Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
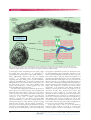
Molecular survival strategies of Borrelia burgdorferi Review Molecular survival strategies of the Lyme disease spirochete Borrelia burgdorferi Sunit Kumar Singh and Hermann Josef Girschick Lyme disease is a tick-transmitted disease caused by the spirochete Borrelia burgdorferi. The bacterium adopts different strategies for its survival inside the immunocompetent host from the time of infection until dissemination in different parts of body tissues. The success of this spirochete depends on its ability to colonise the host tissues and counteract the host’s defence mechanisms. During this process borrelia seems to maintain its vitality to ensure long-term survival in the host. Borrelia’s proteins are encoded by plasmid and chromosomal genes. These genes are differentially regulated and expressed by different environmental factors in ticks as well as in the mammalian host during infection. In addition, antigenic diversity enables the spirochete to escape host defence mechanisms and maintain infection. In this review we focus on the differential expression of proteins and genes, and further molecular mechanisms used by borrelia to maintain its survival in the host. In light of these pathogenetic mechanisms, further studies on spirochete host interaction are needed to understand the complex interplay that finally lead to host autoimmunity. Lancet Infect Dis 2004; 4: 575-83 Lyme disease is caused by a group of related tick-borne spirochetes classified as Borrelia burgdorferi sensu lato (including B burgdorferi sensu stricto, Borellia afzelii, and Borellia garinii). They are transmitted by ticks of the Ixodes species including Ixodes ricinus in Europe and Asia, Ixodes persulcatus in eastern Europe and Asia, and Ixodes scapularis and Ixodes pacificus in North America. After attachment, ticks induce local inflammatory and immunological responses against the components present in saliva of the tick.1 To overcome the host immune responses, tick saliva also contains substances that suppress or divert these responses.1,2 In addition to these vector/host interactions, B burgdorferi undergoes dynamic changes within the vector during both acquisition and transmission, and likewise within vertebrate hosts after transmission. To survive in nature, B burgdorferi must sense its environment and synthesise appropriate proteins for interactions with the different tick and mammalian tissues. Although much progress has been made in characterisation of the host immune reaction, spirochetal factors responsible for infectivity, immune evasion, and disease pathogenesis remain largely obscure. Several studies have demonstrated that different borrelia isolates are highly heterogeneous in their phenotypic and genotypic properties. A high degree of Infectious Diseases Vol 4 September 2004 variability exists in plasmid and protein profiles of different B burgdorferi isolates.3 In this review, the role of major proteins, their differential expression in different environments, and protective molecular strategies adopted by B burgdorferi to cope with the host’s immunological attack are reviewed and discussed. Biology and genomic organisation B burgdorferi is a microaerophilic, wavelike shaped bacterium. It is generally 20–30 m in length and 0·2–0·5 m in width. In addition to a typical bacterial plasma membrane, it has an outer lipid membrane termed an outer membrane sheath (figure 1). The space between the protoplasmic cell cylinder and the outer membrane sheath is called the periplasm, which contains the flagella.4,5 The protoplasmic cylinder contains the cytoplasm. The genome size is relatively small, about 1·5 mb.6 Borrelia has a linear chromosome of 950 kb in length, and at least 21 extra chromosomal DNA elements or plasmids.7 Since genes for biosynthetic reactions are absent, growth of B burgdorferi in vitro needs tissue culture medium supplemented with mammalian serum.8 In addition to the linear chromosome, B burgdorferi isolate B31 contains 12 linear plasmids (lp) (lp5, lp17, lp21, lp25, lp28-1, lp28-2, lp28-3, lp28-4, lp36, lp38, lp54, and lp56), and nine circular plasmids (cp) (cp9, cp26, cp32-1, cp32-3, cp32-4, cp32-6, cp32-7, cp32-8, and cp32-9).8 B burgdorferi has the largest number of plasmids known for any bacterium. Genes related with pathogenicity including outer surface protein genes (osp) are primarily located on the plasmids. Outer surface proteins The borrelia strains possess several outer surface lipoproteins (Osps). The first major Osps isolated were termed alphabetically—namely OspA,9 OspB, OspC,10 OspD,11 OspE, and OspF.11–16 Afterwards these proteins were termed according to their function or by their molecular mass—eg, decorin binding protein, or lp6·6 for a 6·6 kDa protein. Osps seem to interact with cellular and interstitial components of the tick and the mammalian tissue. During feeding of the tick in addition to migration and transmission SKS is at the Department of Paediatric Rheumatology, Children’s Hospital, University of Würzburg, Germany. Correspondence: Dr Hermann Girschick, Paediatric Rheumatology, Children’s Hospital, University of Würzburg, Josef-Schneider-Str 2, 97080 Würzburg, Germany. Tel +49 931 201 27728; fax +49 931 201 27242; email [email protected] http://infection.thelancet.com 575 For personal use. Only reproduce with permission from Elsevier Ltd. Review Molecular survival strategies of Borrelia burgdorferi Outer membrane Multiple wavelike-shaped periplasmic flagella Transmission electron micrographs of Borrelia burgdorferi Protoplasmic cylinder Embedded proteins Outer membrane Periplasmic flagella Protoplasmic cylinder FlaB FlaA Anchor of flagella in cell membrane Figure 1. Ultrastructural morphology of B burgdorferi electron micrographs and schematic line drawings illustrate the structural morphology of B burgdorferi. The spirochetes consist of a protoplasmic cylinder covered by the cell membrane. A second outer membrane covers the periplasm containing the flagella. Flagella consist of two major proteins, flagellin A and flagellin B (FlaA and FlaB). of B burgdorferi, many borrelial lipoproteins (OspC, OspE, and OspF) have been shown to be upregulated17 or downregulated by B burgdorferi (OspA, OspB, and lp6·6).18 Many differentially expressed proteins are plasmid encoded.18,19 B burgdorferi lacks lipopolysaccharide,20 suggesting that Osps, which possess a potent stimulatory capacity on mammalian cells,21 are in part responsible for the inflammation associated with an infection. Among these are the OspE, OspF, and the outer surface E/F-like leader peptide (Elp) paralogues,18,22 which together have been termed OspE/F-related proteins (Erps).7,8,18 The most studied B burgdorferi membrane lipoprotein is OspA expressed on borrelia in resting (unfed) nymphal and adult ticks and in-vitro culture.23 OspA serves as an anchor for the spirochete in the midgut of the unfed tick. Digestion of blood is done intracellularly by tick gut cells using endocytosis as the mode of uptake. OspA-mediated adherence of spirochetes to the surface of gut cells might prevent internalisation of spirochetes during initial blood meal digestion (figure 2).6 Host antibody responses to OspA might therefore interfere with the midgut colonisation.24 Some reports show that spirochetes in the midgut of feeding ticks start to downregulate OspA.17,25 Therefore, suppression of OspA expression during the tick’s blood meal could help 576 B burgdorferi’s detachment from the gut. Transversion into the haemolymphatic fluid, subsequent migration to the salivary glands, and ultimately transmission to the vertebrate host could be helped.26 The role of OspA in transmission of the pathogen is further illustrated by a report demonstrating, that mice immunised with OspA were not protected from infection when they were challenged with skin implants of infected mice containing B burgdorferi. This study indirectly suggested that host adapted spirochetes no longer produced OspA.27 This finding was confirmed by in-vivo culture experiments on B burgdorferi with dialysis membrane chambers.28 During the blood meal, the population of the spirochetes becomes more heterogeneous, because Osp expression is quite variable in one single strain. After transmission, the role of OspA expressed on B burgdorferi in the mammalian host is not known. A correlation between the presence of OspA specific antibodies and T lymphocytes with primary stages of Lyme borreliosis as well as with chronic antibiotic-resistant Lyme arthritis have been shown. These studies indirectly suggested that OspA might again be present at some stages inside the mammalian host.29 Another Osp seems to have a role in spirochete/host interaction. OspC synthesis was shown to correlate with borrelia’s migration from the tick midgut to the salivary Infectious Diseases Vol 4 September 2004 http://infection.thelancet.com For personal use. Only reproduce with permission from Elsevier Ltd. Molecular survival strategies of Borrelia burgdorferi Mammalian host OspA OspC Tick OspC Midgut pC Os OspA OspA Erp Salivary glands Erp and CRASPs bind complement Factor H FHL-1 regulators Factor H Skin and FHL-1 Factor H FHL-1 Complement C3 C3 C3 C3 components are C3 recruited to FactorH/FHL-1 C3 and subsequently C3 inactivated C3 Spirochetal complement resistance CRASPs Figure 2. Regulation of gene expression in B burgdorferi. Borrelia residing in the midgut of ticks show upregulated outer surface protein, Osp A and downregulated OspC expression on their surface. OspA is of major relevance for the colonisation of the tick midgut. During tick feeding OspA is downregulated, while OspC is upregulated when the spirochetes move from the midgut into the salivary glands. Inside the host OspA is downregulated, whereas OspC is upregulated, but shows a complex time and location dependent expression pattern shown by three arrows indicating variable upregulation Erp genes generally are upregulated in the mammalian host. Erp proteins, in addition to complement acquiring surface proteins (CRASPs), are of importance to inhibit complementmediated killing by the host’s innate immune system. Using Erp and CRASP proteins, spirochetes are able to bind complement regulatory proteins, Factor H and FHL-1 present in the host’s serum to their own surface. Subsequently, complement components C3b, c, and d are bound to Factor H and FHL-1 and are inactivated. Impaired activation of the complement cascade in the alternative pathway represents an important mechanism in spirochete transmission, and propagation of the infection. gland, suggesting a role in transmission.30 OspC is not expressed by most spirochetes in unfed ticks.31 The synthesis of OspC is upregulated by B burgdorferi in the midgut during tick feeding.17,32,33 Once the spirochetes reach the tick’s salivary gland, they begin to downregulate OspC production again (figure 2). Antibodies against OspC block the movement of spirochetes from the gut to the tick’s salivary glands.30 Even though most bacteria evading the salivary glands and entering the host did not produce OspC any more, small numbers of bacteria still could have OspC on their surface. This possibility might be sufficient to stimulate the early OspC antibody response that is regularly reported in early stages of Lyme disease.34 Each B burgdorferi spirochete contains a single ospC gene copy.35 By contrast, individual spirochetes carry many Erp operons on the cp32 plasmid.36,37 As many as ten different cp32 family members have been identified in clonal populations of B burgdorferi.38 A single bacterium can express its entire Erp repertoire simultaneously.39 A variety of names were used to describe erp genes including ospE, ospF, elp, p21, bbk2·10, bbk2·11, pG, and upstream homology box (UHB)-flanked genes.40,41 The last term was introduced Infectious Diseases Vol 4 September 2004 Review because these families share a common feature. They are flanked at the 5 end by a highly conserved sequence called UHB.42 The erp genes seen within an individual bacterium often exhibit a considerable range of sequence variation.7 Hefty and colleagues39 classified the Erp polypeptides of B burgdorferi strain 297 into three separate groups: OspErelated orthologues, OspF-related orthologues, and Elps, which contain OspE/F-like leader peptides. Erp proteins are located on the bacterial outer surface and have an important role in complement resistance of B burgdorferi .43–45 Strategies of antigenic variation and immune escape Immunodominant antigens are commonly used to distinguish strains of a pathogen. These antigens can vary from one strain to another resulting in strain-specific immune responses of vertebrate hosts.46 Different strains of B burgdorferi express different OspC proteins in rodents.46 Even though each B burgdorferi spirochete contains one single ospC gene copy, sequences are diverse and immune responses against OspC seem to influence selection of the pathogen.46 This diversity of an immunodominant antigen between strains is called antigenic variation. The adaptive immune system of infected vertebrates mounts an immune response against the original infecting serotype, but this specific response might be ineffective against new emerging variants.46 Genetic recombination at the vls gene locus Another mechanism that could contribute to B burgdorferi survival is the recombination at the variable major protein like sequence (vls) gene locus.47,48 The vlsE (vls expression site) encodes a surface protein of 34 kDa with three defined domains: two constant regions at the amino and carboxyl termini and an internal variable segment.48,49 It has been postulated that infection induces sequence changes and thus alters the antigenic properties of the vlsE, and leads to immune evasion through antigenic variation. The generation of new antigenic variants is thought to occur through the exchange of DNA cassettes by the process of recombination. This recombination could potentially help spirochetes to escape antibody-mediated defence against the vlsE protein variants arising during infection.47,48 The vls gene cluster consists of the single vlsE located near the right telomere of lp28-1 and 15 silent cassettes upstream.48,50 The infectivity of B burgdorferi strain B31 diminished after the loss of vlsE bearing lp25 and lp28-1 plasmids51,52 suggesting that these plasmids encode virulence factors. Unidirectional gene conversion events between the silent cassettes and the vlsE gene generate new vlsE variants.53 Tick feeding stimulates the recombination process at the vlsE locus.34 The three-dimensional structure of the vlse outer surface lipoprotein, VlsE, shows that the most distal outer part of the protein consists primarily of variable regions. The variable regions on the outermost surface could thus mask the invariant regions of the protein. By producing a myriad of VlsE variants that do not bind effectively to anti-VlsE antibodies elicited by previous versions of the protein, B burgdorferi can stay one step ahead of the mammalian http://infection.thelancet.com 577 For personal use. Only reproduce with permission from Elsevier Ltd. Review Molecular survival strategies of Borrelia burgdorferi immune response.54 VlsE gene expression is downregulated during tick feeding and then upregulated upon establishment of infection in the mammalian host.55 Lateral gene transfer Bacteriophages have a role in naturally occurring lateral gene transfer. They have been reported in association with many spirochetes,56 including Borrelia species.57 Eggers and coworkers reported on a bacteriophage, which packaged the 32-kb circular plasmids (cp32, table) of the B burgdorferi genome, which was used to transduce cp32 to other borrelia strains. Transfer of cp32 DNA via phage particles from one spirochete to another could be an important mechanism of pathogenesis. In addition, lateral gene transfer between ospC genes within the species, and between B burgdorferi sensu stricto, B garinii, and B afzelii has been reported.58 Genes for ospC from eight isolates of Borrelia valaisiana were cloned and compared with the ospC from B burgdorferi sensu lato species.59 Frequent ospC gene transfer was shown between B valaisiana and B garinii, as well as B valaisiana and B afzelii.60 Simultaneous infection of B valaisiana and B garinii or B afzelii in ticks and birds61 could explain the high frequency of gene exchange between different Borrelia species. A high frequency of gene exchange could enable the spirochetes to react effectively to environmental selection pressures. Lipoprotein polymorphism Molecular changes in genes encoding surface antigens could provide sufficient antigenic variation to support persistence of the pathogen.62 OspA shows little heterogeneity within species, supposedly because it is not under immune selection pressure by vertebrate hosts.63 OspC as an antigen is, however, much more polymorphic than OspA.16,63 OspC diversity can be prompted by several mechanisms, including host-stimulated immunological selection, gene transfer, intragenomic gene recombination, and effects of environmental constraints. Of these factors, selection pressure might be the major force in maintaining the variation of OspC.64,17 To escape selective immunological pressure caused by the host, individual spirochetal strains can change their ospC gene sequences, and thus cause variation of the ospC gene. By contrast with ospC genes, erp gene sequences inside individual strains remain considerably constant. Erp loci of borrelia re-isolated from laboratory mice after 1 year of infection were identical to those of the inoculated bacteria. This finding shows that the erp gene sequences remain considerably stable during the infection of an individual mammal.38,41 However, comparative sequence analysis of ospE gene family members from various borrelia strains show the existence of hypervariable regions, indicating that these sequences have been influenced to some extent by different molecular events like recombinations or mutations acting in an evolutionary time frame.42,65 Thus, the high variation in protein sequence translated from erp genes, compared with ospC, is based on the high number of different but related genes arranged on up to at least ten different cp32 plasmids. There are several different gene families in Lyme disease spirochetes that enhance the antigenic diversity during infection in their natural reservoirs or in the human host, either by genetic variation (eg, ospC), or by recruiting a variety of closely related genes out of one family (eg, erp). Continual genomic change in its gene repertoire, differential expression of gene families,66,67 and the diversity introduced by lateral transfer of plasmids15 could provide the Lyme disease spirochetes with sufficient genetic and antigenic diversity to be able to react to the host’s immune attack and thus maintain chronic infection. Kinetics of expression, location, and function of B burgdorferi genes Gene/Gene product OspA lp54 Predominant expression Function in location Tick Anchors spirochetes to tick midgut OspC cp26 Tick/mammal Migration of spirochete from tick midgut to salivary gland ErpA/I/N, ErpP, ErpC, and ErpX DbpA DbpB p66 vls BgP CRASP cp32 Mammal lp54 lp54 Chromosome lp28-1 Chromosome Lp54 and cp32 Mammal Mammal Mammal Mammal Mammal Mammal oppA-IV oppA-V Bdr D cp26 lp54 cp32 Mammal Mammal Tick/mammal Bdr E cp32 Tick/mammal spoT Chromosome Tick/mammal? Inactivation of the host alternative complement pathway Binds decorin Binds decorin Binds integrin Promotes spirochetal immune evasion Binds glycosaminoglycans Blocks complement activation of the host Environmental adaptation Environmental adaptation Sensing and transducing environmental signals Sensing and transducing environmental signals Important role during fatty acid starvation ? 578 Gene location Kinetics/pathogenetic mechanisms Expressed in unfed tick and downregulated during tick feeding Upregulated during migration from tick midgut to salivary gland but downregulated during dissemination in mammalian host Expressed/upregulated in mammalian host/infection Responsible for colonisation of the host Responsible for colonisation of the host Responsible for colonisation of the host Recombination leads to antigenic variation Responsible for colonisation of the host Upregulated in mammalian host Upregulated with temperature Unclear Expressed in fed and unfed ticks, induced by serum deprivation Expressed in fed and unfed ticks, induced by serum deprivation Upregulated during fasting/fatty acid starvation Infectious Diseases Vol 4 September 2004 http://infection.thelancet.com For personal use. Only reproduce with permission from Elsevier Ltd. Molecular survival strategies of Borrelia burgdorferi Complement regulator-acquiring surface proteins (CRASPs) The complement system of the host’s innate immunity is important in the first-line defence against invading microbes. Complement activation destroys bacterial pathogens by coating them with opsonising molecules (C1q, C3b, and iC3b) after entry in the human host. This process can be initiated by antibodies or antibody-independent mechanisms via direct activation of the classic or alternative pathways. Assembly of the complement membrane attack complex usually leads to lysis of Gram-negative bacteria; however, microbes have evolved effective means to escape this attack. Microorganisms can evade the destructive effects of this powerful defence system by binding host complement regulators of the alternative pathway directly to their surfaces, leading to inhibition of the complement activation cascade (figure 2).68,69 B burgdorferi expresses up to five distinct proteins on its surface (CRASP-1, CRASP-2, CRASP-3, CRASP-4, and CRASP-5), which display different and unique binding properties for host immune regulators.70 CRASPs bind to two major human fluid-phase complement regulators of the alternative pathway, factor H and factor H like protein1 (FHL-1/reconnectin). This binding can inactivate newly formed C3b (figure 2). CRASPs of borrelia have been given names according to the species of origin, Ba for B afzelii and Bb for B burgdorferi. Both B afzelii and B burgdorferi contain proteins of identical binding profiles.69 BaCRASP-1, BbCRASP-1, and BbCRASP-2 bind FHL-1/reconectin strongly but show weak interaction with factor H.69 By contrast, both of the B afzelii proteins (BaCRASP-4 and BaCRASP-5) and the three B burgdorferi proteins (BbCRASP-3, BbCRASP-4, and BbCRASP-5) bind to factor H, but not FHL-1/reconectin.69 CRASP proteins do orient amino complementary domains of attached FHL1/reconectin and factor H in such a way that they do not loose their complement regulatory function.71 This function prevents the formation of cytolytic complement activation products on the borrelial surface. Erp proteins are also able to bind complement inhibitory factor H (figure 2). Like CRASPs, Erp proteins (OspE, OspF,62,72 Elps,22 p21,73 ErpA, and ErpP73) can interact with complement regulators.74 Each Erp protein exhibits different relative affinities for the complement inhibitors of various potential animal hosts. The presence of many Erp proteins on the surface can allow a single B burgdorferi bacterium to resist complement-mediated killing in any of the potential hosts that it might infect.75 Kurtenbach and co-workers76 reported that B burgdorferi also binds to other complement control proteins in the tick midgut and thus is protected against complement-mediated lysis. Complement resistance might therefore represent one major pathogenic factor favouring spirochete transmission to the host. Of note, exposure to the host’s complement cascade takes place not only in the gut, but also in the subsequent steps of spirochete transmission into the host. Role of tissue localisation of B burgdorferi in survival and immune escape Persistence of B burgdorferi in bradytrophic tissue, deep invaginations of the cell membrane, or even in the cytosol of Infectious Diseases Vol 4 September 2004 Review local joint cells, might contribute to the chemotherapeutic resistance and interference with immune clearance.77 Peripheral blood fibrocytes are a circulating leucocyte subpopulation.78 B burgdorferi was shown to bind to fibrocytes in vitro, and to reside within deep invaginations on the cell surface without being phagocytosed.63,79 This process of hiding could be a mechanism of protection for B burgdorferi avoiding the attack of the immune system.63 B burgdorferi can be recovered a long time after initial infection, even in ceftriaxone-treated fibroblast cultures, suggesting that it can resist eradication by host defence mechanisms as well as antibiotics.80 We have reported that human synovial cells could be a target for intracellular persistence of B burgdorferi after the bacteria have passed through the endothelial barriers.81,82 Ma and colleagues83 reported the intracellular localisation of B burgdorferi within human endothelial cells. They likewise reported that internalisation of B burgdorferi into endothelial cells seemed to be mediated by a constitutively expressed endothelial receptor, but not by adhesion molecules.83 Montgomery and colleagues84 also reported that nonopsonised B burgdorferi could enter macrophages rapidly, resulting mainly in degradation but occasionally in apparent intracellular persistence. Persistence of spirochetes within macrophages might provide a possible reservoir for chronic or recurrent Lyme disease in human beings.84 In an experimental murine Lyme disease model spirochetes have been detected intracellularly in cardiac myocytes.85 Even though there is experimental evidence suggesting intracellular persistence from in vitro and in vivo studies, reports on patients and human tissues supporting these findings are very rare.86 On the other hand, it has been reported several times that borrelia can be detected extracellularly in bradytrophic tissue like ligaments and collagen tissue of Lyme disease patients, or in vitro models.85,87–89 Molecular analysis for B burgdorferi DNA in synovial fluid or tissue samples revealed a higher detection rate in the tissue samples.90 PCR of synovial tissue biopsies, however, does not give information on a potential intracellular or extracellular location. In addition, there are reports of extracellular matrix localisation of B burgdorferi in tissues other than joints.91,92 Thus, in conclusion, B burgdorferi in general seems to reside extracellularly. So far, intracellular location or persistence seems to be a rare event. However, because of the scarcity of spirochetes in affected tissues, and the difficulty in detecting them, further studies are needed to assess modes of spirochetal long-term persistence. Adaptive responses of B burgdorferi to variable environmental factors During ticks feeding, the spirochetes encounter environmental changes caused by the influx of mammalian blood into the midgut. Spirochetes encounter a rise in temperature and a fall in pH from 7·4 to 6·8.93 At the time of tick feeding the spirochetes change their surface expression of the outer surface lipoproteins,17,94 as shown in figure 2. The synthesis of OspC,17,95 Bbk32, and Bbk50 (B burgdorferi fibronectin binding protein) proteins94 is induced specifically during the course of the tick feeding. To survey more broadly B burgdorferi’s adaptation, various in-vitro model systems have been used to http://infection.thelancet.com 579 For personal use. Only reproduce with permission from Elsevier Ltd. Review Molecular survival strategies of Borrelia burgdorferi mimic certain environmental conditions. For example, raised temperature, reduced pH, and an expansion in B burgdorferi cell density are conditions that ostensibly mimic those during tick engorgement. They all have been shown to induce a downregulation of OspA, lipoprotein P22, and an upregulation of OspC, decorin binding protein A, multicopy lipoprotein, and the alternative sigma factor RpoS96 (RNA polymerase sigma S subunit) (table). Carroll and colleagues97 reported that a lower pH of the culture affects the transcription of ospC. This report confirmed earlier findings showing that OspC was downregulated in the midgut of unfed ticks.17 This showed that ospC gene expression could be under the coordinate regulation of temperature and pH. Kraiczy et al,69 noted an upregulation of BaCRASP-1, BaCRASP-2, and BaCRASP-5 in cultures of B afzelii by changing the temperature from 33°C to 37°C. He suggested that the upregulation of CRASPs leads to complement resistance and, thus, could be relevant for maintaining bacterial integrity during adaptation to the human host. Erp proteins, which are regulated by temperature as well,98 do not show any significant change in their expression at different pH values, suggesting that not all proteins under temperature regulation are likewise regulated by pH.41,97 Other stimulating mammalian factors, that have a role in regulation of Erp proteins, have been shown by comparing the transcription of erp genes in spirochetes cultivated within the mammalian host environment with solely temperature-shifted spirochetes cultivated in vitro.39 These findings suggested that host-specific signals could further enhance the positive stimulus provided by raised temperature.99 Of interest, even inside the erp lipoprotein family the expression of members in the midgut of the tick varied during feeding. Some family members were expressed early, while other members were expressed late during transmission39 suggesting that there might be factors other than temperature contributing to gene expression. The induction of lipoprotein genes at different points in the borrelial enzootic cycle shows that individual members of a protein/gene family have evolved differently.39 It is likely that B burgdorferi grown in BSK-H medium are receiving a set of regulatory signals. Therefore, studies done in vitro can not truly mimic the conditions experienced by B burgdorferi during the infectious cycle. One striking observation is that B burgdorferi does not possess genes for the synthesis of aminoacids, fatty acids, and other essential elements.8,100 Thus, the organism is dependent on its environment to supply these essential nutrients. Other bacteria typically possess many peptide transporter systems with different specificities to help the use of peptides as a source for aminoacids.100 However, the genome of B burgdorferi seems to code for a single oligo peptide transport system only (Opp).100 It is reported that a chromosomal locus and two plasmid loci (on cp26 and lp54) in B burgdorferi encode homologues of oligopeptide permease components.101 During its natural life cycle B burgdorferi needs to adapt to different hosts, because it moves between the tick and the mammalian host. Bono and co-workers101 reported that expression of oligopeptide permease system A (oppA-ll and oppA-lV) are upregulated 580 during periods of stable nutrient availability. Expression of oppA-ll seems to be specifically downregulated in spirochetes grown in ticks compared with in vitro.100 By contrast, oppAlV gene expression is substantially upregulated in spirochetes located in the mouse.100 Thus, expression of oppA-lV rises with temperature. Differences in the expression and in substrate specificity of OppA proteins have evolved to allow the spirochete to adapt to different environmental niches and requirements. Another gene family of B burgdorferi has been identified, the bdr (borrelia direct repeat-related gene) family. It has been postulated that bdr proteins have a role in sensing and transducing environmental signals.102 There are three distinct subfamilies known as bdrD, bdrE, and bdrF genes.103–105 Production of bdr proteins is influenced to various degrees by environmental conditions.105 Serum deprivation induced the production of bdr proteins in B burgdorferi. The loss of one or more bdr encoding plasmids did not alter the production level of other bdr genes. In this condition the loss of a plasmid could be complemented by a different plasmid. Morphological changes of B burgdorferi cells in response to adverse environmental conditions (changes in pH, depletion of metabolites, ageing, and exposure to antibiotics) have been reported.106–108 The shape of the spirochetes changes to blebs or cysts. Blebs have been shown to contain DNA108,109 and might be implicated in the exchange of genetic information.108 Unlike cysts, blebs are not viable and not capable of transforming back into motile spirochetes.108 In addition, B burgdorferi responded to serum starvation by inducing the synthesis of two starvationinduced proteins (SSP),108 SSP o, which is a homologue of OspA, and SSP s, a homologue of vlsE.108 An upregulation of ospA was also reported during starvation. Upregulation of the spoT gene (guanosine 3,5bispyrophosphate [ppGpp] synthase/pyrophosphohydrolase gene) was shown during starvation. It expresses a protein that acts as a bifunctional enzyme associated in both the degradation and synthesis of ppGpp. PpGpp is an effector molecule implicated in the physiological response of some bacteria to nutritional stress.108 By contrast, serum feeding suppressed spoT expression in B burgdorferi.110 B burgdorferi has to adapt tremendously to survive in its life cycle. The ability to adapt to different environmental conditions like temperature, pH, and cell density enables the spirochetes to effectively cope with environmental changes encountered in the transition from the cold-blooded tick to the warm-blooded mammals. Since B burgdorferi mainly seems to be located in bradytrophic mammalian tissue, genes and their products implicated in nutritional transport have to be used by the spirochete to survive in nonfavourable conditions and locations. On the other hand, these locations might shelter the spirochetes to some extent from the attack of the host’s immune system. Proposed regulatory networks of gene expression in B burgdorferi To date, there is restricted information available on the mechanisms of gene regulation in B burgdorferi. Infectious Diseases Vol 4 September 2004 http://infection.thelancet.com For personal use. Only reproduce with permission from Elsevier Ltd. Review Molecular survival strategies of Borrelia burgdorferi RpoN-RpoS regulatory pathway RpoS and RpoN (RNA polymerase sigma S/N subunit) are stress-induced sigma factors of eubacteria and are implicated in modulating the gene expression of bacterial virulence in different environmental conditions.11,112 Therefore, the RpoS and RpoN pathway represent a key regulatory network where the RpoN protein controls the transcription of rpoS, which ultimately governs the expression of OspC and decorin binding protein A.93,112 Yang and co-workers113 reported the involvement of the RpoN-RpoS pathway in the expression of multicopy lipoproteins. Environmental signals appearing during the tick’s blood meal could activate the RpoN activator protein, which likely binds to an enhancer region upstream of the rpoN gene leading to the synthesis of rpoS mRNA112 and the further expression of important outer surface proteins. Quorum sensing in B burgdorferi To differentially synthesise proteins during its infection cycle, B burgdorferi must possess regulatory networks to sense its environment and regulate the expression of required genes.114 Quorum sensing is a mechanism by which bacteria monitor the presence of other bacteria in their surrounding by producing and responding to signalling molecules known as autoinducers.115 The sensory reception of autoinducers can coordinate the production of virulence factors during host infection.114,116 Two general types of autoinducers have been reported: those that are specific for the species producing it (eg, homoserine lactones or certain polypeptides) and autoinducer type 2 (AI-2). The latter molecules are conserved across the species. In B burgdorferi a gene (BB0377) that shows homology to a quorum sensor has likewise been identified and called luxS. The functional LuxS enzyme enables B burgdorferi to synthesise an AI-2.114 AI-2mediated quorum sensing can function in both the vertebrate host and the arthropod vector. Therefore it could regulate the expression of a different set of proteins in different environments.114 Miller and colleagues117 suggested that during feeding of nymphal ticks, B burgdorferi produce AI-2 to coordinate expression of Erp lipoproteins. The complex life cycle and mechanism of pathogenesis means that B burgdorferi precisely senses its environment and regulates protein expression. An individual bacterium must interact with many different tissue types during its cycle, derive nutrition from warm-blooded hosts as well as from the vector tick, and avoid clearance by the host’s and the vector’s immune systems. Through quorum sensing, a whole population of Lyme disease spirochetes can synchronise production of proteins needed for infection and survival.118 Role of DNA supercoiling in gene regulation DNA supercoiling can affect gene expression in bacteria. It can serve as a signal transducer between sensing the environmental conditions and altering the gene expression. It has been reported that the temperature-mediated changes References 1 2 Singh SK, Girschick HJ. Tick-host interactions and their immunological implications in tick-borne diseases. Current Science 2003; 85: 101–115. 3 Search strategy and selection criteria Data for this review were identified by searches of Medline and references from relevant articles. Many articles were identified through searches of the files of the authors. Search terms were “Lyme borreliosis”, “outer surface proteins AND Borrelia”, “Borrelia burgdorferi AND immune evasion”, “spirochetal persistence”, “Erp proteins AND Borrelia burgdorferi”, “CRASP AND Borrelia burgdorferi”, “Borrelia burgdorferi AND quorum sensing”, “antigenic variation AND Borrelia”, “genetic recombination AND Borrelia”, “Borrelia AND environment”, “OspC AND Borrelia”, “OspA AND Borrelia”, “tick borne diseases”, “vector and host interaction AND borrelia”, “intracellular persistence AND borrelia”, “extracellular persistence AND borrelia”, “environmental adaptation AND Borrelia burgdorferi”, “Gene regulation AND Borrelia burgdorferi”, “host adaptation AND Borrelia burgdorferi”, “spirochetal pathogenicity”, “gene transfer AND Borrelia burgdorferi”, AND “protein polymorphism AND Borrelia burgdorferi”. English language papers were reviewed. of plasmid-encoded gene expression are because of changes in DNA supercoiling.119,120 The rise in culture temperature can reduce DNA twists, whereas a temperature fall can enhance DNA twists.121 Thus, environmental factors that differ between ticks and mammals, like temperature, influence DNA supercoiling of B burgdorferi.122–124 The study done by Alverson and colleagues,124 showed that the temperature-induced changes in the expression of genes ospA and ospC are a result of changes in DNA supercoiling. This phenomenon suggested that DNA supercoiling is a component of the transcriptional regulatory network. Conclusion B burgdorferi has a strong potential for acclimatisation in the invertebrate as well as in the vertebrate host. During this process it adopts different molecular strategies needed for the survival in these different environments. The Lyme spirochete B burgdorferi challenges the host immune system through a broad variety of molecular mechanisms that include complex regulation of differential gene expression. The knowledge of mechanisms of spirochetal gene regulation influencing spirochete/host interactions will open new doors in the understanding of the molecular pathogenesis of Lyme disease. Strategies in the treatment of chronic, antibiotic-resistant Lyme disease might evolve based on future research in this respect. Acknowledgments We acknowledge the continuous support of J Hacker, M Frosch, C Faber, H Morbach, and CP Speer during the preparation of this manuscript. We would particularly like to thank Gundula Girschick for carefully reading the manuscript. Financial support by IZKF grant A26 of the University of Würzburg, Germany is acknowledged. Conflicts of interest None declared. Wikel SK. Tick modulation of host immunity: an important factor in pathogen transmission. Int J Parasitol 1999; 29: 851–59. Hughes CA, Kodner CB, Johnson RC. DNA analysis of Borrelia burgdorferi NCH-1, the first northcentral Infectious Diseases Vol 4 September 2004 http://infection.thelancet.com 4 US human Lyme disease isolate. J Clin Microbiol 1992; 30: 698–703. Charon NW, Goldstein SF. Genetics of motility and chemotaxis of a fascinating group of bacteria: the spirochetes. Annu Rev Genet 2002; 36: 47–73. 581 For personal use. Only reproduce with permission from Elsevier Ltd. Review 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 Goldstein SF, Buttle KF, Charon NW. Structural analysis of the Leptospiraceae and Borrelia burgdorferi by high-voltage electron microscopy. J Bacteriol 1996; 178: 6539–545. Pal U, Fikrig E. Adaptation of Borrelia burgdorferi in the vector and vertebrate host. Microbes Infect 2003; 5: 659–66. Casjens S, Palmer N, van Vugt R, et al. A bacterial genome in flux: the twelve linear and nine circular extrachromosomal DNAs in an infectious isolate of the Lyme disease spirochete Borrelia burgdorferi. Mol Microbiol 2000; 35: 490–516. Fraser CM, Casjens S, Huang WM, et al. Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi. Nature 1997; 390: 580–86. Li H, Dunn JJ, Luft BJ, Lawson CL. Crystal structure of Lyme disease antigen outer surface protein A complexed with an Fab. Proc Natl Acad Sci USA 1997; 94: 3584–89. Wilske B, Preac-Mursic V, Jauris S, et al. Immunological and molecular polymorphisms of OspC, an immunodominant major outer surface protein of Borrelia burgdorferi. Infect Immun 1993; 61: 2182–911. Norris SJ, Carter CJ, Howell JK, Barbour AG. Lowpassage-associated proteins of Borrelia burgdorferi B31: characterization and molecular cloning of OspD, a surface-exposed, plasmid-encoded lipoprotein. Infect Immun 1992; 60: 4662–72. Lam TT, Nguyen TP, Montgomery RR, Kantor FS, Fikrig E, Flavell RA. Outer surface proteins E and F of Borrelia burgdorferi, the agent of Lyme disease. Infect Immun 1994; 62: 290–98. Barbour AG. Antigenic variation of surface proteins of Borrelia species. Rev Infect Dis 1988; 10 (suppl 2): S399–402. Marconi RT, Samuels DS, Garon CF. Transcriptional analyses and mapping of the ospC gene in Lyme disease spirochetes. J Bacteriol 1993; 175: 926–32. Jauris-Heipke S, Liegl G, Preac-Mursic V, et al. Molecular analysis of genes encoding outer surface protein C (OspC) of Borrelia burgdorferi sensu lato: relationship to ospA genotype and evidence of lateral gene exchange of ospC. J Clin Microbiol 1995; 33: 1860–66. Wilske B, Jauris-Heipke S, Lobentanzer R, et al. Phenotypic analysis of outer surface protein C (OspC) of Borrelia burgdorferi sensu lato by monoclonal antibodies: relationship to genospecies and OspA serotype. J Clin Microbiol 1995; 33: 103–09. Schwan TG, Piesman J, Golde WT, Dolan MC, Rosa PA. Induction of an outer surface protein on Borrelia burgdorferi during tick feeding. Proc Natl Acad Sci USA 1995; 92: 2909–13. Hefty PS, Jolliff SE, Caimano MJ, Wikel SK, Akins DR. Changes in temporal and spatial patterns of outer surface lipoprotein expression generate population heterogeneity and antigenic diversity in the Lyme disease spirochete, Borrelia burgdorferi. Infect Immun 2002; 70: 3468–78. Wallich R, Brenner C, Kramer MD, Simon MM. Molecular cloning and immunological characterization of a novel linear-plasmid-encoded gene, pG, of Borrelia burgdorferi expressed only in vivo. Infect Immun 1995; 63: 3327–35. Takayama K, Rothenberg RJ, Barbour AG. Absence of lipopolysaccharide in the Lyme disease spirochete, Borrelia burgdorferi. Infect Immun 1987; 55: 2311–13. Ma Y, Weis JJ. Borrelia burgdorferi outer surface lipoproteins OspA and OspB possess B-cell mitogenic and cytokine-stimulatory properties. Infect Immun 1993; 61: 3843–53. Akins DR, Caimano MJ, Yang X, Cerna F, Norgard MV, Radolf JD. Molecular and evolutionary analysis of Borrelia burgdorferi 297 circular plasmidencoded lipoproteins with OspE- and OspF-like leader peptides. Infect Immun 1999; 67: 1526–32. Barbour AG, Tessier SL, Todd WJ. Lyme disease spirochetes and ixodid tick spirochetes share a common surface antigenic determinant defined by a monoclonal antibody. Infect Immun 1983; 41: 795–804. Anguita J, Hedrick MN, Fikrig E. Adaptation of Borrelia burgdorferi in the tick and the mammalian host. FEMS Microbiol Rev 2003; 27: 493–504. de Silva AM, Telford SR, 3rd, Brunet LR, Barthold SW, Fikrig E. Borrelia burgdorferi OspA is an arthropod-specific transmission-blocking Lyme disease vaccine. J Exp Med 1996; 183: 271–75. Pal U, de Silva AM, Montgomery RR, Fish D, Anguita J, Anderson JF, et al. Attachment of Borrelia burgdorferi within Ixodes scapularis mediated by outer surface protein A. J Clin Invest 2000; 106: 561–69. 582 Molecular survival strategies of Borrelia burgdorferi 27 Barthold SW, Fikrig E, Bockenstedt LK, Persing DH. Circumvention of outer surface protein A immunity by host-adapted Borrelia burgdorferi. Infect Immun 1995; 63: 2255–61. 28 Akins DR, Bourell KW, Caimano MJ, Norgard MV, Radolf JD. A new animal model for studying Lyme disease spirochetes in a mammalian host-adapted state. J Clin Invest 1998; 101: 2240–50. 29 Akin E, McHugh GL, Flavell RA, Fikrig E, Steere AC. The immunoglobulin (IgG) antibody response to OspA and OspB correlates with severe and prolonged Lyme arthritis and the IgG response to P35 correlates with mild and brief arthritis. Infect Immun 1999; 67: 173–81. 30 Gilmore RD Jr., Piesman J. Inhibition of Borrelia burgdorferi migration from the midgut to the salivary glands following feeding by ticks on OspCimmunized mice. Infect Immun 2000; 68: 411–14. 31 Fingerle V, Hauser U, Liegl G, Petko B, Preac-Mursic V, Wilske B. Expression of outer surface proteins A and C of Borrelia burgdorferi in Ixodes ricinus. J Clin Microbiol 1995; 33: 1867–69. 32 Montgomery RR, Malawista SE, Feen KJ, Bockenstedt LK. Direct demonstration of antigenic substitution of Borrelia burgdorferi ex vivo: exploration of the paradox of the early immune response to outer surface proteins A and C in Lyme disease. J Exp Med 1996; 183: 261–69. 33 Indest KJ, Ramamoorthy R, Philipp MT. Transcriptional regulation in spirochetes. J Mol Microbiol Biotechnol 2000; 2: 473–81. 34 Ohnishi J, Piesman J, de Silva AM. Antigenic and genetic heterogeneity of Borrelia burgdorferi populations transmitted by ticks. Proc Natl Acad Sci USA 2001; 98: 670–75. 35 Sadziene A, Wilske B, Ferdows MS, Barbour AG. The cryptic ospC gene of Borrelia burgdorferi B31 is located on a circular plasmid. Infect Immun 1993; 61: 2192–95. 36 Stevenson B, Tilly K, Rosa PA. A family of genes located on four separate 32-kilobase circular plasmids in Borrelia burgdorferi B31. J Bacteriol 1996; 178: 3508–16. 37 Suk K, Das S, Sun W, et al. Borrelia burgdorferi genes selectively expressed in the infected host. Proc Natl Acad Sci USA 1995; 92: 4269–73. 38 Stevenson B, Miller JC. Intra- and interbacterial genetic exchange of Lyme disease spirochete erp genes generates sequence identity amidst diversity. J Mol Evol 2003; 57: 309–24. 39 Hefty PS, Jolliff SE, Caimano MJ, Wikel SK, Radolf JD, Akins DR. Regulation of OspE-related, OspF-related, and Elp lipoproteins of Borrelia burgdorferi strain 297 by mammalian host-specific signals. Infect Immun 2001; 69: 3618–27. 40 Stevenson B, Zuckert, WR, Akins DR. Repetition, conservation and variation: the multiple cp32 plasmids of Borrelia species. In: Saier MH, GarciaLara, J, eds. The spirochetes: molecular and cellular biology. Oxford: Horizon press, 2001: 87–100. 41 Stevenson B. Borrelia burgdorferi erp (ospE-related) gene sequences remain stable during mammalian infection. Infect Immun 2002; 70: 5307–11. 42 Marconi RT, Sung SY, Hughes CA, Carlyon JA. Molecular and evolutionary analyses of a variable series of genes in Borrelia burgdorferi that are related to ospE and ospF, constitute a gene family, and share a common upstream homology box. J Bacteriol 1996; 178: 5615–26. 43 El-Hage N, Babb K, Carroll JA, et al. Surface exposure and protease insensitivity of Borrelia burgdorferi Erp (OspEF-related) lipoproteins. Microbiology 2001; 147(Pt 4): 821–30. 44 Stevenson B, Casjens S, Rosa P. Evidence of past recombination events among the genes encoding the Erp antigens of Borrelia burgdorferi. Microbiology 1998; 144 (Pt 7): 1869–79. 45 Stevenson B, Zuckert WR, Akins DR. Repetition, conservation, and variation: the multiple cp32 plasmids of Borrelia species. J Mol Microbiol Biotechnol 2000; 2: 411–22. 46 Barbour AG, Restrepo BI. Antigenic variation in vector-borne pathogens. Emerg Infect Dis 2000; 6: 449–57. 47 Zhang JR, Hardham JM, Barbour AG, Norris SJ. Antigenic variation in Lyme disease borreliae by promiscuous recombination of VMP-like sequence cassettes. Cell 1997; 89: 275–85. 48 Anguita J, Thomas V, Samanta S, Persinski R, Hernanz C, Barthold SW, et al. Borrelia burgdorferiinduced inflammation facilitates spirochete adaptation and variable major protein-like sequence locus recombination. J Immunol 2001; 167: 3383–90. 49 Liang FT, Jacobs MB, Philipp MT. C-terminal invariable domain of VlsE may not serve as target for protective immune response against Borrelia burgdorferi. Infect Immun 2001; 69: 1337–43. 50 Wang G, van Dam AP, Dankert J. Analysis of a VMPlike sequence (vls) locus in Borrelia garinii and Vls homologues among four Borrelia burgdorferi sensu lato species. FEMS Microbiol Lett 2001; 199: 39–45. 51 Labandeira-Rey M, Skare JT. Decreased infectivity in Borrelia burgdorferi strain B31 is associated with loss of linear plasmid 25 or 28-1. Infect Immun 2001; 69: 446–55. 52 Purser JE, Norris SJ. Correlation between plasmid content and infectivity in Borrelia burgdorferi. Proc Natl Acad Sci USA 2000; 97: 13865–870. 53 Zhang JR, Norris SJ. Gentic variation of the Borrelia burgdorferi gene vlse involves cassette specific, segmental gene conversion. Infect Immun 1998; 66: 3698–3704. 54 Eicken C, Sharma V, Klabunde T, et al. Crystal structure of Lyme disease variable surface antigen VlsE of Borrelia burgdorferi. J Biol Chem 2002; 277: 21691–96. 55 McDowell JV, Sung SY, Hu LT, Marconi RT. Evidence that the variable regions of the central domain of VlsE are antigenic during infection with lyme disease spirochetes. Infect Immun 2002; 70: 4196–203. 56 Eggers CH, Casjens S, Hayes SF, et al. Bacteriophages of spirochetes. J Mol Microbiol Biotechnol 2000; 2: 365–73. 57 Eggers CH, Samuels DS. Molecular evidence for a new bacteriophage of Borrelia burgdorferi. J Bacteriol 1999; 181: 7308–13. 58 Balmelli T, Piffaretti JC. Analysis of the genetic polymorphism of Borrelia burgdorferi sensu lato by multilocus enzyme electrophoresis. Int J Syst Bacteriol 1996; 46: 167–72. 59 Wang G, van Dam AP, Schwartz I, Dankert J. Molecular typing of Borrelia burgdorferi sensu lato: taxonomic, epidemiological, and clinical implications. Clin Microbiol Rev 1999; 12: 633–53. 60 Azulay RD, Azulay-Abulafia L, Sodre CT, Azulay DR, Azulay MM. Lyme disease in Rio de Janeiro, Brazil. Int J Dermatol 1991; 30: 569–71. 61 Kirstein F, Rijpkema S, Molkenboer M, Gray JS. Local variations in the distribution and prevalence of Borrelia burgdorferi sensu lato genomospecies in Ixodes ricinus ticks. Appl Environ Microbiol 1997; 63: 1102–6. 62 Sung SY, McDowell JV, Carlyon JA, Marconi RT. Mutation and recombination in the upstream homology box-flanked ospE- related genes of the Lyme disease spirochetes result in the development of new antigenic variants during infection. Infect Immun 2000; 68: 1319–27. 63 Nordstrand A, Barbour AG, Bergstrom S. Borrelia pathogenesis research in the post-genomic and postvaccine era. Curr Opin Microbiol 2000; 3: 86–92. 64 Wang IN, Dykhuizen DE, Qiu W, Dunn JJ, Bosler EM, Luft BJ. Genetic diversity of ospC in a local population of Borrelia burgdorferi sensu stricto. Genetics 1999; 151: 15–30. 65 Sung SY, McDowell JV, Carlyon JA, Marconi RT. Mutation and recombination in the upstream homology box-flanked ospE-related genes of the Lyme disease spirochetes result in the development of new antigenic variants during infection. Infect Immun 2000; 68: 1319–27. 66 Akins DR, Porcella SF, et al. Evidence for in vivo but not in vitro expression of a Borrelia burgdorferi outer surface protein F (OspF) homologue. Mol Microbiol 1995; 18: 507–20. 67 Champion CI, Blanco DR, Skare JT, et al. A 9.0kilobase-pair circular plasmid of Borrelia burgdorferi encodes an exported protein: evidence for expression only during infection. Infect Immun 1994; 62: 2653–61. 68 Zipfel PF, Skerka C. FHL-1/reconectin: a human complement and immune regulator with celladhesive function. Immunol Today 1999; 20: 135–40. 69 Kraiczy P, Skerka C, Brade V, Zipfel PF. Further characterisation of complement regulator-acquiring surface proteins of Borrelia burgdorferi. Infect Immun 2001; 69: 7800-809. 70 Zipfel PF, Skerka C, Hellwage J, et al. Factor H family proteins: on complement, microbes and human diseases. Biochem Soc Trans 2002; 30 (Pt 6): 971–78. 71 Kraiczy P, Skerka C, Kirschfink M, Brade V, Zipfel PF. Immune evasion of Borrelia burgdorferi by acquisition of human complement regulators FHL1/reconectin and Factor H. Eur J Immunol 2001; 31: 1674–84. 72 Stevenson B, Bono JL, Schwan TG, Rosa P. Borrelia burgdorferi erp proteins are immunogenic in mammals infected by tick bite, and their synthesis is inducible in cultured bacteria. Infect Immun 1998; 66: 2648–54. Infectious Diseases Vol 4 September 2004 http://infection.thelancet.com For personal use. Only reproduce with permission from Elsevier Ltd. Molecular survival strategies of Borrelia burgdorferi 73 Alitalo A, Meri T, Ramo L, et al. Complement evasion by Borrelia burgdorferi: serum-resistant strains promote C3b inactivation. Infect Immun 2001; 69: 3685–91. 74 Hellwage J, Meri T, Heikkila T, et al. The complement regulator factor H binds to the surface protein OspE of Borrelia burgdorferi. J Biol Chem 2001; 276: 8427–35. 75 Stevenson B, El-Hage N, Hines MA, Miller JC, Babb K. Differential binding of host complement inhibitor factor H by Borrelia burgdorferi Erp surface proteins: a possible mechanism underlying the expansive host range of Lyme disease spirochetes. Infect Immun 2002; 70: 491–97. 76 Kurtenbach K, De Michelis S, Etti S, et al. Host association of Borrelia burgdorferi sensu lato—the key role of host complement. Trends Microbiol 2002; 10: 74–79. 77 Dorward DW, Fischer ER, Brooks DM. Invasion and cytopathic killing of human lymphocytes by spirochetes causing Lyme disease. Clin Infect Dis 1997; 25 (suppl 1): S2–8. 78 Grab DJ, Salim M, Chesney J, Bucala R, Lanners HN. A role for peripheral blood fibrocytes in Lyme disease? Med Hypotheses 2002; 59: 1–10. 79 Grab DJ, Lanners H, Martin LN, et al. Interaction of Borrelia burgdorferi with peripheral blood fibrocytes, antigen-presenting cells with the potential for connective tissue targeting. Mol Med 1999; 5: 46-54. 80 Georgilis K, Peacocke M, Klempner MS. Fibroblasts protect the Lyme disease spirochete, Borrelia burgdorferi, from ceftriaxone in vitro. J Infect Dis 1992; 166: 440–44. 81 Girschick HJ, Huppertz HI, Russmann H, Krenn V, Karch H. Intracellular persistence of Borrelia burgdorferi in human synovial cells. Rheumatol Int 1996; 16: 125-32. 82 Girschick HJ, Meister S, Karch H, Huppertz HI. Borrelia burgdorferi downregulates ICAM-1 on human synovial cells in vitro. Cell Adhes Commun 1999; 7: 73–83. 83 Ma Y, Sturrock A, Weis JJ. Intracellular localisation of Borrelia burgdorferi within human endothelial cell. Infect Immunol 1991; 59: 671–78. 84 Montgomery RR, Nathanson MH, Malawista SE. The fate of Borrelia burgdorferi, the agent for Lyme disease, in mouse macrophages. Destruction, survival, recovery. J Immunol 1993; 150: 909–15. 85 Pachner AR, Basta J, Delaney E, Hulinska D. Localisation of Borrelia burgdorferi in murine Lyme borreliosis by electron microscopy. Am J Trop Med Hyg 1995; 52: 128–33. 86 Chary-Valckenaere I, Jaulhac B, Champigneulle J, Piemont Y, Mainard D, Pourel J. Ultrastructural demonstration of intracellular localisation of Borrelia burgdorferi in Lyme arthritis. Br J Rheumatol 1998; 37: 468–70. 87 Dejmkova H, Hulinska D, Tegzova D, Pavelka K, Gatterova J, Vavrik P. Seronegative Lyme arthritis caused by Borrelia garinii. Clin Rheumatol 2002; 21: 330–34. 88 Haupl T, Hahn G, Rittig M, et al. Persistence of Borrelia burgdorferi in ligamentous tissue from a patient with chronic Lyme borreliosis. Arthritis Rheum 1993; 36: 1621–26. 89 Franz JK, Fritze O, Rittig M, et al. Insights from a novel three-dimensional in vitro model of lyme arthritis: standardized analysis of cellular and molecular interactions between Borrelia burgdorferi and synovial explants and fibroblasts. Arthritis Rheum 2001; 44: 151–62. 90 Priem S, Burmester GR, Kamradt T, Wolbart K, Rittig MG, Krause A. Detection of Borrelia burgdorferi by polymerase chain reaction in synovial membrane, but not in synovial fluid from patients with persisting Lyme arthritis after antibiotic therapy. Ann Rheum Dis 1998; 57: 118–21. 91 Cadavid D, Pachner AR, Estanislao L, Patalapati R, Barbour AG. Isogenic serotypes of Borrelia turicatae show different localisation in the brain and skin of mice. Infect Immun 2001; 69: 3389–397. 92 Cadavid D, O'Neill T, Schaefer H, Pachner AR. Localisation of Borrelia burgdorferi in the nervous system and other organs in a nonhuman primate model of lyme disease. Lab Invest 2000; 80: 1043–54. 93 Yang X, Goldberg MS, Popova TG, et al. Interdependence of environmental factors influencing reciprocal patterns of gene expression in virulent Borrelia burgdorferi. Mol Microbiol 2000; 37: 1470–79. 94 Fikrig E, Feng W, Barthold SW, Telford SR 3rd, Flavell RA. Arthropod- and host-specific Borrelia burgdorferi bbk32 expression and the inhibition of spirochete transmission. J Immunol 2000; 164: 5344–51. 95 Schwan TG, Piesman J. Temporal changes in outer surface proteins A and C of the lyme diseaseassociated spirochete, Borrelia burgdorferi, during the chain of infection in ticks and mice. J Clin Microbiol 2000; 38: 382–88. 96 Revel AT, Talaat AM, Norgard MV. DNA microarray analysis of differential gene expression in Borrelia burgdorferi, the Lyme disease spirochete. Proc Natl Acad Sci USA 2002; 99: 1562–67. 97 Carroll JA, Garon CF, Schwan TG. Effects of environmental pH on membrane proteins in Borrelia burgdorferi. Infect Immun 1999; 67: 3181–87. 98 Babb K, El-Hage N, Miller JC, Carroll JA, Stevenson B. Distinct regulatory pathways control expression of Borrelia burgdorferi infection-associated OspC and Erp surface proteins. Infect Immun 2001; 69: 4146–53. 99 Fikrig E, Chen M, Barthold SW, et al. Borrelia burgdorferi erpT expression in the arthropod vector and murine host. Mol Microbiol 1999; 31: 281–90. 100 Wang XG, Lin B, Kidder JM, Telford S, Hu LT. Effects of environmental changes on expression of the oligopeptide permease (opp) genes of Borrelia burgdorferi. J Bacteriol 2002; 184: 6198–206. 101 Bono JL, Tilly K, Stevenson B, Hogan D, Rosa P. Oligopeptide permease in Borrelia burgdorferi: putative peptide-binding components encoded by both chromosomal and plasmid loci. Microbiology 1998; 144 ( Pt 4): 1033–44. 102 Roberts DM, Theisen M, Marconi RT. Analysis of the cellular localization of Bdr paralogs in Borrelia burgdorferi, a causative agent of lyme disease: evidence for functional diversity. J Bacteriol 2000; 182: 4222–26. 103 Carlyon JF, Roberts DM, Marconi RT. Evolutionary and molecular analyses of the Borrelia bdr super gene family: delineation of distinct sub-families and demonstration of the genus wide conservation of putative functional domains, structural properties and repeat motifs. Microb Pathog 2000; 28: 89–105. 104 Zuckert WR, Meyer J, Barbour AG. Comparative analysis and immunological characterisation of the Borrelia Bdr protein family. Infect Immun 1999; 67: 3257–66. Infectious Diseases Vol 4 September 2004 http://infection.thelancet.com Review 105 Roberts DM, Caimano M, McDowell J, et al. Environmental regulation and differential production of members of the Bdr protein family of Borrelia burgdorferi. Infect Immun 2002; 70: 7033–41. 106 Barbour AG, Hayes SF. Biology of Borrelia species. Microbiol Rev 1986; 50: 381–400. 107 Brorson O, Brorson SH. Transformation of cystic forms of Borrelia burgdorferi to normal, mobile spirochetes. Infection 1997; 25: 240–6. 108 Alban PS, Johnson PW, Nelson DR. Serumstarvation-induced changes in protein synthesis and morphology of Borrelia burgdorferi. Microbiology 2000; 146 (Pt 1): 119–27. 109 Garon CF, Dorward DW, Corwin MD. Structural features of Borrelia burgdorferi—the Lyme disease spirochete: silver staining for nucleic acids. Scanning Microsc Suppl 1989; 3: 109–15. 110 Concepcion MB, Nelson DR. Expression of spoT in Borrelia burgdorferi during serum starvation. J Bacteriol 2003; 185: 444–52. 111 Wosten MM. Eubacterial sigma-factors. FEMS Microbiol Rev 1998; 22: 127–50. 112 Hubner A, Yang X, Nolen DM, Popova TG, Cabello FC, Norgard MV. Expression of Borrelia burgdorferi OspC and DbpA is controlled by a RpoN-RpoS regulatory pathway. Proc Natl Acad Sci USA 2001; 98: 12724–729. 113 Yang XF, Hubner A, Popova TG, Hagman KE, Norgard MV. Regulation of expression of the paralogous Mlp family in Borrelia burgdorferi. Infect Immun 2003; 71: 5012–20. 114 Stevenson B, Babb K. LuxS-mediated quorum sensing in Borrelia burgdorferi, the lyme disease spirochete. Infect Immun 2002; 70: 4099–105. 115 Taga ME, Bassler BL. Chemical communication among bacteria. Proc Natl Acad Sci USA 2003; 100 (suppl 2): 14549–554. 116 Schauder S, Bassler BL. The languages of bacteria. Genes Dev 2001; 15: 1468–80. 117 Miller JC, Stevenson B. Increased expression of Borrelia burgdorferi factor H-binding surface proteins during transmission from ticks to mice. Int J Med Microbiol 2004; 293 (suppl 37): 120–5. 118 Stevenson B, von Lackum K, Wattier RL, McAlister JD, Miller JC, Babb K. Quorum sensing by the Lyme disease spirochete. Microbes Infect 2003; 5: 991–7. 119 Samuels DS, Garon CF. Coumermycin A1 inhibits growth and induces relaxation of supercoiled plasmids in Borrelia burgdorferi, the Lyme disease agent. Antimicrob Agents Chemother 1993; 37: 46–50. 120 El-Hage N, Stevenson B. Simultaneous coexpression of Borrelia burgdorferi Erp proteins occurs through a specific, erp locus-directed regulatory mechanism. J Bacteriol 2002; 184: 4536–43. 121 Rohde JR, Fox JM, Minnich SA. Thermoregulation in Yersinia enterocolitica is coincident with changes in DNA supercoiling. Mol Microbiol 1994; 12: 187–99. 122 Tse-Dinh YC, Qi H, Menzel R. DNA supercoiling and bacterial adaptation: thermotolerance and thermoresistance. Trends Microbiol 1997; 5: 323–6. 123 Lopez-Garcia P, Forterre P. DNA topology and the thermal stress response, a tale from mesophiles and hyperthermophiles. Bioessays 2000; 22: 738–46. 124 Alverson J, Bundle SF, Sohaskey CD, Lybecker MC, Samuels DS. Transcriptional regulation of the ospAB and ospC promoters from Borrelia burgdorferi. Mol Microbiol 2003; 48: 1665–77. 583 For personal use. Only reproduce with permission from Elsevier Ltd.