Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
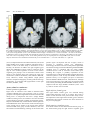
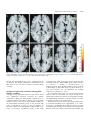
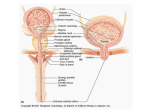
Brain (1998), 121, 2033–2042 Brain activation during micturition in women Bertil F. M. Blok, Leontien M. Sturms and Gert Holstege Department of Anatomy and Embryology, Faculty of Medical Sciences, University of Groningen, Groningen, The Netherlands Correspondence to: Bertil F. M. Blok, Department of Anatomy and Embryology, Faculty of Medical Sciences, University of Groningen, Oostersingel 69, 9713 EZ Groningen, The Netherlands. E-mail: [email protected] Summary Experiments in the cat have led to a concept of how the CNS controls micturition. In a previous study this concept was tested in a PET study in male volunteers. It was demonstrated that specific brainstem and forebrain areas are activated during micturition. It was unfortunate that this study did not involve women, because such results are important for understanding urge incontinence, which occurs more frequently in women than in men. Therefore, a similar study was done in 18 right-handed women, who were scanned during the following four conditions: (i) 15 min prior to micturition (urine withholding); (ii) during micturition; (iii) 15 min after micturition; and (iv) 30 min after micturition. Of the 18 volunteers, 10 were able to micturate during scanning and eight were not, despite trying vigorously. Micturition appeared to be associated with significantly increased blood flow in the right dorsal pontine tegmentum and the right inferior frontal gyrus. Decreased blood flow was found in the right anterior cingulate gyrus during urine withholding. The eight volunteers who were not able to micturate during scanning did not show significantly increased regional cerebral blood flow in the right dorsal, but did so in the right ventral pontine tegmentum. In the cat this region controls the motor neurons of the pelvic floor. In the same unsuccessful micturition group, increased blood flow was also found in the right inferior frontal gyrus. In all 18 volunteers, decreased blood flow in the right anterior cingulate gyrus was found during the period when they had to withhold their urine prior to the micturition condition. The results suggest that in women and in men the same specific nuclei exist in the pontine tegmentum responsible for the control of micturition. The results also indicate that the cortical and pontine micturition sites are more active on the right than on the left side. Keywords: pontine micturition centre; M-region; pontine storage centre; L-region; anterior cingulate gyrus; inferior frontal gyrus Abbreviations: BA 5 Brodmann area; PAG 5 periaqueductal grey; PMC 5 pontine micturition centre; rCBF 5 regional cerebral blood flow; SPM 5 statistical parametric mapping Introduction Micturition or urination is a co-ordinated action between the urinary bladder and its external urethral sphincter. When the bladder contracts, the sphincter relaxes. Although the motor neuronal cell groups of both bladder and sphincter are located in the sacral spinal cord, their co-ordination takes place in the pons. This brainstem organization is best shown in patients with spinal cord injuries above the sacral level. They have great difficulty emptying the bladder, because when their bladder contracts their urethral sphincter also contracts, a disorder called detrusor–sphincter dyssynergia. Such disorders never occur in patients with neurological lesions rostral to the pons, which indicates that the co-ordinating neurons are located in the pontine tegmentum (Blaivas, 1982). As early as 1925, Barrington showed in the cat that the neurons involved in the control of micturition are probably © Oxford University Press 1998 located in the dorsolateral part of the pontine tegmentum, because bilateral lesions in this area produced an inability to empty the bladder, leading to urinary retention. Tracing studies in cat (Holstege et al., 1979) and rat (Loewy et al., 1979) revealed that a distinct cell group in the dorsal pontine tegmentum, called Barrington’s area or the pontine micturition centre (PMC) or M-region, projects to the sacral cord intermediolateral cell column. Blok and Holstege (1997) have shown that this projection is excitatory in nature and contacts dendrites and somata of parasympathetic preganglionic bladder motor neurons. Electrical or chemical stimulation in the PMC produces bladder contractions (Holstege et al., 1986; Mallory et al., 1991) and bilateral destruction of the PMC leads to chronic urinary retention (Griffiths et al., 1990). Another area, important during the filling phase, is located 2034 B. F. M. Blok et al. more ventrally and laterally in the dorsolateral pontine tegmentum. This area, called the L-region, maintains direct projections to the nucleus of Onuf in the sacral cord (Holstege et al., 1979, 1986). In the cat (Sato et al., 1978; Kuzuhara et al., 1980), monkey (Roppolo et al., 1985) and humans (Onufrowicz, 1899; Schröder, 1981), Onuf’s nucleus contains motor neurons innervating the pelvic floor, including the anal and urethral sphincters. Stimulation of the L-region in the cat results in a contraction of the pelvic floor, including the external urethral sphincter (Holstege et al., 1986). Bilateral lesions in the L-region cause an extreme form of ‘urge’ incontinence (Griffiths et al., 1990). Experiments in the cat have led to a concept about the basic micturition control systems in the CNS (Fig. 1). Information about the degree of bladder filling is conveyed by the pelvic nerve to neurons in the lumbosacral cord (Morgan et al., 1981), which in turn project to the periaqueductal grey (PAG) (Noto et al., 1991; Blok et al., 1995; Vanderhorst et al., 1996). When the bladder is filled to such a degree that voiding is appropriate, the PAG activates neurons in the PMC (Blok and Holstege, 1994), which in turn excite the sacral preganglionic parasympathetic bladder motor neurons and inhibit the bladder sphincter motor neurons. The bladder excitation is achieved by way of the direct projection to the parasympathetic motor neurons (Blok and Holstege, 1997), and the sphincter inhibition by way of PMC projections to GABAergic interneurons in the sacral cord dorsal grey commissure (Blok et al., 1997a; Blok and Holstege, 1998). These GABA cells in turn project to the motor neurons in Onuf’s nucleus (Konishi et al., 1985; Nadelhaft et al., 1996). A recent PET study (Blok and Holstege, 1995, 1996; Blok et al., 1997c) in healthy human male volunteers revealed the same brainstem areas to be associated with micturition as in the cat. The first example is the dorsal pontine tegmentum, where the PMC is located in the cat (see also Fukuyama et al., 1996). Other activated areas are the midbrain PAG, the hypothalamus, the right inferior frontal gyrus and the right anterior cingulate gyrus. Withholding of urine, despite vigorous attempts to micturate, was associated with increased regional cerebral blood flow (rCBF) in the ventral pontine tegmentum, an area corresponding to the L-region in the cat, and in the inferior frontal gyrus and anterior cingulate gyrus, all regions on the right side only. PET scan studies are important for our understanding of the micturition control system and its abnormalities. A PET scan study on micturition in women is needed because in women the incidence of neurogenic-related micturition disorders is much greater than in men (Resnick et al., 1989). In particular urge incontinence, in which a patient senses the urge to void but is unable to delay micturition long enough to reach a toilet, is most frequently seen in the elderly population, mostly females (Jewett et al., 1981; Haschek, 1984; Resnick et al., 1989). Since animal studies (e.g. Raisman and Field, 1971; Gorski et al., 1978; Breedlove, 1980) have provided ample evidence for important sex Fig. 1 Schematic overview of pathways between spinal and supraspinal structures involved in the control of micturition based on experiments in cats and men. The locations of the micturition control areas (see text) in the brainstem and diencephalon were used in the null hypotheses for the present study in women. Ascending (sensory-related) and descending (motor-related) pathways are indicated on the respective left and right sides only. BC 5 brachium conjunctivum; CA 5 anterior commissure; IC 5 inferior colliculus; OC 5 optic chiasm; PON 5 pontine nuclei; SC 5 superior colliculus; S2 5 second sacral segment; (1) 5 excitatory effect; (2) 5 inhibitory effect. differences in the structural organization of neuronal cell groups in the CNS, the present study in women was designed to identify the brain regions involved in micturition and to compare the results with the PET scan findings in men, and the anatomical and physiological findings in the cat. Method Experimental design The volunteers were adult women between 20 and 51 years of age (mean 27 years). The subjects completed a general health questionnaire. Volunteers reporting a history of neurological, psychiatric or gastroenterological illness were PET study on micturition in women excluded from the study (n 5 3). The remaining 18 subjects were right-handed, and gave their written informed consent according to the declaration of Helsinki. The protocol of the study was approved by the research ethics committee of the University Hospital of Groningen. During each scan the lights were dimmed, the subjects had their eyes closed and did not move. Each scanning session consisted of four measurements and lasted 1.5 h in total. Experimental protocol and training The volunteers were scanned during the following four conditions: (i) filled bladder, (ii) micturition, (iii) empty bladder and (iv) empty bladder. Eight seconds before the second scan and 15 s after the injection of the H2150 bolus, the right index finger of the volunteer was touched to let her know that she could start micturition. Prior to the other three scans no specific assignment was given. The urine was collected with a special urological device (Femicep bedpan; SIMS Portex Inc., Hythe, UK) attached to a plastic urine reservoir. The device was positioned close to the urethral orifice during all four scans. The floor of the Femicep bedpan was equipped with a self-made battery-driven urine detector, which, during the second condition, indicated the onset of micturition with a small red light. A few days before the scanning session, the volunteers were asked to practise at home, urinating horizontally using the Femicep bedpan. When individual practice was successful, the PET scan session during micturition was simulated at the subjects’ home under the guidance of one of the authors (L.M.S.). Volunteers who were not able to micturate during this session at home were excluded from the study. They were asked to volunteer in a PET study on the voluntary control of the pelvic floor musculature (Blok et al., 1997b). About 70 min prior to the first condition of the actual scanning session, the urine volume of the bladder was measured with the use of an ultrasonic device, called the Bladder Manager (Diagnostic Ultrasound Corporation, Seattle, Wash., USA). If the Bladder Manager indicated that the bladder was hardly filled (,250 ml) and the volunteer affirmed that she did not have a sensation of a filled bladder, she was asked to drink an additional glass of water. When there was successful micturition during condition 2, the bladder volume was measured for a second time to determine the residual urine volume. When the bladder appeared to contain .200 ml, the subject was asked to micturate again. One volunteer was unable to do so, and she was catheterized by a trained physician. Catheterization also took place in the eight volunteers in whom micturition during condition 2 was not successful. 2035 parallel to and 5 cm below the OM (orbitomeatal) line, as determined by external examination. An individually constructed head mould was used to prevent substantial changes in head position. Because of the technical characteristics of the PET camera, the most caudal limit of the scanned area was the pons and the most rostral was the cingulate gyrus, which meant that images were obtained from an axial field of view of 28 mm below to 48 mm above the intercommissural plane. This implied, for example, that the sensorimotor cortex could not be investigated. In order to correct for absorption of γ-radiation by surrounding tissue, a 20-min transmission scan was made at the beginning of the scanning session. The relative attenuation factors, obtained from this scan, were used for correcting the subsequent emission scans and for image reconstruction. After the transmission scan, the subjects were given 1.85 GBq of H215O diluted in saline for each of the four scans. The H215O bolus, followed by 40 ml saline, was injected in the right brachial vein using an automatic pump. Data acquisition continued for 90 s and began 23 s after the beginning of the injection, at which time the peak in radioactivity was assumed to have reached the cerebral circulation. To allow the radiation to return to the background level, there was an interval of 15 min between the injections. Data analysis The data of each scan were summed and the resulting images were centred to prevent loss of information during sampling. Prior to the statistical procedure the data were sampled to a voxel size of 2.2 3 2.2 3 2.4 mm. The data were further analysed using the statistical parametric mapping (SPM) procedure (SPM95 from the Wellcome Department of Cognitive Neurology, London, UK) implemented in Matlab (Mathworks, Sherborn, Mass., USA) on a SPARC workstation (Sun Microsystems, Bagshot, UK). The SPM95 software was used for anatomical realignment, normalization, smoothing and statistical analysis. Realignment corrected the images for the translational and rotational movements of the head, using the first scan as reference. Normalizing spatial transformation matches each scan to a reference or template, which conforms to the stereotaxic standard space (Talairach and Tournoux, 1988). Finally, the images were smoothed with a Gaussian filter of 8 3 8 3 8 mm3 (full width half maximum in the x, y and z axes, respectively). This relatively small filter was used because the main part of the study was aimed at the relatively small brainstem and diencephalon sites that have been found to be involved in the central control of micturition in men (Blok et al., 1997c). Statistical analysis Data acquisition The subject was placed in a horizontal position in the PET camera (Siemens-CTI 951/31, Knoxville, Tenn., USA) The differences in global activity within and between subjects were removed by ANCOVA (analysis of covariance) on a pixel-by-pixel basis with global count as the covariate. For 2036 B. F. M. Blok et al. each pixel in stereotaxic space, the ANCOVA generated a condition-specific adjusted mean rCBF value (normalized to 50 ml/dl/min) and an associated adjusted error variance. A repeated measures ANCOVA was used for the comparison of the four conditions. The following contrasts for both groups were planned: (i) scan 2 (successful/unsuccessful micturition) minus scan 3 (empty bladder), (ii) scan 2 (successful/unsuccessful micturition) minus scan 1 (filled bladder) and (iii) scan 1 (filled bladder) minus the other conditions. The differences between conditions were assessed by weighting each condition with an appropriate contrast. The significance of each contrast was assessed with a statistic whose distribution had Student’s t distribution under the null hypothesis. For each contrast a t statistic was computed for every voxel to form a statistical parametric map [SPM{t}; Friston et al., 1991]. Finally, the SPM{t} was transformed to the unit normal distribution (SPM{Z}). Since the location of the expected micturition related areas was predicted a priori on the basis of the PET study in males, an uncorrected threshold of P , 0.001 was used for these areas. Trends towards activation in the expected micturition control areas were reported when they reached a significance level of P , 0.005 or a Z value of 2.5. This level of significance gives sufficient protection against false positives (Kosslyn et al., 1994; Warburton et al., 1996). The activation in brain areas other than those predicted to be activated during micturition was considered statistically significant only after correction for multiple comparisons. This correction is necessary because, with so many voxelby-voxel comparisons, many t values will reach a conventional level of significance by chance. The problem was resolved by using a Bonferroni-like correction for the number of voxels studied and reporting only those voxels that achieved a corrected level of significance of P , 0.05 after such a correction (Friston et al., 1991). Results Ten of the 18 volunteers were able to micturate within 15 s after the beginning of scan 2. The collected urine volume of this group of volunteers was 423 6 97 ml (mean 6 SD). One successful volunteer, who urinated 320 ml of urine during the second scan, had to be catheterized to remove a urine volume of 380 ml prior to scan 3. The results obtained in this group will be referred to as ‘successful micturition’. The other eight volunteers tried to micturate during scanning but did not succeed (the ‘unsuccessful micturition’ group). Of the unsuccessful micturition group, all were catheterized (urine volume 780 6 162 ml). The data from the two groups are reported separately. Successful micturition group In the 10 subjects who were able to micturate during scanning, the sites showing significant activation (uncorrected P , 0.001) in brain areas previously implicated in micturition control are presented in Table 1. During micturition (scan 2), the right dorsal pontine tegmentum (Fig. 2, left) and the right inferior frontal gyrus were significantly activated compared with the empty bladder condition (scan 3). Using the same comparison, trends towards activation (uncorrected P value between 0.001 and 0.005) were observed in the most caudal extension of the PAG (uncorrected P , 0.002) and the rostral hypothalamus (uncorrected P , 0.004). Other regions, previously not implicated in micturition control, were not found to be significantly activated (corrected P , 0.05). Comparing the micturition condition (scan 2) with the filled bladder condition (scan 1), increased rCBF was observed in the right inferior frontal gyrus with micturition (Fig. 3). A trend towards activation was found in the dorsal pontine tegmentum (uncorrected P , 0.005). With this comparison, other regions that have not previously been implicated in micturition control were not found to be significantly activated (corrected P , 0.05). The rCBF in the right anterior cingulate gyrus (Fig. 3) was significantly decreased during the filled bladder condition (scan 1) compared with the micturition condition (scan 2), but also compared with the empty bladder conditions (scans 3 and 4). Similar significant decreases in rCBF (corrected P , 0.05) were observed during micturition (scan 2) compared with the other three conditions in the left medial frontal gyrus and the right inferior frontal gyrus. Interestingly, the right anterior insula and/or the right frontal operculum were strongly activated (corrected P , 0.003) during the filled bladder condition (scan 1) compared with the empty bladder conditions, and, to a lesser degree, compared with the micturition condition (Z score 5 4.3). Unsuccessful micturition group In the eight subjects who were unable to micturate during scanning, the sites showing significant activation are presented in Table 2. Comparing the second condition (unsuccessful micturition; scan 2) with the condition whilst the bladder was empty (scan 3), significantly increased rCBF (uncorrected P , 0.001) was found in the right ventral pontine tegmentum (Fig. 2, right) and a trend towards significant activation (P , 0.002) in the right inferior frontal gyrus. Comparison of the first condition (withholding of urine) with the unsuccessful micturition condition (scan 2) showed a significant decrease in rCBF (uncorrected P , 0.001) in the right anterior cingulate gyrus. A similar decrease was also found in a comparison with the empty bladder conditions (scans 3 and 4). A trend towards significant activation (P , 0.004) was found in the right inferior frontal gyrus, but not in the ventral pontine tegmentum. The ventral pontine tegmentum was also strongly activated (corrected P , 0.05) during the filled bladder condition (scan 1) compared with the empty bladder conditions (scans 3 and 4). The right anterior insula and/or the right frontal operculum PET study on micturition in women 2037 Table 1 Regional differences in cerebral blood flow during successful micturition Coordinates of peak activation (mm) Micturition, successful (scan 2) minus empty bladder (scan 3) Increased rCBF Right inferior frontal gyrus (BA 44 and 45) Dorsal pontine tegmentum Periaqueductal grey Hypothalamus Decreased rCBF Right inferior frontal gyrus (BA 44 and 45) Left medial frontal gyrus Micturition, successful (scan 2) minus withholding urine (scan 1) Increased rCBF Right inferior frontal gyrus (BA 45 and 46) Right anterior cingulate gyrus (BA 24 and 32) Dorsal pontine tegmentum Decreased rCBF Right inferior frontal gyrus (BA 44 and 45) Left medial frontal gyrus Withholding urine (scan 1) minus micturition, successful (scan 2) Increased rCBF Right frontal operculum and/or anterior insula Decreased rCBF Right anterior cingulate gyrus (BA 24 and 32) Withholding urine (scan 1) minus empty bladder (scan 3) Increased rCBF Right frontal operculum and/or anterior insula Decreased rCBF Right anterior cingulate gyrus (BA 24 and 32) Z score x y z 152 112 12 28 124 236 238 110 112 228 224 28 3.1 3.6 2.8 2.6 146 242 114 140 124 120 4.3 3.6 152 114 116 122 130 240 14 116 228 4.4* 3.6 2.5 146 242 114 140 120 120 4.7* 4.5* 138 110 18 4.3 118 138 18 3.9 138 110 112 5.1* 116 132 112 3.8 Peak activations are indicated by x, y and z coordinates according to the stereotaxic atlas of Talairach and Tournoux (1988). BA 5 estimated Brodmann area. *Significant after multiple comparisons correction with a threshold of corrected P , 0.05. were also strongly activated (corrected P , 0.005) during the filled bladder condition (scan 1) compared with the empty bladder conditions (scan 3 and 4), but not in comparison with the unsuccessful micturition condition (scan 2). Discussion The present study was designed to determine the brain regions activated during micturition in women and to compare the results with those obtained in the previous study in men. It appeared that the micturition-related brain regions in the pons and cerebral cortex in men and women are the same, and that the PAG and the hypothalamus, which have been found to be significantly activated during micturition in men, showed trends towards increased activation in women. It should be stated that the axial field of view of the camera did not involve the more superior parts of the cerebral cortex. Areas related to micturition Dorsal pontine tegmentum A distinct area in the dorsal pontine tegmentum was activated during micturition (scan 2) when compared with the empty bladder condition (scan 3). The location of this pontine area is similar to that of the pontine region found to be activated in men. Since this dorsal pontine region in the cat represents the pontine micturition centre, it seems most likely that a similar group of neurons in the dorsal pons exists in humans (men and women). Right inferior frontal gyrus The activation of the right inferior frontal gyrus during micturition (scan 2) in comparison with a full (scan 1) or empty bladder (scan 3) is the same in men and women, although it is much less extensive in women. The inferior frontal gyrus, in addition to the dorsolateral prefrontal cortex, is involved in attention mechanisms (Pardo et al., 1991) and response selection (Jenkins et al., 1994). With respect to micturition, the area might play a role in deciding whether or not micturition can take place. The observation that this region is activated during micturition is in agreement with specific involvement of the right prefrontal cortex in micturition control (Kuroiwa et al., 1987; Griffiths, 1998). Right anterior cingulate gyrus The right anterior cingulate gyrus showed a significantly decreased rCBF during the withholding of urine condition 2038 B. F. M. Blok et al. Fig. 2 Left: significant differences in rCBF in the right dorsal pontine tegmentum (indicated by pmc 5 pontine micturition centre) after the comparison between conditions ‘successful micturition’ (scan 2) and ‘empty bladder’ (scan 3). Right: significant differences in rCBF in the right ventral pontine tegmentum (indicated by L-region) after the comparison between the conditions ‘unsuccessful micturition’ (scan 2) and ‘empty bladder’ (scan 3). The threshold used for display is uncorrected P , 0.005. The number 228 refers to the distance in millimetres relative to the horizontal plane through the anterior and posterior commissures (z direction). The numbers on the colour scale refer to the corresponding Z scores. Areas with significant activity are superimposed on averaged MRI scans (from six normal subjects) which have been transformed stereotactically to fit a standard atlas. L 5 left side of the brain; R 5 right side. (scan 1) compared with the successful micturition (scan 2) and empty bladder (scans 3 and 4) conditions. This observation is similar to that in the PET study on micturition in men. Possibly, the decrease of rCBF in the right cingulate gyrus during the withholding of urine results in a decrease in the urge to void. Another argument in favour of the anterior cingulate gyrus playing a role in micturition control is that lesions in the forebrain have been reported to cause urge incontinence (Andrew and Nathan, 1964; Maurice-Williams, 1974). Moreover, studies using SPECT (single photon emission computed tomography) scanning indicate that urge incontinence is associated with hypoperfusion of the right forebrain (Griffiths, 1998). Areas related to continence Ventral pontine tegmentum Eight of the 18 volunteers were unable to micturate when requested, although they tried vigorously (scan 2). In this group the ventral pontine tegmentum showed increased rCBF during scan 2 in comparison with scan 3 (empty bladder condition). During scan 2, the volunteers were not successful in their attempt to micturate, probably for emotional reasons. Despite a full bladder they contracted their urethral sphincter and withheld their urine. The location of the activated region in the ventral pons is similar to the region which showed increased rCBF in the group of male volunteers, who were also unable to micturate during scanning. In the cat this same pontine region corresponds with the L-region, which is involved in continence control (see Introduction). Interestingly, this same area in the ventral pontine tegmentum also showed strongly increased rCBF during the filled bladder condition (scan 1) compared with the empty bladder conditions (scans 3 and 4). Activation of the ventral pons, including the L-region, is probably responsible for the increased activity of the striated urethral sphincter during the continence phase in order to keep the bladder closed. This sphincter activity is at its highest just prior to micturition when the bladder is filled (de Groat, 1990). However, although in the successful group one would expect an increased rCBF in the ventral pons during the filled bladder condition (scan 1) compared with the empty bladder conditions (scans 3 and 4), such an increase was not found. Right inferior frontal gyrus The right inferior frontal gyrus was activated during unsuccessful micturition (scan 2) in women. This result is similar to that observed in the previous PET study in men. The importance of the right inferior frontal gyrus in attention mechanisms and response selection has been discussed in the previous paragraph. Right anterior cingulate gyrus In a similar manner to the successful micturition group, in the unsuccessful group the right anterior cingulate gyrus PET study on micturition in women 2039 Fig. 3 Significant differences in rCBF in cortical areas after the comparison between the conditions ‘successful micturition’ (scan 2) and ‘urine withholding’ (scan 1). Note the activations in the right anterior cingulate gyrus (acg) in z planes 18 to 116, and the right inferior frontal gyrus (gfi) in z planes 0 to 112. For other details see legend to Fig. 2. showed decreased rCBF during scan 1 compared with the second scan. This finding is identical to that obtained in the PET study in men who were not able to micturate during scanning. Insular and opercular activation during filled bladder condition The right frontal operculum and/or the right anterior insula were significantly activated (corrected for multiple comparisons) during the filled bladder condition (scan 1) compared with the other three conditions in the successful micturition group, and with the empty bladder condition in the unsuccessful micturition group. In the previous PET study on micturition in men we did not comment on activation related to the filled bladder condition (Blok et al., 1997). In retrospect, unpublished results of this study revealed that the rCBF in the right anterior insula was also increased during the filled bladder phase (peak activation x 5 132 mm, y 5 124 mm, z 5 112 mm; Z score 5 4.1) compared with the empty bladder condition, which is similar to the findings in the present study in women. However, in men this activation was not significant for multiple comparisons (corrected P , 0.05). In a recent PET study (Aziz et al., 1997) the right human anterior insula was associated with the processing of nonpainful and painful sensation from the oesophagus. Possibly, the bladder-filling information is another example of visceral sensation processed by the anterior insula. Alternatively, activation of the right human insula results in increased sympathetic tone (Oppenheimer et al., 1992). Activation of sympathetic fibres has been shown to inhibit mechanoreceptor discharge in the bladder wall (Vaughan and Satchell, 1992). The result is a relaxation of the bladder wall 2040 B. F. M. Blok et al. Table 2 Regional differences in cerebral blood flow during unsuccessful micturition Coordinates of peak activation (mm) Z score x y z Micturition, unsuccessful (scan 2) minus empty bladder (scan 3) Increased rCBF Right inferior frontal gyrus (BA 45) Ventral pontine tegmentum (5 L-region) 136 18 120 222 116 228 2.8 3.8 Micturition, unsuccessful (scan 2) minus withholding urine (scan 1) Increased rCBF Right anterior cingulate gyrus (BA 32) Right inferior frontal gyrus (BA 47) 116 144 136 144 116 112 3.2 2.6 136 18 122 224 112 228 5.1* 4.5* 118 134 120 3.7 Withholding urine (scan 1) minus micturition, unsuccessful (scan 3) Increased rCBF Right frontal operculum and/or anterior insula Ventral pontine tegmentum (5L-region) Decreased rCBF Right anterior cingulate gyrus (BA 24 and 32) For details see Table 1. leading to increased bladder capacity. This is exactly what happened during the first condition of the present study, when the volunteers had a filled bladder but were not allowed to micturate. Predominance of the right brain in micturition control The micturition-related brain regions in women (frontal and cingulate cortices) were located predominantly on the right side of the brain, which is similar to the result of the previous study in men. This finding corresponds with studies indicating that urge incontinence is specifically correlated with lesions in the right hemisphere (Maurice-Williams, 1974; Kuroiwa et al., 1987; Griffiths, 1998). Although this right-sided predominance was also found in the pontine tegmentum, it should be kept in mind that, at least in cats, bilateral lesions of the PMC are necessary to induce urinary retention. Unilateral lesions were not sufficient to produce this effect (Griffiths et al., 1990; Mallory et al., 1991). It is, therefore, premature to conclude that only one side of the brain controls micturition. Acknowledgements We wish to thank all the participating volunteers and members of the Groningen PET centre involved in the micturition project, and A. Nieuwenburg of the Department of Urology and Dr A. M. Blok-Korenhof of the University Hospital of Groningen for performing the catheterization. The Femicep devices were a generous gift from Laprolan B. V. Beuningen, The Netherlands. This research was supported by a grant entitled ‘Neuronal control of micturition (SO-95)’ by the Faculty of Medical Sciences of the University of Groningen. References Andrew J, Nathan PW. Lesions of the anterior frontal lobes and disturbances of micturition and defaecation. Brain 1964; 87: 233–62. Aziz Q, Andersson JL, Valind S, Sundin A, Hamdy S, Jones AK, et al. Identification of human brain loci processing esophageal sensation using positron emission tomography. Gastroenterology 1997; 113: 50–9. Barrington FJF. The effect of lesions of the hind- and mid-brain on micturition in the cat. Q J Exp Physiol Cogn Med Sci 1925; 15: 81–102. Blaivas JG. The neurophysiology of micturition: a clinical study of 550 patients. J Urol 1982; 127: 958–63. Blok BFM, Holstege G. Direct projections from the periaqueductal gray to the pontine micturition center (M-region). An anterograde and retrograde tracing study in the cat. Neurosci Lett 1994; 166: 93–6. Blok BFM, Holstege G. Mapping micturation control areas in the central nervous system with positron emission tomography (PET). Soc Neurosci Abstr 1995; 21: 1972. Blok BFM, Holstege G. The human brain in the control of micturition and urine storage: a positron emission tomography study [abstract]. Neurourol Urodyn 1996; 15: 261. Blok BFM, Holstege G. Ultrastructural evidence for a direct pathway from the pontine micturition center to the parasympathetic preganglionic motoneurons of the bladder of the cat. Neurosci Lett 1997; 222: 195–8. PET study on micturition in women Blok BFM, Holstege G. The central nervous system control of micturition in cats and humans. Behav Brain Res 1998; 92: 119–25. Blok BFM, De Weerd H, Holstege G. Ultrastructural evidence for a paucity of projections from the lumbosacral cord to the pontine micturition center or M-region in the cat. A new concept for the organization of the micturition reflex with the periaqueductal gray as central relay. J Comp Neurol 1995; 359: 300–9. Blok BFM, De Weerd H, Holstege G. The pontine micturition center projects to sacral cord GABA immunoreactive neurons in the cat. Neurosci Lett 1997a; 233: 109–12. 2041 Konishi A, Itoh K, Sugimoto T, Yasui Y, Kaneko T, Takada M, et al. Leucine-enkephalin-like immunoreactive afferent fibers to pudendal motoneurons in the cat. Neurosci Lett 1985; 61: 109–13. Kosslyn SM, Alpert NM, Thompson WL, Chabris CF, Rauch SL, Anderson AK. Identifying objects seen from different viewpoints. A PET investigation. Brain 1994; 117: 1055–71. Kuroiwa Y, Tohgi H, Ono S, Itoh M. Frequency and urgency of micturition in hemiplegic patients: relationship to hemisphere laterality of lesions. J Neurol 1987; 234: 100–2. Blok BFM, Sturms LM, Holstege G. A PET study on cortical and subcortical control of pelvic floor musculature in women. J Comp Neurol 1997b; 389: 535–44. Kuzuhara S, Kanazawa I, Nakanishi T. Topographical localization of the Onuf’s nuclear neurons innervating the rectal and vesical striated sphincter muscles: a retrograde fluorescent double labeling in cat and dog. Neurosci Lett 1980; 16: 125–30. Blok BFM, Willemsen AT, Holstege G. A PET study on brain control of micturition in humans. Brain 1997c; 120: 111–21. Loewy AD, Saper CB, Baker RP. Descending projections from the pontine micturition center. Brain Res 1979; 172: 533–8. Breedlove SM, Arnold AP. Hormone accumulation in a sexually dimorphic motor nucleus of the rat spinal cord. Science 1980; 210: 564–6. Mallory BS, Roppolo JR, de Groat WC. Pharmacological modulation of the pontine micturition center. Brain Res 1991; 546: 310–20. de Groat WC. Central neural control of the lower urinary tract. [Review]. Ciba Found Symp 1990; 151: 27–44. Maurice-Williams RS. Micturition symptoms in frontal tumours. J Neurol Neurosurg Psychiatry 1974; 37: 431–6. Friston KJ, Frith CD, Liddle PF, Frackowiak RS. Comparing functional (PET) images: the assessment of significant change. J Cereb Blood Flow Metab 1991; 11: 690–9. Morgan C, Nadelhaft I, de Groat WC. The distribution of visceral primary afferents from the pelvic nerve to Lissauer’s tract and the spinal gray matter and its relationship to the sacral parasympathetic nucleus. J Comp Neurol 1981; 201: 415–40. Fukuyama H, Matsuzaki S, Ouchi Y, Yamauchi H, Nagahama Y, Kimura J, et al. Neural control of micturition in man examined with single photon emission computed tomography using 99mTcHMPAO. Neuroreport 1996; 7: 3009–12. Nadelhaft I, Vera PL. Neurons in the rat brain and spinal cord labeled after pseudorabies virus injected into the external urethral sphincter. J Comp Neurol 1996; 375: 502–17. Gorski RA, Gordon JH, Shryne JE, Southam AM. Evidence for a morphological sex difference within the medial preoptic area of the rat brain. Brain Res 1978; 148: 333–46. Noto H, Roppolo JR, Steers WD, de Groat WC. Electrophysiological analysis of the ascending and descending components of the micturition reflex pathway in the rat. Brain Res 1991; 549: 95–105. Griffiths D. Clinical studies of cerebral and urinary tract function in elderly people with urinary incontinence. Behav Brain Res 1998; 92: 151–6. Onufrowicz B. Note on the arrangement and function of the cell groups in the sacral region of the spinal cord. J Nerv Ment Dis 1899; 26: 498–504. Griffiths D, Holstege G, Dalm E, De Wall H. Control and coordination of bladder and urethral function in the brainstem of the cat. Neurourol Urodyn 1990; 9: 63–82. Oppenheimer SM, Gelb A, Girvin JP, Hachinski VC. Cardiovascular effects of human insular cortex stimulation. Neurology 1992; 42: 1727–32. Haschek H. Urinary incontinence in the aged. [German]. Z Gerontol 1984; 17: 233–8. Pardo JV, Fox PT, Raichle ME. Localization of a human system for sustained attention by positron emission tomography. Nature 1991; 349: 61–4. Holstege G, Kuypers HG, Boer RC. Anatomical evidence for direct brain stem projections to the somatic motoneuronal cell groups and autonomic preganglionic cell groups in cat spinal cord. Brain Res 1979; 171: 329–33. Raisman G, Field PM. Sexual dimorphism in the preoptic area of the rat. Science 1971; 173: 731–3. Holstege G, Griffiths D, de Wall H, Dalm E. Anatomical and physiological observations on supraspinal control of bladder and urethral sphincter muscles in the cat. J Comp Neurol 1986; 250: 449–61. Resnick NM, Yalla SV, Laurino E. The pathophysiology of urinary incontinence amongst institutionalized elderly persons [see comments]. N Engl J Med 1989; 320: 1–7. Comment in: N Engl J Med 1989; 320: 1421–2. Jenkins IH, Brooks DJ, Nixon PD, Frackowiak RS, Passingham RE. Motor sequence learning: a study with positron emission tomography. J Neurosci 1994; 14: 3775–90. Roppolo JR, Nadelhaft I, de Groat WC. The organization of pudendal motoneurons and primary afferent projections in the spinal cord of the rhesus monkey revealed by horseradish peroxidase. J Comp Neurol 1985; 234: 475–88. Jewett MA, Fernie GR, Holliday PJ, Pim ME. Urinary dysfunction in a geriatric long-term care population. Prevalence and patterns. J Am Geriatr Soc 1981; 29: 211–4. Sato M, Mizuno N, Konishi A. Localization of motoneurons innervating perineal muscles: a HRP study in cat. Brain Res 1978; 140: 149–54. 2042 B. F. M. Blok et al. Schröder HD. Onuf’s nucleus X: a morphological study of a human spinal nucleus. Anat Embryol (Berl) 1981; 162: 443–53. the feline continence process under natural filling conditions. J Neurophysiol 1992; 68: 1842–9. Talairach J, Tournoux P. Co-planar stereotaxic atlas of the human brain. Stuttgart: Thieme; 1988. Warburton E, Wise RJ, Price CJ, Weiller C, Hadar U, Ramsay S, et al. Noun and verb retrieval by normal subjects. [Review]. Brain 1996; 119: 159–79. Vanderhorst VG, Mouton LJ, Blok BF, Holstege G. Distinct cell groups in the lumbosacral cord of the cat project to different areas in the periaqueductal gray. J Comp Neurol 1996; 376: 361–85. Vaughan CW, Satchell PM. Role of sympathetic innervation in Received December 30, 1997. Revised June 3, 1998. Accepted June 24, 1998