Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Paolo Macchiarini wikipedia , lookup
Development of the nervous system wikipedia , lookup
Sexual reproduction wikipedia , lookup
Cell culture wikipedia , lookup
Cell encapsulation wikipedia , lookup
Somatic cell nuclear transfer wikipedia , lookup
Regeneration in humans wikipedia , lookup
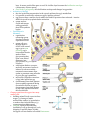
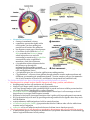
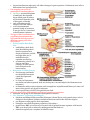
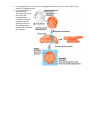
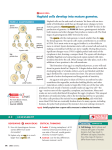
Stephanie Bobbitt Chapter 47) Animal Development p. 998-1017 - The Stages of Early Embryonic Development o From egg to organism, an animal’s form develops gradually: the concept of epigenesis preformation: old idea that the embryo already had everything it needed, and development just meant it got bigger; also included the idea that the embryo contains all of its descendents epigenesis: the form of an animal comes gradually from a formless egg development includes cell division and differentiation and morphogenesis (process where an animal takes shape); overall development process is epigenesis o Fertilization activates the egg and brings together the nuclei of sperm and egg purpose of fertilization is to combine haploid sets of chromosomes into one diploid cell when sperm makes contact with egg’s surface, metabolic reactions are initiated in the egg to start embryonic development The Acrosomal Reaction acrosomal reaction (in sea urchins): sperm cell meets jelly coat around an egg vesicle at sperm tip called acrosome discharges contents by exocytosis; reaction releases hydrolytic enzymes that enable the acrosomal process structure to penetrate the jelly coat lock-and-key recognition ensures eggs are fertilized by sperm of same species membranes of the sperm and egg fuse together ion channels open in egg cell’s plasma membrane allows sodium ions to flow into cell fast block to polyspermy: aka depolarization that was described above; prevents more than one sperm from fusing with the egg’s membrane The Cortical Reaction cortical reaction: series of changes in cortex of egg cytoplasm fusion of egg and sperm trigger egg’s ER to release calcium into cytosol; released calcium causes a change in cortical granules cortical granules fuse with plasma membrane and release contents into space between membrane and vitelline o layer causes perivitelline space to swell vitelline layer becomes the fertilization envelope (resists entry of more sperm) slow block to polyspermy: this fertilization envelope and changes in egg surface Activation of the Egg the rise in calcium concentration in the cytosol quickens the egg’s metabolism it is possible to artificially activate an egg by injecting calcium egg does not have a nucleus, but it makes new kinds of proteins when activated – inactive mRNA is stored in cytoplasm before activation sperm nucleus swells and merges with egg nucleus, then DNA synthesis begins Fertilization in Mammals capacitation: enhancing sperm function in female reproductive tract sperm cell goes through layer of follicle cells around egg to get to zona pellucida: egg’s extracellular matrix made of three glycoproteins that form cross-linked filaments (one glycoprotein, ZP3, is a sperm receptor) when sperm binds to receptor molecule, acrosome of sperm is induced to release contents – enzymes from acrosome allow sperm to penetrate zona pellucida to get to the egg’s membrane microvillia (fingerlike extensions of egg) take sperm into egg chromosomes from both gametes share a spindle apparatus during first mitotic division of zygote – chromosomes from both parents do not come together until after this first division Cleavage partitions the zygote into many smaller cells building animal’s body occurs in three stages: (1) cleavage to make multicellular embryo (2) gastrulation to make a three-layered embryo (3) organogenesis to make organs cleavage: rapid cell division after fertilization; cells go through cell cycle, skipping G1 and G2 phases; divides the zygote cell into smaller cells called blastomeres yolk: stored nutrients; influences pattern of cleavage vegetal pole has a bigger concentration of yolk than the opposite pole called animal pole (see figure on previous page) plasma membrane and cortex rotate to where sperm entered exposes a region of the cytoplasm called gray crescent, which marks the dorsal side of embryo yolk slows down cell division, so cleavage occurs faster in animal hemisphere morula: solid ball of cells produced by continued cleavage blastocoel: fluid-filled cavity forms in morula – making hollow ball called blastula meroblastic cleavage: incomplete division of a yolk-rich egg (ex: in birds) holoblastic cleavage: complete division of eggs with small or moderate amount of yolk (ex: in frogs) Gastrulation rearranges the blastula to form a three-layered embryo with a primitive gut gastrulation: major rearrangement of cells of blastula; involves changes in cell motility, cell shape, and cellular adhesion to other cells and molecules gastrula: three-layered embryo made by gastrulation ectoderm: outer layer of gastrula; later develops into nervous system and epidermis endoderm: lines embryonic digestive tract; later develops into digestive tract and related organs mesoderm: partly fills space between ectoderm and endoderm; later develops into organs like kidney, heart, muscles, etc. sea urchin gastrulation begins at vegetal pole where some cells detach from blastula wall and go to blastocoel as cells called mesenchyme cells; other cells flatten to make a plate that buckles inward (process is called invagination) buckled plate’s cells are rearranged that transforms the invagination into a deep pouch called the archenteron (open end of this is called the blastopore and will later become the anus) an opening forms at the other end of archenteron which will be the mouth end of the digestive tube right figure shows sea urchin gastrulation figure on next page shows frog gastrulation frog gastrulation starts with a small crease on one side of blastula that is caused by invagination that becomes the dorsal side of blastopore (called dorsal lip; forms where gray crescent was in zygote) involution occurs (cells on embryo surface roll into embryo interior and move away from blastopore cells or organized into the mesoderm and endoderm layers yolk plug made of big cells move inward and cells on outside develop into the ectoderm) In organogenesis, the organs of the animal body form from the three embryonic germ layers organogenesis: regions of the layers develop into organs figure on next page shows early organogenesis in frogs notochord: made from dorsal mesoderm that condenses, eventually become the vertebral discs neural tube starts off as a plate of dorsal ectoderm which will fold inward to become the neural tube, which becomes the central nervous system (brain and spinal cord) somites: blocks that some mesoderm separates into; cells from these give rise to vertebrae and make muscles associated with axial skeleton neural crest: band of cells that migrate to parts of embryo and form pigment cells of skin, skull, teeth, adrenal glands, etc. o o o Amniote embryos develop in a fluidfilled sac within a shell or uterus all vertebrate embryos need an aqueous environment in order to develop amniotes: vertebrates with amnions (sac of fluid) Avian Development see figure on next page blastodisc: cap of cells made by early cleavage divisions blastomeres sort into the epiblast and hypoblast layers gastrulation involves cells of the epiblast to move to the middle of the blastodisc, detach, and move toward yolk – this movement makes groove the primitive streak (marks where anterior-posterior axis will be) all cells that make the embryo come from the epiblast; hypoblast directs formation of primitive streak and its cells later make a sac around yolk and a stalk that connects the yolk to the embryo borders of embryonic disc fold down to pinch embryo into a three-layered tube joined in the middle to the yolk tissues outside the embryo develop into four extraembryonic membranes for further development (four are yolk sac, amnion, chorion, and allantois – see figure on next page) yolk sac digests yolk, amnion protects embryo from drying out, chorion cushions embryo against mechanical shocks, and allantois is a disposal sac for uric acid Mammalian Development cleavage in mammals is slower compaction: process that tightly sticks cells together; involves making new proteins on cell surface like cadherins see diagram on next page of development (1) a cluster of cells called the inner cell mass protrudes into blastocyst (stage where embryo has over 100 cells surrounding a central cavity); trophoblast surrounds the cavity; trophoblast + mesoderm = fetal part of placenta (2) trophoblast secretes enzymes that let the blastocyst go through the endometrium and expands to extend fingerlike projections into maternal tissue; blastocyst makes disc with epiblast (upper layer of cells) and hypoblast (lower) (3) trophoblast gives rise to chorion; epiblast starts making amnion (4) gastrulation – cells move from epiblast through primitive streak to make mesoderm and endoderm; 4 extraembryonic membranes formed – chorion, amnion, yolk sac (for mammals site of early formation of blood cells), and allantois (incorporated into umbilical cord) The Cellular and Molecular Basis of Morphogenesis and Differentiation in Animals o Morphogenesis in animals involves specific changes in cell shape, position, and adhesion morphogenesis involves cell movement only in animals changes in cell shape usually involve reorganizing cytoskeleton cells crawl through embryo with cytoskeletal fibers to extend and retract cellular protrusions that are usually flat sheets (lamellipodia) or spikes (filopodia) convergent extension: morphogenic movement where a tissue layer’s cells rearrange so the cell sheet narrows (converges) and elongates (extends) ECM may be involved in convergent extension since it guides cells in morphogenetic movements EC glycoproteins help cells move by providing anchorage and hold cells together when they reach their destination certain substances inhibit migration of cells in certain directions cell adhesion molecules (CAMs): glycoproteins that bind to CAMs on other cells for stable tissue structure and cell migration cadherins: cell-to-cell adhesion molecule that need calcium ions to function correctly o The developmental fate of cells depends on cytoplasmic determinants and cell-cell induction: a review in many animal species, heterogeneous distribution of cytoplasmic determinants in unfertilized egg leads to regional differences in early embryo - interactions between embryonic cells induce changes in gene expression ultimately cause cells to differentiate into specialized cells Fate mapping can reveal cell genealogies in chordate embryos fate maps: territorial diagrams of embryonic development – shows which parts of embryo will be derived from the parts of the zygote or blastula early cells make specific tissues of the older embryo a cell’s developmental potential (range of structures it can give rise to) is restricted as development continues The eggs of most vertebrates have cytoplasmic determinants that help establish the body axes and differences among cells of the early embryo Polarity and the Basic Body Plan establishing a basic body plan (determining where anterior-posterior axis, dorsal-ventral axis, and left and right sides are) is needed before the organism can develop in mammals, polarity isn’t obvious until after cleavage; in other species, basic instructions are known earlier Restriction of Cellular Potency first cleavage could make two identical blastomeres (equal developmental potential) ex: first two blastomeres are similar and when separated, both can develop into a normal tadpole (aka blastomeres are totipotent) in mammals, embryos stay totipotent until arranged into trophoblast and blastocyst’s inner cell mass; other species, only zygote is totipotent tissue-specific fates of cells in late gastrulae are fixed Inductive signals drive differentiation and pattern formation in vertebrates induction: one group of cells influences development of another The “Organizer” of Spemann and Mangold Spemann and Mangold discovered that blastopore’s dorsal lips in early gastrula plays a role in embryonic development (initiation inductions that form neural tube and other organs) see diagram on next page for their experiment blastopore was called the primary organizer of the embryo growth factor BMP-4 is active only on ventral side of gastrula because organizer cells inactive BMP-4 on dorsal side or embryo by making proteins that bind to it inductions typically are a sequence of inductive steps o o o (1) transplant piece of dorsal lip of a nonpigmented newt gastrula to ventral side of early gastrula of pigmented newt (2) recipient embryo made a second notochord and neural tube where the transplantation occurred; cells in the recipient were still made from itself, so the nonpigmented embryo had organized the recipient