subsequently, the authors examined a mouse that lacks Shh expression at early stages of development and demonstrated that the aortic arch fails to cross the midline and both carotid arteries develop on one side of the esophagus. They found that the single thyroid lobe in these mice is always on the same side as and in close contact with the mislocated carotid arteries. The fact that Shh expression is specifically excluded from the thyroid primordium suggests that downregulation of Shh expression may be an important step in its specification similar to the specification of the pancreatic domain (see below). This idea is supported by the finding of small foci of ectopic thyroid tissue in the tracheal tube at 15.5–17.5 days post coitum in the Shh⫺/⫺ mouse (44), suggesting that Shh acts to suppress thyroid cell fate in the trachea and its loss may be critical to thyroid primordium formation. Another similarity between the development of the thyroid and pancreas is the important role of the endothelium in budding (see below), suggesting that a signal derived from endothelial cells may be involved in the restriction of Shh expression from the thyroid and pancreatic buds. In conclusion, Shh suppresses thyroid cell fate specification in the trachea, and its expression is excluded from the thyroid primordium, possibly by signals from the endothelium of the aortic sac. Although the Shh⫺/⫺ mouse has a thyroid phenotype, this phenotype is indirect and the result of defective patterning of the cervical vasculature. C) PANCREAS. The pancreas is a gland with both endocrine and exocrine functions. Most pancreatic tissue (⬃95–99%) is composed of exocrine acinar cells that produce digestive enzymes and pancreatic ducts that are lined by duct cells. The rest of the pancreas consists of small clusters of endocrine cells or islets of Langerhans. Each cluster of endocrine cells has a core of insulinproducing -cells surrounded by glucagon-producing ␣-cells, somatostatin-producing ␦-cells, and pancreaticpolypeptide cells. The pancreas develops from two distinct domains of ventral and dorsal foregut endoderm. The dorsal endodermal domain involved in the formation of the pancreas is specified by signals from the notochord, which remains in close contact to the dorsal endoderm until E8.5 and induces the expression of transcription factor Pdx1 (61, 1353 1354 GIJS R. VAN DEN BRINK TABLE 1. Endodermal appendage phenotypes in Hedgehog pathway mutant mice Airways Shh⫺/⫺ Ihh⫺/⫺ Gli1⫺/⫺ Gli2⫺/⫺ Gli3⫺/⫺ Gli2⫺/⫺ Gli3⫹/⫺ Gli2⫺/⫺ Gli3⫺/⫺ Hhip (170), but injection of Shh increases the number of pancreatic precursor cells (170) whereas Shh and Smo mutant zebrafish show diminished numbers of pancreatic precursor cells (35, 170). Experiments by DiIorio et al. (35) suggest that this role of Hedgehog signaling in early specification is mediated during gastrulation. Thus Hedgehog signaling differentially regulates early pancreatic development in the mouse and zebrafish. Hedgehog signaling inhibits early pancreatic development in the mouse, and the exclusion of Shh and Ihh expression from the pancreatic primordia is critical to normal murine pancreas development. D) LIVER. Shh is expressed in the liver primordium in the ventral foregut endoderm (34). It is not clear if there is a role for Hedgehog signaling in liver budding and development as liver development seems to progress normally in both Hedgehog and Gli mutant mice (117, 138) and, for example, in Xenopus injected with a constitutively active form of Smo (213). In conclusion, instructive mesenchymal signals play an essential role in the establishment of the AP axis during early gut development. This is most evident from the role of mesenchymal factors in the appropriate formation of endodermal appendages. See Table 1 for a summary of phenotypes in Hedgehog pathway mutant mice. Much of this instructive signaling seems to act by modulating the expression of Hedgehogs in the endoderm, which subsequently acts on the mesenchyme in a reciprocal manner. ⫺/⫺ Thyroid Hypoplastic trachea, severely hypoplastic lungs No gross abnormalities Single-lobed thyroid gland, thyroid tissue in trachea Grossly normal Hypoplastic trachea, hypoplastic lungs, lobulation defect right lung Hypoplastic lungs Hypoplastic trachea, severely hypoplastic lungs Complete tracheal atresia, no lungs Branching defect Grossly normal ND ND Pancreas Grossly normal Liver Grossly normal Annular pancreas, increased numbers of endocrine cells Grossly normal ND ND Grossly normal ND ND ND ND ND ND Normal hepatic bud ND ND Normal hepatic bud ND Loss of endocrine cells, exocrine cells normal Intestinal type pancreatic mesenchyme, disorganization of pancreatic endoderm Reduced pancreatic mass, intestinal type pancreatic mesenchyme, loss of endocrine cells, loss of exocrine cells ND Pdx1-Shh Pax4-Shh and Ihh ND, not determined. Note: no gastrointestinal phenotypes are available of the Shh Ihh double-knockout and Ptc and Smo knockout mice as they are early embryonic lethal. Physiol Rev • VOL 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 signaling during early pancreatic development is essential for normal development of both the pancreatic epithelium and mesoderm. Data from Xenopus further support this concept (213). Xenopus injected with a constitutive active form of the Hedgehog receptor Smo completely lack a pancreas, a phenotype more severe than that of the Pdx1Shh mouse. This may be due to the fact that the transgenic mouse expressed Shh behind a pancreas specific promoter and that the ectopic Shh transcription therefore starts after the pancreatic domain has already been specified. Similar to the role of Shh in suppressing thyroid cell fate in the trachea, expression of Shh in the endoderm surrounding the pancreatic primordium may be necessary to suppress pancreatic cell fate in the duodenum and stomach. Treatment of chick embryos with the Hedgehog inhibitor cyclopamine in ovo resulted in ectopic pancreatic tissue formation in the duodenum and stomach (103). Cyclopamine acts at the level of Smo and therefore inhibits the activity of all Hedgehogs (29, 75). No pancreatic differentiation was observed in the stomach or intestine of Shh null mice (62), but this may be related to the fact that Shh and Ihh have overlapping functions in this domain (13, 164). The role of Hedgehog signaling in pancreatic development in zebrafish seems to diverge from that in the mouse in some aspects. Evidence has been presented that in contrast to the mouse, Hedgehog signaling is required for the early stages of pancreas development in the zebrafish (35, 170). As in the mouse, Shh expression is excluded from the pancreatic anlage in the zebrafish 1355 HEDGEHOG SIGNALING D. Endodermal Cytodifferentiation and Radial Patterning of the Gut Tube Endodermal cytodifferentiation is complete at birth in humans. It is important to realize that in contrast to humans, the epithelium is still immature at birth in mice and rats and that its development continues in the first four postnatal weeks. Here I discuss the changes that occur in the first three postnatal weeks as part of the developing gastrointestinal tract and consider the gastrointestinal tract as “adult” after completion of postnatal development. The gut tube phenotypes of Hedgehog pathway mutant mice are summarized in Table 2. Differentiation of the esophageal endodermal layer is poorly studied in the mouse. A morphological study by Raymond et al. (165) suggests that differentiation initiates around E15. At E17, the epithelium is composed of three to four cell layers and contains both squamous and ciliated cells. Only cells in the basal most epithelial layer proliferate, and thus a division in a basal compartment of precursor cells and superficial compartment of differen- TABLE 2. Gut tube phenotypes of Hedgehog pathway mutant mice Esophagus Shh⫺/⫺ Ihh⫺/⫺ Gli1⫺/⫺ Gli2⫺/⫺ Gli3⫺/⫺ Gli2⫺/⫺ Gli3⫹/⫺ Gli2⫺/⫺ Gli3⫺/⫺ Stomach Intestine Esophagus and trachea fail Anterior expansion of Gut malrotation, duodenal to separate, glandular stomach villus overgrowth, tracheoesophageal increased gland abnormal neuronal fistula fission differentiation No gross abnormalities No gross Gut malrotation, abnormalities hypoplastic villi, segmental absence of neurons, Hirschsprung’s-like colonic phenotype Grossly normal Grossly normal Grossly normal Hypoplastic trachea and No gross ND esophagus, hypoplastic abnormalities lungs, lobulation defect right lung ND Anterior expansion of ND glandular stomach, increased glandular growth Esophagus and trachea fail ND ND to separate, tracheoesophageal fistula Complete atresia of ND ND esophagus Hypoplastic branching villi, precursor cell accumulation, crypts on villi Villln-Hip Anorectum Reference Nos. Imperforate anus, fused 99, 117, 136, 153, 164 anorectum-urinary tract No gross abnormalities 164 Grossly normal 150 Imperforate anus, fecto- 99, 136, 138 vesical fistula Anal stenosis 99, 136, 138 Persistent cloaca, common outlet guturinary tract 136, 138 Persistent cloaca, common outlet guturinary tract 136, 138 122 ND, not determined. Note: no phenotypes are available of the Shh⫺/⫺ Ihh⫺/⫺ double-knockout and Ptc and Smo knockout mice as they are early embryonic lethal. Physiol Rev • VOL 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 1. Esophageal development and the role of Hedgehog signaling tiating cells has been established along the vertical axis at this point in time. The ciliated cells rapidly disappear postnatally and are absent 1 wk after birth. The mechanism of this loss of ciliated cells seems to be through desquamation (134). In contrast to the human esophagus, which is not keratinized, murine esophageal epithelium keratinizes ⬃1 mo after birth (37). Hedgehog signaling plays a critical role in the normal development of the esophagus (Fig. 10). Shh is initially expressed throughout the developing esophagus but restricted to the distal esophagus at later stages (117, 164). The Gli genes are mesodermally expressed, indicating that Hedgehog signaling is from the endoderm to the mesoderm (74). At E17.5, the proximal esophagus of Shh⫺/⫺ mice is hypoplastic and fails to separate completely from the developing trachea; more distally there is no discernible remaining esophagus in Shh⫺/⫺ mice at this point in development (117). Similar to the trachea, the developing lungs [which originate from two lung buds on each side of the tracheal bud (19)] fail to separate correctly from the gut and are still connected to the gut tube at E17.5. The importance of Hedgehog signaling to normal esophageal development was confirmed in Gli mutant mice. In the Gli2⫺/⫺ mouse, the esophagus has a very 1356 GIJS R. VAN DEN BRINK small lumen and little mesenchymal cells and fails to develop a smooth muscle layer. In Gli2⫺/⫺ Gli3⫹/⫺ mutant mice, most of the proximal esophagus is degraded, and only a very small proximal esophageal remnant can be observed in the carefully made whole-mount preparations shown by Motoyama et al. (138). A single tube connects the trachea and lungs to the stomach in Gli2⫺/⫺ Gli3⫹/⫺. Although this has not been carefully examined, this is most likely the remnant distal esophagus similar to human with esophageal atresia and tracheoesophageal fistula (Fig. 10). In Gli2⫺/⫺ Gli3⫺/⫺ mice, the foregut was hypoplastic and failed to form the appendages for the trachea and lungs at E9.5. Gli2⫺/⫺ Gli3⫺/⫺ embryos that survived until E14.5 had a very small remaining proximal endodermal tube and lacked both trachea and lungs. The phenotype of these Hedgehog pathway mutant mice indicates that Hedgehog signaling is critical to the maintenance of the developing esophagus and the formation of a tracheal-esophageal septum. Both phenotypes seem to result from a block in the mesenchymal growth that is necessary to support the developing esophagus and separate the gut tube from the airways. This is therefore a nice example of the critical role of endodermal-derived Hedgehog in radial patterning of the gut. The role of Hedgehog signaling in esophageal cytodifferentiation remains to be determined as the severe esophageal phenotype of most Hedgehog mutant mice precludes evaluation of esophageal cytodifferentiation, and the histology of the esophageal epithelium of Gli2⫺/⫺ mice has not been reported. Physiol Rev • VOL 2. Gastric development Gastric epithelial cytodifferentiation is a highly complex process during which the gastric epithelium is patterned into three distinct zones along the proximal-distal axis and in precursor cell and differentiated cell compartments along the vertical axis. Whereas the whole stomach of humans consists of glandular epithelium, mice develop a squamous epithelium in the first proximal one-third of the stomach. The distal two-thirds of the stomach consist of columnar glandular epithelium. A proximal fundic or zymogenic and distal antral zone can be recognized in the adult glandular epithelium based on histological features. Cytodifferentiation of the gastric epithelium is initiated around E13.5. At E16.5, the epithelium of the forestomach is a squamous multilayer, whereas the epithelium of the glandular stomach is a monolayer of columnar cells that have formed primitive epithelial invaginations (48). Recombination experiments (48) suggest that the endoderm of the E14.5 stomach does not require mesenchymal instructive signals for appropriate AP patterning, indicating that the AP pattern has been correctly specified at this point in time (as is the case in the intestine, see below). It should be noted, however, that no markers of differentiation were used and that epithelial phenotype was judged by histology. The authors also performed experiments with E11.5 and E12.5 endoderm and showed that the endoderm may still have some plasticity at this stage as 7 out of 10 E11.5/12.5 gastric endoderm explants keratinized when cultured with E14.5 forestomach mesenchyme, 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 FIG. 10. Hedgehog signaling is critical to normal foregut development. A schematic depiction of the phenotype of patients with tracheoesophageal fistula (A), wild-type (B), and Gli2⫺/⫺ Gli3⫹/⫺ (C) mice at E11.5. A: in the most common form of esophageal atresia and tracheoesophageal fistula, the proximal esophagus is missing and the distal esophagus connects to the lower trachea. C: In Gli2⫺/⫺ Gli3⫹/⫺ mice, the trachea and lungs fail to separate from the developing gut tube. The proximal esophagus becomes progressively hypoplastic until only a small remnant is present at E11.5. As a result, the trachea and lungs are connected to the gut tube at E11.5. In humans, a transition from tracheal tissue to esophageal tissue is found at the point indicated with an arrow in A and C. It is not entirely clear if the tube distal from the lungs, which connects the trachea and lungs to the stomach, consists of esophageal tissue in Gli2⫺/⫺Gli3⫹/⫺ mice as in humans. [B and C redrawn from Motoyama et al. (138).] HEDGEHOG SIGNALING 3. Hedgehog signaling in gastric development From early in stomach development at E11.5 until after the onset of gastric epithelial cytodifferentiation at E15.5, Shh is expressed at high levels in the forestomach and lower levels in the hindstomach, whereas Ihh is expressed in the hindstomach (Fig. 11) (13, 178). Ihh expression in the glandular stomach depends on Fgf signaling as both Fgfr2b⫺/⫺ and Fgf10⫺/⫺ mice lack expression of Ihh in the stomach (178). Expression of Ptc1 is in the mesoderm and mirrors that of Shh. There is only very low expression of Ptc1 in the distal hindstomach at this point FIG. 11. Hedgehog signaling during early gastric development. A: Shh is expressed in a gradient with high expression in the forestomach and low to absent expression in the hindstomach. Shh specifies the zone that will be occupied by squamous forestomach epithelium. Although Ihh is expressed in the hindstomach, it is probably not translated or otherwise inactive as the expression of Ptc in the mesoderm mirrors that of Shh. B: expression of Shh is restricted to the forestomach by FGF10 secreted from the mesoderm, which acts on the FGFR2b in the endoderm. FGF signaling lies upstream of another negative regulator of Shh expression, transcription factor Gata4. Activin signaling is another factor necessary to restrict Shh expression that could hypothetically act downstream of FGF signaling and upstream of Gata4. Physiol Rev • VOL in time, which suggests that Ihh may not be translated or active (178). Indeed, no gross abnormalities are reported in the stomachs of Ihh⫺/⫺ mice, although gland formation may be somewhat reduced [see Fig. 2I in Ramalho-Santos et al. (164)]. Expression of Shh expands to the hindstomach at later stages of development as both Shh and Ihh are expressed in the glandular stomach at E18.5 (164). At this stage of development, Hedgehog signaling may no longer be exclusive to the mesenchyme as both Ptc1 and Gli1 seem to be expressed also in the epithelial layer (164). This is similar to the situation in the adult stomach where Ptc1 is most highly expressed in epithelial gland cells (8, 199). Hedgehog expression in the forestomach has not been described during later stages of gastric development. The expression of Hedgehogs during epithelial cytodifferentiation and compartmentalization in the first four postnatal weeks has similarly not been examined. The analysis of gastric development in Hedgehog pathway mutant mice is limited by the fact that these mutants die well before or just after birth, whereas major events in cytodifferentiation occur postnatally. In Shh⫺/⫺ mice, the area occupied by the squamous epithelium of forestomach is reduced (99, 164). This forestomach phenotype is in accordance with the Shh expression pattern and suggests a role for Shh signaling in the specification of the forestomach domain. Indeed, in mice with mutant Activin receptors, the domain of Shh expression is expanded from the anterior to the posterior stomach with a concomitant posterior expansion of the squamous epithelium of the forestomach (101). A similar posterior expansion of Shh expression with posterior expansion of the squamous epithelium of the forestomach has been observed in Gata4⫺/⫺, Fgfr2b⫺/⫺, and Fgf10⫺/⫺ mice (78, 178). Fgf10 is expressed in the mesenchyme of the glandular stomach, whereas Fgfr2b and Gata4 are expressed by the endoderm of the glandular stomach (178). As Gata4 expression is lacking in both Fgfr2b⫺/⫺ and Fgf10⫺/⫺ mice (178), this places Fgf10 signaling from the mesoderm to the endoderm upstream of Gata4 in the restriction of Shh expression to the forestomach. It is not clear if FGF signaling interacts with Activin signaling in the suppression of Shh expression, but in the developing eyelid, Fgf10 is necessary for the expression of Activin (186) and endodermal Gata4 expression is induced by TGF- family members (1, 169). It is therefore possible that Fgf10 signaling induces Activin expression in the endoderm through Fgfr2b, which would in turn induce the expression of Gata4 in the glandular stomach. Shh also plays an important role in the glandular stomach as glandular growth is increased in Shh⫺/⫺ mice, resulting in mucosal hyperplasia of the glandular stomach (99, 164). Kim et al. (99) found that the gastric histology of Gli3⫺/⫺ mouse phenocopies that of the Shh⫺/⫺ mouse, indicating that the Shh signal is mainly transduced by Gli3 in the developing stomach. This shows that although Gli3 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 whereas 1 out of 9 were keratinized when cultured with mesenchyme from the glandular stomach. At birth, ⬃90% of the cells in the rudimentary gastric units are precursor cells; this percentage is reduced to 20% in the first 7 days postnatally (P1–P7) as cellular differentiation occurs while gland size remains stable (89). The second week after birth (P8 –P14) is marked by an increase of the number of all cell types and glandular growth. Between P15 and P28, further cellular differentiation and glandular growth occurs, and the cells in the glands are compartmentalized (89). Compartmentalization of the gastric glands results in the formation of a precursor cell compartment (isthmus) somewhere halfway up the unit from where a bidirectional migration of differentiating cells occurs. Cells that migrate to the luminal surface form the pit or foveolus, whereas cells that migrate to the bottom of the gland form the gland proper. 1357 1358 GIJS R. VAN DEN BRINK Physiol Rev • VOL gland formation completely, resulting in a squamous epithelium that lacks glands. In the glandular stomach much lower levels of Shh are expressed due to inhibition of Shh expression by Fgf10 secreted from the mesenchyme. The resulting low level of Shh in the glandular stomach allows gland formation but restricts gland branching. The development of gastric epithelial cell lineages seems relatively undisturbed in the absence of Shh, but there is a defect in the appropriate restriction of the expression of markers of the gastric cell lineage. Since Ihh is also expressed in the glandular stomach, there is redundancy in the Hedgehog pathway, and the phenotype in the absence of both Shh and Ihh may be more severe. Since the stomachs of Shh/Ihh double-knockout mice cannot be examined due to early lethality, further analysis of the role of Hedgehog signaling awaits conditional inactivation of Hedgehog pathway components in the stomach endoderm. 4. Intestinal development Differentiation of the intestinal endodermal layer occurs in two phases in rodents. The first phase is initiated at E15 in the mouse and lasts until birth. During this phase, a wave of differentiation occurs from the duodenum to the colon, transforming the pseudostratified cuboidal endodermal layer into a polarized columnar epithelial monolayer. The epithelium compartmentalizes into differentiating and proliferating cell populations. In the small intestine, this occurs by villus formation along the vertical axis of the gut. Differentiating cells are located on these fingerlike structures that project into the lumen of the gut and are extended by upward growth of the underlying mesenchyme. The formation of villi is initiated in the proximal duodenum at E14 and occurs in a proximal to distal wave similar to and just preceding cytodifferentiation (90). Villus morphogenesis is preceded by the formation of clusters of condensed mesenchymal cells that underlie the site of eventual villus formation (90). The villi are separated from each other by small patches of intervillus epithelium that contain a population of proliferating precursor cells. The colon lacks villi but develops epithelial folds at E16.5 (151) at the onset of colonic endodermal cytodifferentiation. These folds form in a manner that resembles small intestinal villus formation with similar mesenchymal condensation and subepithelial expansion (27). The epithelial folds lie more separated in the proximal colon (and therefore more villuslike), whereas they are increasingly packed together towards the distal colon. The endoderm holds all necessary information for correct patterning of the small intestine along the AP axis at the onset of cytodifferentiation at E14. Experiments using E15 whole intestinal explants that were transplanted subcutaneously in young adult syngenic mice (45) or nude mice (38, 171) have shown that the information 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 may often act as a truncated protein that represses targets of Hedgehog signaling, it seems to act mainly as a fulllength transcriptional activator in gastric development. This was further illustrated by the fact that the expression of both Ptc1 and Gli1, two transcriptional targets of Hedgehog signaling, was reduced in the stomach of Gli3 mutant mice (101). No change was found in endodermal proliferation in the gastric mucosa of Shh⫺/⫺ and Gli3⫺/⫺ mice between E14.5 and E18.5 that could account for the increased mucosal thickness (99, 164). Two different findings can explain this apparent discrepancy. It was found that apoptosis may be reduced in Shh⫺/⫺ and Gli3⫺/⫺ mice, which could result in net increased glandular growth. An alternative reason for the mucosal hyperplasia may be the strongly increased gland branching (99). As proliferation is judged per gland, an increase in gland branching will underestimate epithelial proliferation in Shh⫺/⫺ and Gli3⫺/⫺ mice. Ramalho-Santos et al. (164) reported histology reminiscent of intestinal transformation of the stomach epithelium in the Shh⫺/⫺ mouse. If true intestinal transformation (or in the adult: intestinal metaplasia) of the stomach would occur in Shh⫺/⫺ mouse, the mucosa should show the histological hallmark of intestinal transformation, the presence of goblet cells (211). However, there are no goblet cells in the stomach of Shh⫺/⫺ mice (99), and one can therefore not compare the phenotype to intestinal metaplasia of the stomach as it is known in the adult. The authors do show that there is expression of markers of intestinal enterocytes (alkaline phosphatase expression and binding of Wisteria floribunda-agglutinin lectin) in the gastric epithelium, but it was not examined if there is loss of gastric epithelial cell markers as is characteristic for intestinal metaplasia in the adult. In fact, Kim et al. (99) found that all gastric markers examined are normally expressed in the stomachs of Shh⫺/⫺ and Gli3⫺/⫺ mice. They further show that gastric epithelial cells do not have a brush border (see Ref. 99, Fig. 1i-l) as would be expected in the case of intestinal transformation. Also, there is no expression of intestinal enterocyte marker villin, intestinal homeobox gene Cdx2 (a marker of intestinal metaplasia in the adult), or goblet cell marker Muc2 in the stomachs of Shh⫺/⫺ and Gli3⫺/⫺ mutant mice (99). Together these data suggest that Shh signaling may be important to appropriately restrict the expression of markers of the gastric epithelial cell lineage but that there is no evidence of intestinalization or intestinal metaplasia. This loss of appropriate restriction of the expression of cell lineage markers has some similarities with our finding that in rats treated with Hedgehog inhibitor cyclopamine colonic enterocytes aberrantly express a marker of the goblet cell lineage [see Fig. 1u-x in van den Brink et al. (197)]. In conclusion, the available evidence suggests that high levels of Shh expression in the forestomach inhibit HEDGEHOG SIGNALING 5. Hedgehog signaling in intestinal development At E11.5 and E12.5, both Shh and Ihh are expressed throughout the endoderm of developing intestine (13). Shh expression is downregulated in the small intestine at E14.5 around the moment of intestinal villus formation Physiol Rev • VOL and cytodifferentiation, with the highest remaining expression in the duodenum (13). Ihh remains expressed in the developing small intestine and colon throughout development (13, 164). At E18.5, the expression of both genes is restricted to the intervillus epithelium in the small intestine (122, 164). In the colon Shh is expressed at the base of the crypts, whereas Ihh is expressed throughout the epithelium (164). Hedgehog signaling in the developing small intestine and colon is from the epithelium to the mesenchyme as Ptc1 and Gli1 are exclusively expressed in the mesenchyme during development and directly after birth at P0 (13, 122, 164). Both Shh and Ihh mutant mice show malrotation of the gut (164). Although there is no reversion of gut situs in Shh⫺/⫺ or Ihh⫺/⫺ mice, aberrant gut looping may be related to the role of Hedgehog signaling in left-right axis formation that is described above. Hedgehog signaling plays an important role in radial patterning in the developing intestine as it regulates villus formation, smooth muscle layer formation, and development of the enteric nervous system. Shh and Ihh seem to play a distinct role in villus formation. Shh is mainly expressed in the intervillus epithelium of the duodenum. In the duodenum of the Shh⫺/⫺ mouse, villi appear overgrown and clog the duodenal lumen, indicating that Shh may act to restrict villus growth in the duodenum. In contrast, villi in Ihh⫺/⫺ mice are decreased in number and hypoplastic. Madison et al. (122) used a 12.4-kb fragment of the villin gene to drive expression of Hhip in the small intestinal epithelium. The 12.4-kb villin promoter drives the expression of transgenes in the crypts and villi of the small intestine from E12.5 onwards (123). Unfortunately, the colon cannot be analyzed using the villin promoter as transgene expression is mostly absent or very low (123). The phenotype of villin-Hhip transgenic mice was more severe than that of Ihh⫺/⫺ mice. In mice with high levels of transgene expression, villus formation was completely abrogated, and regions of epithelium were observed that failed to organize in a single layer of polarized columnar epithelial cell but seemed instead to have remained pseudocolumnar. As the villin-Hhip transgene presumably blocks signaling by both Shh and Ihh, this indicates that the main role for Hedgehog signaling is in villus growth and development. However, the phenotype of the Shh null mouse suggests that Shh may play some role in restricting duodenal villus growth after their establishment. In mice with lower villin-Hhip transgene levels, the villi were thickened and showed extensive branching (Fig. 12). Branching was caused by the presence of dispersed cryptlike pockets of proliferating cells on the villi from which new villi seemed to originate. Thus Hedgehog signaling not only plays a role in induction of villus growth, but it also seems to act to restrict precursor cells to the intervillus epithelium. Interestingly, the phenotype of ectopic crypt formation on the villi was remark- 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 needed to regulate epithelial differentiation is intrinsic to the gut tube and independent of luminal content or developmental stage-specific hormonal regulation. It was even shown that when small intestinal segments of E14 rats were transplanted under the skin of nude mice, the timing of epithelial differentiation of the segments occurs according to the correct anterior-posterior wave of differentiation (38). Also, as discussed above, endoderm-mesenchymal recombination experiments indicate that the small intestinal endoderm differentiates according to its original position along the AP axis and independent from the type of mesenchyme with which it is recombined. Only the colonic endoderm inappropriately expresses small intestinal enzymes when recombined with small intestinal mesenchyme, indicating an instructive influence of the colonic mesenchyme at later stages of development. Interestingly, the immature colon epithelium expresses several markers of small intestinal differentiation and shows absorptive and digestive functions that are typical for small intestinal enterocytes. Apparently, instructive signals from the colonic mesenchyme are necessarily late in colonic development to suppress small intestinal cell fate. The second phase of small intestinal epithelial differentiation occurs after birth, extending from postnatal day (P) 1 to P28. During the first two postnatal weeks, the crypts of Lieberkühn are generated in the small intestine. These crypts replace the intervillus epithelium in a poorly understood process. The distance between the base of the intervillus epithelium and the circular smooth muscle layer remains constant during crypt formation (17). Crypt formation therefore seems to involve mesenchymal enveloping of the nascent crypt and upwards migration of the junction between the precursor cell compartment of the intervillus epithelium and the differentiated cells on the villi (17). This period coincides with the formation of morphologically recognizable Paneth cells (15). The number of crypts increases rapidly in the third postnatal week through crypt fissioning (17). This process of crypt multiplication involves the branching of a crypt to form two independent crypts and seems to result from symmetric stem cell division (54). At P28, crypt morphogenesis and multiplication is complete and the crypt contains a few Paneth cells at its base (in increasing number from the duodenum to ileum), one or more stem cells just above the Paneth cells and/or in between the Paneth cells, and a number of more or less committed precursor cells above the stem cell position. 1359 1360 GIJS R. VAN DEN BRINK FIG. 12. Hedgehog signaling regulates villus formation in the small intestine. A schematic representation of the P0 small intestine of wild-type (A) and villin-Hhip transgenic and anti-Hedgehog antibody-treated mice (B) (122, 204). In the wild-type newborn intestine, proliferating and differentiated cells have compartmentalized. Differentiated enterocytes populate the villi, whereas proliferating precursor cells (characterized by activity of the Wnt pathway) localize to the epithelium between the villi. In villin-Hhip mice with low transgene expression, this compartmentalization is disturbed, and ectopic patches of precursor cells are found on the villi resulting in a branching phenotype. Physiol Rev • VOL ated via an effect on the precursor cells or the differentiated cells. It could be, for example, that Hedgehog signaling is necessary to repress Wnt signaling and that precursor cells fail to exit from the cell cycle in the absence of Hedgehog signaling. Alternatively, Hedgehog signaling may be necessary for the development and/or survival of cells that have already exited the cell cycle. That would result in a similar accumulation of precursor cells in the epithelium of the villin-Hip mouse secondary to a lack of differentiated cells. An experiment in Xenopus may point towards the first possibility. In Xenopus that were injected with a constitutive active form of Smo, the endoderm was completely absent from the small intestine, suggesting that Hedgehog signaling may upregulate a factor in the mesenchyme that suppresses endodermal precursor cell fate. A major target for Hedgehog signaling at later stages of intestinal development may be the intestinal subepithelial myofibroblasts (ISEMFs) (158). ISEMFs are ␣-smooth muscle antigen (␣-SMA)-positive fibroblast-like cells that lie closely apposed to the basal membrane of the intestinal epithelium of the colon and small intestine. Madison et al. (122) have shown that ISEMFs respond to Hedgehog signaling in vitro. The authors further showed that ISEMFs are increased in number in the mucosa of villin-Hhip mice and abnormally localized in the villi where they were found in close proximity to the ectopic cryptlike pockets of precursor cells (122). These findings have an interesting link with very elegant work performed by Ormestad et al. (146). Ormestad and colleagues showed that Foxf1 and Foxf2 transcription factors are mesenchymal Hedgehog target genes that play a critical role in ISEMFs. At E18.5, Foxf1 and FoxF2 mutant mice show impaired extracellular matrix formation, loss of endodermal differentiation, accumulation of epithelial precursor cells characterized by increased endodermal Wnt signaling, and accumulation of ISEMFs (146). In all of these aspects, the phenotype strongly resembles the phenotype of the villin-Hhip 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 ably similar to the phenotype described in mice that expressed the BMP antagonist noggin behind a villin promoter and mice in which the Bmp receptor 1a was deleted (58, 60). It has been shown that Bmp4 is a mesenchymal target of Hedgehog signaling in the small intestine of the chick (167, 168) and mouse (123, 202), suggesting that epithelial Hedgehog may be upstream of mesenchymal Bmp4 in the control of precursor cell compartmentalization. One finding that argues against this possibility is that the ectopic crypt phenotype in the villin-Noggin mouse only becomes evident 3 wk after birth, directly after the completion of intestinal epithelial maturation and crypt formation (however two different villin promoters were used in these studies) (58). The epithelial phenotype of the villin-Hhip mouse showed a substantial reduction of the number of differentiating cells and accumulation of precursor cells very similar to phenotype of postnatal mice treated with a Hedgehog-blocking antibody that had been described before (204) (Fig. 12, discussed below). In the mice that had lower Hip transgene expression, a normal presence was observed of the two secretory cell lineages that are present in the first postnatal week (Goblet cells and endocrine cells), whereas differentiation of the enterocyte cell lineage was reduced with a poorly developed brush border and loss of expression of brush-border markers. This important role of Hedgehog signaling in enterocyte differentiation correlates with our own experiments (197) in which we have shown that Hedgehog signaling regulates enterocyte differentiation in the distal colon of the adult rat (see below). The epithelial phenotype is indirect, since Hedgehog signaling is exclusively from the epithelium to the mesenchyme in the developing and early postnatal small intestine and therefore mediated via an as yet unidentified mesenchymal factor that is controlled by Hedgehog signaling. An important question is if the increase in the number of precursor cells in the villin-Hhip mouse is medi- HEDGEHOG SIGNALING Physiol Rev • VOL mal cytodifferentiation. Both Bmp4 and Wnt5a are good candidates as mediators of this reciprocal signal. 6. Hedgehog signaling in anorectal development The anorectum develops with a complex interplay of all three germ layers as it forms at the junction of the endoderm and ectoderm (32, 168). Hedgehog signaling is critical to normal anorectal development. Shh⫺/⫺ (and not Ihh⫺/⫺) mice maintained by Ramalho-Santos et al. (164) displayed an imperforate anus phenotype. The colon of Shh⫺/⫺ mice ended in a blind sack and did not form a connection with the perineum. Shh⫺/⫺ mice maintained by Mo et al. (136) displayed a failure to separate the anorectum from the lower urinary tract, that drained in a common cavity or outlet. Gli2⫺/⫺ mice have an imperforate anus with a rectovesical fistula (between the distal intestine and the bladder) and a single urethral opening in the perineum [see also Kim et al. (98)]. The phenotype of Gli3⫺/⫺ mice is very mild with slight anal stenosis. In Gli2⫺/⫺ Gli3⫹/⫺ and Gli2⫹/⫺ Gli3⫺/⫺ compound mutant mice, a persistent cloaca was observed and a common outlet for the digestive and urogenital tract (136). These phenotypes indicate that although Gli3 does play a role in the transduction of the Shh signal to the anorectal mesenchyme, the contribution of Gli2 is much more important similar to the role of Gli signaling in foregut development. 7. Hedgehog signaling in pancreas development As discussed above, exclusion of Shh and Ihh expression from the prospective pancreatic endoderm plays a critical role in early pancreatic development. Hedgehog signaling does play a role at later stages in development however. Although no Shh is expressed during pancreatic development, Ihh and Ptc1 are expressed in the pancreas as detected by RT-PCR from E13.5 onwards, but their expression has not been localized (and Dhh has not been examined). Both Ihh and Dhh are expressed in the islets of the adult pancreas (62, 191). Ptc1 is expressed in the islets and ducts of the adult pancreas (62), suggesting that Hedgehog signaling in the islets is autocrine. Ihh seems to play a role in the morphogenetic movements of the ventral and dorsal pancreatic buds as almost half of Ihh⫺/⫺ mice develop an annular pancreas in which an extension of the ventral pancreas encircles the duodenum (62, 164). Histologically, the pancreas of Ihh mutant mice appears intact and contains normal islets. Since it is not known if Dhh is expressed during the later stages of development of the pancreas, it is not clear if presence of Dhh may rescue a phenotype. More careful analysis of pancreatic cellular differentiation in Shh⫺/⫺ and Ihh⫺/⫺ mice appeared to show a relative increase in the number of endocrine cells in the pancreas, especially in the Ihh⫺/⫺ mice. The authors of the study expressed the number of 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 mouse, suggesting that Foxf1 and Foxf2 may be the major effectors of Hedgehog signaling in the mesenchyme. Interestingly, in Foxf2⫺/⫺ mice, Bmp4 is completely absent from the mesenchyme, suggesting that loss of mesenchymal Bmp4 expression may indeed explain the endodermal phenotype in villin-Hhip transgenic mice as discussed above. Together these data suggest that endodermal Hedgehog signaling induces Foxf transcription factors in the mesenchyme, which in turn induce mesenchymal Bmp4 expression which directly acts on the epithelium to restrict endodermal precursor cell accumulation and stimulate endodermal differentiation. An alternative scenario is discussed by Ormestad et al. (135); they show that mesenchymal Bmp4 acts to suppress mesenchymal Wnt5a expression and that Wnt5a expression is strongly induced in Foxf2⫺/⫺ mice. The authors suggest that this excess Wnt5a may directly be responsible for increased endodermal Wnt signaling. This is an interesting possibility that remains to be tested as Wnt5a can both activate and repress canonical Wnt signaling depending on the receptor context (135). Both Ihh and Shh mutant mice display a reduced thickness of the circular smooth muscle layer that surrounds the small intestine and distinct defects in the development of the enteric nervous system (164). Both phenotypes are not present in the villin-Hhip mouse. As a result, the Hirschsprung’s diseaselike phenotype that has been described in Ihh⫺/⫺ mutant mice (164) is absent from villin-Hhip mice. The most likely explanation for this discrepancy is that the effect of Hedgehog signaling on the size of the developing smooth muscle layer and development of the enteric nervous system is mainly mediated before expression of the Hhip transgene (i.e., before E12.5). Another possibility is that long-range Hedgehog signaling to the smooth muscle layer and developing enteric nervous system is not affected by the Hhip transgene. Wang et al. (204) have studied the role of Hedgehog signaling in postnatal development of the small intestine by injecting newborn pups with a Hedgehog blocking or control antibody. Anti-Hedgehog antibody-treated mice developed steatorrhea due to lipid malabsorption and a runting phenotype. The effect on villus morphology was similar to that later described by Madison et al. (122) in the prenatal small intestine with shortened villi that show increased branching, precursor cell accumulation in the intervillus epithelium, and spreading of precursor cells on the villus (Fig. 12). In conclusion, Hedgehog signaling acts on the mesenchyme in the developing small intestine. It regulates the differentiation of ISEMFs, the smooth muscle layer, and the developing enteric nervous system. Hedgehog signaling induces the expression of a mesenchymal factor that acts reciprocally on the developing endoderm and plays a critical role in precursor cell homeostasis and/or endoder- 1361 1362 GIJS R. VAN DEN BRINK 8. Hedgehog signaling and the VACTERL association Many of the developmental abnormalities that have been described in Hedgehog mutant mice above are also found in human patients. These patients have a variety of nonrandomly associated developmental abnormalities that occur sporadically and are known under the acronym VACTERL. VACTERL stands for vertebral defects (V), anal atresia (A), cardiac defects (C), tracheoesophageal fistula (TE), renal anomalies (R), and limb defects (L) (97, 131, 161, 166). Although these abnormalities are often found together, they are found in varying constellations and the VACTERL association is not recognized as a specific syndrome. One study that examined 2,493,999 births only reported a single case with combined recognized deformities in all 6 organs (166). However, all of these defects have been found in one or more of the Physiol Rev • VOL several known human and murine mutants of the Hedgehog pathway (73, 86, 100, 117, 136, 138, 162, 164) [see Kim et al. for review (98)]. The atresia or stenosis of the esophagus with associated tracheoesophageal (TE) fistula that was found in Hedgehog mutant mice is observed in ⬃1 in 2,000 –5,000 live human births (97). Just like in the Hedgehog mutants this anomaly can be found in association with other gastrointestinal malformations in humans such as anal atresia, midgut malrotation, duodenal atresia, and annular pancreas (3) and with malformations of other organs as part of the VACTERL association mentioned above. Even though a strong similarity exists between patients with the VACTERL association and Hedgehog mutant mice, mutations have not yet been described in patients. The VACTERL association shows considerable overlap with the Feingold syndrome (21). Feingold syndrome is caused by germline mutations in N-myc (195), which is an established target of Hedgehog signaling (96). Although no mutations in N-myc have been found in patients with the VACTERL association, this suggests that mutations in the Hedgehog pathway or important targets of Hedgehog signaling may indeed be its cause. IV. HEDGEHOG SIGNALING AND HOMEOSTASIS OF THE ADULT GUT A. Morphogenesis Versus Morphostasis As discussed above, the developing embryo is patterned along several different axes. This patterning involves inductive signals that are exchanged between different germ layers through morphogenetic signaling pathways that are highly conserved throughout evolution. It is important to realize that cell fate regulation along different spatial axes is not a feature unique to the developing embryo. Cell fate is similarly regulated in a positiondependent manner along different axes in rapidly regenerating epithelial tissues such as those of the gastrointestinal tract (63). Equally spaced stem cell niches along the length of the gut continuously generate a fresh supply of epithelial cells. Cell fate of stem cells and their descendants is regulated along the vertical axis of the gut. The second axis of cell fate determination is the longitudinal axis (esophagus, stomach, small intestine, colon), which determines the phenotype of epithelial cells that are generated by a particular stem cell. It is increasingly being recognized that morphogens not only play a role in the spatial regulation of cell fate during morphogenesis but also play a critical role in the maintenance of tissue homeostasis or morphostasis in the adult along these two different axes. This also explains why mutations in morphogenetic pathways such as the Wnt and BMP pathway can initiate carcinogenesis (10, 58, 60, 69). Knowledge of 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 endocrine cells relative to body weight (62). This distorts the calculations as body weight was much reduced in Shh⫺/⫺ mice, whereas the pancreas was of normal size (as may be expected given the absence of Shh expression at any point in pancreatic development). If the number of endocrine cells is expressed per pancreas weight, their relative number is only modestly increased in Shh⫺/⫺ mice (22% increase), whereas the number of endocrine cells/pancreas weight is increased by 75% in the Ihh⫺/⫺ mice (62). This seems more consistent with a role for Ihh signaling in later pancreatic development and suggests that Ihh acts as a negative regulator of the endocrine cell fate in the developing pancreas. This finding was corroborated by the phenotype of mice that lack the Hedgehog antagonist hedgehog interacting protein (Hhip). In Hhip⫺/⫺ mice, excess Hedgehog signaling in the pancreas results in loss of endocrine cells (92). Similarly, overexpression of Shh or Ihh in the pancreas from E13.0 onwards using a Pax4 promoter resulted in excess mesenchymal growth, inhibition of epithelial expansion, and loss of endocrine cells. At E18.5, this resulted in a pancreas that was ⬃60% reduced in size mainly due to a dramatic reduction in epithelial cell content (93). In conclusion, all mouse mutants in the Hedgehog pathway that have been analyzed show that Hedgehog signaling represses endocrine cell fate at later stages of pancreatic development. Interestingly, Pax6, a gene that is repressed by Hedgehog signaling in the developing neural tube (42), pituitary gland (104), and retina (154), plays a critical role in pancreatic development. Pax6 is a transcription factor that is expressed at least from E9.0 onwards in the pancreatic endoderm (180), and mice that lack Pax6 have reduced numbers of endocrine cells and disturbed islet morphology (172, 180). It would be interesting to see if the observed negative regulatory effect of Hedgehog signaling may (in part) be mediated through repression of Pax6 expression. HEDGEHOG SIGNALING the role of Hedgehog signaling in the adult gut is still limited, since it has received little attention to date. Here I will discuss what is currently known about the role of Hedgehog signaling in adult gut and how this pathway may be deregulated during gastrointestinal carcinogenesis. B. Stomach Physiol Rev • VOL Recently, Fukaya et al. (49) performed a careful analysis of Hedgehog pathway component mRNAs in microdissected pit isthmus and gland cells of the human stomach. Unfortunately, the exact provenance of the specimens in this study was not specified; it is therefore not entirely clear if all specimens shown are of fundic origin, which makes the interpretation of some of the results somewhat difficult. Their results showed that all three Hedgehogs can be detected by PCR in gastric epithelium in humans. The authors found Ihh expression in gastric pit cells and Shh and Dhh expression in the gastric gland. Ihh was expressed by the differentiated pit cells as determined by immunohistochemistry, whereas both Shh and Dhh localized to parietal cells immunohistochemically. Given the fact that the Dhh antibody used in this study recognizes the 19-kDa active fragment of Dhh which is extremely conserved between all Hedgehogs, it may also be that the anti-Dhh antibody cross-reacts with Shh in parietal cells, and this result therefore needs to be confirmed by in situ hybridization. In the hands of Fukaya et al. (49), a Ptc1 antibody reacted only with gastric pit cells. This result contradicts previously obtained results with two different Ptc1 antibodies (C-20 from Santa Cruz and an antibody obtained from Dr. R. Toftgard) (36, 199), in situ hybridization (141), and a Ptc1-LacZ reporter mouse (8) which all localized Ptc1 to the gastric glands of the gastric fundus. Together these results may suggest that Ptc1 is mainly expressed in the fundic glands but that low levels may be present in the fundic pit cells where it is induced by Ihh that is produced by differentiated pit cells. This conclusion is supported by the observation that cyclopamine treatment depletes pit cells from primary cultures of mouse gastric epithelial cells (49). Although Hedgehog signaling maintains proliferation of some gastric cancer cells (see below), we found that treatment of mice with cyclopamine increased epithelial proliferation in vivo by 60 –70% (199). Similarly, Fukaya et al. (49) observed that treatment of primary cultures of normal mouse gastric epithelial cells with Smo antagonist cyclopamine doubled their proliferation. This may indicate that there is a difference in the role of Hedgehog signaling in cell cycle regulation between normal and cancer cells. More likely, one of the factors that may explain the difference observed in the response of gastric carcinoma cells and primary gastric epithelial cells in vivo and in culture is that the effects on cancer cells have been observed in pure populations of cultured cancer cells, whereas both our in vivo experiment and the primary cultures of gastric epithelial cells of Fukaya et al. included all gastric cell types and stromal cells. The fact that Ptc1 is not expressed in the isthmus (49) (which contains the precursor cells) suggests that Hedgehogs may not act directly on precursor cells and that the observed effects on proliferation are indirect and mediated via effects of 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 One month after birth a stable regenerating system is established in which mucus-secreting pit cells migrate towards the lumen from the precursor cell compartment in the isthmus (87, 109). The proximal glandular stomach (fundus) has small pit regions and large glands, whereas the distal stomach (antrum) has large pit regions and small glands. Glands are composed of several cell types that migrate down to the gland base from the stem cell position. Fundic glands are composed of parietal cells which secrete acid, endocrine cells, and mucous neck cells, a cell type with uncertain function and which transdifferentiates when halfway down the gland into zymogenic cells that secrete digestive enzymes (88). Antral glands are exclusively composed of mucous and endocrine cell types (110). We found that Shh protein and mRNA were expressed in the parietal cells of the fundic glands in humans (198, 199). The protein formed a concentration gradient with highest expression in the parietal cells closest to the isthmus and gradually decreasing expression as the parietal cell matured towards the base of the gland (199). In contrast to the restricted expression pattern in humans, we found Shh in a variety of gland cells in rats and mice (199), a finding that was confirmed by in situ hybridization for Shh in the Mongolian gerbil (181). No Shh protein or mRNA is found in the glands of the antrum (198). Hedgehog receptor and transcriptional target Ptc1 was similarly expressed in the epithelium of the gastric fundic glands (199). Treatment of mice with Smo antagonist cyclopamine resulted in downregulation of potential Hedgehog targets such as islet-1, Bmp4, and Foxa2 (Hnf3). Bmp4 was expressed in mesenchymal fibroblast-like cells and Foxa2 mainly by parietal cells with much weaker staining in other gastric epithelial cell types. These results indicated that fibroblasts and parietal cells were the major targets of Hedgehog signaling in the fundus. The role of Hedgehog signaling in parietal cell physiology was further explored in isolated canine parietal cells by Stepan et al. (179). It was shown that Shh expression in parietal cells could be strongly induced by EGF and that Shh positively regulates the expression of the H⫹-K⫹-ATPase ␣-subunit through Gli binding elements in the promoter region and potentiates histamineinduced gastric acid secretion. Treatment with cyclopamine completely blocked EGF-induced H⫹-K⫹-ATPase expression but not induction of the c-Fos promoter, indicating that induction of Shh mediates some but not all aspects of EGF signaling in parietal cells. 1363 1364 GIJS R. VAN DEN BRINK Hedgehog signaling on stromal cell types or on the differentiated epithelial cells. In conclusion, much work remains to be done in understanding the role of Hedgehog signaling in the adult stomach. Progress will be made by more careful analysis of expression patterns of Hedgehog pathway components by in situ hybridization and the study of inducible mutant mice of different components of the Hedgehog pathway. So far the available data have shown a role for Shh signaling in parietal cell physiology and suggest that Ihh may be involved in the differentiation and maintenance of the pit cell lineage. The adult small intestinal epithelium is established after the completion of crypt formation in the third postnatal week. A stem cell just above the base of the crypts generates enterocytes, goblet cells, and endocrine cells that migrate up towards the villus and Paneth cells that migrate down and fill the base of the crypts (52). Very little is known about the role of Hedgehog signaling in the adult small intestine. We found that low levels of Shh mRNA can be detected just above the Paneth cell position in the crypt, but expression levels are too low to be detected by immunohistochemistry when using gastric specimens as a positive control (198). This low expression of Shh in the small intestine and colon compared with the stomach was confirmed by others by quantitative PCR (181). In situ hybridization by Batts et al. (6) showed that Ihh is expressed at the crypt-villus junction with gradually diminishing expression towards the tip of the villus, whereas Ptc1 is expressed at low levels in the mesenchyme (Fig. 13). Ihh protein is mainly expressed by the enterocytes on the upper half of the villus (80). This partially overlapping expression pattern of mRNA and protein is typical for many enterocyte genes and explained by the fact that cells that have moved to the top of the villus often no longer transcribe mRNA from a gene but still express the translated protein. Immunohistochemical analysis suggests that rare Ptc1-positive epithelial cells may also exist around the stem cell position just above the Paneth cell (202). Varnat et al. (202) found Ihh protein expression in the Paneth cells at the base of the crypts; it is not entirely clear how this can be reconciled with the in situ hybridization and immunohistochemistry done by others. This issue needs to be resolved by further experimentation that would also take into account the exact provenance of the small intestinal tissue along its AP axis as the concentration of Hedgehogs diminishes towards the distal small intestine (204), and differences in the expression patterns of different Hedgehog pathway components may exist along the AP axis of the small intestine. Physiol Rev • VOL FIG. 13. Hedgehog signaling in the adult small intestine. Ihh is the predominant Hedgehog expressed in the adult small intestine. Ihh mRNA is expressed at highest levels at the crypt villus junction, with diminishing levels towards the top of the crypt. Ihh protein is expressed from halfway up the villus to its tip. Ptc mRNA expression is mainly in the mesenchyme, but rare Ptc protein-expressing cells may exist in the crypts. Shh mRNA is expressed by a few epithelial cells in each crypt. We found that treatment of mice with cyclopamine results in a modest ⬃10% reduction of proliferation in the duodenum. The only available data concerning the role of Hedgehog signaling in the adult small intestine are from a study by Varnat et al. (202). These authors studied the role of Ppar (⫽Ppar␦) in small intestinal homeostasis and showed that Ppar is important for Paneth cell differentiation. The expression of Ihh mRNA and protein was induced approximately threefold in Ppar null mice, and Ihh mRNA could be strongly reduced by treatment of wild-type mice with a Ppar agonist, showing that Ppar negatively regulates Ihh expression. Treatment of wildtype and Ppar⫺/⫺ mutant mice with cyclopamine increased the number of Paneth cells in the duodenum, suggesting that at least part of the effect of Ppar is mediated through repression of Ihh expression. This effect was recapitulated in vitro as the expression of lysozyme, a Paneth cell marker, was reduced in HT-29 colon cancer cells by treatment with recombinant Shh, whereas treatment with cyclopamine increased lysozyme expression (202). D. Colon The mucosa of the colon is covered by a flat epithelium that lacks a villus structure but contains numerous crypts. Three cell types exist in the adult colon: absorptive enterocytes, goblet cells, and endocrine cells. The 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 C. Small Intestine HEDGEHOG SIGNALING Physiol Rev • VOL showed loss of markers of enterocyte differentiation such as brush-border staining for villin and carbonic anhydrase IV but inappropriately expressed intestinal trefoil factor (Itf), a marker of the goblet cell lineage. These data suggested that Hedgehog signaling is necessary for enterocyte maturation and the appropriate restriction of the enterocyte transcriptome to markers of the enterocyte lineage. This phenomenon of the inappropriate induction of Itf expression in the enterocyte lineage may be similar to the induction of alkaline phosphatase expression in the gastric epithelium that has been observed in the developing stomach of Hedgehog mutant mice (discussed above). Interestingly, since Ptc1 seems to be expressed uniquely in the mesenchyme underlying the differentiated enterocytes (at least in the mouse), this effect on enterocyte differentiation is likely indirect similar to the effect on epithelial differentiation in the developing small intestine observed in the villin-Hip mouse (discussed above). We have confirmed the role of Hedgehog signaling in enterocyte maturation in the HT-29 cell differentiation model. This model is based on the observation that HT-29 cells grow into a confluent monolayer and can be made to differentiate upon glucose withdrawal or stimulation with butyrate, a short-chain fatty acid (5, 218). We found that butyrate induces Hedgehog expression in HT-29 cells. We further demonstrated that Hedgehog signaling is necessary and sufficient for induction of differentiation markers p21 (WAF/CIP1) and villin in HT-29 cells, experiments that have been confirmed by Varnat et al. (202). HT-29 cells do express PTC1 and GLI1 mRNA (160, 217), explaining the observed effects of Hedgehog on HT-29 cells. It is, however, questionable if experiments examining a direct effect of Hedgehog signaling on epithelial cells are of any relevance since we fail to detect Ptc1 mRNA in the colonic epithelium of mice in vivo (unpublished observations discussed above), making it much more likely that a so far unidentified factor from the mesenchyme mediates the observed effects on enterocyte differentiation in vivo. Clearly additional experiments are required to reassess PTC1 expression and examine GLI1 expression in normal human colonic epithelium and colorectal cancers using in situ hybridization to avoid problems of antibody aspecifity. In rats treated with cyclopamine, proliferation increased by ⬃50% similar to the effect observed in the mouse stomach and in contrast to the effect on the small intestine where we observed a modest reduction of epithelial proliferation. As in the stomach, this probably reflects the fact that differentiating epithelial cells and not the precursor cells are the major target of Hedgehog signaling in vivo or that the effect on proliferation is indirectly mediated via the stroma in vivo as our recent in situ hybridization for Ptc1 seems to suggest. In contrast to the inhibitory effect of Hedgehog signaling on colonic precursor cells studied in a whole tissue context in vivo, 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 proximal (ascending) and distal (descending) colon have a different histology. Histology of the proximal colon of the mouse has been poorly studied; the mucosa is organized in large folds, and epithelial precursor cells are localized halfway up the crypts, indicating that some cells migrate down towards the base of the crypt. In the distal colon precursor cells localize to the bottom of the crypt, and the different cell lineages are present in gradients (22). Enterocytes differentiate in a gradient from the base to the top of the crypt, and differentiated enterocytes are found at the top of the crypt and on the intercrypt tables. Goblet cells are mainly found in the midcrypt region, whereas most endocrine cells are found at the base of the crypt (22). We found very low levels of Shh at the base of the crypts by in situ hybridization. Similar to the small intestine, we have not been able to detect Shh protein in these cells by immunohistochemistry, using gastric biopsies as positive control (198). As mentioned above, this correlates with data from Suzuki et al. (181), who have shown by quantitative PCR that levels of Shh are very low in the small intestine and colon compared with the stomach. Using different antibodies we detected a strong signal for Hedgehog protein in the absorptive enterocytes of the distal colon (Fig. 14) (197). These same cells were positive for Ihh by in situ hybridization showing that differentiated enterocytes of the distal colon produce Ihh. We have previously described Ptc1 protein expression in the epithelium and mesenchyme of the human, rat, and mouse colon (197) and Ptc1 mRNA in the epithelium and mesenchyme of the distal human colon (141), suggesting that Hedgehog signaling acts both paracrine and autocrine in the adult colon. As some doubts were raised about the specificity of the anti-Ptc1 antibodies, we have now performed novel in situ hybridizations for Ptc1 in the mouse colon and anorectum and find that although Ptc1 mRNA is expressed in the epithelium of the anorectum, no Ptc1 mRNA is detected in colonic epithelium (unpublished observations). Instead, a clear Ptc1 mRNA signal is detected in the mesenchyme underlying the Ihh-positive differentiated colonic enterocytes in the mouse. This clearly suggests that Hedgehog signaling in the adult colon of the mouse is uniquely from the epithelium to the mesenchyme similar to the adult and developing small intestine (discussed above) and not partially autocrine as we have suggested previously based on immunohistochemical results (197). We will also reexamine the human colon as it is possible that differences between species exist and that PTC1 mRNA is expressed in the human colonic epithelium as we have found previously (141). Additionally, Gli1 localization will have to be examined by in situ hybridization in both mice and humans. In rats treated with Smo antagonist cyclopamine, enterocyte maturation was disturbed in the distal colon, whereas goblet cells matured normally. The enterocytes 1365 1366 GIJS R. VAN DEN BRINK Hedgehog signaling seems to stimulate proliferation if primary colonic enterocytes are isolated from their context and cultured in pure populations (145). This opposite effect of Hedgehog signaling on isolated precursor cells and precursor cells in vivo is similar to the situation in the stomach discussed above. It indicates that some effects of Hedgehog signaling on epithelial cells may be indirect and mediated via the stroma and that care has to be taken in the interpretation of results when isolated gastrointestinal precursor cells are studied. One of the effects we observed when looking for possible targets of Hedgehog signaling in the distal colon Physiol Rev • VOL was that the expression of Bmp4 was increased in the epithelium of cyclopamine-treated rats. In the distal colon Bmp4 is an epithelial Wnt target that is expressed mainly at the base of the crypt (196). Signaling by the Wnt pathway is critical in the regulation of intestinal precursor cell maintenance and proliferation (55, 196). When we examined the Bmp4 expression pattern in cyclopamine-treated rats, we observed that the normal expression gradient was lost and high Bmp4 expression extended all the way up to the abnormally differentiated enterocytes at the top of the crypt (197). These data suggested that Hedgehog signaling may act to restrict Wnt signaling to the base of 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 FIG. 14. Hedgehog signaling in the adult colon. Hedgehog protein is expressed by the differentiated colonic enterocytes at the top of the crypt in the human (A) and rat (B) distal colon. C–J show colon of control rats (C, E, G, I) and rats treated with Smo inhibitor cyclopamine (D, F, H, J). C: in controls, nuclei of the differentiated enterocytes at the top of the crypt have a dense slender appearance. D: in cyclopamine-treated rats, differentiation of the enterocytes is disturbed (open arrowheads). A few remaining normal enterocytes can be observed (closed arrowhead). E: normal colonic enterocytes express the brush-border enzyme carbonic anhydrase IV (CAIV; arrows). F: expression of CAIV is lost in cyclopaminetreated rats (remaining normal cells with brush-border staining indicated with arrows). G and I: in control rats, the expression of intestinal trefoil factor (ITF) is appropriately restricted to the goblet cell lineage (asterisk in I indicates ITF positive goblet cell). H and J: in cyclopamine-treated animals, expression of ITF is induced in the enterocyte lineage (arrows indicate ITF positive enterocytes). [From van den Brink et al. (197), with permission from Nature Publishing Group.] HEDGEHOG SIGNALING the crypt. Experiments are currently underway in our laboratory to confirm this negative effect of Hedgehog signaling on the Wnt pathway in conditional Hedgehog pathway mutant mice in vivo. In conclusion, differentiated colonic enterocytes of the distal colon produce Ihh which plays a role in their appropriate maturation. Given our recent finding that Ptc1 is expressed in the mesenchyme and not the epithelium of the mouse colon (unpublished observations), this effect of enterocyte differentiation is likely the result of epithelial-mesenchymal interactions, but additional experiments are required to address this issue. A. Evidence for a Role for Hedgehog Signaling in Initiation of Gastrointestinal Carcinogenesis? The role of Hedgehog signaling in carcinogenesis of several tissues has become evident from the analysis of patients with inherited cancer syndromes and genetically modified mice. Mutations in the Hedgehog receptor PTC1 lead to constitutive activity of the Hedgehog pathway and cause basal cell carcinomas (BCC) and medulloblastomas in patients with Gorlin syndrome (57, 79), a phenotype that is recapitulated in mice with the same mutations. The relevance of PTC1 mutations for the biology of these cancers is underscored by the fact that these activating mutations in the Hedgehog pathway are also found in sporadically occurring cases of the same cancers (155, 163, 194, 209). Two germline and two sporadic mutations in the intracellular Hedgehog signaling antagonist Suppressor of Fused have been described in 46 children with medulloblastoma (187). Subsequent screening for SUFU mutations in 2 independent sets of 134 (105) and 33 medulloblastomas (140) found 0 and 2 SUFU mutations, respectively, suggesting that SUFU mutations exist but are a rare phenomenon. The Hedgehog mutant mice that have been analyzed thus far do not develop gastrointestinal cancers except in the pancreas, indicating that it is unlikely that overactivation of the Hedgehog pathway is sufficient to initiate carcinogenesis in most of the gastrointestinal tract. In fact, a mouse with an inducible constitutive active form of Smo developed rhabdomyosarcoma, BCC, and medulloblastoma as expected, but no neoplastic lesions were observed in the esophagus, stomach, biliary epithelium, small intestine, or colon (128). The only gastrointestinal organ with a role for Hedgehog signaling in tumor initiation may be the pancreas. Thayer et al. (190) have shown that Shh is overexpressed in human preneoplastic pancreatic lesions and reported that a mouse that expressed Shh behind a pancreas specific promoter developed preneoPhysiol Rev • VOL plastic lesions, so-called PanIN lesions [for review, see Hezel et al. (64)]. In a recent consensus report on mouse models of pancreatic cancer it was concluded that although the pancreas of the Pdx1-Shh mouse shows intestinal metaplasia and cellular atypia, no epithelial lesions were observed that resemble human PanIN lesions (70). A mouse with inducible activation of a dominant active GLI2 gene (152) developed undifferentiated pancreatic tumors that showed invasive growth. Similar to the Pdx1Shh mouse, no lesions formed in these mice that resembled human preneoplastic lesions, arguing for an alternative pathway of tumorigenesis. Activating mutations in RAS play an important role in pancreatic carcinogenesis in humans and mice, with such mutations recapitulating many aspects of the disease, including the formation of PanIN lesions (64, 70). When the dominant active GLI2 inducible mice were crossed with mice that express an inducible active Ras mutant (KrasG12D) in a pancreas specific manner, dominant active GLI2 increased the formation of PanIN lesions, and mice succumbed from invasive cancers within weeks after birth (152). This suggests that activation of Hedgehog signaling is not an initiator of pancreatic carcinogenesis but plays an early role in pancreatic tumor progression in humans. Interestingly, Pasca di Magliano et al. (152) found that mutant Ras strongly induced pancreatic expression of both Shh and Ihh. This suggests that Hedgehog ligand overexpression may be one of the first steps that facilitate Ras-mediated carcinogenesis in the pancreas. In contrast, in two organs that seem to express high levels of Hedgehog protein in the adult gut, expression is lost rather than increased in preneoplastic lesions. In the proximal stomach, expression of SHH is reduced by Helicobacter pylori infection and lost in intestinal metaplasia and gastric atrophy, factors that predispose to gastric carcinogenesis (36, 175, 198). A similar loss of Shh expression was observed in the Mongolian gerbil model of Helicobacter pylori-associated gastric carcinogenesis (181). Likewise, in the distal colon, IHH expression is lost very early in the process of the adenoma to carcinoma sequence of colorectal cancer (197). There is therefore no evidence that mutations in or altered regulation of the Hedgehog pathway can initiate gastrointestinal carcinogenesis. As discussed below, there is important evidence that Hedgehog signaling may play a role in the maintenance of tumor cell viability at later stages of tumorigenesis. B. Hedgehog Signaling in Cancers of the Proximal Gastrointestinal Tract In an important study by Berman et al. (8), it was shown that cancer cell lines derived from the esophagus, stomach, pancreas, and biliary tract showed high auton- 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 V. HEDGEHOG SIGNALING AND GASTROINTESTINAL CARCINOGENESIS 1367 1368 GIJS R. VAN DEN BRINK Physiol Rev • VOL C. Hedgehog Signaling in Colorectal Cancer As discussed above, we have shown that Ihh signaling regulates differentiation of the absorptive enterocyte lineage in the colon of the rat. We further found that IHH expression is lost very early in the process of colorectal carcinogenesis in humans (197). We have studied neoplastic lesions in patients with familial adenomatous polyposis (FAP) who carry a germline mutation in the adenomatous polyposis coli (APC) gene (50). These patients develop hundreds to thousands of polyps in adolescence upon loss of expression of the second APC allele in intestinal epithelial precursor cells. Loss of APC expression results in uncontrolled activity of the Wnt signaling pathway, which results in clonal precursor cell accumulation (55). Although FAP may be a rare syndrome, it is believed that up to 90% of colorectal cancers are similarly caused by mutations in APC or other mutations that result in uncontrolled Wnt pathway activity (55). We found that IHH expression was lost from polyps in patients with FAP and in flat lesions that precede the polyp stage. These data indicated that loss of IHH expression may be associated with one of the first steps in colorectal carcinogenesis and a direct result from the overactivation of the Wnt pathway. Similar to the results in APC mutant polyps, we were able to show that IHH expression is very low in DLD-1 colorectal cancer cells which carry a mutation in the APC gene. Expression of IHH could be restored in DLD-1 cells by blocking Wnt pathway activation (197). These results indicated that loss of IHH expression in APC mutant polyps indeed resulted directly from Wnt pathway activation. There is little evidence for activation of Hedgehog signaling at later stages of colorectal carcinogenesis. In the study by Berman et al. (8), it was found that in contrast to proximal gastrointestinal cancer cells, colon cancer cells did not depend on Hedgehog signaling for their survival. The authors showed that this lack of sensitivity of colorectal cancer cells to cyclopamine correlated with the absence of active Hedgehog signaling in these cells. This corroborates our findings that Hedgehog signaling is low to absent in undifferentiated colorectal cancer cells. We similarly found that inhibition of Hedgehog signaling with cyclopamine in colorectal cancer cells did not affect their viability or proliferation (unpublished observations). In contrast, Qualtrough et al. (160) observed increased cell death in a variety of colonic epithelial cell lines treated with cyclopamine; these experiments included HT-29 cells that have also been used by us. We currently have no explanation for this difference, but the sensitivity of the viability of colorectal cancer cells to the effects of cyclopamine may depend on cell culture conditions, such as the level of confluence. Berman et al. (8) did not detect any PTC1 mRNA in colorectal cancer cells. However, GLI1 expression correlated with PTC1 expres- 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 omous Hedgehog pathway activity. This was supported by the demonstration that PTC1 mRNA expression was highly induced in gastric and pancreatic carcinomas compared with adjacent normal tissue, indicating increased pathway activity. It was shown that cell lines and cells derived from pancreas carcinoma xenografts that express high PTC1 levels depended on Hedgehog signaling for their survival in vitro. The overactivity of the Hedgehog pathway resulted from overexpression of Hedgehog ligand as it could be blocked by both the SMO antagonist cyclopamine and a Hedgehog blocking monoclonal antibody. The data correlated with a jointly published study by Thayer et al. (190) in which it was found that expression of SHH and PTC1 protein is low or absent from the normal pancreas, induced at advanced stage preneoplastic lesions, and highest in adenocarcinomas. Six out of 13 pancreatic cell lines were sensitive to treatment with cyclopamine in this study when grown in vitro or as shown for some as xenografts in nude mice. Importantly, some of the cell lines that were unresponsive to cyclopamine treatment did express high levels of Hedgehog target genes, suggesting that the pathway may be activated at a level downstream of SMO in these cells. It is important in the interpretation of all of the available data to realize that the role of Hedgehog signaling on gastrointestinal carcinoma cells has so far been studied in isolated populations and not in the context of the tumor stroma (the stroma in xenografts is unlikely to adequately reflect the stroma found in real cancers). As discussed above in section IVB, the effect of Hedgehog signaling on epithelial cells may depend on the presence of stromal cells. It is therefore now important to examine the role of Hedgehog signaling in these tumors in their in vivo context for example in genetically modified mice. Samples of large patient cohorts should be examined for Hedgehog pathway activation, to further address the role of Hedgehog signaling in proximal gastrointestinal carcinomas. Little data are currently available to accurately assess the frequency of Hedgehog pathway activation in proximal gastrointestinal carcinomas. Ma et al. (120) examined 99 gastric adenocarcinomas and found evidence for Hedgehog activity (PTC1 expression detected by in situ hybridization) in ⬃65% of all gastric carcinomas except signet-ring cell carcinomas. The frequency of Hedgehog activation in tumors correlated with advanced stage and poor differentiation (120). In conclusion, important evidence indicates that Hedgehog signaling is active at later stages of tumorigenesis of the proximal gastrointestinal tract. Although this may often be associated with ligand overexpression, the data by Thayer et al. (190) suggest that activation at a level below Smo may also play a role. HEDGEHOG SIGNALING D. Therapeutic Targeting of the Hedgehog Pathway The spectacular effect of SMO inhibition on the viability of many gastrointestinal cancer cell lines suggests that the Hedgehog pathway may be an attractive target for cancer therapy. It is important to further examine in large numbers of cancers if Hedgehog pathway activity is mostly due to ligand overexpression (8) or if pathway Physiol Rev • VOL activation at a level downstream of Smo plays a role in some tumors as the data of Thayer et al. suggest (190). It seems preferable to try to target Hedgehog signaling as far downstream as possible. This allows treatment of cancers in which the pathway is activated at a level downstream of Smo and may help to minimize the chance of the development of drug resistance. The Gli transcription factors may therefore ultimately be the ideal target for therapy. This may be a difficult target for drug development, however, as this would probably require a drug that interferes with protein-DNA binding. Several compounds have now been described that inhibit Hedgehog signaling at the level of Smo (24, 47, 207). Hopefully one or more of these compounds may have favorable pharmacokinetic properties and a low toxicity profile that would allow the first trials in human cancer patients. VI. CONCLUSION The Hedgehog signaling pathway is a highly conserved signaling pathway that plays an important role in pattern formation in many, if not most, of the organs of the body. The pathway affects patterning of the gut along all of its axes. The first effect mediated by Hedgehog signaling that is of relevance for the developing gut is the regulation of left-right axis formation. Hedgehog signaling allows the establishment of this axis by establishing a midline barrier that is necessary to contain long-range acting signals to one side of the body to allow the induction of left-right specific gene expression. During the early formation of the gut tube, instructive signals from the mesenchyme establish specific domains of Hedgehog expression along the AP axis that are critical to the normal outgrowth of endodermal appendages, such as the airways and pancreas. At a later stage, Hedgehog signaling affects, for example, AP axis formation of the stomach by specifying the location of the forestomach-glandular stomach boundary. Hedgehog affects the radial axis by regulating the size of the smooth muscle layer of the gut, development of the intestinal nervous system, and villus formation in the small intestine. Analysis of the role of Hedgehog signaling in the adult has thus far been hampered by the embryonic lethality of most of the Hedgehog pathway mutant mice. More knowledge will hopefully soon be available by the generation of inducible mutant mice. From the analysis of the expression of Hedgehog pathway components, it seems that at least two domains exist in the adult gut with Hedgehog pathway activity: the epithelium of the proximal stomach and the distal colon. In the stomach, Hedgehog signaling seems to play a role in parietal cell physiology and possibly pit cell survival and/or differentiation. In the distal colon, Hedgehog signaling regulates colonic enterocyte differentiation. 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 sion in all cell lines examined except in the colorectal cancer cells. Correlation between PTC1 and GLI1 expression may be expected as both are transcriptional targets of Hedgehog signaling. Also, since SMO is expressed in colorectal cancer cells (160, 217), absence of PTC1 expression would be expected to activate the pathway and would not result in low to absent activity of Hedgehog signaling as observed by Berman et al. (8). Indeed, several other labs have shown that PTC1 mRNA is expressed in colorectal cancer cells. Qualtrough et al. (160) demonstrated PTC1 expression in two out of two colon adenoma cell lines and three out of three colorectal cancer cell lines. Zhu et al. (217) detected PTC1 mRNA in six out of eight colorectal cancer cell lines. Both Zhu et al. and Qualtrough et al. demonstrated PTC1 expression in HT-29 cells that were used in our experiments. Furthermore, PTC1 mRNA has been demonstrated by PCR in normal colonic epithelial cells on microdissected cells (145) and by in situ hybridization in normal human colon (141). As discussed above, we have recently performed in situ hybridizations for Ptc1 mRNA in the mouse colon, since questions arose about the specificity of the anti-Ptc1 antibodies we previously used. Although we found that Ptc1 mRNA is expressed in the epithelium of the anorectum, we did not detect any Ptc1 mRNA in the epithelium of the mouse colon where it was exclusively expressed in the mesenchyme underlying the differentiated enterocytes (unpublished observations). Expression of PTC1 in colorectal cancer cells may therefore reflect their loss of differentiation along the anterior-posterior axis of the gastrointestinal tract (colorectal cancer cells are, for example, known to express several markers of gastric epithelium). We will reassess PTC1 mRNA expression in the normal colonic epithelium and in colorectal cancers in humans. It remains questionable, however, if PTC expression and Hedgehog responsiveness in colorectal cancer cell lines is of any relevance if Hedgehog signaling is mainly towards the mesenchyme in vivo. Clearly, additional experiments are required to resolve this issue, and these are currently underway in our laboratory. In conclusion, the relevance of the loss of Hedgehog signaling in colorectal carcinogenesis is not yet clear. It may be involved in the loss of epithelial differentiation that is a hallmark of adenomatous polyps. 1369 1370 GIJS R. VAN DEN BRINK ACKNOWLEDGMENTS Address for reprint requests and other correspondence: G. R. van den Brink, Dept. of Gastroenterology & Hepatology, Leiden University Medical Center, Building 1, C4-P012, PO Box 9600, 2300 RC Leiden, The Netherlands (e-mail: [email protected]). GRANTS This study was supported by a VENI grant from the Netherlands Organization for Scientific Research. REFERENCES 1. Afouda BA, Ciau-Uitz A, Patient R. GATA4, 5 and 6 mediate TGFbeta maintenance of endodermal gene expression in Xenopus embryos. Development 132: 763–774, 2005. 2. Alt B, Elsalini OA, Schrumpf P, Haufs N, Lawson ND, Schwabe GC, Mundlos S, Gruters A, Krude H, Rohr KB. Arteries define the position of the thyroid gland during its developmental relocalisation. Development 133: 3797–3804, 2006. 3. Andrassy RJ, Mahour GH. Gastrointestinal anomalies associated with esophageal atresia or tracheoesophageal fistula. Arch Surg 114: 1125–1128, 1979. 4. Apelqvist A, Ahlgren U, Edlund H. Sonic hedgehog directs specialised mesoderm differentiation in the intestine and pancreas. Curr Biol 7: 801– 804, 1997. Physiol Rev • VOL 5. Augeron C, Laboisse CL. Emergence of permanently differentiated cell clones in a human colonic cancer cell line in culture after treatment with sodium butyrate. Cancer Res 44: 3961–3969, 1984. 6. Batts LE, Polk DB, Dubois RN, Kulessa H. Bmp signaling is required for intestinal growth and morphogenesis. Dev Dyn 235: 1563–1570, 2006. 7. Beck F, Erler T, Russell A, James R. Expression of Cdx-2 in the mouse embryo and placenta: possible role in patterning of the extra-embryonic membranes. Dev Dyn 204: 219 –227, 1995. 8. Berman DM, Karhadkar SS, Maitra A, Montes De Oca R, Gerstenblith MR, Briggs K, Parker AR, Shimada Y, Eshleman JR, Watkins DN, Beachy PA. Widespread requirement for Hedgehog ligand stimulation in growth of digestive tract tumours. Nature 425: 846 – 851, 2003. 9. Bhatia N, Thiyagarajan S, Elcheva I, Saleem M, Dlugosz A, Mukhtar H, Spiegelman VS. Gli2 is targeted for ubiquitination and degradation by beta-TrCP ubiquitin ligase. J Biol Chem 281: 19320 –19326, 2006. 10. Bienz M, Clevers H. Linking colorectal cancer to Wnt signaling. Cell 103: 311–320, 2000. 11. Bijlsma MF, Peppelenbosch MP, Spek CA. Hedgehog morphogen in cardiovascular disease. Circulation 114: 1985–1991, 2006. 12. Bijlsma MF, Spek CA, Zivkovic D, van de Water S, Rezaee F, Peppelenbosch MP. Repression of smoothened by patched-dependent (Pro-) vitamin D3 secretion. PLoS Biol 4, 2006. 13. Bitgood MJ, McMahon AP. Hedgehog and Bmp genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev Biol 172: 126 –138, 1995. 14. Briscoe J, Chen Y, Jessell TM, Struhl G. A hedgehog-insensitive form of patched provides evidence for direct long-range morphogen activity of sonic hedgehog in the neural tube. Mol Cell 7: 1279 –1291, 2001. 15. Bry L, Falk P, Huttner K, Ouellette A, Midtvedt T, Gordon JI. Paneth cell differentiation in the developing intestine of normal and transgenic mice. Proc Natl Acad Sci USA 91: 10335–10339, 1994. 16. Bumcrot DA, Takada R, McMahon AP. Proteolytic processing yields two secreted forms of sonic hedgehog. Mol Cell Biol 15: 2294 –2303, 1995. 17. Calvert R, Pothier P. Migration of fetal intestinal intervillous cells in neonatal mice. Anat Rec 227: 199 –206, 1990. 18. Capdevila J, Vogan KJ, Tabin CJ, Izpisua Belmonte JC. Mechanisms of left-right determination in vertebrates. Cell 101: 9 –21, 2000. 19. Cardoso WV, Lu J. Regulation of early lung morphogenesis: questions, facts and controversies. Development 133: 1611–1624, 2006. 20. Caspary T, Garcia-Garcia MJ, Huangfu D, Eggenschwiler JT, Wyler MR, Rakeman AS, Alcorn HL, Anderson KV. Mouse Dispatched homolog1 is required for long-range, but not juxtacrine, Hh signaling. Curr Biol 12: 1628 –1632, 2002. 21. Celli J, van Bokhoven H, Brunner HG. Feingold syndrome: clinical review and genetic mapping. Am J Med Genet A 122: 294 –300, 2003. 22. Chang WW, Leblond CP. Renewal of the epithelium in the descending colon of the mouse. I. Presence of three cell populations: vacuolated-columnar, mucous and argentaffin. Am J Anat 131: 73–99, 1971. 23. Chen JK, Taipale J, Cooper MK, Beachy PA. Inhibition of Hedgehog signaling by direct binding of cyclopamine to Smoothened. Genes Dev 16: 2743–2748, 2002. 24. Chen JK, Taipale J, Young KE, Maiti T, Beachy PA. Small molecule modulation of Smoothened activity. Proc Natl Acad Sci USA 99: 14071–14076, 2002. 25. Chen MH, Li YJ, Kawakami T, Xu SM, Chuang PT. Palmitoylation is required for the production of a soluble multimeric Hedgehog protein complex and long-range signaling in vertebrates. Genes Dev 18: 641– 659, 2004. 26. Chuang PT, McMahon AP. Vertebrate Hedgehog signalling modulated by induction of a Hedgehog-binding protein. Nature 397: 617– 621, 1999. 27. Colony PC, Conforti JC. Morphogenesis in the fetal rat proximal colon: effects of cytochalasin D. Anat Rec 235: 241–252, 1993. 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 Hedgehog signaling is initially lost in preneoplastic lesions of the stomach and colon. It seems, however, that at later stages of carcinogenesis there is a gain of Hedgehog signaling that maintains viability of carcinoma cells in vitro and when grown as xenografts in nude mice. This gain of Hedgehog signaling may be both at the level of Hedgehog ligand overexpression and activation downstream of Smo, which has important implications for possible therapeutic strategies. Many important questions remain, especially for our understanding of the role of Hedgehog signaling in the adult gut. What exactly is the role of Hedgehog signaling in parietal cell physiology in vivo? Hedgehog signaling is lost early in the process of neoplastic change of the distal colon. Is this an epiphenomenon or does it play a causal role in loss of cellular differentiation? Is Hedgehog signaling in the colon exclusively from the epithelium to the mesenchyme or also partially autocrine? Hedgehog signaling seems to antagonize Wnt signaling in the distal colon; does this mean that Hedgehog signaling can act as a tumor suppressor in the colon? Hedgehog signaling seems essential to maintain the viability of cell lines derived from cancers of the proximal gastrointestinal tract in vitro and when grown as xenografts. Is this reflective of effects on these tumor cells when they are in their natural environment of tumor stroma which is known to play a critical role in carcinoma behavior? Will it be possible to treat patients with systemic Hedgehog antagonists without major side effects? In the coming years many of these questions will be addressed in experiments in animal models and hopefully also in the first clinical trials in human cancer patients. HEDGEHOG SIGNALING Physiol Rev • VOL 50. 51. 52. 53. 54. 55. 56. 57. 58. 59. 60. 61. 62. 63. 64. 65. 66. 67. 68. 69. 70. moda T, Nimura Y, Yoshida T, Sasaki H. Hedgehog signal activation in gastric pit cell and in diffuse-type gastric cancer. Gastroenterology 131: 14 –29, 2006. Galiatsatos P, Foulkes WD. Familial adenomatous polyposis. Am J Gastroenterol 101: 385–398, 2006. Goodrich LV, Johnson RL, Milenkovic L, McMahon JA, Scott MP. Conservation of the hedgehog/patched signaling pathway from flies to mice: induction of a mouse patched gene by Hedgehog. Genes Dev 10: 301–312, 1996. Gordon JI, Hermiston ML. Differentiation and self-renewal in the mouse gastrointestinal epithelium. Curr Opin Cell Biol 6: 795– 803, 1994. Grapin-Botton A. Antero-posterior patterning of the vertebrate digestive tract: 40 years after Nicole Le Douarin’s PhD thesis. Int J Dev Biol 49: 335–347, 2005. Greaves LC, Preston SL, Tadrous PJ, Taylor RW, Barron MJ, Oukrif D, Leedham SJ, Deheragoda M, Sasieni P, Novelli MR, Jankowski JA, Turnbull DM, Wright NA, McDonald SA. Mitochondrial DNA mutations are established in human colonic stem cells, and mutated clones expand by crypt fission. Proc Natl Acad Sci USA 103: 714 –719, 2006. Gregorieff A, Clevers H. Wnt signaling in the intestinal epithelium: from endoderm to cancer. Genes Dev 19: 877– 890, 2005. Gualdi R, Bossard P, Zheng M, Hamada Y, Coleman JR, Zaret KS. Hepatic specification of the gut endoderm in vitro: cell signaling and transcriptional control. Genes Dev 10: 1670 –1682, 1996. Hahn H, Wicking C, Zaphiropoulous PG, Gailani MR, Shanley S, Chidambaram A, Vorechovsky I, Holmberg E, Unden AB, Gillies S, Negus K, Smyth I, Pressman C, Leffell DJ, Gerrard B, Goldstein AM, Dean M, Toftgard R, Chenevix-Trench G, Wainwright B, Bale AE. Mutations of the human homolog of Drosophila patched in the nevoid basal cell carcinoma syndrome. Cell 85: 841– 851, 1996. Haramis AP, Begthel H, van den Born M, van Es J, Jonkheer S, Offerhaus GJ, Clevers H. De novo crypt formation and juvenile polyposis on BMP inhibition in mouse intestine. Science 303: 1684 –1686, 2004. Haycraft CJ, Banizs B, Aydin-Son Y, Zhang Q, Michaud EJ, Yoder BK. Gli2 and Gli3 localize to cilia and require the intraflagellar transport protein polaris for processing and function. PLoS Genet 1: e53, 2005. He XC, Zhang J, Tong WG, Tawfik O, Ross J, Scoville DH, Tian Q, Zeng X, He X, Wiedemann LM, Mishina Y, Li L. BMP signaling inhibits intestinal stem cell self-renewal through suppression of Wnt-beta-catenin signaling. Nat Genet 36: 1117–1121, 2004. Hebrok M, Kim SK, Melton DA. Notochord repression of endodermal Sonic hedgehog permits pancreas development. Genes Dev 12: 1705–1713, 1998. Hebrok M, Kim SK, St Jacques B, McMahon AP, Melton DA. Regulation of pancreas development by hedgehog signaling. Development 127: 4905– 4913, 2000. Hermiston ML, Wong MH, Gordon JI. Forced expression of E-cadherin in the mouse intestinal epithelium slows cell migration and provides evidence for nonautonomous regulation of cell fate in a self-renewing system. Genes Dev 10: 985–996, 1996. Hezel AF, Kimmelman AC, Stanger BZ, Bardeesy N, Depinho RA. Genetics and biology of pancreatic ductal adenocarcinoma. Genes Dev 20: 1218 –1249, 2006. Hirokawa N, Tanaka Y, Okada Y, Takeda S. Nodal flow and the generation of left-right asymmetry. Cell 125: 33– 45, 2006. Hogan BL. Morphogenesis. Cell 96: 225–233, 1999. Hooper JE, Scott MP. Communicating with Hedgehogs. Nat Rev Mol Cell Biol 6: 306 –317, 2005. Horne-Badovinac S, Rebagliati M, Stainier DY. A cellular framework for gut-looping morphogenesis in zebrafish. Science 302: 662– 665, 2003. Howe JR, Bair JL, Sayed MG, Anderson ME, Mitros FA, Petersen GM, Velculescu VE, Traverso G, Vogelstein B. Germline mutations of the gene encoding bone morphogenetic protein receptor 1A in juvenile polyposis. Nat Genet 28: 184 –187, 2001. Hruban RH, Adsay NV, Albores-Saavedra J, Anver MR, Biankin AV, Boivin GP, Furth EE, Furukawa T, Klein A, Klimstra DS, Kloppel G, Lauwers GY, Longnecker DS, Luttges 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 28. Cooper AF, Yu KP, Brueckner M, Brailey LL, Johnson L, McGrath JM, Bale AE. Cardiac and CNS defects in a mouse with targeted disruption of suppressor of fused. Development 132: 4407– 4417, 2005. 29. Cooper MK, Porter JA, Young KE, Beachy PA. Teratogenmediated inhibition of target tissue response to Shh signaling. Science 280: 1603–1607, 1998. 30. Corbit KC, Aanstad P, Singla V, Norman AR, Stainier DY, Reiter JF. Vertebrate Smoothened functions at the primary cilium. Nature 437: 1018 –1021, 2005. 31. De Felice M, Di Lauro R. Thyroid development and its disorders: genetics and molecular mechanisms. Endocr Rev 25: 722–746, 2004. 32. De Santa Barbara P, Roberts DJ. Tail gut endoderm and gut/ genitourinary/tail development: a new tissue-specific role for Hoxa13. Development 129: 551–561, 2002. 33. Denef N, Neubuser D, Perez L, Cohen SM. Hedgehog induces opposite changes in turnover and subcellular localization of patched and smoothened. Cell 102: 521–531, 2000. 34. Deutsch G, Jung J, Zheng M, Lora J, Zaret KS. A bipotential precursor population for pancreas and liver within the embryonic endoderm. Development 128: 871– 881, 2001. 35. diIorio PJ, Moss JB, Sbrogna JL, Karlstrom RO, Moss LG. Sonic hedgehog is required early in pancreatic islet development. Dev Biol 244: 75– 84, 2002. 36. Dimmler A, Brabletz T, Hlubek F, Hafner M, Rau T, Kirchner T, Faller G. Transcription of sonic hedgehog, a potential factor for gastric morphogenesis and gastric mucosa maintenance, is upregulated in acidic conditions. Lab Invest 83: 1829 –1837, 2003. 37. Duan H, Gao F, Li S, Nagata T. Postnatal development and aging of esophageal epithelium in mouse: a light and electron microscopic radioautographic study. Cell Mol Biol 39: 309 –316, 1993. 38. Duluc I, Freund JN, Leberquier C, Kedinger M. Fetal endoderm primarily holds the temporal and positional information required for mammalian intestinal development. J Cell Biol 126: 211–221, 1994. 39. Dwyer JR, Sever N, Carlson M, Nelson SF, Beachy PA, Parhami F. Oxysterols are novel activators of the hedgehog signaling pathway in pluripotent mesenchymal cells. J Biol Chem. In press. 40. Echelard Y, Epstein DJ, St-Jacques B, Shen L, Mohler J, McMahon JA, McMahon AP. Sonic hedgehog, a member of a family of putative signaling molecules, is implicated in the regulation of CNS polarity. Cell 75: 1417–1430, 1993. 41. Ehlen HW, Buelens LA, Vortkamp A. Hedgehog signaling in skeletal development. Birth Defects Res C Embryo Today 78: 267– 279, 2006. 42. Ericson J, Rashbass P, Schedl A, Brenner-Morton S, Kawakami A, van Heyningen V, Jessell TM, Briscoe J. Pax6 controls progenitor cell identity and neuronal fate in response to graded Shh signaling. Cell 90: 169 –180, 1997. 43. Fagman H, Andersson L, Nilsson M. The developing mouse thyroid: embryonic vessel contacts and parenchymal growth pattern during specification, budding, migration, and lobulation. Dev Dyn 235: 444 – 455, 2006. 44. Fagman H, Grande M, Gritli-Linde A, Nilsson M. Genetic deletion of sonic hedgehog causes hemiagenesis and ectopic development of the thyroid in mouse. Am J Pathol 164: 1865–1872, 2004. 45. Falk P, Roth KA, Gordon JI. Lectins are sensitive tools for defining the differentiation programs of mouse gut epithelial cell lineages. Am J Physiol Gastrointest Liver Physiol 266: G987– G1003, 1994. 46. Farrington SM, Belaoussoff M, Baron MH. Winged-helix, Hedgehog and Bmp genes are differentially expressed in distinct cell layers of the murine yolk sac. Mech Dev 62: 197–211, 1997. 47. Frank-Kamenetsky M, Zhang XM, Bottega S, Guicherit O, Wichterle H, Dudek H, Bumcrot D, Wang FY, Jones S, Shulok J, Rubin LL, Porter JA. Small-molecule modulators of Hedgehog signaling: identification and characterization of Smoothened agonists and antagonists. J Biol 1: 10, 2002. 48. Fukamachi H, Mizuno T, Takayama S. Epithelial-mesenchymal interactions in differentiation of stomach epithelium in fetal mice. Anat Embryol 157: 151–160, 1979. 49. Fukaya M, Isohata N, Ohta H, Aoyagi K, Ochiya T, Saeki N, Yanagihara K, Nakanishi Y, Taniguchi H, Sakamoto H, Shi- 1371 1372 71. 72. 73. 74. 76. 77. 78. 79. 80. 81. 82. 83. 84. 85. 86. 87. 88. 89. 90. 91. J, Maitra A, Offerhaus GJ, Perez-Gallego L, Redston M, Tuveson DA. Pathology of genetically engineered mouse models of pancreatic exocrine cancer: consensus report and recommendations. Cancer Res 66: 95–106, 2006. Huangfu D, Anderson KV. Signaling from Smo to Ci/Gli: conservation and divergence of Hedgehog pathways from Drosophila to vertebrates. Development 133: 3–14, 2006. Huangfu D, Liu A, Rakeman AS, Murcia NS, Niswander L, Anderson KV. Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature 426: 83– 87, 2003. Hui CC, Joyner AL. A mouse model of greig cephalopolysyndactyly syndrome: the extra-toesJ mutation contains an intragenic deletion of the Gli3 gene. Nat Genet 3: 241–246, 1993. Hui CC, Slusarski D, Platt KA, Holmgren R, Joyner AL. Expression of three mouse homologs of the Drosophila segment polarity gene cubitus interruptus, Gli, Gli-2, Gli-3, in ectoderm- and mesoderm-derived tissues suggests multiple roles during postimplantation development. Dev Biol 162: 402– 413, 1994. Incardona JP, Gaffield W, Kapur RP, Roelink H. The teratogenic Veratrum alkaloid cyclopamine inhibits sonic hedgehog signal transduction. Development 125: 3553–3562, 1998. Incardona JP, Gruenberg J, Roelink H. Sonic hedgehog induces the segregation of patched and smoothened in endosomes. Curr Biol 12: 983–995, 2002. Ingham PW, Placzek M. Orchestrating ontogenesis: variations on a theme by sonic hedgehog. Nat Rev Genet 7: 841– 850, 2006. Jacobsen CM, Narita N, Bielinska M, Syder AJ, Gordon JI, Wilson DB. Genetic mosaic analysis reveals that GATA-4 is required for proper differentiation of mouse gastric epithelium. Dev Biol 241: 34 – 46, 2002. Johnson RL, Rothman AL, Xie J, Goodrich LV, Bare JW, Bonifas JM, Quinn AG, Myers RM, Cox DR, Epstein EH Jr, Scott MP. Human homolog of patched, a candidate gene for the basal cell nevus syndrome. Science 272: 1668 –1671, 1996. Jones RG, Li X, Gray PD, Kuang J, Clayton F, Samowitz WS, Madison BB, Gumucio DL, Kuwada SK. Conditional deletion of 1 integrins in the intestinal epithelium causes a loss of Hedgehog expression, intestinal hyperplasia, early postnatal lethality. J Cell Biol 175: 505–514, 2006. Jung J, Zheng M, Goldfarb M, Zaret KS. Initiation of mammalian liver development from endoderm by fibroblast growth factors. Science 284: 1998 –2003, 1999. Kaestner KH. The making of the liver: developmental competence in foregut endoderm and induction of the hepatogenic program. Cell Cycle 4: 1146 –1148, 2005. Kalderon D. The mechanism of hedgehog signal transduction. Biochem Soc Trans 33: 1509 –1512, 2005. Kang JS, Mulieri PJ, Hu Y, Taliana L, Krauss RS. BOC, an Ig superfamily member, associates with CDO to positively regulate myogenic differentiation. EMBO J 21: 114 –124, 2002. Kang JS, Mulieri PJ, Miller C, Sassoon DA, Krauss RS. CDO, a robo-related cell surface protein that mediates myogenic differentiation. J Cell Biol 143: 403– 413, 1998. Kang S, Graham JM Jr, Olney AH, Biesecker LG. GLI3 frameshift mutations cause autosomal dominant Pallister-Hall syndrome. Nat Genet 15: 266 –268, 1997. Karam SM, Leblond CP. Dynamics of epithelial cells in the corpus of the mouse stomach. II. Outward migration of pit cells. Anat Rec 236: 280 –296, 1993. Karam SM, Leblond CP. Identifying and counting epithelial cell types in the “corpus” of the mouse stomach. Anat Rec 232: 231–246, 1992. Karam SM, Li Q, Gordon JI. Gastric epithelial morphogenesis in normal and transgenic mice. Am J Physiol Gastrointest Liver Physiol 272: G1209 –G1220, 1997. Karlsson L, Lindahl P, Heath JK, Betsholtz C. Abnormal gastrointestinal development in PDGF-A and PDGFR-␣ deficient mice implicates a novel mesenchymal structure with putative instructive properties in villus morphogenesis. Development 127: 3457–3466, 2000. Kaufman M. The Atlas of Mouse Development. San Diego, CA: Academic, 1992. Physiol Rev • VOL 92. Kawahira H, Ma NH, Tzanakakis ES, McMahon AP, Chuang PT, Hebrok M. Combined activities of hedgehog signaling inhibitors regulate pancreas development. Development 130: 4871– 4879, 2003. 93. Kawahira H, Scheel DW, Smith SB, German MS, Hebrok M. Hedgehog signaling regulates expansion of pancreatic epithelial cells. Dev Biol 280: 111–121, 2005. 94. Kawakami T, Kawcak T, Li YJ, Zhang W, Hu Y, Chuang PT. Mouse dispatched mutants fail to distribute hedgehog proteins and are defective in hedgehog signaling. Development 129: 5753–5765, 2002. 95. Kedinger M, Simon-Assmann P, Bouziges F, Arnold C, Alexandre E, Haffen K. Smooth muscle actin expression during rat gut development and induction in fetal skin fibroblastic cells associated with intestinal embryonic epithelium. Differentiation 43: 87–97, 1990. 96. Kenney AM, Widlund HR, Rowitch DH. Hedgehog and PI-3 kinase signaling converge on Nmyc1 to promote cell cycle progression in cerebellar neuronal precursors. Development 131: 217–228, 2004. 97. Khoury MJ, Cordero JF, Greenberg F, James LM, Erickson JD. A population study of the VACTERL association: evidence for its etiologic heterogeneity. Pediatrics 71: 815– 820, 1983. 98. Kim J, Kim P, Hui CC. The VACTERL association: lessons from the Sonic hedgehog pathway. Clin Genet 59: 306 –315, 2001. 99. Kim JH, Huang Z, Mo R. Gli3 null mice display glandular overgrowth of the developing stomach. Dev Dyn 234: 984 –991, 2005. 100. Kim SK, Hebrok M. Intercellular signals regulating pancreas development and function. Genes Dev 15: 111–127, 2001. 101. Kim SK, Hebrok M, Li E, Oh SP, Schrewe H, Harmon EB, Lee JS, Melton DA. Activin receptor patterning of foregut organogenesis. Genes Dev 14: 1866 –1871, 2000. 102. Kim SK, Hebrok M, Melton DA. Notochord to endoderm signaling is required for pancreas development. Development 124: 4243– 4252, 1997. 103. Kim SK, Melton DA. Pancreas development is promoted by cyclopamine, a hedgehog signaling inhibitor. Proc Natl Acad Sci USA 95: 13036 –13041, 1998. 104. Kioussi C, O’Connell S, St-Onge L, Treier M, Gleiberman AS, Gruss P, Rosenfeld MG. Pax6 is essential for establishing ventraldorsal cell boundaries in pituitary gland development. Proc Natl Acad Sci USA 96: 14378 –14382, 1999. 105. Koch A, Waha A, Hartmann W, Milde U, Goodyer CG, Sorensen N, Berthold F, Digon-Sontgerath B, Kratzschmar J, Wiestler OD, Pietsch T. No evidence for mutations or altered expression of the Suppressor of Fused gene (SUFU) in primitive neuroectodermal tumours. Neuropathol Appl Neurobiol 30: 532– 539, 2004. 106. Lammert E, Cleaver O, Melton D. Induction of pancreatic differentiation by signals from blood vessels. Science 294: 564 –567, 2001. 107. Lammert E, Cleaver O, Melton D. Role of endothelial cells in early pancreas and liver development. Mech Dev 120: 59 – 64, 2003. 108. Lawson KA, Pedersen RA. Cell fate, morphogenetic movement and population kinetics of embryonic endoderm at the time of germ layer formation in the mouse. Development 101: 627– 652, 1987. 109. Lee ER. Dynamic histology of the antral epithelium in the mouse stomach. III. Ultrastructure and renewal of pit cells. Am J Anat 172: 225–240, 1985. 110. Lee ER, Leblond CP. Dynamic histology of the antral epithelium in the mouse stomach. IV. Ultrastructure and renewal of gland cells. Am J Anat 172: 241–259, 1985. 111. Lee JJ, Ekker SC, von Kessler DP, Porter JA, Sun BI, Beachy PA. Autoproteolysis in hedgehog protein biogenesis. Science 266: 1528 –1537, 1994. 112. Lee JJ, von Kessler DP, Parks S, Beachy PA. Secretion and localized transcription suggest a role in positional signaling for products of the segmentation gene hedgehog. Cell 71: 33–50, 1992. 113. Lewis PM, Dunn MP, McMahon JA, Logan M, Martin JF, St-Jacques B, McMahon AP. Cholesterol modification of sonic hedgehog is required for long-range signaling activity and effective modulation of signaling by Ptc1. Cell 105: 599 – 612, 2001. 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 75. GIJS R. VAN DEN BRINK HEDGEHOG SIGNALING Physiol Rev • VOL 135. Mikels AJ, Nusse R. Purified Wnt5a protein activates or inhibits beta-catenin-TCF signaling depending on receptor context. PLoS Biol 4: e115, 2006. 136. Mo R, Kim JH, Zhang J, Chiang C, Hui CC, Kim PC. Anorectal malformations caused by defects in sonic hedgehog signaling. Am J Pathol 159: 765–774, 2001. 137. Mohler J, Vani K. Molecular organization and embryonic expression of the hedgehog gene involved in cell-cell communication in segmental patterning of Drosophila. Development 115: 957–971, 1992. 138. Motoyama J, Liu J, Mo R, Ding Q, Post M, Hui CC. Essential function of Gli2 and Gli3 in the formation of lung, trachea and oesophagus. Nat Genet 20: 54 –57, 1998. 139. Motoyama J, Takabatake T, Takeshima K, Hui C. Ptch2, a second mouse Patched gene is co-expressed with Sonic hedgehog. Nat Genet 18: 104 –106, 1998. 140. Ng D, Stavrou T, Liu L, Taylor MD, Gold B, Dean M, Kelley MJ, Dubovsky EC, Vezina G, Nicholson HS, Byrne J, Rutka JT, Hogg D, Reaman GH, Goldstein AM. Retrospective family study of childhood medulloblastoma. Am J Med Genet A 134: 399 – 403, 2005. 141. Nielsen CM, Williams J, van den Brink GR, Lauwers GY, Roberts DJ. Hh pathway expression in human gut tissues and in inflammatory gut diseases. Lab Invest 84: 1631–1642, 2004. 142. Nonaka S, Tanaka Y, Okada Y, Takeda S, Harada A, Kanai Y, Kido M, Hirokawa N. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 95: 829 – 837, 1998. 143. Nusslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila. Nature 287: 795– 801, 1980. 144. Okada Y, Takeda S, Tanaka Y, Belmonte JC, Hirokawa N. Mechanism of nodal flow: a conserved symmetry breaking event in left-right axis determination. Cell 121: 633– 644, 2005. 145. Oniscu A, James RM, Morris RG, Bader S, Malcomson RD, Harrison DJ. Expression of Sonic hedgehog pathway genes is altered in colonic neoplasia. J Pathol 203: 909 –917, 2004. 146. Ormestad M, Astorga J, Landgren H, Wang T, Johansson BR, Miura N, Carlsson P. Foxf1 and Foxf2 control murine gut development by limiting mesenchymal Wnt signaling and promoting extracellular matrix production. Development 133: 833– 843, 2006. 147. Osterlund T, Kogerman P. Hedgehog signalling: how to get from Smo to Ci and Gli. Trends Cell Biol 16: 176 –180, 2006. 148. Pan Y, Bai CB, Joyner AL, Wang B. Sonic hedgehog signaling regulates Gli2 transcriptional activity by suppressing its processing and degradation. Mol Cell Biol 26: 3365–3377, 2006. 149. Panakova D, Sprong H, Marois E, Thiele C, Eaton S. Lipoprotein particles are required for Hedgehog and Wingless signalling. Nature 435: 58 – 65, 2005. 150. Park HL, Bai C, Platt KA, Matise MP, Beeghly A, Hui CC, Nakashima M, Joyner AL. Mouse Gli1 mutants are viable but have defects in SHH signaling in combination with a Gli2 mutation. Development 127: 1593–1605, 2000. 151. Park YK, Franklin JL, Settle SH, Levy SE, Chung E, Jeyakumar LH, Shyr Y, Washington MK, Whitehead RH, Aronow BJ, Coffey RJ. Gene expression profile analysis of mouse colon embryonic development. Genesis 41: 1–12, 2005. 152. Pasca di Magliano M, Sekine S, Ermilov A, Ferris J, Dlugosz AA, Hebrok M. Hedgehog/Ras interactions regulate early stages of pancreatic cancer. Genes Dev 20: 3161–3173, 2006. 153. Pepicelli CV, Lewis PM, McMahon AP. Sonic hedgehog regulates branching morphogenesis in the mammalian lung. Curr Biol 8: 1083–1086, 1998. 154. Perron M, Boy S, Amato MA, Viczian A, Koebernick K, Pieler T, Harris WA. A novel function for Hedgehog signalling in retinal pigment epithelium differentiation. Development 130: 1565–1577, 2003. 155. Pietsch T, Waha A, Koch A, Kraus J, Albrecht S, Tonn J, Sorensen N, Berthold F, Henk B, Schmandt N, Wolf HK, von Deimling A, Wainwright B, Chenevix-Trench G, Wiestler OD, Wicking C. Medulloblastomas of the desmoplastic variant carry mutations of the human homologue of Drosophila patched. Cancer Res 57: 2085–2088, 1997. 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 114. Li H, Arber S, Jessell TM, Edlund H. Selective agenesis of the dorsal pancreas in mice lacking homeobox gene Hlxb9. Nat Genet 23: 67–70, 1999. 115. Li Y, Zhang H, Litingtung Y, Chiang C. Cholesterol modification restricts the spread of Shh gradient in the limb bud. Proc Natl Acad Sci USA 103: 6548 – 6553, 2006. 116. Litingtung Y, Chiang C. Specification of ventral neuron types is mediated by an antagonistic interaction between Shh and Gli3. Nat Neurosci 3: 979 –985, 2000. 117. Litingtung Y, Lei L, Westphal H, Chiang C. Sonic hedgehog is essential to foregut development. Nat Genet 20: 58 – 61, 1998. 118. Liu A, Wang B, Niswander LA. Mouse intraflagellar transport proteins regulate both the activator and repressor functions of Gli transcription factors. Development 132: 3103–3111, 2005. 119. Lum L, Beachy PA. The Hedgehog response network: sensors, switches, routers. Science 304: 1755–1759, 2004. 120. Ma X, Chen K, Huang S, Zhang X, Adegboyega PA, Evers BM, Zhang H, Xie J. Frequent activation of the hedgehog pathway in advanced gastric adenocarcinomas. Carcinogenesis 26: 1698 –1705, 2005. 121. Ma Y, Erkner A, Gong R, Yao S, Taipale J, Basler K, Beachy PA. Hedgehog-mediated patterning of the mammalian embryo requires transporter-like function of dispatched. Cell 111: 63–75, 2002. 122. Madison BB, Braunstein K, Kuizon E, Portman K, Qiao XT, Gumucio DL. Epithelial hedgehog signals pattern the intestinal crypt-villus axis. Development 132: 279 –289, 2005. 123. Madison BB, Dunbar L, Qiao XT, Braunstein K, Braunstein E, Gumucio DL. Cis elements of the villin gene control expression in restricted domains of the vertical (crypt) and horizontal (duodenum, cecum) axes of the intestine. J Biol Chem 277: 33275–33283, 2002. 124. Mahlapuu M, Enerback S, Carlsson P. Haploinsufficiency of the forkhead gene Foxf1, a target for sonic hedgehog signaling, causes lung and foregut malformations. Development 128: 2397–2406, 2001. 125. Mahlapuu M, Ormestad M, Enerback S, Carlsson P. The forkhead transcription factor Foxf1 is required for differentiation of extra-embryonic and lateral plate mesoderm. Development 128: 155–166, 2001. 126. Manley NR. Thymus organogenesis and molecular mechanisms of thymic epithelial cell differentiation. Semin Immunol 12: 421– 428, 2000. 127. Mann RK, Beachy PA. Novel lipid modifications of secreted protein signals. Annu Rev Biochem 73: 891–923, 2004. 128. Mao J, Ligon KL, Rakhlin EY, Thayer SP, Bronson RT, Rowitch D, McMahon AP. A novel somatic mouse model to survey tumorigenic potential applied to the Hedgehog pathway. Cancer Res 66: 10171–10178, 2006. 129. Marcotullio LD, Ferretti E, Greco A, De Smaele E, Po A, Sico MA, Alimandi M, Giannini G, Maroder M, Screpanti I, Gulino A. Numb is a suppressor of Hedgehog signalling and targets Gli1 for Itch-dependent ubiquitination. Nat Cell Biol 8: 1415–1423, 2006. 130. Marigo V, Roberts DJ, Lee SM, Tsukurov O, Levi T, Gastier JM, Epstein DJ, Gilbert DJ, Copeland NG, Seidman CE, Jenkins NA, Seidman JG, McMahon AP, Tabin C. Cloning, expression, chromosomal location of SHH and IHH: two human homologues of the Drosophila segment polarity gene hedgehog. Genomics 28: 44 –51, 1995. 131. Martinez-Frias ML, Bermejo E, Rodriguez-Pinilla E. Anal atresia, vertebral, genital, urinary tract anomalies: a primary polytopic developmental field defect identified through an epidemiological analysis of associations. Am J Med Genet 95: 169 –173, 2000. 132. May SR, Ashique AM, Karlen M, Wang B, Shen Y, Zarbalis K, Reiter J, Ericson J, Peterson AS. Loss of the retrograde motor for IFT disrupts localization of Smo to cilia and prevents the expression of both activator and repressor functions of Gli. Dev Biol 287: 378 –389, 2005. 133. McGrath J, Somlo S, Makova S, Tian X, Brueckner M. Two populations of node monocilia initiate left-right asymmetry in the mouse. Cell 114: 61–73, 2003. 134. Menard D, Arsenault P. Maturation of human fetal esophagus maintained in organ culture. Anat Rec 217: 348 –354, 1987. 1373 1374 GIJS R. VAN DEN BRINK Physiol Rev • VOL 176. 177. 178. 179. 180. 181. 182. 183. 184. 185. 186. 187. 188. 189. 190. 191. 192. 193. 194. progressing to gastric cancer. Am J Gastroenterol 100: 581–587, 2005. Shook D, Keller R. Mechanisms, mechanics and function of epithelial-mesenchymal transitions in early development. Mech Dev 120: 1351–1383, 2003. Soriano L. Differentiation of epithelium of the digestive tube in vitro. J Embryol Exp Morphol 14: 119 –128, 1965. Spencer-Dene B, Sala FG, Bellusci S, Gschmeissner S, Stamp G, Dickson C. Stomach development is dependent on fibroblast growth factor 10/fibroblast growth factor receptor 2b-mediated signaling. Gastroenterology 130: 1233–1244, 2006. Stepan V, Ramamoorthy S, Nitsche H, Zavros Y, Merchant JL, Todisco A. Regulation and function of the sonic hedgehog signal transduction pathway in isolated gastric parietal cells. J Biol Chem 280: 15700 –15708, 2005. St-Onge L, Sosa-Pineda B, Chowdhury K, Mansouri A, Gruss P. Pax6 is required for differentiation of glucagon-producing alphacells in mouse pancreas. Nature 387: 406 – 409, 1997. Suzuki H, Minegishi Y, Nomoto Y, Ota T, Masaoka T, van den Brink GR, Hibi T. Down-regulation of a morphogen (sonic hedgehog) gradient in the gastric epithelium of Helicobacter pyloriinfected Mongolian gerbils. J Pathol 206: 186 –197, 2005. Svard J, Heby-Henricson K, Persson-Lek M, Rozell B, Lauth M, Bergstrom A, Ericson J, Toftgard R, Teglund S. Genetic elimination of Suppressor of fused reveals an essential repressor function in the mammalian Hedgehog signaling pathway. Dev Cell 10: 187–197, 2006. Tabata T, Eaton S, Kornberg TB. The Drosophila hedgehog gene is expressed specifically in posterior compartment cells and is a target of engrailed regulation. Genes Dev 6: 2635–2645, 1992. Taipale J, Cooper MK, Maiti T, Beachy PA. Patched acts catalytically to suppress the activity of Smoothened. Nature 418: 892– 897, 2002. Tanaka Y, Okada Y, Hirokawa N. FGF-induced vesicular release of Sonic hedgehog and retinoic acid in leftward nodal flow is critical for left-right determination. Nature 435: 172–177, 2005. Tao H, Shimizu M, Kusumoto R, Ono K, Noji S, Ohuchi H. A dual role of FGF10 in proliferation and coordinated migration of epithelial leading edge cells during mouse eyelid development. Development 132: 3217–3230, 2005. Taylor MD, Liu L, Raffel C, Hui CC, Mainprize TG, Zhang X, Agatep R, Chiappa S, Gao L, Lowrance A, Hao A, Goldstein AM, Stavrou T, Scherer SW, Dura WT, Wainwright B, Squire JA, Rutka JT, Hogg D. Mutations in SUFU predispose to medulloblastoma. Nat Genet 31: 306 –310, 2002. Tempe D, Casas M, Karaz S, Blanchet-Tournier MF, Concordet JP. Multisite protein kinase A and glycogen synthase kinase 3beta phosphorylation leads to Gli3 ubiquitination by SCFbetaTrCP. Mol Cell Biol 26: 4316 – 4326, 2006. Tenzen T, Allen BL, Cole F, Kang JS, Krauss RS, McMahon AP. The cell surface membrane proteins Cdo and Boc are components and targets of the Hedgehog signaling pathway and feedback network in mice. Dev Cell 10: 647– 656, 2006. Thayer SP, di Magliano MP, Heiser PW, Nielsen CM, Roberts DJ, Lauwers GY, Qi YP, Gysin S, Fernandez-del Castillo C, Yajnik V, Antoniu B, McMahon M, Warshaw AL, Hebrok M. Hedgehog is an early and late mediator of pancreatic cancer tumorigenesis. Nature 425: 851– 856, 2003. Thomas MK, Rastalsky N, Lee JH, Habener JF. Hedgehog signaling regulation of insulin production by pancreatic beta-cells. Diabetes 49: 2039 –2047, 2000. Thomas PQ, Brown A, Beddington RS. Hex: a homeobox gene revealing peri-implantation asymmetry in the mouse embryo and an early transient marker of endothelial cell precursors. Development 125: 85–94, 1998. Tsukui T, Capdevila J, Tamura K, Ruiz-Lozano P, RodriguezEsteban C, Yonei-Tamura S, Magallon J, Chandraratna RA, Chien K, Blumberg B, Evans RM, Belmonte JC. Multiple leftright asymmetry defects in Shh(⫺/⫺) mutant mice unveil a convergence of the shh and retinoic acid pathways in the control of Lefty-1. Proc Natl Acad Sci USA 96: 11376 –11381, 1999. Unden AB, Holmberg E, Lundh-Rozell B, Stahle-Backdahl M, Zaphiropoulos PG, Toftgard R, Vorechovsky I. Mutations in the 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 156. Porter JA, Ekker SC, Park WJ, von Kessler DP, Young KE, Chen CH, Ma Y, Woods AS, Cotter RJ, Koonin EV, Beachy PA. Hedgehog patterning activity: role of a lipophilic modification mediated by the carboxy-terminal autoprocessing domain. Cell 86: 21–34, 1996. 157. Porter JA, von Kessler DP, Ekker SC, Young KE, Lee JJ, Moses K, Beachy PA. The product of hedgehog autoproteolytic cleavage active in local and long-range signalling. Nature 374: 363–366, 1995. 158. Powell DW, Adegboyega PA, Di Mari JF, Mifflin RC. Epithelial cells and their neighbors. I. Role of intestinal myofibroblasts in development, repair, cancer. Am J Physiol Gastrointest Liver Physiol 289: G2–G7, 2005. 159. Preat T. Characterization of Suppressor of fused, a complete suppressor of the fused segment polarity gene of Drosophila melanogaster. Genetics 132: 725–736, 1992. 160. Qualtrough D, Buda A, Gaffield W, Williams AC, Paraskeva C. Hedgehog signalling in colorectal tumour cells: induction of apoptosis with cyclopamine treatment. Int J Cancer 110: 831– 837, 2004. 161. Quan L, Smith DW. The VATER association. Vertebral defects, Anal atresia, T-E fistula with esophageal atresia, Radial and Renal dysplasia: a spectrum of associated defects. J Pediatr 82: 104 –107, 1973. 162. Radhakrishna U, Bornholdt D, Scott HS, Patel UC, Rossier C, Engel H, Bottani A, Chandal D, Blouin JL, Solanki JV, Grzeschik KH, Antonarakis SE. The phenotypic spectrum of GLI3 morphopathies includes autosomal dominant preaxial polydactyly type-IV and postaxial polydactyly type-A/B; no phenotype prediction from the position of GLI3 mutations. Am J Hum Genet 65: 645– 655, 1999. 163. Raffel C, Jenkins RB, Frederick L, Hebrink D, Alderete B, Fults DW, James CD. Sporadic medulloblastomas contain PTCH mutations. Cancer Res 57: 842– 845, 1997. 164. Ramalho-Santos M, Melton DA, McMahon AP. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development 127: 2763–2772, 2000. 165. Raymond C, Anne V, Millane G. Development of esophageal epithelium in the fetal and neonatal mouse. Anat Rec 230: 225–234, 1991. 166. Rittler M, Paz JE, Castilla EE. VACTERL association, epidemiologic definition and delineation. Am J Med Genet 63: 529 –536, 1996. 167. Roberts DJ, Johnson RL, Burke AC, Nelson CE, Morgan BA, Tabin C. Sonic hedgehog is an endodermal signal inducing Bmp-4 and Hox genes during induction and regionalization of the chick hindgut. Development 121: 3163–3174, 1995. 168. Roberts DJ, Smith DM, Goff DJ, Tabin CJ. Epithelial-mesenchymal signaling during the regionalization of the chick gut. Development 125: 2791–2801, 1998. 169. Rossi JM, Dunn NR, Hogan BL, Zaret KS. Distinct mesodermal signals, including BMPs from the septum transversum mesenchyme, are required in combination for hepatogenesis from the endoderm. Genes Dev 15: 1998 –2009, 2001. 170. Roy S, Qiao T, Wolff C, Ingham PW. Hedgehog signaling pathway is essential for pancreas specification in the zebrafish embryo. Curr Biol 11: 1358 –1363, 2001. 171. Rubin DC, Swietlicki E, Roth KA, Gordon JI. Use of fetal intestinal isografts from normal and transgenic mice to study the programming of positional information along the duodenal-to-colonic axis. J Biol Chem 267: 15122–15133, 1992. 172. Sander M, Neubuser A, Kalamaras J, Ee HC, Martin GR, German MS. Genetic analysis reveals that PAX6 is required for normal transcription of pancreatic hormone genes and islet development. Genes Dev 11: 1662–1673, 1997. 173. Scholey JM, Anderson KV. Intraflagellar transport and ciliumbased signaling. Cell 125: 439 – 442, 2006. 174. Serls AE, Doherty S, Parvatiyar P, Wells JM, Deutsch GH. Different thresholds of fibroblast growth factors pattern the ventral foregut into liver and lung. Development 132: 35– 47, 2005. 175. Shiotani A, Iishi H, Uedo N, Ishiguro S, Tatsuta M, Nakae Y, Kumamoto M, Merchant JL. Evidence that loss of sonic hedgehog is an indicator of Helicobater pylori-induced atrophic gastritis HEDGEHOG SIGNALING 195. 196. 197. 199. 200. 201. 202. 203. 204. Physiol Rev • VOL 205. Wells JM, Melton DA. Early mouse endoderm is patterned by soluble factors from adjacent germ layers. Development 127: 1563– 1572, 2000. 206. Wells JM, Melton DA. Vertebrate endoderm development. Annu Rev Cell Dev Biol 15: 393– 410, 1999. 207. Williams JA, Guicherit OM, Zaharian BI, Xu Y, Chai L, Wichterle H, Kon C, Gatchalian C, Porter JA, Rubin LL, Wang FY. Identification of a small molecule inhibitor of the hedgehog signaling pathway: effects on basal cell carcinoma-like lesions. Proc Natl Acad Sci USA 100: 4616 – 4621, 2003. 208. Wolpert L. One hundred years of positional information. Trends Genet 12: 359 –364, 1996. 209. Xie J, Murone M, Luoh SM, Ryan A, Gu Q, Zhang C, Bonifas JM, Lam CW, Hynes M, Goddard A, Rosenthal A, Epstein EH Jr, de Sauvage FJ. Activating Smoothened mutations in sporadic basal-cell carcinoma. Nature 391: 90 –92, 1998. 210. Yao S, Lum L, Beachy P. The ihog cell-surface proteins bind Hedgehog and mediate pathway activation. Cell 125: 343–357, 2006. 211. Yuasa Y. Control of gut differentiation and intestinal-type gastric carcinogenesis. Nat Rev Cancer 3: 592– 600, 2003. 212. Zeng X, Goetz JA, Suber LM, Scott WJ Jr, Schreiner CM, Robbins DJ. A freely diffusible form of Sonic hedgehog mediates long-range signalling. Nature 411: 716 –720, 2001. 213. Zhang J, Rosenthal A, de Sauvage FJ, Shivdasani RA. Downregulation of Hedgehog signaling is required for organogenesis of the small intestine in Xenopus. Dev Biol 229: 188 –202, 2001. 214. Zhang W, Kang JS, Cole F, Yi MJ, Krauss RS. Cdo functions at multiple points in the Sonic Hedgehog pathway, Cdo-deficient mice accurately model human holoprosencephaly. Dev Cell 10: 657– 665, 2006. 215. Zhang XM, Ramalho-Santos M, McMahon AP. Smoothened mutants reveal redundant roles for Shh and Ihh signaling including regulation of L/R symmetry by the mouse node. Cell 106: 781–792, 2001. 216. Zhu AJ, Zheng L, Suyama K, Scott MP. Altered localization of Drosophila Smoothened protein activates Hedgehog signal transduction. Genes Dev 17: 1240 –1252, 2003. 217. Zhu Y, James RM, Peter A, Lomas C, Cheung F, Harrison DJ, Bader SA. Functional Smoothened is required for expression of GLI3 in colorectal carcinoma cells. Cancer Lett 207: 205–214, 2004. 218. Zweibaum A, Pinto M, Chevalier G, Dussaulx E, Triadou N, Lacroix B, Haffen K, Brun JL, Rousset M. Enterocytic differentiation of a subpopulation of the human colon tumor cell line HT-29 selected for growth in sugar-free medium and its inhibition by glucose. J Cell Physiol 122: 21–29, 1985. 87 • OCTOBER 2007 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.3 on June 17, 2017 198. human homologue of Drosophila patched (PTCH) in basal cell carcinomas and the Gorlin syndrome: different in vivo mechanisms of PTCH inactivation. Cancer Res 56: 4562– 4565, 1996. Van Bokhoven H, Celli J, van Reeuwijk J, Rinne T, Glaudemans B, van Beusekom E, Rieu P, Newbury-Ecob RA, Chiang C, Brunner HG. MYCN haploinsufficiency is associated with reduced brain size and intestinal atresias in Feingold syndrome. Nat Genet 37: 465– 467, 2005. Van de Wetering M, Sancho E, Verweij C, de Lau W, Oving I, Hurlstone A, van der Horn K, Batlle E, Coudreuse D, Haramis AP, Tjon-Pon-Fong M, Moerer P, van den Born M, Soete G, Pals S, Eilers M, Medema R, Clevers H. The beta-catenin/TCF-4 complex imposes a crypt progenitor phenotype on colorectal cancer cells. Cell 111: 241–250, 2002. Van den Brink GR, Bleuming SA, Hardwick JC, Schepman BL, Offerhaus GJ, Keller JJ, Nielsen C, Gaffield W, van Deventer SJ, Roberts DJ, Peppelenbosch MP. Indian Hedgehog is an antagonist of Wnt signaling in colonic epithelial cell differentiation. Nat Genet 36: 277–282, 2004. Van den Brink GR, Hardwick JC, Nielsen C, Xu C, ten Kate FJ, Glickman J, van Deventer SJ, Roberts DJ, Peppelenbosch MP. Sonic hedgehog expression correlates with fundic gland differentiation in the adult gastrointestinal tract. Gut 51: 628 – 633, 2002. Van den Brink GR, Hardwick JC, Tytgat GN, Brink MA, Ten Kate FJ, Van Deventer SJ, Peppelenbosch MP. Sonic hedgehog regulates gastric gland morphogenesis in man and mouse. Gastroenterology 121: 317–328, 2001. Varjosalo M, Li SP, Taipale J. Divergence of hedgehog signal transduction mechanism between Drosophila and mammals. Dev Cell 10: 177–186, 2006. Varjosalo M, Taipale J. Hedgehog signaling. J Cell Sci 120: 3– 6, 2007. Varnat F, Heggeler BB, Grisel P, Boucard N, Corthesy-Theulaz I, Wahli W, Desvergne B. PPARbeta/delta regulates paneth cell differentiation via controlling the hedgehog signaling pathway. Gastroenterology 131: 538 –553, 2006. Wang B, Fallon JF, Beachy PA. Hedgehog-regulated processing of Gli3 produces an anterior/posterior repressor gradient in the developing vertebrate limb. Cell 100: 423– 434, 2000. Wang LC, Nassir F, Liu ZY, Ling L, Kuo F, Crowell T, Olson D, Davidson NO, Burkly LC. Disruption of hedgehog signaling reveals a novel role in intestinal morphogenesis and intestinal-specific lipid metabolism in mice. Gastroenterology 122: 469 – 482, 2002. 1375