Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
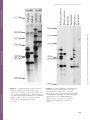
Journal of Heredity 2004:95(5):436–444 DOI: 10.1093/jhered/esh057 ª 2004 The American Genetic Association A Deletion Affecting Several Gene Candidates is Present in the Evergrowing Peach Mutant D. G. BIELENBERG, Y. WANG, S. FAN, G. L. REIGHARD, R. SCORZA, AND A. G. ABBOTT Address correspondence to D. G. Bielenberg, at the above address, or e-mail: [email protected]. Abstract Evergrowing (EVG ) peach is one of only two described mutants affecting winter dormancy in woody perennial species. EVG peach does not set terminal buds, cease new leaf growth, nor enter into a dormant resting phase in response to winter conditions. The EVG mutation segregates in F2 progeny as a single recessive nuclear gene. A local molecular genetic linkage map around EVG was previously developed using amplified fragment length polymorphism (AFLP) and simple sequence repeat (SSR) markers, and a bacterial artificial chromosome (BAC) contig that contains the EVG mutation was assembled. A MADS box coding open reading frame (ORF) was found in a BAC of this contig and used as a probe. The probe detected a polymorphism between the wild-type and mutant genomes, and the polymorphism is indicative of a deletion in EVG peach. The EVG gene region contained six potential MADS-box transcription factor sequences, and the deletion in EVG affected at least four of these. The deletion was bracketed using RFLP analysis, which showed that it is contained within a segment of the genome no greater than 180 kb. Perennial woody plants are the dominant species in many ecosystems of the world and have significant ecological and economic importance. A major life-history trait of perennial species is the ability to become physiologically dormant in order to avoid unfavorable climatic conditions, such as the low temperatures and limited water availability associated with winter. Mutants that fail to cease growth and enter dormancy under dormancy-inducing conditions have been described in only two woody perennial species, Corylus avellana L. (Thompson et al. 1985) and Prunus persica (L.) Batsch (Rodriguez et al. 1994). The best described mutant of these two species is the Evergrowing (previously known as Evergreen, USDA PI442380) peach mutant, a nondormant genotype identified from southern Mexico, where killing frosts do not occur (Rodriguez et al. 1994; Werner and Okie 1998). In Mexico, terminal growth on Evergrowing trees is continuous under favorable environmental conditions, and leaves are retained until they are lost due to drought and/or disease (Diaz 1974). At more northern latitudes, the Evergrowing peach does not appear to respond to winter dormancy cues, exhibiting persistent shoot growth and a lack of leaf abscission when experiencing short days and low temperatures in the fall until these tissues are killed by freezing temperatures (Rodriguez et al. 1994). In addition, 436 the frost hardiness of Evergrowing trees is half that of wildtype dormant trees (Rodriguez et al. 1994). For example, Evergrowing trees show some cold acclimation and accumulation of bark storage proteins and dehydrins; this occurs later in the fall and to a lesser degree than in wild-type trees (Arora and Wisniewski 1996; Arora et al. 1992, 1996). A number of crosses of Evergrowing (nondormant) trees with different wild-type dormant trees were made by Rodriguez et al. (1994), and all nine of the resulting F2 progenies fit a 3:1 (dormant:nondormant) ratio, suggesting that the Evergrowing phenotype is controlled by a single recessive nuclear gene. A genetic map of the Evergrowing (EVG ) locus was previously generated from a segregating F2 population derived from a cross between wild-type dormant (‘‘Empress’’ dwarf) and mutant, nondormant (Evergrowing, PI442380) parents and reported by Wang et al. (2002b). Subsequently a physical map of the EVG region comprised of bacterial artificial chromosomes (BACs) was initiated from the closest sequence tagged site (STS) marker found in that study (Wang et al. 2002a). Analysis of simple sequence repeat (SSR) markers developed from this contig confirmed the positions of the genetic makers relative to one another (Wang et al. 2002a). BAC PpN018F12 (Prunus persica ‘‘Nemared’’ 018F12) Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 From the Department of Horticulture, Clemson University, Clemson, SC 29634 (Bielenberg, Wang, and Reighard); Department of Genetics, Biochemistry, and Life Science Studies, Clemson University, Clemson, SC 29634 (Fan and Abbott); and USDA-ARS Appalachian Fruit Research Station, Kearneysville, WV 25430 (Scorza). Bielenberg et al. A Deletion in Evergrowing Peach Methods Genomic DNA Isolation Peach leaves for DNA extraction were obtained from individuals of an F2 mapping population and from moderate chilling cultivars (Musser Fruit Research Center, Clemson University, Clemson, SC). All other peach tissue was shipped overnight on ice from the USDA-ARS Southeastern Fruit and Nut Research Laboratory (Byron, GA). Leaves were weighed as 1.0 g fresh weight samples, wrapped in aluminum foil, frozen in liquid N2, and stored at 808C. Total DNA was isolated from the frozen leaves using a CTAB (hexadecyltrimethylammonium bromide) extraction buffer protocol modified from Doyle and Doyle (1990). For each sample 1.0 g of frozen leaf tissue was ground in a mortar and pestle under liquid N2. This ground tissue was transferred to 15 ml CTAB buffer [2% (w/v) CTAB, 1.4 M NaCl, 20 mM EDTA, 100 mM Tris-HCL pH 8.0] in a 50 ml plastic culture tube at 608C and incubated for 30 min at 608C. After incubation, one volume of chloroform:isoamyl alcohol (24:1) was added and the contents gently mixed. The sample was then clarified by centrifugation at 1600g for 10 min at 48C. The upper aqueous phase was transferred to a clean 50 ml culture tube and another volume of chloroform:isoamyl alcohol (24:1) was added and gently mixed. The sample was again clarified by centrifugation at 1600g for 10 min at 48C, and the resulting upper, aqueous phase transferred to a clean 50 ml tube. The DNA was precipitated from this solution by the addition of one-tenth volume 3.3 M sodium acetate (pH 5.2) and one volume of cold isopropanol. Recovered nucleic acids were transferred to a 2.0 ml microcentrifuge tube, washed in 70% (v/v) ethanol, and resuspended in 1.0 ml TE. RNase A (Sigma-Aldrich Corp., St. Louis, MO) was added to a concentration of 10 lg/ml and the sample was incubated at 378C for 30 min. RNase A-treated samples were ethanol precipitated, resuspended in 500 ll of 0.13 TE, and quantified by spectrophotometry. BAC DNA Isolation All BAC clones used in this study were obtained from a genomic library of the peach rootstock Nemared (wildtype dormant), digested with HindIII and cloned into the pBeloBAC11 vector (Georgi et al. 2002). BAC DNA was isolated from Escherichia coli cultured overnight at 378C in 10 ml LB media using the alkaline lysis protocol for large insert clones described by Sambrook and Russell (2002). DNA quality and quantity was assessed by spectrophotometry. BAC End Sequencing Cesium chloride-purified BAC DNA was used as a template for end sequencing of PpN018G07 and PpN089G02. Sequencing reactions were performed using Big Dye v2.0 terminator chemistry (Applied Biosystems, Inc., Foster City, CA) according to the manufacturer’s instructions for large insert DNA. Sequencing reactions were analyzed on an ABI Prism 377 sequencer. BAC end sequences were submitted to GenBank and assigned the following accession numbers: PpN018G07SP6 (CL311195), PpN018G07T7 (CL311196), PpN089G02SP6 (CL311194), and PpN089G02T7 (CL311193). 437 Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 was found to contain the EAT/MCAC marker (Wang et al. 2002a), which was mapped at a distance of 1 cM from the EVG gene (Wang et al. 2002b). BAC PpN109L12 was shown to overlap with the T7 end of BAC PpN018F12. An SSR marker (pchgms29) was developed from BAC PpN109L12 and mapped 2.9 cM from the EAT/MCAC marker, in the opposite direction from the map location of the EVG gene (Wang 2002). The position of pchgms29 confirms the contig walk initiated from the SP6 end of BAC PpN018F12 is in the direction of the EVG gene (Wang 2002). End sequencing of PpN018F12 revealed a potential open reading frame (ORF) with homology to a MADS box-type transcription factor and was designated bes_18F12B01 (BAC end sequence of BAC PpN018F12 clone B01) (Georgi et al. 2003). MADS box transcription factors are found in a wide variety of organisms and regulate important developmental processes (Garcia-Maroto et al. 2003). In plants, MADS box genes have been shown to regulate reproductive traits such as the timing of meristem transition from vegetative to reproductive growth in response to environmental conditions, floral structure, fruit ripening, and abscission of mature fruit (Fang and Fernandez 2002; Lohmann and Weigel 2002; Mao et al. 2000; Mouradov et al. 2002; Vrebalov et al. 2002). In addition, a number of plant MADS box genes have been found that are expressed in vegetative tissues (Alvarez-Buylla et al. 2000; Garcia-Maroto et al. 2000; Johansen et al. 2002; Kim et al. 2002; van der Linden et al. 2002). However, with the exception of ANR1 in Arabidopsis, which is in the signaling pathway for increased root elongation in response to nitrate (Zhang and Forde 2000), and POTM1, which is involved in suppression of axillary bud growth in potato (Rosin et al. 2003), most of these genes do not yet have a defined function. Nonetheless, the role that MADS box genes play in major developmental transitions in plants made the MADS sequence found in PpN018F12 a candidate for the Evergrowing mutation. Here we report that a hybridization probe developed from the MADS box coding sequence found in bes_18F12B01 is polymorphic between genomic DNA from wild-type dormant and mutant nondormant EVG peach trees. Southern analyses revealed a cluster of six genes homologous to the sequenced ORF and four of these genes were missing in the mutant, indicating that a possible deletion of these genes was present in the Evergrowing peach mutant. In addition, we demonstrated that these six MADS box genes were located within the approximately 180 kb region spanned by three overlapping BAC clones and that the hypothesized deletion was bracketed by the ends of this BAC contig. Journal of Heredity 2004:95(5) Probe Amplification and Labeling Figure 1. Sequence of peach BAC partial sequence bes_18F12B01. Sequence in uppercase is the 196 bp MADS box ORF. Underlined and bolded sequences denote the primers used to generate the 121 bp DNA fragment used in subsequent Southern hybridizations. Southern Analysis Genomic or BAC DNA was digested at 378C overnight with 10 units of enzyme (HindIII, EcoRI, or BamHI; Promega Corp.) and size fractionated in a 0.8% (w/v) SeaKem LE agarose gel (BioWhittaker Molecular Applications, Rockland, ME) using 13 TAE (40 mM Tris-acetate, 1 mM EDTA, pH 8.0) as a running buffer. After separation was completed, gels were depurinated by incubation in 0.125 N HCl followed by a 30 min incubation in denaturing buffer (1.5 M NaCl, 0.5 M NaOH), and finally a 30 min incubation in neutralization buffer (1.5 M NaCl, 0.5 M Tris pH 7.5). DNA was transferred to Hybond-XL membrane (Amersham Pharmacia Biotech, Little Chalfort, Buckinghamshire, England) by capillary blot using 103 SSC (pH 7.0) as a transfer buffer. Following transfer, DNA was fixed to the membrane by baking at 808C for 2 h. Membranes were prehybridized at 658C with a 0.5 M sodium phosphate, 7% sodium dodecylsulfate (SDS) hybridization buffer for a minimum of 30 min. Radioactively labeled and denatured DNA probe was added directly to the prehybridization solution, and the membranes were hybridized with the probe overnight at 658C. All hybridized membranes were washed at 658C twice with 23 SSC (pH 7.0), 1% (w/v) SDS for 30 min, and twice with 13 SSC, 0.1% (w/v) SDS for 30 min. The Southern blot containing DNA from nonpeach species were prehybridized and washed at 508C. Membranes were then exposed to autoradiographic film (Kodak X-Omat Blue XB-1, Perkin-Elmer Life Sciences, Inc., Boston, MA) for 3–10 days as determined by signal strength. Results Primers were designed for the conserved regions of the MADS box sequence found in bes_18F12B01 (BAC end sequence of BAC PpN018F12 clone B01) (Figure 1). A 119 bp fragment of the MADS box sequence was amplified by PCR from PpN018F12 for use as a DNA probe (Figure 1). 438 The amplified probe was hybridized to restriction enzymedigested DNA from wild-type dormant and nondormant mutant trees from the F2 population used to map the locus. In the HindIII digested wild-type DNA, the probe detected what appeared to be five strongly hybridizing bands, one each at 3.3 kb, 2.7 kb, 2.4 kb, 2.1 kb, and 1.5 kb (Figure 2). However, the smallest hybridizing band was of greater intensity and wider spread than the other four bands, indicating that it was either the sequence of highest similarity to the hybridizing probe or potentially two or more overlapping bands (Figure 2). In the HindIII digest of the mutant DNA, the two bands present corresponded in size to two of the bands in the wild-type DNA—2.7 kb and 1.5 kb (Figure 2). The 1.5 kb band in the HindIII-digested mutant DNA was of lower intensity and spread than the band at the same position in the wild-type DNA. This interpretation is consistent with the presence of two overlapping bands at 1.5 kb in the wild-type DNA and the presence of only one, slight larger band in the mutant DNA (Figure 2). The EcoRI-digested wild-type DNA showed six strongly hybridizing bands, one each at approximately 12.0 kb, 7.2 kb, 4.9 kb, 3.5 kb, 3.3 kb, and 2.2 kb (Figure 2). In the EcoRIdigested EVG mutant DNA, one band corresponded in size to a band existing in the wild-type digestion, that is, 4.9 kb, but the larger band did not correspond to one in the wild type (Figure 2). Three overlapping BAC clones from a previously created EVG locus physical contig (Wang et al. 2002a) were digested with HindIII and separated on a gel with HindIII-digested DNA from wild-type dormant and nondormant mutant trees, and hybridized with the amplified MADS box probe (Figure 3). HindIII was used for analysis of the BAC clones because the BAC clones were prepared from partial HindIIIdigested fragments of the peach genome and therefore should produce a banding pattern identical to the genomic DNA. The three BAC clones together possessed bands corresponding to all six of the HindIII-digested MADS box hybridizing bands in the wild-type DNA (Figure 3). These Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 Forward and reverse PCR primers for each of the amplifications were designed from BAC end sequences using Primer3_www.cgi v 0.2 (Rozen and Skaletsky 2000) and obtained from Integrated DNA Technologies (Coralville, IA). The BAC from which primer pairs were designed was used as a template to amplify probes. Amplification products were size fractionated on an agarose gel (PpN089G02SP6, 260 bp; PpN018G07SP6, 235 bp; bes_18F12B01, 119 bp). The amplification products were excised from the gel and purified using the QIAquick Gel Extraction kit (Qiagen, Inc., Valencia, CA). DNA probes were labeled with a32P-dCTP (PerkinElmer Life Sciences, Inc., Boston, MA) following denaturing and incubation with random DNA hexamers and the Klenow fragment of DNA polymerase (Promega Corp., Madison, WI) at 378C for 3 h (Sambrook and Russell 2002). Bielenberg et al. A Deletion in Evergrowing Peach Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 Figure 2. Southern hybridization of digested genomic DNA from wild-type dormant (DE) and Evergrowing mutant (EVG ) F2 siblings. Genomic DNA samples (10 lg) were digested with HindIII, EcoRI, or BamHI. Membranes were probed with DNA amplified from the bes_18F12B01 MADS box ORF (121 bp). Lines represent positions of bands from the HindIII-digested k DNA size marker. Figure 3. Southern hybridization of HindIII-digested genomic DNA from wild-type dormant (DE) and nondormant mutant (EVG) peaches and of three BACs from the EVG contig (PpN089G02, PpN018F12, and PpN018G07). Membranes were probed with DNA amplified from the bes_18F12B01 MADS box ORF, as in Figure 2. Lines represent positions of bands from the HindIII-digested k DNA size marker. 439 Journal of Heredity 2004:95(5) bands were labeled, from largest to smallest, as A, 3.3 kb; B, 2.7 kb; C, 2.4 kb; D, 2.1 kb; E, 1.5 kb; and F, 1.5 kb. The four missing bands in the nondormant mutant DNA (A, C, D, and F) corresponded to fragments observed in PpN018F12 or PpN089G02, or both. The hybridizing fragment (E) observed in PpN018G07 was not altered between the wildtype and mutant genomic DNAs (Figure 3). EcoRI digestion of the BACs and subsequent hybridization with the MADS box probe resulted in nonidentical band patterns relative to the genomic DNA (data not shown). Therefore the analysis of this digest could not be used to confirm the molecular structure of the EVG locus in the wild-type and mutant genomes. The SP6 and T7 ends of BACs PpN089G02 and PpN018G07 were sequenced, and these sequences were used to identify amplification primers for the production of BAC end specific probes for the EVG contig. T7 ends of PpN018G07 and PpN089G02 were previously determined to overlap with PpN018F12 by chromosomal walking during the generation of the BAC contig (Figure 4) (Wang et al. 2002a). Therefore the extreme ends of the BAC contig are represented by the SP6 ends of PpN018G07 and PpN089G02. The probes were separately hybridized onto a Southern membrane containing HindIII-digested wild-type genomic DNA, mutant genomic DNA, and the three BACs in the EVG contig (Figure 5). The PpN089G02-SP6 end probe hybridized to a fragment of similar size (1.9 kb) in the wildtype genomic DNA, mutant genomic DNA, and PpN089G02, but did not hybridize to a fragment in either of the other two BACs (Figure 5A). The PpN018G07-SP6 end probe hybridized to a fragment of similar size (0.7 kb) in the wild-type genomic DNA, mutant genomic DNA, and PpN018G07, but did not hybridize to a fragment in either of the other two BACs (Figure 5B). Thus the BAC contig, which was developed from wild-type dormant Nemared genomic DNA, spans the deletion in the mutant EVG genome. In order to determine if multiple copies of this gene sequence were characteristic of the specific peach cultivars used in this study or a more general characteristic of peach, 440 Figure 5. Southern hybridization of HindIII-digested genomic DNA from wild-type dormant (DE) and nondormant mutant (EVG) peach and of three BACs from the EVG gene contig. (A) Membranes hybridized with a DNA probe amplified from the SP6 end of BAC PpN089G02. (B) Membranes hybridized with a DNA probe amplified from the SP6 end of BAC PpN018G07. Lines represent positions of bands from the HindIII-digested k DNA size marker. EcoRI-digested genomic DNA from several peach cultivars, along with a limited selected of individuals from the Evergrowing F2 mapping population, were hybridized with the bes_018F12 MADS box probe (Figure 6). EcoRI was selected for use in the genomic analyses due to the clearer separation of hybridizing bands observed with this enzyme (see Figure 2). All wild-type dormant peach DNA analyzed had from five to seven fragments that hybridized with the MADS box probe (Figure 6). The individuals from the F2 population used to map the Evergrowing gene all showed the same pattern of hybridization, six fragments in the wild-type dormant trees and two fragments in the mutant nondormant trees (Figure 6). Additional cultivars (i.e., Carogem, Biscoe, Maycrest, Goldcrest, Contender, and Peen-To) were also analyzed and the hybridization patterns were similar to the patterns of the other wild-type dormant trees (data not shown). EcoRI-digested and HindIII-digested genomic DNA from four additional species within the Rosaceae that enter a winter dormant state were hybridized with the bes_018F12 MADS box probe to determine if these species also contained multiple copies of a similar sequence. Woody perennial species related to peach (plum [Prunus domestica L.], apricot [Prunus armeniaca L.], cherry [Prunus avium L.], and apple [Malus domestica Borkh.]) contained a number of DNA fragments hybridizing to the MADS box probe from bes_018F12B01 (Figure 7). Discussion The MADS box coding sequence from bes_018F12B01 hybridized to six fragments in EcoRI- and HindIII-digested wild-type DNA and showed polymorphism between the Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 Figure 4. Structure of the BAC contig covering the EVG gene region. Lines designating the individual BACs are scaled to show their relative sizes with respect to one another. The degree of overlap is not known. Letters corresponding to bands in the HindIII digestion of wild-type genomic DNA are show in the positions along the contig deduced from Southern hybridization (see Figure 3). Letters positioned above overlaps are found on both BACs. The parentheses enclosing A and C signify that their positions could be reversed. Bielenberg et al. A Deletion in Evergrowing Peach wild-type and mutant DNAs (Figure 2). The presence of six hybridizing bands in the EcoRI digestion and the absence of an EcoRI restriction site in the DNA probe provide confirmation that there are six bands in the HindIII-digested wild-type DNA as well, with the smallest band actually being a doublet. RFLP analysis showed the complete absence of four of the bands in the mutant. Sequence polymorphisms that result merely in the gain or loss of restriction enzyme recognition sites would be expected to produce bands of different sizes, as the restricted DNA fragment to which the probe hybridizes would be smaller or larger in size. However, in the case presented here, the complete loss of hybridization to the genomic DNA suggested that the genomic DNA to which the probe has homology was not present in the genome of the EVG mutant (Figure 2). The four MADS box genes, 441 Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 Figure 6. Southern hybridization of EcoRI-digested genomic DNA from peach cultivars of varying chilling requirements with selected individuals from the EVG F2 mapping population. Cultivar name or sample identification is at the top of each lane. F2 individuals are designated by a number and noted as either wild-type or mutant phenotype. Membranes were probed with DNA amplified from the bes_18F12B01 MADS box ORF (121 bp). Lines represent positions of bands from the HindIII-digested k DNA size marker (from top: 9.416 kb, 6.557 kb, 4.361 kb, 2.322 kb, and 2.027 kb). similar to the MADS box probe from bes_018F12B01, are therefore the first candidate genes for the Evergrowing mutation. However, sequencing of the EVG gene region will certainly uncover more candidates for this mutation. The nature of the mutation that has resulted in the loss of the four MADS box hybridizing fragments cannot be definitively determined from the experiments we present here. A number of structural rearrangements could be responsible for the absence of hybridization bands in the mutant (Figure 2). Since we have not performed comprehensive hybridizations with probes from throughout the BACs in our contig, we cannot definitively state the nature of the deletion(s). These sequences may be a series of tandem duplicated loci, of which certain pairs may be independently lost in the mutant trees. Such a series of closely linked small deletions would still potentially result in the simple 3:1 Mendelian inheritance of the Evergrowing trait (Rodriguez et al. 1994) seen in the F2 mapping population. However, a simpler explanation would be presence of a single large deletion in the mutant that results in the four missing bands (Figure 3). We have based our analyses of the bracketing of the mutation on this assumption, but are cognizant that subsequent sequencing of the genomic region of the mutant will provide ultimate proof of the structural arrangement of the region. The sequence of the MADS box probe from bes_018F12B01 was very similar to that of other MADS box transcription factors (Table 1). The most similar gene sequences were all MADS genes of the MIKC structural class ( Johansen et al. 2002), however, since the probe sequence is comprised of only the highly conserved MADS box coding region, these results may change upon sequencing of the complete genes from the EVG region. Two of the top five paralogous genes have been well studied: Arabidopsis Agamous-Like 24 (AGL24) and Tomato Jointless (LeJointless). AGL24 is involved in the vernalization pathways of Arabidopsis, which regulate the timing of the terminal meristem transition from vegetative to reproductive growth in response to cold treatments (Yu et al. 2002). LeJointless regulates the formation of abscission zones in the pedicels of tomato fruit (Mao et al. 2000). Both of these traits deal with reproductive events in annual species, while the Evergrowing mutant is disrupted in its ability to undergo winter dormancy, a vegetative trait in a perennial, indicating that there may not be useful analogies to be drawn between the three pathways, despite apparent similarities between the protein sequences. The six related MADS box coding sequences detected in the RFLP analysis appear to be grouped in a relatively small region of the genome. We confirmed this grouping by hybridizing the bes_018F12B01 MADS box probe to three HindIII-digested BACs (PpN089G02, PpN018F12, and PpN018G07), which were previously determined to form a contig through the EVG region, digested wild-type, and EVG mutant genomic DNA (Figure 3). Despite the better separation of the hybridizing MADS box probe achieved with the use of EcoRI as a restriction enzyme (Figure 2), HindIII was used as an enzyme in the BAC analysis. The Nemared BAC library was prepared using partial Journal of Heredity 2004:95(5) HindIII-digested fragments (Georgi et al. 2002), therefore digested fragments present in the genomic DNA should all be the same size in the BAC clones. Across the BAC contig, all six of the hybridizing fragments that were seen in the genomic wild-type DNA were accounted for in the BAC clones (Figure 3). Therefore the MADS box hybridizing sequences detected in the genomic Southern analysis appear to be grouped in one discrete region spanned by the BAC contig. However, it is not known whether there are full gene structures associated with the MADS box sequences or whether these sequences are simply gene fragments. Full sequencing of the BACs in the contig will be required to determine the true arrangement and structure of the genomic region. Arabidopsis contains more than 100 predicted MADS box transcription factors (Garcia-Maroto et al. 2003). Despite this high number, there is only one cluster of four MADS box genes, the MADS affecting flowering (MAF ) gene family (Ratcliffe et al. 2003). In tomato, the Rin (ripening inhibitor) locus was discovered to contain a pair of closely spaced MADS box genes that had become fused due to a deletion of 2.6 kb of intervening sequence (Vrebalov et al. 2002). The six putative MADS box genes reported here would be the first group of clustered MADS box genes identified in a tree species. The proposed arrangement of the BAC contig created for this region positions PpN089G02 and PpN018G07 overlapping with PpN018F12, but no overlap with each other (Figure 5). Therefore these BACs extend away from PpN018F12 in opposite directions (Wang et al. 2002a). The MADS box probe hybridization supported this arrangement. PpN018F12 shared two hybridization bands (D, 2.1 kb; F, 1.5 kb) with PpN089G02 that were not found in PpN018G07 (Figures 3 and 4), and PpN018F12 shared one hybridization band (E, 1.5 kb) with PpN018G07 that was not found in PpN089G02 (Figures 3 and 4). In addition, there were three bands (A, 3.3 kb; B, 2.7 kb; C, 2.1 kb) in PpN018F12 that were not represented in either PpN089G02 Table 1. The top five most similar protein sequences to the translated MADS box ORF from bes_18F12B01 using tBLASTp from the National Center for Biotechnology Information (NCBI) database e-value 4.0 7.0 9.0 9.0 2.0 3 3 3 3 3 442 10 10 10 10 10 20 20 20 20 19 Gene Species GenBank accession MADS box transcription factor MADS box transcription factor MADS box protein AGL24 MADS box transcription factor JOINTLESS Putative MADS domain transcription factor MpMADS1 Canavalia lineata Ipomoea batatas Arabidopsis thaliana Lycopersicon esculentum Magnolia praecocissima AF144623.1 AAK27150.1 CAB79364.1 AAG09811.1 BAB70736.1 Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 Figure 7. Southern hybridization of HindIII- and EcoRI-digested genomic DNA from selected species probed with DNA amplified from the bes_18F12B01 MADS box ORF (121 bp). The probe was hybridized at 508C. Lines represent positions of bands from the HindIII-digested k DNA size marker (from top: 9.416 kb, 6.557 kb, 4.361 kb, 2.322 kb, and 2.027 kb). Bielenberg et al. A Deletion in Evergrowing Peach the Clemson University Experiment Station. This material is based on work supported by the CSREES/USDA under project no. SC-2003055. References Alvarez-Buylla ER, Liljegren SJ, Pelaz S, Gold SE, Burgeff C, Ditta GS, Vergara-Silva F, and Yanofsky MF, 2000. MADS-box gene evolution beyond flowers: expression in pollen, endosperm, guard cells, roots and trichomes. Plant J 24:457–466. Arora R and Wisniewski M, 1996. Accumulation of a 60-kD dehydrin protein in peach xylem tissues and its relationship to cold acclimation. HortScience 31:923–925. Arora R, Wisniewski M, and Rowland LJ, 1996. Cold acclimation and alterations in dehydrin-like and bark storage proteins in the leaves of sibling deciduous and evergreen peach. J Am Soc Hort Sci 121:915–919. Arora R, Wisniewski ME, and Scorza R, 1992. Cold-acclimation in genetically related (sibling) deciduous and evergreen peach (Prunus persica L Batsch). 1. Seasonal changes in cold hardiness and polypeptides of bark and xylem tissues. Plant Physiol 99:1562–1568. Diaz MD, 1974. Vegetative and reproductive growth habits of evergreen peach trees in Mexico. In: XIXth International Horticultural Congress, Warsaw, Poland (Zych CC, ed). Warsaw: Wroclawska Drukarnia Naukowa; 525. Doyle JJ and Doyle JL, 1990. Isolation of plant DNA from fresh tissue. Focus 12:13–15. Fang SC and Fernandez DE, 2002. Effect of regulated overexpression of the MADS domain factor AGL15 on flower senescence and fruit maturation. Plant Physiol 130:78–89. Garcia-Maroto F, Ortega N, Lozano R, and Carmona MJ, 2000. Characterization of the potato MADS-box gene STMADS16 and expression analysis in tobacco transgenic plants. Plant Mol Biol 42:499–513. Garcia-Maroto F, Carmona MJ, Garrido JA, Vilches-Ferron M, RodriguezRuiz J, and Alonso DL, 2003. New roles for MADS-box genes in higher plants. Biol Plant 46:321–330. Georgi LL, Wang Y, Yvergniaux D, Ormsbee T, Inigo M, Reighard G, and Abbott AG, 2002. Construction of a BAC library and its application to the identification of simple sequence repeats in peach Prunus persica (L.) Batsch. Theor Appl Genet 105:1151–1158. Georgi LL, Wang Y, Reighard GL, Mao L, Wing RA, and Abbott AG, 2003. Comparison of peach and Arabidopsis genomic sequences: fragmentary conservation of gene neighborhoods. Genome 46:268–276. Johansen B, Pedersen LB, Skipper M, and Frederiksen S, 2002. MADS-box gene evolution-structure and transcription patterns. Mol Phylogenet Evol 23:458–480. Kim SH, Mizuno K, and Fujimura T, 2002. Isolation of MADS-box genes from sweet potato (Ipomoea batatas (L.) lam.) expressed specifically in vegetative tissues. Plant Cell Physiol 43:314–322. Lohmann JU and Weigel D, 2002. Building beauty: the genetic control of floral patterning. Dev Cell 2:135–142. Mao L, Begum D, Chuang HW, Budiman MA, Szymkowiak EJ, Irish EE, and Wing RA, 2000. JOINTLESS is a MADS-box gene controlling tomato flower abscission zone development. Nature 406:910–913. Mouradov A, Cremer F, and Coupland G, 2002. Control of flowering time: interacting pathways as a basis for diversity. Plant Cell 14:S111–S130. Acknowledgments Ratcliffe OJ, Kumimoto RW, Wong BJ, and Riechmann JL, 2003. Analysis of the Arabidopsis MADS AFFECTING FLOWERING gene family: MAF2 prevents vernalization by short periods of cold. Plant Cell 15:1159–1169. The authors would like to thank Dr. W. R. Okie for providing peach cultivar leaf tissue for Southern analysis and D. Ouellette and K. Brock for invaluable plant culture assistance. This report is Technical Contribution no. 4982 of Rodriguez J, Sherman WB, Scorza R, Wisniewski M, and Okie WR, 1994. Evergreen peach, its inheritance and dormant behavior. J Am Soc Hort Sci 119:789–792. 443 Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 or PpN018G07, confirming that there was a middle region of the contig where PpN018F12 had no overlap with the other two BACs (Figures 3 and 4). The hybridizing bands were assigned a putative order within the contig based on the hybridization results (from the SP6 end of PpN089G02 to the SP6 end of PpN018G07): D, F, A/C or C/A, B, and E (Figure 4). The limit of the deletion present in the EVG trees was bracketed. Polymorphism between the wild-type and mutant DNA for a probe, which falls within the BAC contig (bes_18F12B01 MADS box), showed that our BAC contig covered the region affected by the mutation (Figure 3). The lack of polymorphism observed between wild-type and mutant DNA for probes covering the ends of the BAC contig (Figure 5) indicated that these probes hybridized outside the region affected by the mutation and that there was at least one HindIII restriction site unaffected by the mutation between the probes used and the mutation. Therefore the mutation (i.e., deletion) appeared to be contained wholly within the region spanned by the three overlapping BAC clones. The size of the region that may contain the EVG gene can be estimated from the size of the BAC contig, which spans the region. PpN089G02, PpN018F12, and PpN018G07 have been sized to approximately 70 kb, 60 kb, and 50 kb, respectively, by NotI digestion and subsequent pulse field gel electrophoresis (data not shown). Therefore, assuming minimal overlap between BACs, the mutation has been narrowed to a region of approximately 180 kb. Gene sequences hybridizing to the MADS box ORF found in the EVG locus were present in a wide variety of peach cultivars (Figure 6). Some variation in hybridizing fragment size exists between the cultivars, but the number of fragments was relatively similar between wild-type cultivars (Figure 7). The cultivars used in this analysis display a wide range of behaviors in dormancy traits such as chilling requirement, date of bud break, and bloom date. None of these traits appeared to correlate with the number of hybridizing bands in this analysis. In addition, close relatives of peach in the genus Prunus also contain a number of DNA sequences similar to the bes_018F12B01 MADS box probe (Figure 6). We determined that the EVG gene region contains a number of potential MADS box-type transcription factors. A deletion exists in the EVG mutant trees in this region, which affects at least four of the MADS box-type sequences. We have bracketed this possible deletion using RFLP analysis and showed that it is likely contained in a segment of the genome no larger than 180 kb. Complete sequencing and annotation of the three BACs in the contig will allow identification of the structural nature of the deletion and undoubtedly increase the number of candidate genes. Journal of Heredity 2004:95(5) Rosin FM, Hart JK, Van Onckelen H, and Hannapel DJ, 2003. Suppression of a vegetative MADS box gene of potato activates axillary meristem development. Plant Physiol 131:1613–1622. Rozen S and Skaletsky HJ, 2000. Primer3 on the WWW for general users and for biologist programmers. In: Bioinformatics methods and protocols: methods in molecular biology (Misener S, eds).Totowa, NJ: Humana Press; 365–386. Sambrook J and Russell DW, 2002. Molecular cloning: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. Thompson MM, Smith DC, and Burgess JE, 1985. Non-dormant mutants in a temperate tree species, Corylus avellana L. Theor Appl Genet 70: 687–692. van der Linden CG, Vosman B, and Smulders MJM, 2002. Cloning and characterization of four apple MADS box genes isolated from vegetative tissue. J Exp Bot 53:1025–1036. Wang Y, 2002. Characterizing the evergrowing phenotype and mapping the Evergrowing gene in peach (PhD dissertation). Clemson, SC: Clemson University. 444 Wang Y, Georgi LL, Reighard GL, Scorza R, and Abbott AG, 2002b. Genetic mapping of the evergrowing gene in peach Prunus persica (L.) Batsch. J Hered 93:352–358. Werner DJ and Okie WR, 1998. A history and description of the Prunus persica plant introduction collection. HortScience 33:787–793. Yu H, Xu YF, Tan EL, and Kumar PP, 2002. AGAMOUS-LIKE 24, a dosage-dependent mediator of the flowering signals. Proc Natl Acad Sci USA 99:16336–16341. Zhang HM and Forde BG, 2000. Regulation of Arabidopsis root development by nitrate availability. J Exp Bot 51:51–59. Received September 15, 2003 Accepted April 1, 2004 Corresponding Editor: Reid Palmer Downloaded from http://jhered.oxfordjournals.org/ at Pennsylvania State University on February 21, 2013 Vrebalov J, Ruezinsky D, Padmanabhan V, White R, Medrano D, Drake R, Schuch W, and Giovannoni J, 2002. A MADS-box gene necessary for fruit ripening at the tomato ripening-inhibitor (Rin) locus. Science 296:343–346. Wang Y, Garay L, Georgi LL, Reighard GL, Scorza R, and Abbott AG, 2002a. Development of bacterial artificial chromosome contigs in the Evergrowing gene region in peach [Prunus persica (L.) Batsch]. Acta Hort 592:183–189.