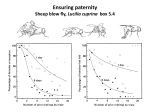
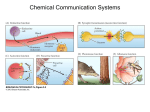
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
J Chem Ecol (2016) 42:631–654 DOI 10.1007/s10886-016-0733-8 Sex and Aggregation-Sex Pheromones of Cerambycid Beetles: Basic Science and Practical Applications Lawrence M. Hanks 1 & Jocelyn G. Millar 2 Received: 6 May 2016 / Revised: 25 June 2016 / Accepted: 21 July 2016 / Published online: 8 August 2016 # Springer Science+Business Media New York 2016 Abstract Research since 2004 has shown that the use of volatile attractants and pheromones is widespread in the large beetle family Cerambycidae, with pheromones now identified from more than 100 species, and likely pheromones for many more. The pheromones identified to date from species in the subfamilies Cerambycinae, Spondylidinae, and Lamiinae are all maleproduced aggregation-sex pheromones that attract both sexes, whereas all known examples for species in the subfamilies Prioninae and Lepturinae are female-produced sex pheromones that attract only males. Here, we summarize the chemistry of the known pheromones, and the optimal methods for their collection, analysis, and synthesis. Attraction of cerambycids to host plant volatiles, interactions between their pheromones and host plant volatiles, and the implications of pheromone chemistry for invasion biology are discussed. We also describe optimized traps, lures, and operational parameters for practical applications of the pheromones in detection, sampling, and management of cerambycids. Keywords Semiochemical . Attractant . Host plant volatile . Invasive species . Detection Submitted to: Journal of Chemical Ecology Electronic supplementary material The online version of this article (doi:10.1007/s10886-016-0733-8) contains supplementary material, which is available to authorized users. * Lawrence M. Hanks [email protected] 1 Department of Entomology, University of Illinois at Urbana-Champaign, Urbana, IL 61801, USA 2 Departments of Entomology, University of California, Riverside, CA 92521, USA Introduction The family Cerambycidae, or the longhorned beetles, is among the largest insect families, with approximately 35,000 described species in the eight subfamilies currently recognized (Švácha and Lawrence 2014). The larvae are endophytic, with the vast majority of species developing in woody plants (Haack 2016). As such, the beetles perform valuable ecosystem services by initiating the degradation of woody materials, and species can be useful indicators of ecosystem health (e.g., Maeto et al. 2002). However, many cerambycid species are important pests in their native regions, damaging and killing trees in natural and managed habitats, and degrading wood destined for lumber (e.g., Solomon 1995). The larvae frequently are long-lived, and consequently are transported readily by global commerce, concealed in wooden products and shipping materials (Haack et al. 2010). As a result, cerambycids are among the most common exotic phytophagous species intercepted in international quarantine (e.g., Brockerhoff et al. 2006; Haack 2006; Liebhold et al. 1995; Nowak et al. 2001). Exotic cerambycids have the potential to become devastating pests when introduced into new regions of the world. For example, the invasion of North America, and the more recent invasion of Europe by the Asian longhorned beetle, Anoplophora glabripennis (Motschulsky) (Meng et al. 2015), represent examples of the invasive potential of cerambycids, and the enormous difficulty and cost in detecting, controlling, or eradicating them from regions they have invaded. On both continents, we are about to witness the huge economic and environmental impact that exotic cerambycids can have on natural and urban forests as A. glabripennis expands its ranges, attacking and killing deciduous trees of many species (Haack et al. 2010). In the context of these and other invasions by cerambycid beetles, pheromone-baited traps that 632 are effective for detecting and sampling the insects would be valuable additions to the otherwise meager tools available for their surveillance, eradication, or management. This review summarizes the rapid progress that has been made since 2004 in the identification and synthesis of cerambycid pheromones, the patterns of pheromone and host plant volatile use by species in the various subfamilies, the development of practical applications of their pheromones, and the implications of pheromone chemistry for invasion biology. Types of Pheromones Used by Cerambycids Despite the economic importance of many cerambycid species in their native countries, and the threat they pose as potential invaders worldwide, remarkably little is known about the basic biology of even common and well-known species (e.g., see Solomon 1995). Until recently, even less was known about their chemical ecology, with attractant pheromones identified from fewer than 10 species (Allison et al. 2004). However, during the past decade, pheromones or likely pheromones have been identified from more than 100 species (Tables 1, S1; Millar and Hanks 2016). The volatile attractant pheromones are of two types: aggregation pheromones (or aggregation-sex pheromones, sensu Cardé 2014), that are produced by males and attract both sexes, or sex pheromones that are produced by females and attract only males (Millar and Hanks 2016). Research to date suggests that the use of these two types of pheromones breaks out along subfamily lines, with species in the Cerambycinae, Lamiinae, and Spondylidinae using aggregation-sex pheromones and species in the Prioninae and Lepturinae using sex pheromones. In general, the chemistries of the two types of pheromones are markedly different, but within each type, there is often remarkable biosynthetic parsimony, with the same or similar pheromone components being shared by species across genera, tribes, and in at least two cases, different subfamilies (Tables 1, S1, discussed below). Moreover, closely related cerambycid species on different continents often have similar, or even identical pheromones, indicating that at least some of these pheromones have persisted unchanged for millions of years. Sex Attractant Pheromones The first female-produced sex attractant pheromone in the Cerambycidae was identified from Prionus californicus Motschulsky (Prioninae) as (3R,5S)-3,5-dimethyldodecanoic acid, given the common name prionic acid (Rodstein et al. 2009, 2011). The pheromone is emitted in very small amounts, likely from an eversible gland on the ovipositor (Barbour et al. 2006). The pheromone is highly attractive, with lures containing submilligram doses remaining attractive for many weeks, and attracting male beetles from hundreds of meters away. Fortunately, the three “non-natural” stereoisomers are not inhibitory (Rodstein et al. 2011), so the relatively inexpensive mixture of all four J Chem Ecol (2016) 42:631–654 stereoisomers can be used in lures. Although it has been possible only to show that prionic acid is present in extracts from females of one other species (Prionus laticollis [Drury], JGM unpub. data), the mixture of stereoisomers is attractive to males of all six North American Prionus species with which it has been tested, as well as the European Prionus coriarius (L.) (Agnello et al. 2011; Barbour et al. 2011). Furthermore, prionic acid was found to be highly attractive to males of the Asian prionine Dorysthenes granulosus (Thomson) (Wickham et al. 2016a), suggesting that the pheromone structure is conserved across at least these two genera. It remains to be seen whether this compound will attract any of the many other species of Dorysthenes that are native to Asia (Löbl and Smetana 2010). A second structural class of female-produced sex pheromones, consisting of 2,3-alkanediols, has been discovered in other prionine genera (Tables 1, S1). Thus, females of several North American species of Tragosoma seem to use various stereoisomers of 2,3-hexanediol as sex attractant pheromones (Ray et al. 2012a), whereas the sex pheromone of the Asian prionine Megopis costipennis (White) consists of (2R,3S)-2,3octanediol (Wickham et al. 2016c). Surprisingly, the same 2,3alkanediol structures are used as aggregation-sex pheromones by many species in the Cerambycinae (see below), and even more remarkable is that the pheromone-producing prothoracic glands seem to be similar between the female prionines and the male cerambycines that produce these types of pheromones (see Ray et al. 2006; Wickham et al. 2016c). Furthermore, both the chemistry of the pheromones and the sites of production are entirely different between species that produce prionic acid and those that produce 2,3-alkanediols (discussed below). Female-produced sex pheromones also have been identified from several species in the Lepturinae (Tables 1, S1). The first such pheromone, identified from Ortholeptura valida (LeConte), consists of cis-vaccenyl acetate. By a biological coincidence, this chemical also is a well-known pheromone component of male fruit flies (Drosophila species; Ray et al. 2011). Another chemical, given the common name (R)desmolactone, is a sex pheromone, or at least sex attractant for several lepturine species in the genus Desmocerus, including the endangered D. californicus dimorphus Fisher (Ray et al. 2012b, 2014). The authors have evidence from coupled gas chromatography-electroantennogram detection (GCEAD) that several other lepturine species have femaleproduced sex pheromones (unpub. data), suggesting that use of sex pheromones is common in the subfamily, but no other structures have been identified yet. Aggregation-Sex Pheromones To date, all volatile pheromones known from species in the Cerambycinae, Lamiinae, and Spondylidinae are male-produced aggregation-sex pheromones (Tables 1, S1). These were the first type of pheromones J Chem Ecol (2016) 42:631–654 Table 1 633 Sources of cerambycid pheromone components, and references to recommended methods of synthesis identified from cerambycids, by Iwabuchi and coworkers, who showed that males of the Asian cerambycine X y l o t re c h u s p y r rh o d e r u s B a t e s p r o d u c e d ( S ) - 2 hydroxyoctan-3-one and (2S,3S)-2,3-octanediol (Iwabuchi et al. 1986; Sakai et al. 1984). The congener X. chinensis Chevrolat subsequently was shown to produce the same two compounds (Iwabuchi et al. 1987). Over the next 15 years, several related structures were identified from males of other 634 Table 1 J Chem Ecol (2016) 42:631–654 (continued) cerambycine species, including (R)-3-hydroxyhexan-2-one, (2R,3R)-2,3-hexanediol, and (2S,3R)-2,3-hexanediol from the European Hylotrupes bajulus (L.), known as the old house borer, and Pyrrhidium sanguinium (L.) (Fettköther et al. 1995; Schröder et al. 1994); (R)-3-hydroxyhexan-2-one and (R)-3hydroxyoctan-2-one from the Asian Anaglyptus subfasciatus J Chem Ecol (2016) 42:631–654 Table 1 635 (continued) Pic (Leal et al. 1995); and (S)-2-hydroxydecan-3-one and 3hydroxydecan-2-one from the Asian Xylotrechus quadripes Chevrolat (Jayarama et al. 1998; Rhainds et al. 2001). The use of these closely related structures by species in different genera and tribes, and from different continents, suggested that the hydroxyketone and 2,3-alkanediol type pheromones were highly conserved within the subfamily, and this has since been borne out by the identification of these types of structures as pheromones or likely pheromones for numerous cerambycine species from different regions of the world (Table S1). The corresponding 2,3-alkanediones are found frequently in headspace volatiles from male beetles, but to date, it seems likely that they are artifacts, because there is no evidence that they are attractants or synergists. In addition, the first variations on the hydroxyketone motif recently have been discovered, such as (2S,4E)-2-hydroxyoct-4-en-3-one and (3R,4E)-3-hydroxyoct-4-en-2-one, which were discovered from Tylonotus bimaculatus Haldeman (Zou et al. 636 2015), i.e., unsaturated analogs of hydroxyoctanone. The first examples of acetylated hydroxyketone structures also have been tentatively identified from cerambycine species (JGM, unpub. data). In addition to the hydroxyketone and 2,3-alkanediol type pheromones, evidence is accumulating that the pheromones of cerambycine species actually may be very diverse (Table S1). For example, very small and simple molecules such as 1-butanol and the enantiomers of 2-methylbutanol have been identified from several different species. Other species such as Megacyllene caryae (Gahan) have pheromones consisting of terpenoids such as geranial and neral (Lacey et al. 2008), whereas Rosalia funebris Motschulsky produces (Z)-3-decenyl (E)-2-hexenoate (Ray et al. 2009a), which resembles compounds from true bugs. Recently, the novel pyrrole structure 1-(1H-pyrrol-2-yl)-1,2-propanedione was found in the headspace volatiles from male beetles in several different cerambycid genera, either alone or in combination with 3-hydroxyhexan-2-one (Table S1; JGM, LMH, and coworkers, unpub. data), and in field trials, it has proven to be a crucial component of attractant blends for at least two invasive Callidiellum species from Asia, C. rufipenne (Motschulsky) (Zou et al. 2016) and C. villosulum (Fairmaire) (Wickham et al. 2016b). Other recently identified pheromones or likely pheromones include the unsaturated straight-chain aldehyde (6E,8Z)-6,8-pentadecadienal (Silva et al. 2016a) and the saturated methyl-branched aldehyde 10-methyldodecanal (Silva et al. 2016b), as well as a pyrone, a tetrahydropyran, and several cyclic terpenoid derivatives (JGM, LMH, and coworkers, unpub. data). Thus, it seems that cerambycine pheromones may be characterized by widely divergent structural types, but with widespread sharing of particular structural types among more closely related species. In contrast, the known or suspected pheromones of species in the Lamiinae fall into two main structural classes. The first class consists of geranylacetone; the corresponding alcohol (E)-6,10-dimethylundeca-5,9-dien-2-ol (known as fuscumol); and its acetate ester (E)-6,10-dimethylundeca-5,9-dien-2-yl acetate (fuscumol acetate), either as single components or as blends (Table S1). The latter two components each have two enantiomeric forms, providing additional opportunities for evolution of species-specific blends. For example, a recent study found evidence in North American lamiines of the relatively rare phenomenon of enantiomeric synergism (Meier et al. 2016): males of Astylidius parvus (LeConte) produce (R)- and (S)-fuscumol + (R)-fuscumol acetate + geranylacetone, whereas males of Lepturges angulatus (LeConte) produce (R)- and (S)-fuscumol acetate + geranylacetone. The fact that several other lamiine species are known to produce nonracemic blends of fuscumol enantiomers (LMH, unpub. Data) suggests that enantiomeric synergism may be common within this subfamily. J Chem Ecol (2016) 42:631–654 The second class of pheromone structures found from lamiine species consists of hydroxyethers and related compounds. The first example, a blend of 4-(heptyloxy)butan-1ol and the corresponding aldehyde, was identified from the notoriously invasive A. glabripennis (Zhang et al. 2002). The alcohol component has now been identified from the Asian congener Anoplophora chinensis (Forster) as well (Hansen et al. 2015). A related pheromone, 2-(undecyloxy) ethanol (known as monochamol) was identified originally from the European Monochamus galloprovincialis Olivier (Pajares et al. 2010), and has since been shown to be a pheromone or likely pheromone for another 13 congeners native to Eurasia and North America (Table S1). Furthermore, during field bioassays in China, monochamol was found to attract species in four other lamiine genera (Acalolepta formosana [Breuning], Pharsalia subgemmata [Thomson], Pseudomacrochenus antennatus [Gahan], and Xenohammus bimaculatus Schwarzer; Wickham et al. 2014), suggesting that it represents another highly conserved pheromone motif. However, the African Monochamus leuconotus (Pascoe) seems to be an exception, because the males produce a different hydroxyether, 2-(heptyloxybutoxy)ethanol (Hall et al. 2006; Pajares et al. 2010). It remains to be seen what other pheromone structures will be found from species in the Lamiinae, and whether they will be as diverse as those turning up among species in the Cerambycinae. To date, all known or suspected pheromones from species in the Spondylidinae are of the geranylacetone structural class shared with some lamiines (Table S1). In fact, fuscumol was first identified from the Palearctic spondylidine Tetropium fuscum (F.) and the North American T. c. cinnamopterum Kirby (Silk et al. 2007; Sweeney et al. 2010). Fuscumol has since been implicated as a pheromone for several other spondylidine species (Table S1). Overall, pheromones or likely pheromones have been identified from only a few species of this relatively small subfamily (see Švácha and Lawrence 2014), and so it is impossible to predict whether this subfamily will turn out to have a diversity of pheromone structures. No pheromones have been identified from species in the remaining three currently recognized subfamilies in the Cerambycidae — the Parandrinae, Necydalinae, and Dorcasominae — all of which are relatively small (Švácha and Lawrence 2014). Conundrums among the Volatile Pheromones of Cerambycidae Over the past decade of studying cerambycid pheromones, the authors and their coworkers have encountered several examples of two currently unexplainable phenomena. First, males of several cerambycine species produce chemicals that are typical pheromone components of cerambycines, but all attempts at attracting the producing species with these compounds have failed. For example, males of the sugar maple pest Glycobius speciosus (Say) and the J Chem Ecol (2016) 42:631–654 important outbreak pest Enaphalodes rufulus (Haldeman) both produce two isomers of 2,3-hexanediol, but all attempts to attract beetles with racemic or chiral compounds, emitted singly or as blends, in traps placed at ground level or in the forest canopy, have failed. Males of Tragidion armatum Linsley produce copious quantities of (R)-3-hydroxyhexan2-one, but no beetles have been attracted to traps by lures containing either the chiral or racemic compound. Finally, males of the very common and abundant locust borer, Megacyllene robiniae (Forster), produce an isomer of 2,3hexanediol, (R)-3-hydroxyhexan-2-one, and (R)-1phenylethan-2-ol, but attempts to trap beetles with all possible blends of components, over trials spanning a decade, have provided no evidence of even the slightest attraction. In all four cases, the compounds are produced sex specifically by males, and their structures are identical to common pheromone components of related species, so it seems unlikely that the compounds might be, for example, defensive compounds instead of pheromones. Conversely, large numbers of both sexes of the invasive species Callidiellum rufipenne were serendipitously attracted to a blend of 3-hydroxyhexan-2-one and 1-(1H-pyrrol-2-yl)1,2-propanedione during a field trial in Japan that targeted a different species (Zou et al. 2016). Although we have found 3hydroxyhexan-2-one in numerous extracts of headspace volatiles from male C. rufipenne, we have never detected the pyrrole in any extracts (Zou et al. 2016). Furthermore, 3hydroxyhexan-2-one alone is not attractive. Thus, it is unclear why both compounds are necessary for attraction of this species when all analyses done to date indicate that the males produce only one of the two components. However, it does suggest that it may be worth field testing the pyrrole as a possible “cryptic” pheromone component for the three currently intractable species described in the previous paragraph. Morphological Structures and Behaviors Associated with Pheromone Use To date, two types of pheromone-producing structures have been identified in various cerambycid species: glands in the prothorax, or glands associated with the ovipositor (Millar and Hanks 2016). Prothoracic glands are common in many species of cerambycines that produce pheromones of the hydroxyalkanone-alkanediol motif, such as Xylotrechus pyrrhoderus (Iwabuchi 1986), Anaglyptus subfasciatus (Nakamuta et al. 1994), Hylotrupes bajulus (Noldt et al. 1995), and Neoclytus a. acuminatus (F.) (Lacey et al. 2007b). Nevertheless, some cerambycines that have the male-specific prothoracic pores have aggregation-sex pheromones with different structures. For example, males of Tylonotus bimaculatus have prominent pores which are absent in females (LMH, unpub. data), and produce a blend of (2S,4E)-2-hydroxyoct-4-en-3-one with smaller amounts of three isomers. This compound is an unsaturated analog of 2hydroxyoctan-3-one, and so does not represent a significant 637 departure from the hydroxyalkanone-alkanediol motif. However, Chlorida festiva (L.) also has the male-specific pores (Ray et al. 2006), but males produce a pheromone of very different structure, (6E,8Z)-6,8-pentadecadienal (Silva et al. 2016a). The pheromone glands of cerambycines are in the endocuticle of the prothorax, and connect by ducts to pores on the cuticular surface (Hoshino et al. 2015). The fact that the prothoracic gland pores are sex specific in cerambycines suggests that they can be used as reliable evidence that a species uses an aggregation-sex pheromone. For example, in a morphological study of cerambycine species, Ray et al. (2006) reported that 49 of the 65 species had the malespecific pores, and indeed, 11 of those species subsequently were found to use aggregation-sex pheromones (Lacey et al. 2008, 2009; Mitchell et al. 2013, 2015; Ray et al. 2015; Zou et al. 2016). In contrast, the absence of male-specific gland pores may indicate either that a species does not use a volatile pheromone, or that the pheromone glands are elsewhere on the body. For example, the cerambycine Rosalia funebris lacks the male-specific gland pores (Ray et al. 2006), and males produce a pheromone with a unique structure ([Z]-3-decenyl [E]-2hexenoate), from as yet unidentified glands (Ray et al. 2009a). Prothoracic gland pores also have been associated with production of female-produced sex pheromones in some species in the Prioninae, with the pores being specific to females. Remarkably, the 2,3-alkanediols used as sex pheromones in these prionine species are the same compounds as are produced by males of many cerambycine species (Table S1). For example, females of the North American Tragosoma depsarium “sp. nov. Laplante” (tribe Meroscelisini) have prothoracic gland pores, and they produce (2R,3R)-2,3-hexanediol as a sex pheromone that attracts only males (Ray et al. 2012a). Similarly, females of the Asian prionine Nepiodes (formerly Megopis) c. costipennis White (tribe Megopidini) have prothoracic gland pores and produce (2R,3S)-2,3-octanediol as a sex pheromone (Wickham et al. 2016c). Another group of species in the Prioninae uses sex pheromones that apparently are produced by an eversible gland on the ovipositor. In female Prionus californicus, extracts of this gland contained prionic acid, as well as several analogs and homologs, although prionic acid alone is both necessary and sufficient for attraction of males (Maki et al. 2011b; Rodstein et al. 2009, 2011). There is little information about biosynthesis of cerambycid pheromones. Kiyota et al. (2009) reasoned that the structural similarity between the (S)-2hydroxyoctan-3-one and (2S,3S)-2,3-octanediol pheromone components of Xylotrechus pyrrhoderus indicated that one was the biosynthetic precursor of the other. They tested this hypothesis by synthesizing deuteriumlabeled standards of each compound, topically applied them to the prothoraces of males, and subsequently 638 collected the chemicals that males produced. This study demonstrated that the ketol was converted to the diol, pinpointing the ketol as the precursor (also see Iwabuchi et al. 2014). For the Spondylidinae, Mayo et al. (2013) suspected that geranylacetone was a biosynthetic precursor of fuscumol for male Tetropium fuscum based on its structural relationship and presence as a minor component in headspace aerations. This was confirmed by topically applying deuterium-labeled geranylacetone to males, resulting in them producing labeled fuscumol. A suspected key biosynthetic enzyme was expressed primarily in the midgut, suggesting that the pheromone was produced there. Zarbin et al. (2013) also applied labeled geranylacetone to different body parts of males of the lamiine Hedypathes betulinus (Klug), demonstrating that the chemical was converted to labeled fuscumol and fuscumol acetate by tissues in the prothorax of males, but there was no such effect in females. Calling behaviors of cerambycids vary with the type of pheromone they produce and the location of their pheromone glands. Males of cerambycines that have the prothoracic gland pores commonly adopt a characteristic “push-up stance” that is thought to aid in dissemination of pheromone: the front legs are extended, elevating the prothorax above the substrate (Lacey et al. 2007a, b, 2009; Ray et al. 2009b). Males of the spondylidine Tetropium fuscum display a similar behavior that is associated with release of the aggregation-sex pheromone fuscumol (Lemay et al. 2010), although the location of the pheromone glands has not yet been verified (Mayo et al. 2013). Female Prionus californicus adopt a very different posture when calling, with the head down, the tip of the abdomen raised, and the ovipositor extended (Barbour et al. 2006). The presumed pheromone gland, on the dorsum of the ovipositor, then is everted. Additional reports of similar postures in female prionines of other species (e.g., Benham and Farrar 1976; Gwynne and Hostetler 1978; Paschen et al. 2012), is further evidence that use of sex pheromones is likely widespread among the prionines, and that pheromones are produced by a gland on the ovipositor. The antennae have been demonstrated to be the organs that cerambycids use to perceive volatile pheromones. The sexually dimorphic, more heavily branched antennae of prionine males suggested that they use sex pheromones to find females (Linsley 1959), in analogy to male Lepidoptera. The role of the antennae as pheromone detectors has been supported by several studies using electroantennography (e.g., Álvarez et al. 2015a; Iwabuchi et al. 1985; MacKay et al. 2015; Ray et al. 2011; Rodstein et al. 2009). To our knowledge, the molecular basis of pheromone reception has been explored only once to date, by Mitchell et al. (2012), who functionally characterized olfactory receptors for three of the nine pheromone components produced by males of the cerambycine Megacyllene caryae. Two of the receptors also were sensitive to isomers J Chem Ecol (2016) 42:631–654 that are pheromone components of other cerambycine species, consistent with the important role of antagonism by components in the pheromone blends of sympatric congeners in averting interspecies attraction (e.g., Mitchell et al. 2015). Nonvolatile Contact Pheromones Once the sexes are in contact, male cerambycids recognize females by contact chemoreception, detecting species- and sex-specific contact pheromones in the cuticular wax layer of females (Hanks and Wang 2016). Several of these contact pheromones have been identified, and they consist of a subset of the cuticular lipids that make up the wax layer (reviewed by Ginzel 2010; Millar and Hanks 2016). Moreover, there is now evidence from several cerambycid species that non-volatile chemicals (possibly lipids) that females deposit on the substrate while walking may act as a chemical trail that males can follow to locate females (Galford 1977; Hoover et al. 2014; Wang 1998, 2002). Implications of Pheromone Chemistry for Invasion Biology The cumulative body of research on cerambycid chemical ecology has shown that the pheromone chemistry of related species often is highly conserved, with many species sharing pheromone components, or even having identical pheromones (Table S1). Moreover, pheromone components may be shared by species on different continents, despite their having diverged many thousands of years ago, notably: 1) hydroxyalkanones and alkanediols among cerambycines; 2) fuscumol, fuscumol acetate, and monochamol among lamiines; 3) fuscumol among spondylidines; and 4) prionic acid and alkanediols among prionines (Table S1). These close similarities in pheromone composition are supported further by the recently discovered 1-(1H-pyrrol-2-yl)-1,2propanedione, which is at least an attractant or synergist for five species of Asian cerambycines (Wickham et al. 2016b; Zou et al. 2016), and a confirmed pheromone component of the North American cerambycine Dryobius sexnotatus Linsley (LMH, unpub. data). This pheromonal parsimony among cerambycids in different parts of the world has profound implications for the successful colonization of exotic species. Specifically, new populations of invaders usually will be founded by a small number of adults emerging from wood products or wooden packing materials. Asynchronous emergence of adults will result in their being widely separated as they disperse into the new region. The dispersion of these few founding adults will be even greater for lamiine species, because adults require a period of maturation feeding before mating. Thus, initial population densities will be very low and it will be difficult for males and females to find one another to mate, the so-called Allee effect (Courchamp et al. 1999). Under these conditions, pheromonal communication for the invading species may be J Chem Ecol (2016) 42:631–654 significantly disrupted by calling adults of much more abundant native species that use the same pheromone components, posing a major barrier to reproduction and establishment. Thus, at least some exotic cerambycid species may fail to establish because their pheromone channel overlaps with those of native species. Conversely, the exotic species that have the best chance of establishing may be those whose pheromone chemistries do not overlap with those of any native species, allowing for efficient and unhindered mate location. In short, the latter species are able to colonize and invade a “pheromone-free space”. It should be noted that the concept of pheromone-free space is by no means exclusive to cerambycids, but likely applies to any invasive insect species that relies on volatile pheromones to bring the sexes together. An example of a successful invader that may have benefited from pheromone-free space is Asian longhorned beetle, the highly polyphagous Asian longhorned borer, Anoplophora glabripennis (Haack et al. 2010). The two-component aggregation-sex pheromone of this species seems to be unique in North America, based on two lines of evidence (Table S1): 1) no native species has yet been found to produce either compound, and 2) no native species have been shown to be significantly attracted by the synthesized pheromones during trapping programs conducted in multiple areas of the eastern USA, and for extended periods of time (DiGirolomo and Dodds 2014; Nehme et al. 2014; unpub. data). Thus, establishment of A. glabripennis may have been favored by its having a unique pheromone, allowing it to independently colonize multiple regions of North America (Haack et al. 2010). In contrast, the European Tetropium fuscum has established in Canada, despite the fact that it uses the same pheromone, fuscumol, as is used by several native spondylidines, including T. c. cinnamopterum. Moreover, it seems that many North American lamiine species share fuscumol as a pheromone component (Table S1). This overlap in pheromone chemistry may explain why the invasion of North America by T. fuscum has stalled, with minimal spread during the 20 years since its introduction, possibly due to low population densities and reproductive failure (Dearborn et al. 2016; Rhainds et al. 2011). It seems possible, and even likely that the geographic range of this invader has been constrained by interference in its pheromone communication by the much more abundant native spondylidines and lamiines that use the same pheromone. Collection, Analysis, and Identification of Cerambycid Pheromones The methods used to identify volatile pheromones of cerambycids depend on the amounts available, which in turn depend to a large extent on the type of pheromone being produced. Specifically, aggregation-sex pheromones generally are produced in microgram or larger amounts, i.e., amounts that are sufficient to obtain NMR spectra. In optimal cases, it 639 has proven possible to simply concentrate and replace the methylene chloride in the extract containing headspace volatiles with deuterated methylene chloride, and run the resulting concentrated extract with microprobe NMR (e.g., Silva et al. 2016a, b). In contrast, female-produced sex pheromones seem to be produced in only nanogram amounts per individual, so all identifications to date have relied on combinations of mass spectral interpretations, microchemical tests, and use of retention indices, because the amounts available were not sufficient for NMR analysis. Below, we describe the methods used to obtain crude pheromone extracts, followed by the specific conditions used for analyses of those extracts. Pheromone Collection Methods The first cerambycid pheromones identified, the aggregation-sex pheromones of Xylotrechus pyrrhoderus and X. chinensis, were collected by confining beetles in glass containers and then rinsing the containers with solvent, or by extracting whole insects or body parts with solvent (Iwabuchi 1986; Iwabuchi et al. 1987; Kuwahara et al. 1987; Sakai et al. 1984). These methods also were used during identification of the pheromone of Hylotrupes bajulus (Fettköther et al. 1995). Crude extracts of most other cerambycid pheromones have been obtained by what has become the standard methodology for collecting odor samples, that is, by passing clean air through a closed chamber containing the insects (with or without host plant material), and trapping the headspace volatiles on an adsorbent such as Tenax (Supelco, Bellefonte, PA, USA), Porapak Q (now Hayesep Q; Sigma-Aldrich, St. Louis, MO, USA), or activated charcoal (Millar and Simms 1998). The trapped volatiles then are eluted with small volumes of a solvent such as methylene chloride. For the aggregation-sex pheromones, aeration samples of a single beetle, if the beetle has produced pheromone at all, usually are dominated by the maleproduced compound(s), so that they are unmistakable, even if the beetle is aerated along with host plant material. In contrast, aeration extracts from species with femaleproduced sex pheromones usually are entirely different, with the pheromone peak being a minor or trace component. Thus, careful comparison of extracts with the corresponding controls, with the analogous extracts from male beetles, or both, is required to locate pheromone candidates. This process is made much more reliable and efficient if the extracts are analyzed by coupled gas chromatography-electroantennography (GCEAD), because the antennae of males respond strongly and selectively to even trace amounts of the pheromone components (e.g., Ray et al. 2011, 2012b, 2014). In the case of Prionus californicus, collections of headspace odors from females did not contain enough material to elicit a detectable GC peak. Instead, a composite sample was finally obtained by inserting a solid phase microextraction fiber into the opening from which the presumed pheromone gland was everted, on the ovipositors of 14 females, followed by thermal desorption of 640 the entire sample directly into the GC column (Rodstein et al. 2009). This method produced a sample with detectable peaks, from which mass spectra were obtained (Rodstein et al. 2009). The identification of the pheromone of P. californicus was particularly difficult because of the high polarity and poor chromatographic properties of the critical compound, the carboxylic acid 3,5-dimethyldodecanoic acid (Rodstein et al. 2009). Analysis of Cerambycid Pheromone Components GCEAD, to detect potential pheromone compounds, and coupled gas chromatography-mass spectrometry (GC-MS), to provide mass spectra and retention indices, are typically the methods used to analyze extracts of insect pheromones, because of the excellent resolution of GC, and the nanogram or less sensitivity of both methods. However, there are two potential pitfalls. First, some cerambycid pheromone components, such as 2methylbutan-1-ol, are very volatile, and so the GC temperature program must have a low initial temperature to ensure that they are not missed, or hidden under the solvent peak. Alternatively, the presence of a very volatile compound can be verified by a solventless collection method, such as solid phase microextraction. Second, the most common types of cerambycine pheromones, the 3-hydroxyalkan-2-ones and the corresponding 2hydroxyalkan-3-ones, are thermally labile and readily interconvert between the two regioisomers in hot GC injector ports or columns (Leal et al. 1995; Sakai et al. 1984). Thus, for analyses of these types of compounds, it is essential to use a relatively low injector temperature (e.g., 100 °C), particularly when running samples in splitless mode where the dwell time of the sample in the injector is many seconds. Alternatively, cool on-column injection (e.g., Rhainds et al. 2001), or programmed temperature vaporization can be used for sample introduction. In parallel, a slow oven temperature program helps to minimize isomerization. Some cerambycid pheromones, such as the hydroxyether compounds produced by species of Monochamus and Anoplophora, are achiral and do not have stereoisomers, and so can be completely identified on the basis of GC retention time and mass spectral matches with standards. However, other types of pheromones may have positional and/or stereoisomers, and so several sequential analyses are required to fully identify the structure, as follows: 1. The enantiomers of 2-methylbutan-1-ol are resolved on a chiral stationary phase Cyclodex B column (J&W Scientific, Folsom, CA, USA), with the (R)-enantiomer eluting first (e.g., Hanks et al. 2007). They also resolve on a Betadex 225 column (J&W Scientific). 2. The positional isomers 2-hydroxyhexan-3-one and 3hydroxyhexan-2-one do not separate well on a nonpolar DB-5 column (J&W Scientific), but they can be distinguished by their mass spectra. Alternatively, all four of the J Chem Ecol (2016) 42:631–654 isomers (i.e., the pairs of enantiomers of each positional isomer) are readily separable on either a Cyclodex B column (elution order 2R:3R:2S:3S) or a Betadex 225 column (retention order 3R:2R:2S:3S). Remarkably, the enantiomers of 3-hydroxyhexan-2-one are separated by several minutes on a Mega-Dex DAC-Beta column (Mega SNC, Legnano, Italy; Andres Gonzalez, pers. comm.). If a chiral stationary phase column is not available, the hydroxyketones can be esterified with (R)- or (S)methoxy(trifluoromethyl)phenyl acetyl chloride to form diastereomeric derivatives that are separable on a standard GC column (e.g., Leal et al. 1995). For the corresponding eight-carbon analogs, the positional isomers again do not separate well on a nonpolar DB-5 column, but are completely separated on a polar DB-Wax column, with the 2-hydroxyoctan-3-one eluting first. On a Cyclodex B column, the elution order is 2R:2S:3R:3S, but the 2S and 3R-stereoisomers nearly overlap. However, by sequential analysis on DB-Wax to determine which positional isomer is present, followed by analysis on Cyclodex B, each isomer can be identified unequivocally. Similarly, the positional isomers of the 10-carbon homologs are not well resolved on a DB-5 column, but they are easily separable on polar GC phases, or they can be readily identified from their mass spectra. The enantiomers of 2hydroxydecan-3-one resolve on a Cyclodex B column (2R elutes first; Rhainds et al. 2001), or on a CP-Chirasil-DexCB column (2R elutes first; Chrompack, Middleberg, The Netherlands; Hall et al. 2006). Similarly, racemic 3hydroxydecan-2-one resolves on Cyclodex B, but we have not yet made enantiomerically enriched standards to determine which peak corresponds to which enantiomer. 3. The stereoisomers of 2,3-hexanediol and 2,3-octanediol are identified readily in two steps. First, the syn- and antidiastereomers of the 2,3-alkanediols are well separated on DB-5 and DB-17 columns, or on polar phases (e.g., Fettköther et al. 1995), with the syn-diastereomers eluting first. Second, all four stereoisomers of each compound separate well on Cyclodex B (elution order for hexanediols and octanediols: 2S,3S: 2R,3R: 2R,3S: 2S,3R). The syn-enantiomers also resolve well on a βcyclodextrin column (SGE, Milton Keynes, UK), with the (2S,3S)-enantiomer eluting first (Hall et al. 2006). 4. The E- and Z-isomers of fuscumol are readily separable by GC; so far, only the E-isomer has been reported as a cerambycid pheromone (Table S1). Fuscumol is not resolved on Cyclodex B or Betadex 225 columns, and is only partially resolved on permethylated cyclodextrin columns (HP-Chiral 20B, Agilent, Santa Clara, CA, USA, Vidal et al. 2010; or β-Dex 120, Supelco, Sweeney et al. 2010). Fuscumol acetate is resolved on Cyclodex B, and J Chem Ecol (2016) 42:631–654 partially resolved on HP-Chiral-20B (Vidal et al. 2010). Alternatively, fuscumol can be derivatized with enantiomerically pure (R)- or (S)-2-acetoxypropionyl chloride (Slessor et al. 1985). The resulting diastereomeric derivatives separate easily on a standard GC column (e.g., Meier et al. 2016; Sweeney et al. 2010). 5. The enantiomers of desmolactone are not resolved on Cyclodex B, but do resolve on β-Dex 225 (Ray et al. 2012b). 6. Prionic acid (3,5-dimethyldodecanoic acid) has four stereoisomers (two diastereomeric pairs of enantiomers). The syn- and anti-diastereomers separate easily on a nonpolar DB-5 column, with syn eluting first. The corresponding methyl esters elute in the same order, with the added advantage that the peak shapes are better, and the retention times are more reproducible than those of the free acids. However, neither of the enantiomeric pairs of methyl esters was resolvable on a Cyclodex B chiral stationary phase column. Thus, the insect-produced enantiomer had to be deduced from bioassays of the two anti enantiomers (i.e., 3R,5S and 3S,5R; Rodstein et al. 2011). Synthesis of Cerambycid Pheromones Table 1 lists all cerambycid pheromones reported to date, where they can be obtained, and/or their recommended methods of synthesis, with the exception of some minor components such as 2-nonanone and some monoterpenes that are readily available from numerous commercial sources. Some further discussion on the syntheses of several of these compounds is warranted. In particular, for practical applications, syntheses of the aggregation-sex pheromones must yield multigram and even kilogram quantities because lures must contain 25–100 mg or more of pheromone components to allow release of several milligrams per day for periods of a few weeks. Thus, syntheses must be short, efficient, and if possible, involve no expensive chromatographic purifications of intermediates or final products. 1. Racemic and (S)-2-methylbutan-1-ol are readily available, but the (R)-enantiomer apparently is not. In our experience, smaller companies purporting to sell the (R)enantiomer actually provided either the (S)-enantiomer or the racemate, i.e., caveat emptor. However, the (R)enantiomer is readily available by enzymatic resolution of the racemate (Bello et al. 2015), or it can be made from commercial (S)-3-bromo-2-methylpropan-1-ol (Mitchell et al. 2012). 2. 2,3-Hexanedione and the corresponding 8- and 10-carbon analogs are seen frequently in aeration extracts of species that produce 2-hydroxyalkan-3-ones or 3-hydroxyalkan-2ones, but to date, have not been shown to be active for any 641 species. Nevertheless, 2,3-hexanedione is commercially available, and analytical standards of the 8- and 10carbon compounds are readily made by oxidation of the corresponding hydroxyketones (e.g., Hall et al. 2006) or permanganate oxidation of 2-alkenes (Rhainds et al. 2001). 3. The hydroxyketone pheromones have been prepared as a mixture of all four stereo- and regioisomers by partial reduction of 2,3-alkanediones using NaBH4 in ethanol (Hanks et al. 2007), or more cleanly by reduction with zinc and ZnCl 2 (Millar et al. 2009). Racemic 3hydroxyalkan-2-ones can be prepared by reaction of the lithium anion of 2-methyl-1,3-dithiane with the appropriate aldehydes, followed by deprotection (Fettköther et al. 1995; Schröder et al. 1994), or by alkylation of trimethylsilyl-protected cyanohydrins with methyllithium followed by hydrolysis of the imine intermediates (Rhainds et al. 2001). However, the recommended route is the one-pot reaction of 3-hydroxy-1-alkynes with BF3 in MeOH with HgO catalyst, followed by aqueous acidic hydrolysis of the intermediate methyl enol (Imrei et al. 2013; Leal et al. 1995). The reaction also can be done in one step using aqueous H2SO4, but larger amounts of the highly toxic HgO catalyst apparently are required (Noseworthy et al. 2012). This reaction uses relatively inexpensive and readily available intermediates, is easily scalable to >100-g quantities; and when reaction conditions are optimized, yields are good to excellent. Enantiomerically enriched 3-hydroxyalkan-2-ones have been prepared by Wacker oxidation of OHprotected chiral 1-alken-3-ols (Schröder et al. 1994), or by ring-opening of protected epoxyalcohols and oxidation, with the epoxyalcohol intermediates being generated from kinetic resolution of allylic alcohols by Sharpless asymmetric epoxidation (Leal et al. 1995). Alternatively, HgO-catalyzed reaction of chiral 1-alkyn-3-ol substrates, as described above, produces the desired hydroxyketones without epimerization (Leal et al. 1995). However, the simplest and recommended route to both enantiomers is by kinetic resolution of the racemate with Amano lipase AK and vinyl acetate, resulting in highly selective acetylation of the (S)-enantiomer. The unchanged (R)-3hydroxyalkanone and the acetylated (S)-enantiomer are separated easily on multigram scale chromatographically (Lacey et al. 2007a). The (S)-enantiomer is recovered by careful alkaline hydrolysis, to minimize epimerization. Alternatively, racemic 3-O-acetylalkan-2-one can be enantioselectively hydrolyzed with the same enzyme in neutral phosphate buffer to produce the (S)-3hydroxyalkan-2-one (Lacey et al. 2007a). Racemic 2-hydroxyalkan-3-ones can be made by reaction of 2-alkyl-1,3-dithianes with acetaldehyde (Fettköther et al. 1995; Schröder et al. 1994), or reaction of the trimethylsilyl- 642 protected cyanohydrin from acetaldehyde with alkyllithiums (Rhainds et al. 2001). However, syntheses of the racemates are probably unnecessary because of the ease with which the enantiomers can be prepared (see below). The enantiomers of 2hydroxyalkan-3-ones have been synthesized by kinetic resolution of allylic alkenols to chiral epoxides followed by ring opening and oxidation (Mori and Otsuka 1985; Rhainds et al. 2001). However, the simplest and cheapest method for synthesis of 2-hydroxyalkan-3-ones is via alkylation of the lithium salts of tetrahydropyran-protected lactic acid enantiomers, a total of four straightforward steps from commercially available (R)- or (S)-ethyl lactates (Hall et al. 2006; Lacey et al. 2008; Sakai et al. 1984). The tetrahydropyran protecting group was found to be best when compared to benzyl or silyl protecting groups (Hall et al. 2006). 4. Mixtures of all four stereoisomers of 2,3-alkanediols can be made by NaBH4 reduction of the corresponding 2,3alkanediones (Hanks et al. 2007) or LiAlH4 or NaBH4 reduction of racemic 2-hydroxyalkan-3-ones or 3hydroxyalkan-2-ones (Fettköther et al. 1995; Lacey et al. 2008). The racemates of the syn- and anti-diastereomers can be prepared in large scale and with good yield in one step by OsO4-catalyzed oxidation of the corresponding (E)- or (Z)-2-alkenes, respectively, with inexpensive N-methylmorpholine N-oxide as the terminal oxidant (Lacey et al. 2004). The individual stereoisomers of 2,3-alkanediols can be prepared in several ways. Historically, several multistep routes were developed based on chiral starting materials (Sakai et al. 1984; Schröder et al. 1994) or the Sharpless asymmetric epoxidation (Mori and Otsuka 1985; Schr öde r et a l. 1 994 ) . Sha r pl e s s a s ym m e t r i c dihydroxylation of stereochemically pure 2-alkenes also has been used to provide a one-step synthesis, but the resulting diols are only ~80–90 % enantiomerically pure (Hall et al. 2006; Lacey et al. 2004). Alternatively, reduction of enantiomerically enriched starting hydroxyketones produces a chromatographically separable mixture of the corresponding syn- and anti-diols (e.g., [R]-3hydroxyhexan-2-one produces a mixture of [2R,3R]- and [2S,3R]-2,3-hexanediols; Hall et al. 2006; Lacey et al. 2008). For multigram scale, kinetic resolution of the racemic anti-diols with Amano lipase PS and vinyl acetate in methyl t-butyl ether is the preferred method, with the (2R,3S)-enantiomer being selectively acetylated (Wickham et al. 2016c). In contrast, the enantiomers of syn-2,3-alkanediols can be prepared in a few steps in multigram amounts from commercially available D- or L-threonine (Mitchell et al. 2012). 5. The enantiomers of fuscumol and fuscumol acetate are prepared easily in multigram scale by kinetic resolution of racemic fuscumol with an immobilized lipase J Chem Ecol (2016) 42:631–654 (Novozyme 435) from the yeast Pseudozyma (=Candida) antarctica (Goto, Sugiy. & Iizuka) Boekhout (Sigma-Aldrich; Hughes et al. 2013; Sweeney et al. 2010; Vidal et al. 2010). Choice of the lipase was important, because other lipases produced much poorer resolution (Sweeney et al. 2010). 6. All of the hydroxyether-type pheromones (e.g., monochamol) are readily prepared by standard Williamson ether syntheses, reacting an alkoxide with an alkyl halide (e.g., Zhang et al. 2002). Only single syntheses of most of the other pheromones shown in Table 1 have been reported, and the citations to those syntheses are listed in the table. Host Plant Volatiles and Bark Beetle Pheromones as Kairomones Many cerambycid beetles mate on the host plants on which the adults feed, or on the host plants of the larvae, suggesting to Linsley (1959) that finding mates and finding host plants are intimately associated. It had long been suspected that the adult beetles primarily use volatile cues to locate host plants (e.g., Beeson 1930), which encouraged some authors to conclude that various species did not use long-range attractant pheromones (e.g., Allison et al. 2004; Hanks et al. 1996; Lu et al. 2007). Much of the research on attraction of cerambycids by plant volatiles has focused on species whose larvae develop in coniferous trees that have been stressed by environmental conditions (e.g., Fan et al. 2007; Hanks 1999; Wood 1982). Because many cerambycid species are polyphagous, it seems likely that the volatile cues used by adults to locate hosts are common to all the host species. Two such compounds, which are broadly attractive to saprophytic insects in general, are ethanol and the monoterpene α-pinene (Millar and Hanks 2016). Volatiles emitted by conifers primarily consist of monoterpenes and sesquiterpenes, with α-pinene being a dominant component (Millar and Hanks 2016). Consequently, αpinene also is a dominant component of the distilled essential oils of conifers known as turpentines (Byers and Zhang 2011; Fan et al. 2007; Kozlowski and Pallardy 1997). In contrast, ethanol is commonly produced by plants that are stressed, damaged, or dying (e.g., from water deficit or fire), and it attracts adults of cerambycid species whose larvae require such hosts (Joseph et al. 2001; Kelsey and Joseph 2003). Ethanol synergizes attraction to α-pinene for some cerambycid species, but not others (e.g., Allison et al. 2001; Chénier and Philogène 1989; Ibeas et al. 2007; Sweeney et al. 2004). Thus, traps baited with either ethanol, α-pinene, or the combination typically attract only a few cerambycid species in large numbers (e.g., Byers 2004; Gardiner 1957; Miller et al. 2015a, b; Montgomery and Wargo 1983). Nevertheless, the combination is commonly, but probably somewhat J Chem Ecol (2016) 42:631–654 erroneously, considered to be a general “plant volatile” attractant for trapping saprophytic insects (e.g., Campbell and Borden 2009; Chénier and Philogène 1989; Costello et al. 2008; Kobayashi et al. 1984). The limited attraction of many cerambycids to blends of ethanol and α-pinene is no doubt due to these compounds representing a crude approximation of the complex volatile profiles of conifers (e.g., Kimmerer and Kozlowski 1982; Yatagai et al. 2002). Ethanol and α-pinene also may fail to attract cerambycids that infest deciduous trees, which have profiles of volatiles that are not dominated by α-pinene. For these and other hosts, other classes of plant volatiles are likely to be more reliable cues for locating larval hosts, and the particular host trees that are in a suitable physiological condition for larval development (see Phillips et al. 1988; Schroeder and Lindelöw 1989). For example, larvae of some cerambycid species develop in fire-damaged conifers, and the adults are attracted to such hosts by the blend of volatiles in smoke (reviewed by Allison et al. 2004; Suckling et al. 2001). For these species, attraction may be antagonized by chemicals that are characteristic of healthy as opposed to fire-damaged trees, i.e., hosts that are not in a condition suitable for the larvae (Suckling et al. 2001; Yamasaki et al. 1997). Similarly, volatiles from tree species that are not natural hosts also may inhibit attraction (Yatagai et al. 2002). Deciduous hosts do indeed produce volatiles that attract adult cerambycids (e.g., Barata and Araújo 2001; Coyle et al. 2015; Fujiwara-Tsujii et al. 2012; Hanks et al. 1996; Liendo-Barandiaran et al. 2010), but there has been little progress in identifying the specific attractive components. Contributing to the problem is the complexity of the blend of volatile chemicals released by host plants. For example, Barata et al. (2000) demonstrated t h a t a n t e n n a e o f th e c er a m b y c i n e P h o r a c a n t h a semipunctata (F.) can detect more than 40 chemicals in odor blends from their eucalypt hosts, but the researchers could not identify two thirds of these chemicals. Adult cerambycids also may use chemical cues to locate host plants for their own feeding. For example, adults of the lamiine Anoplophora malasiaca (Thomson) are attracted to volatiles released from host plants as a result of feeding by conspecifics (see Yasui 2009; Yasui et al. 2007, 2008). Some of these volatiles become absorbed in the beetles’ cuticular lipids, and in turn serve to attract mates. Thus, the host plant volatiles directly and indirectly mediate location of both host plants and mates. Adults of the cerambycine Anaglyptus subfasciatus feed on nectar and pollen, and use floral volatiles to locate their hosts (Ikeda et al. 1993). In a screening trial of floral volatiles, Sakakibara et al. (1998) attracted a few lepturine and cerambycine species to different chemicals, although the vast majority of species in the local community were not attracted by any of the baits. This limited response suggests that the chemicals that were tested may not have 643 simulated the specific blends of floral volatiles that cue attraction of these species. Pheromones of scolytine bark beetles also serve as kairomones for cerambycids (reviewed by Ibeas et al. 2007), guiding them to trees that have had their defenses weakened by being infested by the scolytine larvae. As a bonus, the scolytine larvae are preyed upon facultatively by the cerambycid larvae, providing additional concentrated nutrients (e.g., Allison et al. 2001, 2003; Billings and Cameron 1984; Dodds et al. 2001). Ipsenol and ipsdienol are among the most common bark beetle pheromone components, and the most studied (e.g., Seybold et al. 1995; Wood 1982). A particular bark beetle pheromone may act as a kairomone for one cerambycid species, but antagonize attraction or be entirely neutral for other species (reviewed by Ibeas et al. 2007). The response of a cerambycid species to the pheromone of a particular scolytine species probably is determined by whether they share host species, and by their coevolutionary history. That is, pheromones of a scolytine beetle may serve as kairomones for sympatric cerambycid species, but have no effect on cerambycids native to other parts of the world, or that infest hosts other than those attacked by the particular scolytine species. The response to scolytine pheromones may be influenced further by plant volatiles such as ethanol and α-pinene, which may synergize attraction of some species, but have no effect on others (e.g., Miller et al. 2015a, b). Host Plant Volatiles and Bark Beetle Pheromones as Synergists or Antagonists of Pheromones Plant volatiles and bark beetle pheromones may influence attraction of cerambycids to the aggregation-sex pheromones produced by their males, perhaps by providing an ecological “context” to the chemical signal, indicating that the calling male has discovered a suitable host for the larvae. Again, the effects of host plant volatiles and bark beetle pheromones on responses of cerambycids to their pheromones vary widely (Table S1). Some cerambycid species are not attracted to either their pheromones or host plant volatiles alone, but respond strongly when the two are combined. Others are moderately attracted to their pheromones, with the responses increased additively or synergistically by host plant volatiles. For a third group, and particularly those species using female-produced sex pheromones, the pheromone alone is necessary and sufficient for strong attraction, with host plant or other volatiles apparently playing no role. For example, volatiles associated with spruce trees critically synergize attraction of Tetropium species (Spondylidinae) to their pheromones (Silk et al. 2007; Sweeney et al. 2004). Similarly, ethanol and α-pinene and/or bark beetle pheromones critically synergize attraction of some Monochamus species (Lamiinae) to their pheromones, whereas for other Monochamus species they merely enhance attraction, or seem 644 to have no activity (Allison et al. 2012; Fierke et al. 2012; Hanks et al. 2012; Macias-Samano et al. 2012; Pajares et al. 2010, 2013; Ryall et al. 2015; Teale et al. 2011). For species whose larvae develop in deciduous hosts, ethanol may enhance attraction to aggregation-sex pheromones, whereas monoterpenes such as α-pinene (common volatiles of conifers) may be either neutral or repellent (e.g., Collignon et al. 2016; Hanks et al. 2012). Floral volatiles from host plants of the adults also may influence attraction to pheromone. For example, floral volatiles critically synergized attraction of Anaglyptus subfasciatus to the aggregation-sex pheromone of males, but they were not attractive when tested alone (Nakamuta et al. 1997). Optimization of Pheromone Lures and Pheromone-Baited Traps The development of pheromone-baited traps as effective practical tools for detection and monitoring of cerambycid beetles has been contingent upon several interlinked factors, including design of efficient traps; development of lures with release rates of several milligrams per day; and in some cases, the release of host plant volatiles with the pheromones. The fact that some species require a period of maturation feeding before they produce or respond to pheromones (Hanks and Wang 2016) also may have contributed to earlier failures to recognize that those species were indeed using pheromonal attractants. The iterative optimization of trap and lure design has contributed to the radical paradigm shift that has occurred since 2004, when it was suggested that cerambycids made limited use of volatile pheromones (Allison et al. 2004), to the current situation, where it seems likely that most cerambycid species use volatile sex or aggregation-sex pheromones. As mentioned above, volatile pheromones have been identified from more than 100 species, with likely pheromones known from many more species based on trap catches in pheromone screening trials, and incidental catches in field experiments targeting other species (Table S1). A more detailed discussion of each of these points is useful in understanding why the elucidation of the chemical ecology of the cerambycids has lagged far behind that of many other economically important insects. Trap design is crucial for effective trapping and retention of cerambycids. During our earliest experiments with cerambycid pheromones, we soon discovered that sticky cards and similar traps were ineffective because the relatively large cerambycids either would not walk on the adhesive, or were able to extricate themselves from it (Lacey et al. 2008). Numerous studies have now determined that multiple funnel traps or cross-vane panel traps, both of which mimic the vertical silhouette of tree trunks, are the traps of choice for cerambycids (discussed in Allison et al. 2014; Álvarez et al. 2015b; Graham et al. 2012; Millar and Hanks 2016). J Chem Ecol (2016) 42:631–654 Furthermore, to work effectively, it is essential that all trap surfaces be coated with the Teflon dispersion Fluon, or other lubricants, which render trap surfaces so slippery that the beetles cannot alight such that they immediately fall into the collection bucket (Allison et al. 2011, 2014, 2016; Álvarez et al. 2015b; Graham et al. 2010, 2012). This single factor can increase trap capture more than 10-fold (Graham et al. 2010). Similarly, the inner surface of the collection bucket should be coated with Fluon so that trapped beetles cannot escape, or else the collection bucket should contain a killing and preserving solution such as soapy water, saturated brine, or aqueous propylene glycol (Allison et al. 2014). The Fluon treatment is durable enough to last for several seasons (Allison et al. 2016; Álvarez et al. 2015b). We cannot overemphasize the importance of these factors for effective trapping of cerambycids. Developing lures that can release milligrams of pheromone per day was a second crucial factor in efficient attraction of cerambycids that use aggregation-sex pheromones. In particular, males of these species can release >100 micrograms per male per day of pheromone (e.g., Hanks et al. 2007). However, because males usually release pheromone only for a few of hours per day (during their normal activity period) whereas passive pheromone release devices emit pheromone continuously, the release devices must emit several milligrams per day to match or exceed the release rate from a single male. Thus, pheromone lures for these species typically are made from permeable plastic sachets or similar devices that can achieve these release rates. For example, for research purposes, the authors use small, low-density polyethylene resealable sachets loaded with ethanol or isopropanol solutions of the pheromone as release devices (e.g., Hanks et al. 2014). Adding a cotton wick (1 × 4 cm dental wick) to sachets helps to minimize leakage and to stabilize release rates. These sachets seem to have adequate release rates for many cerambycid pheromones, as judged by trap catches, and they are cheap and readily available almost anywhere in the world. However, these sachets are impractical for operational purposes or commercialization because they must be used immediately after they have been loaded, and so are not suitable for shipping, or storage for extended periods. Nevertheless, for relatively short term field experiments, their simplicity and convenience are major assets. As already discussed, lures must be loaded with tens to hundreds of milligrams of pheromone so that they will emit several milligrams of pheromone per day for periods of a week to several months. Lures that release racemic 3hydroxyhexan-2-one, fuscumol, and other common pheromone components of cerambycid beetles are available from commercial sources (Synergy Semiochemical Corp., Burnaby, BC, Canada; Sylvar Technologies Inc., Fredericton, New Brunswick, Canada; ChemTica Internacional SA, San Jose, Costa Rica; Alpha Scents, West Linn OR), and in some cases have been tested under field conditions (Miller et al. 2015b; J Chem Ecol (2016) 42:631–654 Ryall et al. 2015; Sweeney et al. 2014). Lures that target species of Monochamus with the pheromone monochamol also are available from commercial sources (Alpha Scents, West Linn OR; Econex, Murcia, Spain; SEDQ, Barcelona, Spain; Witasek, Feldkirchen in Kärnten, Austria). The discussion above of high release rates and consequent lure design is primarily applicable to cerambycid species that use aggregation-sex pheromones. The female-produced sex pheromones of species in the Prioninae and Lepturinae are emitted in much smaller amounts, so that for at least some species, lure loadings of a fraction of a milligram remain effective for weeks. In fact, higher loadings may not result in improved trap catch (Rodstein et al. 2011). For species using sex pheromones, the authors have used dilute solutions of the pheromones in plastic sachets as described above, but other devices, such as heat-sealed plastic tubes, also have proven effective (Alston et al. 2015; Meier et al. 2016). In general, it is likely that other commonly used pheromone release devices such as rubber septa would work for these sex pheromones, but this may not always be the case. For example, rubber septa did not work well as release devices for prionic acid, the sex pheromone of Prionus californicus (Rodstein et al. 2009). The height at which pheromone-baited traps are deployed is known to affect catches of lepidopteran species in orchards, where trees are typically no more than a few meters high (e.g., Riedl et al. 1979). This effect may be exacerbated for cerambycids in forests, where the trees are typically much taller, in part as a result of the dynamics of the pheromone plume. However, there is a rapidly growing body of evidence that stratification of different cerambycid species at different heights within forests may be a much more important effect. For example, in field bioassays in east-central Illinois, USA, five cerambycid species were caught in significantly greater numbers by traps that were positioned in the forest canopy (~12 m above the ground) than in the understory, whereas another three species were caught mostly by traps in the understory (1.5 m above the ground; Schmeelk et al. 2016; also see Wong and Hanks 2016). Dodds (2014) reported similar differences among species of cerambycids (and scolytines) as to whether they were caught in greater numbers by traps in the forest canopy versus the understory. Additional indications of stratification were reported in trapping studies in Michigan (Graham et al. 2010) and New Brunswick, Canada (Webster et al. 2016). This stratification in the flight patterns of cerambycids is presumably a consequence of the preferred feeding, mating, and oviposition sites of various species. For example, it would be predicted that adults of species that infest dead wood or roots would be found primarily in the understory, whereas species that feed on foliage of trees and/or whose larvae develop in the branches of living trees would likely spend much of their lives within the canopy (Hanks and Wang 2016). Overall, these studies suggest that unless the preferred height of a target species is known, traps hung at 645 only one height may result in serious underestimation of the local population density. It follows that studies that aim to detect the maximum number of species possible, either for estimating species richness or for detecting invasive species, should deploy traps at several different heights. Failure to do so may result in missing some species altogether. Another useful discovery with major implications for practical applications of pheromone-baited traps was that pheromone components can be blended to create “generic” lures for cerambycids, and these lures attract multiple species simultaneously. This effect first became clear during field bioassays of blends consisting of fewer than 10 cerambycid pheromone components, in which traps baited with these blends in some cases caught many thousands of cerambycid beetles of more than 100 species (Table S1; e.g., USA: Graham et al. 2012; Handley et al. 2015; Hanks and Millar 2013; Hanks et al. 2007, 2012; Australia: Hayes et al. 2016). Ongoing field screening trials conducted by the authors’ collaborators in Asia, Europe, and South America are yielding similar results. Thus, in at least some cases, a series of traps, each of which would have been baited with a single pheromone, now can be replaced by a single trap baited with a blend of pheromones of many species, resulting in cost savings in terms of both materials and labor. Obviously, these generic lures have major implications when the goal is to attract as many species as possible, for example in surveillance efforts to detect incursions of invasive species or in surveys of species richness (e.g., Graham et al. 2012; Handley et al. 2015; Hanks and Millar 2013). For the majority of the known pheromone components, it also is useful that their chemistry is such that they are unlikely to react chemically with each other when mixed in blends, particularly if they are formulated in a diluent such as isopropanol The use of multispecies pheromone blends is most effective when there is the least likelihood of the blend of components producing heterospecific behavioral antagonism, which would otherwise reduce attraction and trap catch of many of the targeted species. Optimal multispecies attraction is therefore achieved when the components are from more distantly related species in different subfamilies (i.e., the chemistry of the pheromones is completely different), and/or when the pheromones are of different types (i.e., aggregation-sex pheromones versus sex pheromones). Using blends of closely related species’ pheromones for multispecies trapping is illadvised because closely related species frequently share pheromone components; recent studies have shown that analogous to the pheromone blends of other insect taxa, the attractiveness of cerambycid pheromone blends can depend strongly on the presence of minor components, either as synergists for conspecifics or as behavioral antagonists reducing the attraction of heterospecifics (e.g., Hanks and Millar 2013; Meier et al. 2016; Mitchell et al. 2015; Ray et al. 2012b, 2015). These synergistic and antagonistic effects provide mechanisms for 646 maintaining the species specificity of signals among sympatric species whose pheromones are dominated by the same single compound. Thus, if the goal is to attract a single species most effectively, then an accurate reconstruction of its pheromone blend should be used. Conversely, if the goal is to attract the maximum number of species with the minimum investment in materials and labor, then formulating mixtures of the pheromone blends of many species will be most cost effective. However, it must be clearly understood that use of such blends may limit or prevent the attraction of species that are strongly inhibited by one or more components. The roles of, and interactions between, minor components of pheromone blends merit further study to assess the extent to which such inhibition may affect trap catches. Applications of Semiochemicals in Detection, Monitoring, and Management of Cerambycid Beetles Pheromones and other semiochemicals have several important uses in insect management, including detection of one or more particular species, monitoring population densities or seasonal phenology of a particular species, and control of a particular species (Millar 2007). The first of these two applications overlap considerably and are thus considered together. Use of Pheromones for Detection and Monitoring of Cerambycids Two of the major advantages of pheromonebaited traps are their sensitivity and their selectivity, both of which have been exploited in the detection of cerambycids. There are several different scenarios in which pheromonebaited traps can be used to advantage in detecting lowdensity populations. First and foremost is the detection of exotic species. The problem of exotic species being inadvertently introduced into new areas of the world is particularly pernicious for cerambycids because their long-lived larvae, concealed within the wood of packing cases, dunnage, pallets, or other wooden materials, are readily transported by international commerce. Thus, species such as the Asian Anoplophora glabripennis have invaded North America and Europe, with the potential for causing devastating losses of hardwoods. Similarly, the Asian species Aromia bungii (Faldermann) has recently invaded Europe (EPPO 2014) and Japan (Kiriyama et al. 2015), where it represents a major threat to fruit trees. For these and numerous other species of cerambycids, semiochemically baited traps may be powerful tools for detecting new incursions as well as monitoring range expansions and the success of eradication efforts. For example, pheromonal attractants are now known for A. glabripennis (Zhang et al. 2002), Callidiellum rufipenne (Zou et al. 2016), and Tetropium fuscum (Sweeney et al. 2010), all of which have invaded North America. Conversely, pheromonal attractants are available for North American species that have invaded Europe, such as J Chem Ecol (2016) 42:631–654 Neoclytus a. acuminatus (Lacey et al. 2004) and several species of Monochamus that have invaded various parts of the world (Table S1). Furthermore, at the time of writing, the authors and their collaborators also have tentatively identified pheromones for several other potentially invasive Asian species, including Anoplophora chinensis (Hansen et al. 2015), Callidiellum villosulum (Wickham et al. 2016b), Xylotrechus rufilius Bates (Narai et al. 2015), as well as A. bungii and Trichoferus campestris (Faldermann) (unpub. data). Semiochemically baited traps also are finding increased use for monitoring populations of species within their native range, such as Monochamus galloprovincialis, the primary vector of the lethal pinewood nematode in Europe (Álvarez et al. 2015b; Rassati et al. 2012). Historically, cerambycids that could cause damage to timber and cut logs have been monitored with traps baited with common host plant volatiles such as α-pinene or turpentine (reviewed in Allison et al. 2004; Millar and Hanks 2016). However, the recent and ongoing identifications of pheromones for several economically important species coupled with concurrent improvements in trap design have improved the efficiency of trapping many fold, and so it seems likely that pheromone-baited traps will rapidly supplant use of traps baited only with host plant compounds or other kairomonal attractants. In addition, pheromone traps may be the most powerful tools available for detecting and monitoring populations of endangered or threatened cerambycid species, such as the iconic Rosalia alpina (L.), a European species that has been red-listed by the International Union for Conservation of Nature (http://www.iucnredlist.org/details/19743/0), or the valley elderberry longhorned borer, Desmocerus californicus dimorphus, in the western USA (Ray et al. 2014). For the latter species, a small survey of several populations with pheromone-baited traps collected more beetles than the entire number held in collections when the species was first listed as threatened by the United States Fish and Wildlife Service (Ray et al. 2014). Similarly, field studies using pheromone-baited traps frequently result in new state records for cerambycid species. For example, of the 69 cerambycid species caught during a study in Delaware, USA using traps baited with a generic pheromone blend, seven species represented new state records (Handley et al. 2015). More generally, cerambycid species perform valuable ecosystem services as primary decomposers of woody biomass and they can be important indicators of ecosystem health (e.g., Maeto et al. 2002). In this context, semiochemically baited traps are powerful tools for conducting surveys of species richness and abundance (e.g., Dodds et al. 2015; Graham et al. 2012; Handley et al. 2015; Hanks and Millar 2013; Hayes et al. 2016; Sweeney et al. 2014; Wickham et al. 2014). They may be particularly useful for surveying species in the forest canopy because these species could easily be missed in ground level surveys conducted by other means. J Chem Ecol (2016) 42:631–654 Use of Pheromones for Control of Cerambycid Beetles For insects in general, three main pheromone-based control strategies have been developed. These include mating disruption with sex pheromones, attract-and-kill using attractants in association with insecticides or pathogens, and mass trapping using traps baited with pheromones, often in combination with host volatiles. The choice of which method to use is dictated by the natural history of the target species, and of course, economics. For example, mating disruption may be most suitable for managing species in which the females produce powerful sex pheromones, because it could prevent them from attracting mates. Nevertheless, mating disruption may still be effective with species that use male-produced aggregation-sex pheromones, as demonstrated in a study in which aerial application of fuscumol-impregnated dispensers over forested areas resulted in significant reductions in populations of Tetropium fuscum (Sweeney et al. 2011a). In terms of economics, the cost of synthesizing, formulating, and deploying pheromone release devices area-wide for mating disruption often may not be competitive with control using insecticides or deployment of plants engineered to produce insecticidal toxins, particularly in crops grown on very large acreages with low profit margins. When viewed in this light, it seems unlikely that pheromone-based mating disruption would be a feasible strategy for use over huge areas, such as in controlling cerambycid species that are pests of forest trees. However, mating disruption could indeed be feasible, and possibly even crucial, for suppression or eradication of newly established invasive species that are limited to small geographic areas. Several pheromone-based strategies have now been tested for control of cerambycid species, with promising results. For instance, mating disruption has been tested twice with Prionus californicus, the females of which produce the powerful sex pheromone prionic acid in nanogram quantities that can attract males from hundreds of meters away. In a preliminary test in hop yards, the number of males that reached a pheromonebaited sentinel trap was reduced by 84 % when the sentinel was encircled by pheromone lures at distances of 18 or 36 m, compared to the number of males reaching a similar sentinel encircled by lures containing only solvent (Maki et al. 2011a). A similar trial in a sweet cherry orchard was even more promising, with pheromone lures reducing by 94 % the number of males reaching a pheromone-baited sentinel trap (J. D. Barbour, pers. comm.). Prionus californicus has a long and asynchronous life cycle (3–5 years), so it may take several years for the effects of mating disruption to be fully realized. Nevertheless, in the latter case, it was noted that after two years, catches of males in monitoring traps had decreased more than seven-fold, suggesting that the strategy was indeed effective in suppressing the local population (Alston et al. 2015). Researchers have been experimenting for several decades with using entomopathogenic fungi (e.g., Hajek and Bauer 647 2009; Petersen-Silva et al. 2015, Sweeney et al. 2013) or nematodes (e.g., Li et al. 1989) for control of cerambycids, including experimenting with attract-and-kill techniques using host plant compounds as the attractant (Li et al. 2007). However, to our knowledge, attract-and-kill using pheromones as attractants and pathogens as the killing agent has not yet been reported for any cerambycid species, although such experiments may be in progress (e.g., see Álvarez-Baz et al. 2015). This technique could prove to be useful for auto-disseminating either fungal conidia or nematodes throughout a population, using either sex or aggregation-sex pheromones as the attractants. In the latter case, both sexes would be contaminated with pathogens upon being attracted to traps, whereas in the former case, dissemination of pathogens to females would occur only during mating with contaminated males. Mass trapping has been explored for control of cerambycid pests in several systems. In a preliminary trial using the female-produced sex pheromone of Prionus californicus, deploying pheromone-baited traps around a central baited sentinel trap decreased by 88 % the number of males caught by the sentinel trap (Maki et al. 2011a). Even more promising, a longer term experiment testing control of P. californicus in sweet cherries showed that after five consecutive years of mass trapping, trap catches in monitoring traps were reduced from 265 to 2 male beetles (Alston et al. 2015). These data suggest that exertion of continuous trapping pressure on this univoltine species, which reproduces relatively slowly due to its long life cycle of several years, can indeed drive populations to low levels, as occurs in mass trapping of large tropical weevils (Faleiro 2006; Oehlschlager 2016, this issue). Furthermore, mass trapping may be a viable strategy for other prionine species that have long life cycles, such as the sugar cane pest Dorysthenes granulosus that is readily caught by using simple pheromone-baited pitfall traps consisting of vertical-sided holes dug in the ground (Wickham et al. 2016a). Mass trapping trials also have been conducted with two species that have aggregation-sex pheromones. Grids of pheromone-baited traps resulted in lower infestation rates of the invasive species Tetropium fuscum in bait logs (Sweeney et al. 2011b). However, this method might only be useful for treating spot infestations because the 10 × 10 m spacing of the traps would likely be prohibitively expensive to deploy over larger areas. The European lamiine Monochamus galloprovincialis, a primary vector of pine wood nematode, also may be targeted for mass trapping using lures consisting of a blend of the pheromone monochamol and bark beetle pheromone synergists, optimized over many years by Pajares and coworkers (e.g., Álvarez et al. 2015b). The lures currently are widely used in Europe to monitor for this pest (e.g., Rassati et al. 2012), but are so effective that trap densities of less than one trap per hectare may reduce beetle population densities by 95 % (Sanchez-Husillos et al. 2015). As an added bonus, 648 because the pheromone is highly conserved within the genus Monochamus (as discussed above), monochamol is the aggregation-sex pheromone of congeners in Europe that also may vector the pinewood nematode. Thus, these species also are strongly attracted to the lure, so the single lure and trap should be effective against multiple species. Summary Based on the studies described herein, it seems likely that cerambycid pheromones will find use in numerous applications, including ecological studies assessing insect communities in different habitats, detection and estimation of the ranges of rare or threatened species, detection and eradication of invasive species, and management of pest species, be they native or exotic. To date, the development of practical applications for cerambycid pheromones has lagged behind that of many other insect groups, for several reasons. For example, it is only in the last decade that it has become clear that, as with so many other insect species, volatile pheromones are crucial mediators of cerambycid reproduction, and these pheromones should be readily exploitable for management of pest species. This paradigm shift has come about in part as a consequence of the “explosion” in the identifications of cerambycid pheromone structures, with sex or aggregation-sex pheromones now known from all five of the major subfamilies within the Cerambycidae. In tandem with the rapidly growing database of pheromone structures (Table S1), great improvements in methods for their practical application have shown that semiochemically based tools can be far more effective than the detection, monitoring, or control methods that they could replace. Few cerambycids are crop pests, so sources of grant funds for studying these insects are less obvious and less available than for important pests of major crops such as corn or soybeans. However, we suggest that the work summarized in this review makes a compelling case for the continued development and incorporation of semiochemicals into programs for detecting, controlling, and where necessary, eradicating exotic cerambycid species. We also suggest several areas for future research. First, it is clear that there are numerous additional pheromone structures waiting to be identified, and there are still no pheromones identified from the small subfamilies Parandrinae, Necydalinae, and Dorcasominae. Further identification of new pheromones has obvious practical implications, but in addition, the new structures will help to flesh out the developing picture of pheromone types and pheromone use within the family, and it is likely that pheromone structures can be used in systematic studies of the evolutionary relationships among species, genera, and tribes. Furthermore, because of the conservation of structures within closely related taxonomic groups, a more detailed plan of pheromone use within the J Chem Ecol (2016) 42:631–654 family will increase our ability to predict whether a species uses volatile pheromones, and if so, whether they are maleproduced aggregation-sex pheromones or female-produced sex pheromones, and what their general structures are likely to be. This information would be invaluable for rapidly developing pheromone-based tools for detecting, tracking, and eradicating new exotic invaders before they have a chance to spread widely. Second, the explosion of identifications of cerambycid semiochemicals is rapidly outpacing our understanding of the roles of these compounds in the ecology of the beetles and the interactions of species with other members of their communities. For example, it remains unclear why the three subfamilies Cerambycinae, Lamiinae, and Spondylidinae seem to have only aggregation-sex pheromones, whereas based on the relatively few pheromones identified from the Prioninae and Lepturinae, these two subfamilies may have only femaleproduced sex pheromones. Numerous other biological and ecological questions abound, such as why some species are attracted to the pheromones of congeners, or why some species produce what seem to be pheromones, but there is no evidence that they are attracted by these compounds. Third, although there have been major advances in trap design, the pheromone lures may still be a weak link in at least some cases. To be widely adopted, pheromone lures must have effective lifetimes of several weeks, and they must be robust, with reasonable shelf life, and be suitable for shipping. This is clearly an area in which improvements can be made. We also suggest that the implications of pheromone chemistry in the success or failure of invasion biology warrants careful investigation. That is, if indeed pheromonal communication by an exotic species is essentially blocked by similar or identical pheromones produced by much more abundant native species, this natural form of mating disruption will reduce the chances of establishment and spread of an invader. Conversely, an exotic species that does not overlap in pheromone chemistry with native species, which therefore is free to invade a “pheromone-free space”, will be more likely to become established. Thus, we suggest that the pheromone chemistry of an exotic species and its overlap in pheromone chemistry with native species should be a factor to consider in developing risk assessments for potentially invasive exotic species. Overall, the rapid expansion of research on cerambycid semiochemicals over the past decade has opened a new window into the biology and ecology of this large and diverse insect family. This new expansion has been most timely because of the increasing problems worldwide with exotic cerambycids as devastating invasive pests. We hope that this review serves as a useful reference for the increasing number of researchers and regulatory personnel working to understand the chemical ecology of cerambycids, and as needed, to manage their populations. J Chem Ecol (2016) 42:631–654 Acknowledgments The authors are grateful for the many students, technicians, postdoctoral and visiting scholars, and collaborators who contributed so extensively to much of the research described herein. We also appreciate helpful comments on the manuscript from reviewer Jon Sweeney. We thank the numerous funding sources which have supported our work on cerambycid semiochemistry over the past 12 years, including the United States Department of Agriculture (USDA) - Animal and Plant Health Inspection Service, the USDA CSREES National Research Initiative, National Institute of Food and Agriculture, Western Regional Integrated Pest Management Fund, North Central Region Integrated Pest Management Center, and the Alphawood Foundation of Chicago. JGM also gratefully acknowledges ongoing funding from Hatch Act project CA-R*ENT-5181-H. References Agnello AM, Loizos L, Gilrein D (2011) A new pheromone for Prionus root-boring beetles. NY Fruit Quart 19:17–19 Allison J, Borden J, McIntosh R, De Groot P, Gries R (2001) Kairomonal response by four Monochamus species to bark beetle pheromones. J Chem Ecol 27:633–646 Allison JD, Morewood WD, Borden JH, Hein KE, Wilson IM (2003) Differential bioactivity of Ips and Dendroctonus (Coleoptera: Scolytidae) pheromone components for Monochamus clamator and M. scutellatus (Coleoptera: Cerambycidae). Environ Entomol 32:23–30 Allison JD, Borden JH, Seybold SJ (2004) A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 14: 123–150 Allison JD, Johnson CW, Meeker JR, Strom BL, Butler SM (2011) Effect of aerosol surface lubricants on the abundance and richness of selected forest insects captured in multiple-funnel and panel traps. J Econ Entomol 104:1258–1264 Allison JD, McKenney JL, Millar JG, McElfresh JS, Mitchell RF, Hanks LM (2012) Response of the woodborers Monochamus carolinensis and Monochamus titillator (Coleoptera: Cerambycidae) to known cerambycid pheromones in the presence and absence of the host plant volatile α-pinene. Environ Entomol 41:1587–1596 Allison JD, Bhandari BD, McKenney JL, Millar JG (2014) Design factors that influence the performance of flight intercept traps for the capture of longhorned beetles (Coleoptera: Cerambycidae) from the subfamilies Lamiinae and Cerambycinae. PloS One 9:e93203 Allison JD, Graham EE, Poland TM, Strom BL (2016) Dilution of fluon before trap surface treatment has no effect on longhorned beetle (Coleoptera: Cerambycidae) captures. J Econ Entomol 109:1215–1219 Alston DG, Barbour JD, Steffen SA (2015) California prionus. Orchard Pest Management Online: 14 November 2015 Álvarez G, Ammagarahalli B, Hall DR, Pajares JA, Gemeno C (2015a) Smoke, pheromone and kairomone olfactory receptor neurons in males and females of the pine sawyer Monochamus galloprovincialis (Olivier) (Coleoptera: Cerambycidae). J Insect Physiol 82:46–55 Álvarez G, Etxebeste I, Gallego D, David G, Bonifacio L, Jactel H, Sousa E, Pajares J (2015b) Optimization of traps for live trapping of pine wood nematode vector Monochamus galloprovincialis. J Appl Entomol 139:618–626 Álvarez-Baz G, Fernández-Bravo M, Pajares J, Quesada-Moraga E (2015) Potential of native Beauveria pseudobassiana strain for biological control of pine wood nematode vector Monochamus galloprovincialis. J Invert Path 132:48–56 Barata EN, Araújo J (2001) Olfactory orientation responses of the eucalyptus woodborer, Phoracantha semipunctata, to host plant in a wind tunnel. Physiol Entomol 26:26–37 649 Barata EN, Pickett JA, Wadhams LJ, Woodcock CM, Mustaparta H (2000) Identification of host and nonhost semiochemicals of eucalyptus woodborer Phoracantha semipunctata by gas chromatography-electroantennography. J Chem Ecol 26:1877–1895 Barbour JD, Cervantes DE, Lacey ES, Hanks LM (2006) Calling behavior in the primitive longhorned beetle Prionus californicus Mots. J Ins Behav 19:623–629 Barbour JD, Millar JG, Rodstein J, Ray AM, Alston DG, Rejzek M, Dutcher JD, Hanks LM (2011) Synthetic 3,5-dimethyldodecanoic acid serves as a general attractant for multiple species of Prionus (Coleoptera: Cerambycidae). Ann Entomol Soc Am 104:588–593 Beeson CFC (1930) Sense of smell of longicorn beetles. Nature 126:12 Bello JE, McElfresh JS, Millar JG (2015) Isolation and determination of absolute configurations of insect-produced methyl-branched hydrocarbons. Proc Natl Acad Sci USA 112:1077–1082 Benham GS, Farrar RJ (1976) Notes on the biology of Prionus laticollis (Coleoptera: Cerambycidae). Can Entomol 108:569–576 Billings RF, Cameron RS (1984) Kairomonal responses of Coleoptera, Monochamus titillator (Cerambycidae), Thanasimus dubius (Cleridae), and Temnochila virescens (Trogositidae), to behavioral chemicals of southern pine bark beetles (Coleoptera: Scolytidae). Environ Entomol 13:1542–1548 Brockerhoff EG, Jones DC, Kimberley MO, Suckling DM, Donaldson T (2006) Nationwide survey for invasive wood-boring and bark beetles (Coleoptera) using traps baited with pheromones and kairomones. For Ecol Manag 228:234–240 Byers JA (2004) Chemical ecology of bark beetles in a complex olfactory landscape. In: Lieutier F, Day K, Battisti A (eds) Bark and wood boring insects in living trees in Europe, a synthesis. Kluwer Academic Publishers, Dordrecht, pp. 89–134 Byers JA, Zhang Q (2011) Chemical ecology of bark beetles in regard to search and selection of host trees. In: Liu T, Kang L, Lei Z, Hernandez R (eds) Recent advances in entomological research. Higher Education Press, Beijing, and Springer-Verlag, Heidelberg and Berlin, pp 150–190 Campbell SA, Borden JH (2009) Additive and synergistic integration of multimodal cues of both hosts and non-hosts during host selection by woodboring insects. Oikos 118:553–563 Cardé RT (2014) Defining attraction and aggregation pheromones: Teleological versus functional perspectives. J Chem Ecol 40:519–520 Chénier JVR, Philogène BJR (1989) Field responses of certain forest Coleoptera to conifer monoterpenes and ethanol. J Chem Ecol 15: 1729–1745 Collignon MR, Swift I, Zou Y, McElfresh JS, Hanks LM, Millar JG (2016) The influence of host plant volatiles on the attraction of longhorned beetles (Coleoptera: Cerambycidae) to pheromones. J Chem Ecol 42:215–229 Costello SL, Negron JF, Jacobi WR (2008) Traps and attractants for wood-boring insects in ponderosa pine stands in the Black Hills, South Dakota. J Econ Entomol 101:409–420 Courchamp F, Clutton-Brock T, Grenfell B (1999) Inverse density dependence and the Allee effect. Trends Ecol Evol 14:405–410 Coyle DR, Brissey CL, Gandhi KJK (2015) Species characterization and responses of subcortical insects to trap-logs and ethanol in a hardwood biomass plantation. Ag For Entomol 17:258–269 Dearborn KW, Heard SB, Sweeney J, Pureswaran DS (2016) Displacement of Tetropium cinnamopterum (Coleoptera: Cerambycidae) by its invasive congener Tetropium fuscum. Environ Entomol doi:10.1093/ee/nvw045 DiGirolomo MF, Dodds KJ (2014) Cerambycidae bycatch for Asian longhorned beetle survey traps placed in forested environs. Northeastern Nat 21:28–34 Dodds KJ (2014) Effects of trap height on captures of arboreal insects in pine stands of northeastern United States of America. Can Entomol 146:80–89 650 Dodds KJ, Graber C, Stephen FM (2001) Facultative intraguild predation by larval Cerambycidae on bark beetle larvae. Comm Ecosyst Ecol 30:17–22 Dodds KJ, Allison JD, Miller DR, Hanavan RP, Sweeney J (2015) Considering species richness and rarity when selecting optimal survey traps: Comparisons of semiochemical baited flight intercept traps for Cerambycidae in eastern North America. Ag For Entomol 17:36–47 EPPO (2014) Pest risk analysis for Aromia bungii. EPPO, Paris. http://www.eppo.int/QUARANTINE/Pest_Risk_Analysis/PRA_ intro.htm. Accessed 2 Apr. 2016 Faleiro J (2006) A review of the issues and management of the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Rhynchophoridae) in coconut and date palm during the last one hundred years. Intl J Trop Ins Sci 26:135–154 Fan J, Sun J, Shi J (2007) Attraction of the Japanese pine sawyer, Monochamus alternatus, to volatiles from stressed host in China. Ann For Sci 64:67–71 Fettköther R, Dettner K, Schröder F, Meyer H, Francke W, Noldt U (1995) The male pheromone of the old house borer Hylotrups bajulus (L.) (Coleoptera: Cerambycidae): identification and female response. Experientia 51:270–277 Fierke MK, Skabeikis DD, Millar JG, Teale SA, McElfresh JS, Hanks LM (2012) Identification of a male-produced aggregation pheromone for Monochamus scutellatus scutellatus and an attractant for the congener Monochamus notatus (Coleoptera: Cerambycidae). J Econ Entomol 105:2029–2034 Fujiwara-Tsujii N, Yasui H, Wakamura S, Hashimoto I, Minamishima M (2012) The white-spotted longicorn beetle, Anoplophora malasiaca (Coleoptera: Cerambycidae), with blueberry as host plant, utilizes host chemicals for male orientation. Appl Entomol Zool 47:103–110 Galford JR (1977) Evidence for a pheromone in the locust borer. USDA For Serv Res Note NE-240:1–3 Gardiner L (1957) Collecting wood-boring beetle adults by turpentine and smoke. Can For Serv, Bi-Monthly Res Note 13:2 Ginzel MD (2010) Hydrocarbons as contact pheromones of longhorned beetles (Coleoptera: Cerambycidae). In: Blomquist GL, Bagnères AG (eds) Insect hydrocarbons: Biology, biochemistry and chemical ecology. Cambridge University Press, Cambridge, pp. 375–389 Graham EE, Mitchell RF, Reagel PF, Barbour JD, Millar JG, Hanks LM (2010) Treating panel traps with a fluoropolymer enhances their efficiency in capturing cerambycid beetles. J Econ Entomol 103:641–647 Graham EE, Poland TM, McCullough DG, Millar JG (2012) A comparison of trap type and height for capturing cerambycid beetles (Coleoptera). J Econ Entomol 105:837–846 Gwynne D, Hostetler BB (1978) Mass emergence of Prionus emarginatus (Say) (Coleoptera: Cerambycidae). Coleopt Bull 32:347–348 Haack RA (2006) Exotic bark- and wood-boring Coleoptera in the United States: Recent establishments and interceptions. Can J For Res 36: 269–288 Haack R (2016) Feeding biology of cerambycids. In: Wang Q (ed) Cerambycidae of the world: Biology and management. CRC Press/Taylor & Francis, Boca Raton (in press) Haack RA, Hérard F, Sun J, Turgeon JJ (2010) Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective. Annu Rev Entomol 55:521–546 Hajek AE, Bauer LS (2009) Use of entomopathogens against invasive wood boring beetles in North America. In: Hajek AE, Glare TR, O'Callaghan MO (eds) Use of microbes for control and eradication of invasive arthropods. Springer Science + Business Media B V, pp 159–179 Hall DR, Cork A, Phythian SJ, Chittamuru S, Jayarama BK, Venkatesha MG, Sreedharan K, Vinod Kumar PK, Seetharama HG, Naidu R (2006) Identification of components of male-produced pheromone of coffee white stem borer, Xylotrechus quadripes. J Chem Ecol 32: 195–219 J Chem Ecol (2016) 42:631–654 Handley K, Hough-Goldstein J, Hanks LM, Millar JG, D’Amico V (2015) Species richness and phenology of cerambycid beetles in urban forest fragments of northern Delaware. Ann Entomol Soc Am 108:251–262 Hanks LM (1999) Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu Rev Entomol 44:483–505 Hanks LM, Millar JG (2013) Field bioassays of cerambycid pheromones reveal widespread parsimony of pheromone structures, enhancement by host plant volatiles, and antagonism by components from heterospecifics. Chemoecology 23:21–44 Hanks LM, Wang Q (2016) Reproductive biology of cerambycid beetles. In: Wang Q (ed) Cerambycidae of the world: Biology and management. CRC Press/Taylor & Francis, Boca Raton (in press) Hanks LM, Millar JG, Paine TD (1996) Mating behavior of the eucalyptus longhorned borer (Coleoptera: Cerambycidae) and the adaptive significance of long “horns”. J Insect Behav 9:383–393 Hanks LM, Millar JG, Moreira JA, Barbour JD, Lacey ES, McElfresh JS, Reuter FR, Ray AM (2007) Using generic pheromone lures to expedite identification of aggregation pheromones for the cerambycid beetles Xylotrechus nauticus, Phymatodes lecontei, and Neoclytus modestus modestus. J Chem Ecol 33:889–907 Hanks LM, Millar JG, Mongold-Diers JA, Wong JCH, Meier LR, Reagel PF, Mitchell RF (2012) Using blends of cerambycid beetle pheromones and host plant volatiles to simultaneously attract a diversity of cerambycid species. Can J For Res 42:1050–1059 Hanks LM, Reagel PF, Mitchell RF, Wong JCH, Meier LR, Silliman CA, Graham EE, Striman BL, Robinson KP, Mongold-Diers JA, Millar JG (2014) Seasonal phenology of the cerambycid beetles of eastcentral Illinois. Ann Entomol Soc Am 107:211–226 Hansen L, Xu T, Wickham J, Chen Y, Hao D, Hanks LM, Millar JG, Teale SA (2015) Identification of a male-produced pheromone component of the citrus longhorned beetle, Anoplophora chinensis. PloS One 10:e0134358 Hayes R, Griffiths MW, Nahrung HF, Arnold PA, Hanks LM, Millar JG (2016) Optimizing generic cerambycid pheromone lures for Australian biosecurity and biodiversity monitoring. J Econ Entomol. doi:10.1093/jee/tow100 Hoover K, Keena M, Nehme M, Wang S, Meng P, Zhang A (2014) Sexspecific trail pheromone mediates complex mate finding behavior in Anoplophora glabripennis. J Chem Ecol 40:169–180 Hoshino K, Nakaba S, Inoue H, Iwabuchi K (2015) Structure and development of male pheromone gland of longicorn beetles and its phylogenetic relationships within the tribe Clytini. J Exp Zool Part B: Mol Dev Evol 324:68–76 Hughes GP, Zou Y, Millar JG, Ginzel MD (2013) (S)-Fuscumol and (S)fuscumol acetate produced by a male Astyleiopus variegatus (Coleoptera: Cerambycidae. Can Entomol 145:327–332 Ibeas F, Gallego D, Diez JJ, Pajares JA (2007) An operative kairomonal lure for managing pine sawyer beetle Monochamus galloprovincialis (Coleoptera: Cerambycidae). J Appl Entomol 131:13–20 Ikeda T, Ohya E, Makihara H, Nakashima T, Saitoh A, Tate K, Kojima K (1993) Olfactory responses of Anaglyptus subfasciatus Pic and Demonax transilis Bates (Coleoptera: Cerambycidae) to flower scents. J Jap For Soc 75:108–112 Imrei Z, Millar JG, Tóth M (2013) Field screening of known pheromone components of longhorned beetles in the subfamily Cerambycinae (Coleoptera: Cerambycidae) in Hungary. Z Naturforsch C 68:236–242 Iwabuchi K (1986) Mating behavior of Xylotrechus pyrrhoderus Bates. III. Pheromone secretion by male. Appl Entomol Zool 21:606–612 I w a b u c h i K , Ta k a h a s h i J , N a k a g a w a Y, S a k a i T ( 1 9 8 5 ) Electroantennogram response of grape borer Xylotrechus pyrrhoderus Bates (Coleoptera: Cerambycidae) to its male sex pheromone components. J Chem Ecol 11:819–828 J Chem Ecol (2016) 42:631–654 Iwabuchi K, Takahashi J, Nakagawa Y, Sakai T (1986) Behavioral responses of female grape borer to synthetic male sex pheromone components. Appl Entomol Zool 21:21–27 Iwabuchi K, Takahashi J, Sakai T (1987) Occurrence of 2,3-octanediol and 2-hydroxy-3-octane, possible male sex pheromone in Xylotrechus chinensis. Appl Entomol Zool 22:110–111 Iwabuchi K, Arakawa M, Kiyota R, Hoshino K, Ando T (2014) Carbonyl reduction in the biosynthesis of a male sex pheromone secreted by the grape borer Xylotrechus pyrrhoderus. J Chem Ecol 40:1146–1151 Jayarama MGV, Souza MV, Naidu R, Hall DR, Cork A (1998) Sex pheromone of coffee white stem borer for monitoring and control is on the anvil. Indian Coffee 62:15–16 Joseph G, Kelsey RG, Peck RW, Niwa CG (2001) Response of some scolytids and their predators to ethanol and 4-allylanisole in pine forests of central Oregon. J Chem Ecol 27:697–715 Kelsey RG, Joseph G (2003) Ethanol in ponderosa pine as an indicator of physiological injury from fire and its relationship to secondary beetles. Can J For Res 33:870–884 Kimmerer TW, Kozlowski TT (1982) Ethylene, ethane, acetaldehyde, and ethanol production by plants under stress. Plant Physiol 69:840–847 Kiriyama S, Iwata R, Kagaya E (2015) Newly discovered populations of Aromia bungii (Faldermann), an invasive cerambycid infesting cherry and Japanese apricot trees in Tatebayashi, Gunma Pref., and Fussa, Tokyo Pref. Plant Protection 69:807–809 (in Japanese) Kiyota R, Yamakawa R, Iwabuchi K, Hoshino K, Ando T (2009) Synthesis of deuterated sex pheromone components of the grape borer, Xylotrechus pyrrhoderus. Biosci Biotech Biochem 73:2252–2256 Kobayashi F, Yamane A, Ikeda T (1984) The Japanese pine sawyer beetle as the vector of pine wilt disease. Annu Rev Entomol 29:115–135 Kozlowski TT, Pallardy SG (1997) Physiology of woody plants, 2nd edn. Academic Press, San Diego Kuwahara Y, Matsuyama S, Suzuki T (1987) Identification of 2,3-octanediol, 2-hydroxy-3-octanone and 3-hydroxy-2-octanone from male Xylotrechus chinensis Chevrolat as possible sex pheromones. Appl Entomol Zool 22:25–28 Lacey ES, Ginzel MD, Millar JG, Hanks LM (2004) Male-produced aggregation pheromone of the cerambycid beetle Neoclytus acuminatus acuminatus. J Chem Ecol 30:1493–1507 Lacey ES, Moreira JA, Millar JG, Ray AM, Hanks LM (2007a) Maleproduced aggregation pheromone of the cerambycid beetle Neoclytus mucronatus mucronatus. Entomol Exp Appl 122:171–179 Lacey ES, Ray AM, Hanks LM (2007b) Calling behavior of the cerambycid beetle Neoclytus acuminatus acuminatus (F.). J Insect Behav 20:117–128 Lacey ES, Moreira JA, Millar JG, Hanks LM (2008) A male-produced aggregation pheromone blend consisting of alkanediols, terpenoids, and an aromatic alcohol from the cerambycid beetle Megacyllene caryae. J Chem Ecol 34:408–417 Lacey ES, Millar JG, Moreira JA, Hanks LM (2009) Male-produced aggregation pheromones of the cerambycid beetles Xylotrechus colonus and Sarosesthes fulminans. J Chem Ecol 35:733–740 Leal WS, Shi XW, Nakamuta K, Ono M, Meinwald J (1995) Structure, stereochemistry, and thermal isomerization of the male sex pheromone of the longhorn beetle Anaglyptus subfasciatus. Proc Natl Acad Sci U S A 92:1038–1042 Lemay MA, Silk PJ, Sweeney J (2010) Calling behavior of Tetropium fuscum (Coleoptera: Cerambycidae: Spondylidinae). Can Entomol 142:256–260 Li W, Wu C, Wang C, Chai C (1989) Application of pathogenic nematode against cerambycids. In: Kobayashi K, Ikeda T, Tanaka K, Nakamuta K (eds) Forest insect pests and tree diseases in the northeast Asia. Proc IUFRO Regional Workshop, Aug. 28-Sept 1, 1989, Tsukuba, Japan, pp 56–65 Li Z, Fan M, Wang S, Ding D (2007) Beauveria bassiana plus chemical attractant: A new approach against pine sawyer? Proc Ann Mtg Soc Invertebr Pathol, Quebec City, Quebec Canada, pp. 113–114 651 Liebhold AM, MacDonald WL, Bergdahl D, Mastro VC (1995) Invasion by exotic forest pests: A threat to forest ecosystems. For Sci Mono 30:1–48 Liendo-Barandiaran CV, Herrera-Malaver B, Morillo F, Sánchez P, Hernández JV (2010) Behavioral responses of Steirastoma breve (Sulzer) (Coleoptera: Cerambycidae) to host plant Theobroma cacao L., brushwood piles, under field conditions. Appl Entomol Zool 45:489–496 Linsley EG (1959) Ecology of Cerambycidae. Annu Rev Entomol 4:99–138 Löbl I, Smetana A (2010). Catalogue of the Palaearctic Coleoptera, Vol. 6: Chrysomeloidea. Apollo Books, Stenstrup, Demark Lu W, Wang Q, Tian MY, He XZ, Zeng XL, Zhong YX (2007) Mate location and recognition in Glenea cantor (Fabr.) (Coleoptera: Cerambycidae: Lamiinae): roles of host plant health, female sex pheromone, and vision. Environ Entomol 36:864–870 Macias-Samano JE, Wakarchuk D, Millar JG, Hanks LM (2012) 2Undecyloxy-1-ethanol in combination with other semiochemicals attracts three Monochamus species (Coleoptera: Cerambycidae) in British Columbia, Canada. Can Entomol 144:764–768 MacKay CA, Sweeney JD, Hillier NK (2015) Olfactory receptor neuron responses of a longhorned beetle, Tetropium fuscum (Fabr.) (Coleoptera: Cerambycidae), to pheromone, host, and non-host volatiles. J Insect Physiol 83:65–73 Maeto K, Sato S, Miyata H (2002) Species diversity of longicorn beetles in humid warm-temperate forests: The impact of forest management practices on old-growth forest species in southwestern Japan. Biodiv Conserv 11:1919–1937 Maki EC, Millar JG, Rodstein J, Hanks LM, Barbour JD (2011a) Evaluation of mass trapping and mating disruption for managing Prionus californicus (Coleoptera: Cerambycidae) in hop production yards. J Econ Entomol 104:933–938 Maki EC, Rodstein J, Millar JG, Barbour KS, Hanks LM, Barbour JD (2011b) Synthesis and field tests of possible minor components of the sex pheromones of Prionus californicus. J Chem Ecol 37:714–716 Mayo PD, Silk PJ, Cusson M, Béliveau C (2013) Steps in the biosynthesis of fuscumol in the longhorn beetles Tetropium fuscum (F.) and Tetropium cinnamopterum Kirby. J Chem Ecol 39:377–389 Meier LR, Zou Y, Millar JG, Mongold-Diers JA, Hanks LM (2016) Synergism between enantiomers creates species-specific pheromone blends and minimizes interspecific attraction for two cerambycid beetle species. J Chem Ecol (in press) Meng PS, Hoover K, Keena MA (2015) Asian longhorned beetle (Coleoptera: Cerambycidae), an introduced pest of maple and other hardwood trees in North America and Europe. J Integ Pest Manag 6:1–13 Millar JG (2007) Insect pheromones for integrated pest management: Promise versus reality. Redia 90:51–55 Millar JG, Hanks LM (2016) Chemical ecology of cerambycid beetles. In: Wang Q (ed) Cerambycidae of the world: Biology and management. CRC Press/Taylor & Francis, Boca Raton (in press) Millar JG, Simms JJ (1998) Preparation, cleanup, and preliminary fractionation of extracts. In: Millar JG, Haines KF (eds) Methods in chemical ecology: Chemical methods. Kluwer Academic Publishers, Norwell, pp. 1–37 Millar JG, Hanks LM, Moreira JA, Barbour JD, Lacey ES (2009) Pheromone chemistry of cerambycid beetles. In: Nakamuta K, Millar JG (eds) Chemical ecology of wood-boring insects. Forestry and Forest Products Research Institute, Ibaraki, Japan, pp. 52–79 Miller DR, Crowe CM, Dodds KJ, Galligan LD, De Groot P, Hoebeke ER, Mayfield AE, Poland TM, Raffa KF, Sweeney JD (2015a) Ipsenol, ipsdienol, ethanol, and α-pinene: trap lure blend for Cerambycidae and Buprestidae (Coleoptera) in pine forests of eastern North America. J Econ Entomol 108:1837–1851 652 Miller DR, Crowe CM, Mayo PD, Silk PJ, Sweeney JD (2015b) Responses of Cerambycidae and other insects to traps baited with ethanol, 2,3-hexanediol, and 3,2-hydroxyketone lures in northCentral Georgia. J Econ Entomol 108:2354–2365 Mitchell RF, Hughes DT, Luetje CW, Millar JG, Soriano-Agatón F, Hanks LM, Robertson HM (2012) Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem Mol Biol 42:499–505 Mitchell RF, Millar JG, Hanks LM (2013) Blends of (R)-3hydroxyhexan-2-one and alkan-2-ones identified as potential pheromones produced by three species of cerambycid beetles. Chemoecology 23:121–127 Mitchell RF, Reagel PF, Wong JCH, Meier LR, Silva WD, MongoldDiers J, Millar JG, Hanks LM (2015) Cerambycid beetle species with similar pheromones are segregated by phenology and minor pheromone components. J Chem Ecol 41:431–440 Montgomery ME, Wargo PM (1983) Ethanol and other host-derived volatiles as attractants to beetles that bore into hardwoods. J Chem Ecol 9:181–190 Mori K, Otsuka T (1985) Synthesis of (2S,3S)-2,3-octanediol and (S)-2hydroxy-3-octanone, the male sex pheromone of the grape borer Xylotrechus pyrrhoderus. Tetrahedron 41:553–556 Nakamuta K, Sato H, Nakashima T (1994) Behavioral and morphological evidence for a male-produced sex pheromone in the Cryptomeria twig borer, Anaglyptus subfasciatus Pic (Coleoptera, Cerambycidae). Jpn. J Entomol 62:371–376 Nakamuta K, Leal WS, Nakashima T, Tokoro M, Ono M, Nakanishi M (1997) Increase of trap catches by a combination of male sex pheromones and floral attractant in longhorn beetle, Anaglyptus subfasciatus. J Chem Ecol 23:1635–1640 Narai Y, Zou Y, Nakamuta K, Mongold-Diers JA, Hanks LM, Millar JG (2015) Candidate aggregation pheromones of two potentially invasive Asian cerambycid species in the genus Xylotrechus. J Econ Entomol 108:1444–1446 Nehme ME, Trotter RT, Keena MA, McFarland C, Coop J, Hull-Sanders HM, Meng P, De Moraes CM, Mescher MC, Hoover K (2014) Development and evaluation of a trapping system for Anoplophora glabripennis (Coleoptera: Cerambycidae) in the United States. Environ Entomol 43:1034–1044 Noldt U, Fettköther R, Dettner K (1995) Structure of the sex pheromoneproducing prothoracic glands of the male old house borer. Intl J Morph Embryol 24:223–234 Noseworthy MK, Humble LM, Sweeney J, Silk P, Mayo P (2012) Attraction of Monarthrum scutellare (Coleoptera: Curculionidae: Scolytinae) to hydroxy ketones and host volatiles. Can J For Res 42:1851–1857 Nowak DJ, Pasek JE, Sequeira RA, Crane DE, Mastro VC (2001) Potential effect of Anoplophora glabripennis (Coleoptera: Cerambycidae) on urban trees in the United States. J Econ Entomol 94:116–122 Oehlschlager AC (2016) Palm weevil pheromones - discovery and use. J Chem Ecol (this issue) Pajares JA, Álvarez G, Ibeas F, Gallego D, Hall DR, Farman DI (2010) Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J Chem Ecol 36:570–583 Pajares JA, Álvarez G, Hall DR, Douglas P, Centeno F, Ibarra N, Schroeder M, Teale SA, Wang Z, Yan S, Millar J, Hanks LM (2013) 2-(Undecyloxy)-ethanol is a major component of the maleproduced aggregation pheromone of Monochamus sutor. Entomol Exp Appl 149:118–127 Paschen MA, Schiff NM, Ginzel MD (2012) Role of volatile semiochemicals in the host and mate location behavior of Mallodon dasystomus (Coleoptera: Cerambycidae). J Insect Behav 25:569–577 Petersen-Silva R, Inácio L, Henriques J, Naves P, Sousa E, Pujade-Villar J (2015) Susceptibility of larvae and adults of Monochamus J Chem Ecol (2016) 42:631–654 galloprovincialis to entomopathogenic fungi under controlled conditions. Intl J Pest Management 61:106–112 Phillips TW, Wilkening AJ, Atkinson TH, Nation JL, Wilkinson RC, Foltz JL (1988) Synergism of turpentine and ethanol as attractants for certain pine-infesting beetles (Coleoptera). Environ Entomol 17:456– 462 Rassati D, Toffolo EP, Battisti A, Faccoli M (2012) Monitoring of the pine sawyer beetle Monochamus galloprovincialis by pheromone traps in Italy. Phytoparasitica 40:329–336 Ray AM, Lacey ES, Hanks LM (2006) Predicted taxonomic patterns in pheromone production by longhorned beetles. Naturwissenschaften 93:543–550 Ray AM, Millar JG, McElfresh JS, Swift IP, Barbour JD, Hanks LM (2009a) Male-produced aggregation pheromone of the cerambycid beetle Rosalia funebris. J Chem Ecol 35:96–103 Ray AM, Swift IP, Moreira JA, Millar JG, Hanks LM (2009b) R)-3hydroxyhexan-2-one is a major pheromone component of Anelaphus inflaticollis (Coleoptera: Cerambycidae). Environ Entomol 38:1462–1466 Ray AM, Žunič A, Alten RL, McElfresh JS, Hanks LM, Millar JG (2011) cis-Vaccenyl acetate, a female-produced sex pheromone component of Ortholeptura valida, a longhorned beetle in the subfamily Lepturinae. J Chem Ecol 37:173–178 Ray AM, Barbour JD, McElfresh JS, Moreira JA, Swift I, Wright IM, Žunič A, Mitchell RF, Graham EE, Alten RL, Millar JG, Hanks LM (2012a) 2,3-Hexanediols as sex attractants and a female-produced sex pheromone for cerambycid beetles in the prionine genus Tragosoma. J Chem Ecol 38:1151–1158 Ray AM, Swift IP, McElfresh JS, Alten RL, Millar JG (2012b) (R)desmolactone, a female-produced sex pheromone component of the cerambycid beetle Desmocerus californicus californicus (subfamily Lepturinae). J Chem Ecol 38:157–167 Ray AM, Arnold RA, Swift I, Schapker PA, McCann S, Marshall CJ, McElfresh JS, Millar JG (2014) (R)-Desmolactone is a sex pheromone or sex attractant for the endangered valley elderberry longhorn beetle Desmocerus californicus dimorphus and several congeners (Cerambycidae: Lepturinae). PloS One 9:e115498 Ray AM, Millar JG, Moreira JA, McElfresh JS, Mitchell RF, Barbour JD, Hanks LM (2015) North American species of cerambycid beetles in the genus Neoclytus share a common hydroxyhexanone-hexanediol pheromone structural motif. J Econ Entomol 108:1860–1868 Rhainds M, Lan CC, King S, Gries R, Mo LZ, Gries G (2001) Pheromone communication and mating behaviour of coffee white stem borer, Xylotrechus quadripes Chevrolat (Coleoptera: Cerambycidae). Appl Entomol Zool 36:299–309 Rhainds M, MacKinnon WE, Porter KB, Sweeney JD, Silk PJ (2011) Evidence for limited spatial spread in an exotic longhorn beetle, Tetropium fuscum (Coleoptera: Cerambycidae). J Econ Entomol 104:1928–1933 Riedl H, Hoying SA, Barnett WW, Detar JE (1979) Relationship of within-tree placement of the pheromone trap to codling moth catches. Environ Entomol 8:765–769 Rodstein J, McElfresh JS, Barbour JD, Ray AM, Hanks LM, Millar JG (2009) Identification and synthesis of a female-produced sex pheromone for the cerambycid beetle Prionus californicus. J Chem Ecol 35:590–600 Rodstein J, Millar JG, Barbour JD, McElfresh JS, Wright IM, Barbour KS, Ray AM, Hanks LM (2011) Determination of the relative and absolute configurations of the female-produced sex pheromone of the cerambycid beetle Prionus californicus. J Chem Ecol 37:114–124 Ryall K, Silk P, Webster RP, Gutowski JM, Meng Q, Li Y, Gao W, Fidgen J, Kimoto T, Scarr T (2015) Further evidence that monochamol is attractive to Monochamus (Coleoptera: Cerambycidae) species, with attraction synergised by host plant volatiles and bark beetle (Coleoptera: Curculionidae) pheromones. Can Entomol 147:564–579 J Chem Ecol (2016) 42:631–654 Sakai T, Nakagawa Y, Takahashi J, Iwabuchi K, Ishii K (1984) Isolation and identification of the male sex pheromone of the grape borer Xylotrechus pyrrhoderus Bates (Coleoptera: Cerambycidae). Chem Lett 1984:263–264 Sakakibara Y, Kikuma A, Iwata R, Yamane A (1998) Performances of four chemicals with floral scents as attractants for longicorn beetles (Coleoptera: Cerambycidae) in a broadleaved forest. J For Res 3:221–224 Sanchez-Husillos E, Etxebeste I, Pajares J (2015) Effectiveness of mass trapping in the reduction of Monochamus galloprovincialis Olivier (Col.: Cerambycidae) populations. J Appl Entomol 10:747–758 Schmeelk TC, Millar JG, Hanks LM (2016) Influence of trap height and bait type on abundance and species diversity of cerambycid beetles captured in forests of east-central Illinois. J Econ Entomol. doi:10.1093/jee/tow102 Schröder F, Fettköther R, Noldt U, Dettner K, König WA, Francke W (1994) Synthesis of (3R)-3-hydroxy-2-hexanone, (2R,3R)-2,3hexanediol and (2S,3R)-2,3-hexanediol, the male sex pheromone of Hylotrupes bajulus and Pyrrhidium sanguineum (Cerambycidae). Liebigs Ann Chem 1994:1211–1218 Schroeder L, Lindelöw Å (1989) Attraction of scolytids and associated beetles by different absolute amounts and proportions of α-pinene and ethanol. J Chem Ecol 15:807–817 Seybold SJ, Ohtsuka T, Wood DL, Kubo I (1995) Enantiomeric composition of ipsdienol: A chemotaxonomic character for North American populations of Ips spp. in the pini subgeneric group (Coleoptera: Scolytidae). J Chem Ecol 21:995–1016 Silk PJ, Sweeney J, Wu J, Price J, Gutowski JM, Kettela EG (2007) Evidence for a male-produced pheromone in Tetropium fuscum (F.) and Tetropium cinnamopterum (Kirby) (Coleoptera: Cerambycidae. Naturwissenschaften 94:697–701 Silva WD, Millar JG, Hanks LM (2016a) (6E,8Z)-6,8-Pentadecadienal, a novel attractant pheromone produced by males of the cerambycid beetles Chlorida festiva and Chlorida costata. J Chem Ecol (in press) Silva WD, Millar JG, Hanks LM (2016b) 10-Methyldodecanal, an attractant pheromone produced by males of the cerambycid species Eburodachrys vittata. PLOS ONE (in press) Slessor KN, King GGS, Miller DR, Winston ML, Cutforth TL (1985) Determination of chirality of alcohol or latent alcohol semiochemicals in individual insects. J Chem Ecol 11:1659–1667 Solomon JD (1995) Handbook 706. Guide to insect borers in North American broadleaf trees and shrubs. USDA For Serv, Washington, DC Suckling DM, Gibb AR, Daly JM, Chen X, Brockerhoff EG (2001) Behavioral and electrophysiological responses of Arhopalus tristis to burnt pine and other stimuli. J Chem Ecol 27:1091–1104 Švácha P, Lawrence JF (2014) Chapter 2.4 Cerambycidae Latreille, 1802. In: Leschen RAB, Beutel RG (eds) Handbook of zoology: Arthropoda: Insecta: Coleoptera, beetles. Vol. 3: Morphology and systematics (Phytophaga). Walter de Gruyter, Berlin/Boston, pp 77–177 Sweeney J, de Groot P, Macdonald L, Smith S, Cocquempot C, Kenis M, Gutowski JM (2004) Host volatile attractants and traps for detection of Tetropium fuscum (F.), Tetropium castaneum (L.) and other longhorned beetles (Coleoptera: Cerambycidae. Environ Entomol 33:844–854 Sweeney JD, Silk PJ, Gutowski JM, Wu J, Lemay MA, Mayo PD, Magee DI (2010) Effect of chirality, release rate, and host volatiles on response of Tetropium fuscum (F.), Tetropium cinnamopterum Kirby, and Tetropium castaneum (L.) to the aggregation pheromone, fuscumol. J Chem Ecol 36:1309–1321 Sweeney JD, Silk P, Hurley JE, Kettela E (2011a) Population suppression of Tetropium fuscum (F.) by pheromone-mediated mating disruption. In: McManus, Gottschalk KA, KW, editors. Proc 22nd U.S. Department of Agriculture interagency research 653 forum on invasive species. USDA Gen Tech Rep NRS-P-52. USDA-Forest Service, Newton Square, p. 91 Sweeney JD, Silk P, Rhainds M, Hurley JE, MacKay W (2011b) Mass trapping for population suppression of an invasive longhorn beetle, Tetropium fuscum (F.) (Coleoptera: Cerambycidae). Proc 22nd U.S. Department of Agriculture interagency research forum on invasive species. USDA Gen Tech Rep NRS-P-52. USDA-Forest Service, Newton Square, p. 92 Sweeney JD, Silk PJ, Hughes C, Lavallée R, Blais M, Guertin C (2013) Auto-dissemination of Beauveria bassiana for control of brown spruce longhorned beetle, Tetropium fuscum (F.), Coleoptera: Cerambycidae. Proc 24nd U.S. Department of Agriculture interagency research forum on invasive species. Forest Health and Technology pub 13–01, USDA-Forest Service, Fort Collins, p 98 Sweeney JD, Silk PJ, Grebennikov V (2014) Efficacy of semiochemicalbaited traps for detection of longhorn beetles (Coleoptera: Cerambycidae) in the Russian far east. Eur J Entomol 111:397–406 Teale SA, Wickham JD, Zhang F, Su J, Chen Y, Xiao W, Hanks LM, Millar JG (2011) A male-produced aggregation pheromone of Monochamus alternatus (Coleoptera: Cerambycidae), a major vector of pine wood nematode. J Econ Entomol 104:1592–1598 Vidal DM, Fonseca MG, Zarbin P (2010) Enantioselective synthesis and absolute configuration of the sex pheromone of Hedypathes betulinus (Coleoptera: Cerambycidae). Tetrahedron Lett 51:6704–6706 Wang Q (1998) Evidence for a contact female sex pheromone in Anoplophora chinensis (Forester) (Coleoptera: Cerambycidae: Lamiinae). Coleop Bull 52:363–368 Wang Q (2002) Sexual selection of Zorion guttigerum Westwood (Coleoptera: Cerambycidae: Cerambycinae) in relation to body size and color. J Ins Behav 15:675–687 Webster RP, Alderson CA, Webster VL, Hughes CC, Sweeney JD (2016) Further contributions to the longhorn beetle (Coleoptera, Cerambycidae) fauna of New Brunswick and Nova Scotia, Canada. ZooKeys 552:109–122 Wickham JD, Harrison RD, Lu W, Guo Z, Millar JG, Hanks LM, Chen Y (2014) Generic lures attract cerambycid beetles in a tropical montane rain forest in southern China. J Econ Entomol 107:259–267 Wickham JD, Lu W, Jin T, Peng Z, Guo D, Millar JG, Hanks LM, Chen Y (2016a) Prionic acid: an effective sex attractant for an important pest of sugarcane, Dorysthenes granulosus (Coleoptera: Cerambycidae: Prioninae. J Econ Entomol 109:484–486 Wickham JD, Lu W, Long-Wa Z, Chen Y, Zou Y, Hanks LM, Millar JG (2016b) Likely aggregation-sex pheromones of the invasive beetle species Callidiellum villosulum, and the related Asian species Allotraeus asiaticus, Semanotus bifasciatus, and Xylotrechus buqueti (Coleoptera: Cerambycidae). J Econ Entomol (in press) Wickham JD, Millar JG, Hanks LM, Zou Y, Wong JCH, Harrison RD, Chen Y (2016c) (2R,3S)-2,3-octanediol, a female-produced sex pheromone of Megopis costipennis (Coleoptera: Cerambycidae: Prioninae). Environ Entomol 45:223–228 Wong JCH, Hanks LM (2016) Influence of fermenting bait and vertical position of traps on attraction of cerambycid beetles to pheromone lures. J Econ Entomol (in press) Wood DL (1982) The role of pheromones, kairomones, and allomones in the host selection and colonization behavior of bark beetles. Annu Rev Entomol 27:411–446 Yamasaki T, Sato M, Sakoguchi H (1997) (−)-Germacrene D: masking substance of attractants for the cerambycid beetle, Monochamus alternatus. Appl Entomol Zool 32:423–429 Yasui H (2009) Chemical communication in mate location and recognition in the white-spotted longicorn beetle, Anoplophora malasiaca (Coleoptera: Cerambycidae). Appl Entomol Zool 44:183–194 Yasui H, Yasuda T, Fukaya M, Akino T, Wakamura S, Hirai Y, Kawasaki K, Ono H, Narahara M, Kousa K, Fukuda T (2007) Host plant chemicals serve intraspecific communication in the white-spotted 654 longicorn beetle, Anoplophora malasiaca (Thomson) (Coleoptera: Cerambycidae). Appl Entomol Zool 42:255–268 Yasui H, Akino T, Fukaya M, Wakamura S, Ono H (2008) Sesquiterpene hydrocarbons: kairomones with a releaser effect in the sexual communication of the white-spotted longicorn beetle, Anoplophora malasiaca (Thomson) (Coleoptera: Cerambycidae). Chemoecology 18:233–242 Yatagai M, Makihara H, Oba K (2002) Volatile components of Japanese cedar cultivars as repellents related to resistance to Cryptomeria bark borer. J Wood Sci 48:51–55 Zarbin PHG, Fonseca MG, Szczerbowski D, Oliveira ARM (2013) Biosynthesis and site of production of sex pheromone components of the cerambycid beetle, Hedypathes betulinus. J Chem Ecol 39:358–363 J Chem Ecol (2016) 42:631–654 Zhang A, Oliver JE, Aldrich JR, Wang B, Mastro VC (2002) Stimulatory beetle volatiles for the Anoplophora glabripennis (Motschulsky). Z Naturforsch 57c:553–558 Zou Y, Millar JG, Blackwood JS, Duzor RV, Hanks LM, MongoldDiers JA, Wong JCH, Ray AM (2015) (2S,4E)-2-hydroxyoct-4en-3-one, a male-produced attractant pheromone of the cerambycid beetle Tylonotus bimaculatus. J Chem Ecol 41: 670–677 Zou Y, Rutledge CF, Nakamuta K, Maier CT, Hanks LM, Richards AB, Lacey ES, Millar JG (2016) Identification of a pheromone component and a critical synergist for the invasive beetle Callidiellum rufipenne (Coleoptera: Cerambycidae). Environ Entomol 45:216– 222