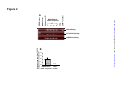
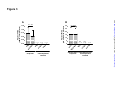
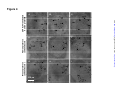
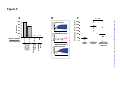
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Blood First Edition Paper, prepublished online November 22, 2013; DOI 10.1182/blood-2013-08-520478 Platelet gene therapy corrects the hemophilic phenotype in immunocompromized hemophilia A mice transplanted with genetically manipulated human cord blood stem cells Qizhen Shi1,2,3,4, Erin L. Kuether1,2,3, Yingyu Chen2,5, Jocelyn A. Schroeder1,2,3, Scot A. Fahs2, and Robert R. Montgomery1,2,3, 4 1Department of Pediatrics, Medical College of Wisconsin; 2Blood Research Institute, BloodCenter of Wisconsin; 3Children’s Research Institute, Children’s Hospital of Wisconsin; 4MACC Fund Research Center, Milwaukee, WI 53226, USA; 5Fujian Institute of Hematology, Fujian Provincial Key Laboratory of Hematology, Fujian Medical University Union Hospital, Fuzhou, Fujian, China. Running title: Human platelet gene therapy of hemophilia A Address correspondence: Qizhen Shi, MD, PhD. Department of Pediatrics, Medical College of Wisconsin, 8701 Watertown Plank Road, Milwaukee, WI 53226. Tel: 414937-3853; Fax: 414-937-6811; e-mail: [email protected] 1 Copyright © 2013 American Society of Hematology From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Key Points • Platelet-specific lentiviral gene delivery to human HSCs can efficiently introduce therapeutic levels of FVIII expression in human platelets • Human platelet-derived FVIII can ameliorate the hemophilic phenotype in an immunocompromised hemophilia A mouse model. 2 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Abstract Our previous studies have demonstrated that platelet-FVIII (2bF8) gene therapy can improve hemostasis in hemophilia A mice even in the presence of inhibitory antibodies, but none of our studies has targeted human cells, only mouse cells. Here we evaluated the feasibility for lentivirus (LV)-mediated human platelet gene therapy of hemophilia A. Human platelet-FVIII expression was introduced by 2bF8LV-mediated transduction of human cord blood (hCB) CD34+ cells followed by xeno-transplantation into immunocompromised NSG mice or NSG mice in a FVIIInull background (NSGF8KO). Platelet-FVIII was detected in all recipients that received 2bF8LV-transduced hCB cells as long as human platelet chimerism persists. Electron microscopy demonstrated that FVIII was colocalized with VWF in 2bF8LV-transduced human platelets. All NSGF8KO recipients (n = 7) that received 2bF8LV-transduced hCB cells survived tail clipping if animals had greater than 2% of platelets derived from 2bF8LV-transduced hCB cells, while 5 of 7 survived when human platelets were 0.3-2%. Whole blood clotting time analysis confirmed that hemostasis was improved in NSGF8KO mice that received 2bF8LV-transduced hCB cells. We demonstrate for the first time the feasibility of 2bF8LV gene delivery to human HSCs to introduce FVIII expression in human platelets and that human platelet-derived FVIII can improve hemostasis in hemophilia A. Key words: Platelet; Gene therapy; hemophilia A; Human cells 3 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Introduction Hemophilia A is a congenital bleeding disorder caused by a deficiency of factor VIII (FVIII). Protein replacement therapy using either plasma derived or recombinant FVIII is effective for the treatment of hemophilia A patients. However, it is expensive and requires frequent infusions owing to the short half-life of the protein. Furthermore, up to 35% of patients will develop anti-FVIII inhibitory antibodies, referred to as inhibitors, after exogenous FVIII replacement therapy.1-3 The clinical hallmark of inhibitor development in hemophilia A patients is failure to respond to routine replacement therapy for bleeding episodes.3-6 Gene therapy is an attractive strategy for hemophilia A treatment. The goal of gene therapy is to introduce long-term expression of therapeutic levels of FVIII in vivo by genetically modifying the target cells resulting in a cure of the disease. Although substantial progress has been achieved in the past decade, potential development of an immune response to transgene product or vector remains a significant concern in genetic therapy.7-9 We have developed a novel clinically translatable platelet-targeted gene therapy approach using lentiviral gene delivery to hematopoietic stem cells (HSCs), in which FVIII expression is under control of the platelet-specific glycoprotein (GP) IIb promoter (2bF8).10 Our previous studies have demonstrated that 2bF8 lentivirus (2bF8LV)-mediated platelet-specific gene therapy can efficiently introduce therapeutic levels of platelet-FVIII in hemophilia A mice with neither inhibitory nor non-inhibitory antibody development.10 Further studies have demonstrated that sustained therapeutic levels of platelet-FVIII are obtained while inhibitor titers decline with time after 2bF8 4 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. gene therapy in hemophilia A mice with pre-existing anti-FVIII immunity.11 However, this approach has not been studied in human cells. Since our ultimate goal is to express FVIII in hemophilia A patient’s platelets, the questions we addressed in the current study include: 1) whether human hematopoietic stem cells (hHSCs) can be transduced by 2bF8LV; 2) whether 2bF8LV-transduced hHSCs can normally give rise to blood cells including the platelet lineage; 3) whether 2bF8LV-mediated gene transfer can efficiently introduce FVIII expression in human platelets; and 4) whether human platelet-derived FVIII can correct the hemophilic bleeding diathesis. We demonstrate for the first time the feasibility of 2bF8 lentiviral gene delivery to hHSCs to introduce FVIII expression in human platelets and that human platelet-derived FVIII can improve hemostasis in hemophilia A. 5 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Materials and methods Mice. Immunocompromised NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ (NSG) mice, which combine the features of the NOD background, the severe combined immune deficiency mutation (scid) and IL2 receptor gamma chain (IL2rg) deficiency,12 were purchased from the Jackson Laboratory (Bar Harbor, ME). The NSG mice have been shown to support greater engraftment of hHSCs than all other strains.12;13 The immunocompromised hemophilia A mouse model (NSGF8KO) was generated by our laboratory by crossing NSG mice into the FVIIInull background. The FVIIInull mice (in C57BL/6 x 129/SV background) were generated by targeted disruption of exon 17 of the F8 gene.14 All animals were kept in non-specific pathogen free microisolator cages at the animal facilities operated by the Medical College of Wisconsin. Animal studies were performed according to a protocol approved by the Institutional Animal Care and Use Committee of the Medical College of Wisconsin. Virus production, purification, and titering. The lentiviral construct, pWPT-2bF8, was generated as previously described.10 Recombinant lentivirus was generated from HEK 293T cells by transient transfection. The procedures for virus production and purification were described in previous reports.10;15 Lentivirus-mediated transduction of human cord blood (hCB) CD34+ cells. Human cord blood (hCB) CD34+ cells were purchased from ALLCELLs (Emeryville, CA) and transduced with 2bF8LV using a protocol similar to that described in previous reports.10;15 The details are provided in the supplemental materials. 6 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Xeno-transplantation. Six to seven week old NSG or NSGF8KO mice were conditioned for xeno-transplantation with busulfan treatment. Busulfan (6 mg/mL) (DSM Pharmacenticals, Inc. Greenville, NA) was diluted 1: 4 in PBS and administered intravenously at 25 mg/kg per day on days -2 and -1. Twenty-four hours after busulfan treatment, 1-2 x 106 2bF8 LV-transduced hCB CD34+ cells were transplanted in a volume of 300 μL of QBSF-60 media per mouse by retro-orbital vein injection. Untransduced cells were transplanted into controls. The animals that received hCB CD34+ cell transplantation (hCBT) are defined as a humanized model. Human cell chimerism analysis. Blood samples from animals were collected by tail bleeds and blood counts were performed using the Scil Vet ABC Plus blood counter (Scil Animal Care Company, Gurnee, IL). The details are provided in the supplemental materials. PCR and quantitative real-time PCR (qPCR) analysis. PCR and qPCR were performed as previously described.10;11 The details are provided in the supplemental materials. Immunogold electron microscopy (EM). EM studies were performed by Clive W. Wells in The Electron Microscopy Facility at the Medical College of Wisconsin. Isolated platelets were processed and stained for FVIII protein as previously described.16 The details are provided in the supplemental materials. 7 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. FVIII activity (FVIII:C) assays. Platelets and plasma were isolated as previously described.17 Functional FVIII:C in platelet lysates or plasma was quantitated by a modified FVIII chromogenic assay as previously described.16;17 Recombinant human Bdomain deleted FVIII (rhFVIII) (Xyntha, Wyeth Pharmaceuticals, Collegeville, PA) was used as the standard. Platelets and plasma from NSG mice, C57BL6 mice, and humans were used as controls. Assessment of phenotypic correction. To evaluate whether 2bF8LV-transduced human platelets can improve hemostasis in hemophilic mice, the tail clip survival test and rotational thromboelastometry (ROTEM) analysis were used. The tail clip tests were performed between 4-6 weeks after transplantation as previously reported.11;16;17 ROTEM analysis of whole blood clotting time (CT) was performed between 6-8 weeks after transplantation as described in our previous report.18 2bF8LV integration site analysis. 2bF8LV integration site analysis was performed on genomic DNA extracted from peripheral blood using non-restrictive LAM-PCR (nrLAMPCR), which is faster and more cost-efficient than LAM-PCR as reported.19 The details are provided in the supplemental materials. Statistical analysis. Data are presented as mean ± SD, and the significance of differences was evaluated by 2-tailed Student t-test. A value of P < .05 was considered statistically significant. 8 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Results Human cell chimerism. Using our culture conditions for in vitro transduction of hCBderived hHSCs, we were able to maintain both a similar cell number and the “stemness” (as indicated by CD34 expression) compared to the initial hCB sample. Greater than 90% of the cells were still CD34+ after 2bF8LV-transduction. To determine whether 2bF8LVtransduced hCB CD34+ cells can differentiate into human platelets in our xenograft model, flow cytometry was used to analyze peripheral blood cells from transplanted recipients. No human cells could be detected 1 week after transplantation, but both human platelets and leukocytes were detected in NSG mice that received 2bF8LVtransduced hCB CD34+ cells at later time points. Three weeks after transplantation, 48.9 ± 13.5% (n = 10) of total platelets and 73.2 ± 9.6% (n = 6) of total leukocytes in peripheral blood were of human origin (Figure 1 A-D). Human platelet chimerism dropped to 7.4 ± 4.3% (n = 6) at week 5 while leukocyte chimerism was still maintained at greater than 47% (Figure 1C and D). Total human platelet number was calculated based on the whole blood cell count and the percent of chimerism determined by flow cytometry analysis. The peak levels of human platelet chimerism in recipients occurred 2-3 weeks after transplantation, while the animals exhibited murine cell thrombocytopenia at that time point (Figure 1E). There were 56.3 ± 27.0 x 103/ μL human platelets in the blood at 2 weeks, but dropped to 12.4 ± 11.9 x 103/ μL at 5 weeks and 1.9 ± 0.6 x 103/ μL at 12 weeks after transplantation. Mouse platelet number was 9.3 ± 5.5 x 103/ μL at 2 weeks after transplantation, which is only about 1% of the platelet number of NSG mice. The mouse platelet number started 9 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. to increase at 3 weeks after transplantation, but never recovered to pre-transplantation levels during the study course (Figure 1E). In contrast to platelets, the peak levels of human leukocyte chimerism occurred at 7 weeks after transplantation and were maintained around that level for the rest of the study course. When we looked at murine leukocytes, the animals had developed severe murine leukopenia at 3 to 5 weeks after transplantation. Compared to untreated NSG mice, there were only about 3% of mouse leukocytes remaining at 3 weeks after transplantation. Mouse leukocyte levels started to recover after 7 weeks. These levels were approximately 21% of normal in NSG mice and were maintained around that level onward during the study course (Figure 1F). Analysis of 2bF8LV transduction in engrafted human cells. To investigate the transduction of viably engrafted 2bF8LV-transduced hCB-derived HSCs, PCR was used to detect the 2bF8 transgene in peripheral blood cell-derived DNA from NSG recipients. Representative results from recipients at least 3 weeks after transplantation are shown in Figure 2A. The 2bF8 transgene was detected in all NSG mice that received 2bF8LV transduced human cord blood CD34+ cells. To determine the average copy number of 2bF8 proviral DNA in human cells, qPCR was used to quantify the 2bF8 expression cassette, normalized to the human GAPDH gene. There were 1.88 ± 0.52 copies of the 2bF8 transgene per human cell in NSG mice that received 2bF8LV-transduced hCB CD34+ cells (Figure 2B). As expected, no 2bF8 proviral DNA was detected in NSG control mice. 10 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Analysis of FVIII expression in 2bF8 LV-transduced human platelets. The levels of platelet-FVIII expression were determined by a chromogenic assay of platelet lysates from recipients at least 3 weeks after transplantation. Functional FVIII activity was detected in the NSG recipients that received 2bF8LV-transduced hCB CD34+ cells, at an average of 0.76 ± 0.43 mU/108 total platelets (n = 6) (Figure 3A). This level of plateletFVIII corresponds to about 17.77 ± 10.36 mU/108 human platelets when calculated according to chimerism levels measured by flow cytometry (Figure 3B). As expected, no FVIII was detected in platelet lysates from control mice (NSG and C57BL6) and human control platelets. Localization of human FVIII transgene protein expression was determined by immunogold electron microscopy. Transgene hFVIII protein was labeled with small colloidal gold particles and endogenous VWF was labeled with large colloidal gold particles. Transgene hFVIII protein was detected in 2bF8LV-transduced human platelets recovered from transplanted NSG mice and was co-localized with endogenous VWF in platelet α-granules (Figure 4 A-C). No FVIII was detected in platelets from untransplanted NSG mice or normal human individuals although a normal distribution of endogenous human VWF (Figure 4 D-F) or murine VWF (Figure 4 G-I) was observed. Insertion site analysis. To investigate the chromosomal insertion sites of 2bF8LV in human cells, we used non-restrictive LAM-PCR. We identified 19 insertion sites in 5 of the NSG recipients and found 13 insertion sites in the introns of genes, 2 in the exons, and 4 in intergenic regions (Table 1). Similar to what we have found in transduced mouse cell studies,11 the integration of 2bF8LV into the first intron of human genes appears to 11 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. have occurred at a higher frequency (27%) than other areas within genes. The frequency of sense versus anti-sense orientation of 2bF8 transgene within human genes appears to be approximately, equal at 47% and 53%, respectively. No identified integration sites were located within known proto-oncogenes in human hematopoiesis. Evaluation of clinical efficacy of human platelet-derived FVIII. Using the NSG mouse model, we have examined the feasibility of targeting FVIII expression to human platelets by 2bF8 lentiviral gene transfer to hCB-derived hHSCs. To explore whether 2bF8LV-transduced human platelets can improve the bleeding diathesis in hemophilia A, we generated an immunocompromised hemophilia A mouse model by crossing NSG and FVIIInull to produce NSGF8KO mice in which the genetic background is a mixture of NOD, 129/SV, and C57BL/6. 2bF8LV-transduced hCB-derived CD34+ cells were transplanted into NSGF8KO mice pre-conditioned with busulfan. Because the IL2rg gene is relatively near the F8 locus (the distance between the two genes is 7.5 centimorgan) on the X-chromosome, there was some difficulty with crossing to achieve linkage of the defective IL2rg gene with the FVIII knockout gene, but after multiple matings, we successfully identified a cross-over between these two genes and obtained NSGF8KO mice for our studies. We found that human platelet chimerism was markedly decreased in recipients that had lost the NOD phenotype during breeding, while leukocyte chimerism was less affected. Three weeks after transplantation, 20.9 ± 23.9% (n = 14) of total platelets in NSGF8KO recipients were of human origin and only 2.1 ± 1.7% (n = 12) at week 5, significantly lower than in NSG recipients (P < .01) (Figure 12 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. 1C). In contrast, human leukocyte chimerism in NSGF8KO recipients was not significantly different from the NSG recipients (Figure 1D). As expected, no FVIII:C was detected in plasmas from NSGF8KO recipients (data not shown). Functional platelet-FVIII activity was detected in NSGF8KO recipients at an average the level of 0.56 ± 0.42 mU/108 total mouse plus human platelets (Figure 3A), which corresponded to 17.72 ± 8.65 mU/108 human platelets (Figure 3B) when calculated based on human platelet chimerism as determined by flow cytometry. There was no significant difference in the level of platelet-FVIII expression between NSG and NSGF8KO groups after xeno-transplantation of 2bF8LV-transduced hCB (Figure 3A and B). Tail clip survival tests were used to assess phenotypic correction of the FVIIInull coagulation defect in NSGF8KO mice. Four to 6 weeks after transplantation, all recipients survived tail clipping if greater than 2% of platelets were derived from 2bF8LV-transduced hCB CD34+ cells; 5 of 7 survived when human platelets were between 0.3% and 2%. In contrast, none of the untransplanted NSGF8KO mice or NSGF8KO mice that received untransduced hCB CD34+ cells survived tail clipping (Figure 5A). To further assess whether 2bF8LV-transduced human platelets improve hemostasis in hemophilia A mice, ROTEM analysis was used to analyze whole blood CT (Figure 5B and C). Six to 8 weeks after transplantation, the whole blood CT was 1296 ± 234 seconds (n = 7) in NSGF8KO/2bF8LV-hCBT recipients, in which human platelet chimerism was 0.4-5.5% at the time of the ROTEM analysis. This whole blood CT was significantly shorter than that obtained from NSGF8KO control mice [2792 ± 220 13 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. seconds (n = 10)], confirming that hemostasis was improved in NSGF8KO mice that received 2bF8 LV-transduced hCB CD34+ cells. Taken together, our results show that 2bF8 lentiviral gene delivery to hHSCs can efficiently introduce FVIII expression in human platelets and that 2bF8LV-transduced human platelets can improve hemostasis in a hemophilia A mouse model. 14 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Discussion Previous studies by our group10;11;16;17 and others20-23 using mouse models have demonstrated that platelet-targeted expression of FVIII may be a promising approach for gene therapy of hemophilia A and hemophilia A with inhibitors. In the current study, we established a novel immunocompromised hemophilia A mouse model and used humanized mouse models to explore the feasibility and efficacy of targeting FVIII expression to human platelets for hemophilia A gene therapy. FVIII expression was introduced by ex vivo transduction of hCB-derived hHSCs followed by xenotransplantation into immunocompromised hemophilia A mice. We conclude that 2bF8 lentiviral gene delivery to hHSCs can efficiently introduce functional FVIII expression in human platelets and that human platelet-derived FVIII can ameliorate the bleeding diathesis in hemophilia A mice. Human thrombocytopoiesis in the 2bF8LV-genetically modified humanized NSG model. Our study demonstrated that 2bF8LV can efficiently transduce hCB CD34+ cells, resulting in successfully genetically manipulated hHSCs. In preliminary experiments, we transplanted the transduced hCB CD34+ cells into NSG mice preconditioned with a sublethal irradiation dose of 200 cGy or 300 cGy total body irradiation, but the mortality was extremely high. It has been reported that busulfan chemotherapy is an efficient preconditioning regimen to achieve successful engraftments in humanized models for white blood cell related studies.24-27 Thus, we for the first time used busulfan chemotherapy as a preconditioning regimen for xeno-transplants to study the feasibility of 2bF8 gene delivery to hHSCs and the clinical efficacy of human platelet-derived FVIII 15 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. in gene therapy of hemophilia A. We found that the transduced repopulating hHSCs were able to successfully engraft into immunocompromised animals preconditioned with busulfan and give rise to human blood cells, including platelets that contain functional FVIII activity. Since the final target for FVIII expression in our gene therapy approach is platelets, it is important to investigate whether 2bF8LV-transdcued hHSCs can differentiate into human platelets. Human platelet chimerism peaked at 2-3 weeks after xeno-transplantation, when up to 93% of platelets were of human origin, but by week 5 they had dropped to 3-18% although leukocyte chimerism was still maintained at greater than 47%. The rapid reduction of human platelet chimerism in NSG recipients also occurs in non-transduced transplants (data not shown), so it is not due to 2bF8 genetic modification. This is a common problem in studies of human thrombocytopoiesis in immunocompromised mouse models and the cause is still unknown.28-31 There was a small population that was positive for both human and mouse platelet markers, which could be due to low level platelet activation during sample processing resulting in human and mouse platelet association. We had monitored some animals up to 7 months after transplantation and found that leukocyte chimerism was sustained, but the reduction of human platelet chimerism between weeks 3 and 7 was remarkable, decreasing by 70 to 95%. 2bF8 proviral DNA was consistently detected in peripheral blood cell-derived DNA at all time points during the study course, indicating that long-term repopulating hHSCs were successfully engrafted and were genetically modified by 2bF8LV. Although the temporal window for 16 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. studying human platelets derived from 2bF8LV-transduced hCBT in the NSG mouse model was narrow, there were up to 133 x 103 human platelets/μL circulating in the peripheral blood in xeno-transplanted recipients with the highest chimerism between weeks 2 and 3, nearly reaching the range of normal platelet number in humans (150 - 450 x 103/μL). Since high levels of white blood cell chimerism were sustained in 2bF8LV-transduced recipients, we suspect the quick reduction of human platelets in xeno-transplant recipients may be due to incompatibility of murine molecular signals involving thrombocytopoiesis or maintenance of the proper hematopoietic microenvironment, for human platelet production. While we did infuse rhTPO into 3 of the NSG mice that received 2bF8LVtransduced hCB CD34+ cells, human platelet chimerism did not significantly increase (data not shown). Recent studies done by Hu et al have demonstrated that poor human platelet reconstitution in their xeno-transplantion models is mainly caused by macrophage-mediated rejection.32 The macrophage rejection mechanism cannot explain the phenotype of blood cells in our xeno-transplanted recipients, in which human platelets peak in the early phase (2 weeks) when there were 12% murine endogenous leukocytes including macrophages and decrease dramatically at 5 weeks when murine endogenous leukocytes are barely detectable (0.9%). These results indicate that there are other mechanisms, which are still unknown, playing roles in human thrombocytopoiesis in immunocompromised mouse models. 17 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Human thrombocytopoiesis in the 2bF8LV-genetically modified humanized NSGF8KO model. While we obtained high levels of human platelet chimerism in NSG mice that received 2bF8LV-transduced hCB CD34+ cell, the human platelet chimerism in NSGF8KO recipients was dramatically lower. This might be owing to loss of the NOD background during crossbreeding of NSG mice to the FVIIInull background as well as scid “leakiness”.26 It is known that the NOD background contributes to the absence of hemolytic complement, defective macrophages, and enhanced human HSC engraftment.33-35 For genotyping of NSGF8KO mice, we used PCR to detect the FVIIInull allele, the scid mutation, and the IL-2Rγ(null) gene. However, there is currently no genotyping approach available to track the NOD background. Thus, the genetic background of NOD was likely diluted during crossbreeding of NSG into FVIIInull, so macrophages from resulting NSGF8KO mice might function normally and phagocytize human platelets in xeno-transplanted recipients. The scid mutation prevents maturation of T and B cells,36 but scid leakiness can result in the production of some functional B and T cells as the mice age,37 causing a rejection of xeno-grafts. It has been reported that such leakiness is higher in mice housed under nonspecific pathogen free conditions, which is the type of facility in which we house our mice, and leakiness is generally higher on the C57BL/6J genetic background.38-40 The FVIIInull mice crossed with NSG mice were on a mixed genetic background of C57BL/6 and 129/SV, so NSGF8KO mice inherited a mixed genetic background of C57BL/6, 129/SV, and NOD. Backcrossing NSGF8KO mice to a pure NOD background will need to be pursued in future studies. 18 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. FVIII expressed in human platelets has functional activity. Although human platelet reconstitution was not sustained in 2bF8LV-transduced hCBT recipients, platelets produced by 2bF8LV-transduced hCB CD34+ cells contained functional FVIII activity. When the levels of platelet-FVIII expression were converted to average per human platelet values according to chimerism levels determined by flow cytometry, the amount of FVIII in 2bF8LV-transduced human platelets (17.77 mU/108 platelets) is significantly greater than those obtained from transduced or transgenic mouse platelets (1.56 mU/108 platelets or 1.41 mU/108 platelets, respectively) in our previous studies.11;16 This phenomena was consistent for both NSG and NSGF8KO mice that received 2bF8LV-transduced hCB CD34+ cells. While it is still unclear why the average amount of FVIII in human platelets derived from 2bF8 genetically engineered hHSCs is greater than in mouse HSCs, there are three potential explanations. First, human platelets are larger than mouse platelets. Human platelets are approximately 2.0-4.0 μm in diameter with a mean volume of 7.8 fL, whereas mouse platelets are 1.0-2.0 μm in diameter with a mean volume of 5.5 fL.41;42 Thus, each 2bF8LV-transduced human platelet has a higher capacity to carry cargo including transgene FVIII protein than mouse platelets. Secondly, 2bF8LV may transduce human cord blood-derived HSCs more efficiently than mouse bone marrowderived HSCs. The vector that we use to deliver 2bF8 transgene to HSCs is a second generation human immunodeficiency virus type 1-based vector, which has been reported to transduce human cells more efficiently than their mouse counterparts.43-45 Indeed, the average copy number of 2bF8 proviral DNA per human cell was significantly higher than 19 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. that obtained from the transduced mouse cells in our previous reports.10;11 A third reason for potentially higher expression (not infection) is that human platelets contain human VWF which works better with human FVIII. Our previous studies have demonstrated that hFVIII expressed in mouse platelets can rescue the bleeding diathesis in hemophilia A mice.10;11;16;46 In the current study using a humanized NSGF8KO model we demonstrated that 2bF8LV can efficiently introduce FVIII expression in human platelets and that human platelet-derived FVIII can restore hemostasis in a hemophilia A mouse model as long as human platelet persist. The bleeding diathesis was significantly improved even with a small proportion of platelets (0.3% - 2%) derived from 2bF8LV-transduced hCB CD34+ cells. Our data suggest that sufficiently high transduction efficiencies can be practically achieved to ensure adequate transgene delivery to hHSCs to produce therapeutic levels of platelet-FVIII using 2bF8 lentiviral gene transfer. In conclusion, we have demonstrated for the first time that 2bF8 lentiviral gene delivery to hHSCs can efficiently introduce therapeutic levels of FVIII expression in human platelets and that this can ameliorate the hemophilic phenotype in an immunocompromised hemophilia A mouse model. Our data strongly suggest that 2bF8 lentiviral gene delivery to hHSCs can be a promising approach for gene therapy of hemophilia A. In the scenario of a hemophilia A patient, autologous cells from cord blood or mobilized HSCs will be transduced with 2bF8LV ex vivo followed by autotransplantation of the transduced cells. 20 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Acknowledgments We thank H. Kazazian at the University of Pensylvania School of Medicine for FVIIInull mice. This work was supported by National Institutes of Health grants HL-102035 (QS) and HL-44612 (RRM), Hemophilia Association of New York grant (QS), and generous gifts from the Children’s Hospital Foundation (QS) and MACC Fund (QS). Authorship Contribution: Q. S. designed and performed research, analyzed data, and wrote the manuscript; E. L. K. and Y. C. performed research and analyzed data; J. A. S., and S. A. F. performed research, analyzed data, and made comments to the manuscript; R. R. M. helped in the design of the project and made comments to the manuscript. Conflict-of-interest disclosure: The authors declare no competing financial interests. 21 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Reference List (1) Hay CR, Brown S, Collins PW, Keeling DM, Liesner R. The diagnosis and management of factor VIII and IX inhibitors: a guideline from the United Kingdom Haemophilia Centre Doctors Organisation. Br J Haematol 2006;133(6):591-605. (2) Gouw SC, van den Berg HM, le CS, van der Bom JG. Treatment characteristics and the risk of inhibitor development: a multicenter cohort study among previously untreated patients with severe hemophilia A. J Thromb Haemost 2007;5(7):1383-1390. (3) Wong T, Recht M. Current options and new developments in the treatment of haemophilia. Drugs 2011;71(3):305-320. (4) Gouw SC, van der Bom JG, Marijke van den BH. Treatment-related risk factors of inhibitor development in previously untreated patients with hemophilia A: the CANAL cohort study. Blood 2007;109(11):4648-4654. (5) Ingerslev J, Sorensen B. Parallel use of by-passing agents in haemophilia with inhibitors: a critical review. Br J Haematol 2011;155(2):256-262. (6) Sultan Y, Algiman M. Treatment of factor VIII inhibitors. Blood Coagul Fibrinolysis 1990;1(2):193-199. (7) High KA. Gene therapy for haemophilia: a long and winding road. J Thromb Haemost 2011;9 Suppl 1:2-11. (8) Chuah MK, Nair N, Vandendriessche T. Recent progress in gene therapy for hemophilia. Hum Gene Ther 2012;23(6):557-565. (9) Scott DW, Lozier JN. Gene therapy for haemophilia: prospects and challenges to prevent or reverse inhibitor formation. Br J Haematol 2012;156(3):295-302. (10) Shi Q, Wilcox DA, Fahs SA et al. Lentivirus-mediated platelet-derived factor VIII gene therapy in murine haemophilia A. J Thromb Haemost 2007;5(2):352-361. (11) Kuether EL, Schroeder JA, Fahs SA et al. Lentivirus-mediated platelet gene therapy of murine hemophilia A with pre-existing anti-factor VIII immunity. J Thromb Haemost 2012;10(8):1570-1580. (12) Spranger S, Frankenberger B, Schendel DJ. NOD/scid IL-2Rg(null) mice: a preclinical model system to evaluate human dendritic cell-based vaccine strategies in vivo. J Transl Med 2012;10:30. (13) McDermott SP, Eppert K, Lechman ER, Doedens M, Dick JE. Comparison of human cord blood engraftment between immunocompromised mouse strains. Blood 2010;116(2):193-200. 22 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. (14) Bi L, Lawler AM, Antonarakis SE, High KA, Gearhart JD, Kazazian HH, Jr. Targeted disruption of the mouse factor VIII gene produces a model of haemophilia A. Nat Genet 1995;10(1):119-121. (15) Shi Q, Wilcox DA, Morateck PA, Fahs SA, Kenny D, Montgomery RR. Targeting platelet GPIbalpha transgene expression to human megakaryocytes and forming a complete complex with endogenous GPIbbeta and GPIX. J Thromb Haemost 2004;2(11):1989-1997. (16) Shi Q, Wilcox DA, Fahs SA et al. Factor VIII ectopically targeted to platelets is therapeutic in hemophilia A with high-titer inhibitory antibodies. J Clin Invest 2006;116(7):1974-1982. (17) Shi Q, Fahs SA, Wilcox DA et al. Syngeneic transplantation of hematopoietic stem cells that are genetically modified to express factor VIII in platelets restores hemostasis to hemophilia A mice with preexisting FVIII immunity. Blood 2008;112(7):2713-2721. (18) Zhang G, Shi Q, Fahs SA, Kuether EL, Walsh CE, Montgomery RR. Factor IX ectopically expressed in platelets can be stored in alpha-granules and corrects the phenotype of hemophilia B mice. Blood 2010;116(8):1235-1243. (19) Paruzynski A, Arens A, Gabriel R et al. Genome-wide high-throughput integrome analyses by nrLAM-PCR and next-generation sequencing. Nat Protoc 2010;5(8):1379-1395. (20) Yarovoi HV, Kufrin D, Eslin DE et al. Factor VIII ectopically expressed in platelets: efficacy in hemophilia A treatment. Blood 2003;102(12):4006-4013. (21) Greene TK, Wang C, Hirsch JD et al. In vivo efficacy of platelet-delivered, high specific activity factor VIII variants. Blood 2010;116(26):6114-6122. (22) Gewirtz JE, Rauova L, Poncz M. Characterizing Factor VIII Inhibitor Resistance of Platelet-Delivered Factor VIII in the Treatment of Hemophilia A. [abstract]Gewirtz JE, Rauova L, Poncz M. Blood 2007;110:240a (23) Damon AL, Scudder LE, Gnatenko DV et al. Altered bioavailability of plateletderived factor VIII during thrombocytosis reverses phenotypic efficacy in haemophilic mice. Thromb Haemost 2008;100(6):1111-1122. (24) Robert-Richard E, Ged C, Ortet J et al. Human cell engraftment after busulfan or irradiation conditioning of NOD/SCID mice. Haematologica 2006;91(10):1384. (25) Choi B, Chun E, Kim M et al. Human B Cell Development and Antibody Production in Humanized NOD/SCID/IL-2Rgamma(null) (NSG) Mice Conditioned by Busulfan. J Clin Immunol 2011;31(2):253-264. 23 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. (26) Hayakawa J, Hsieh MM, Uchida N, Phang O, Tisdale JF. Busulfan produces efficient human cell engraftment in NOD/LtSz-Scid IL2Rgamma(null) mice. Stem Cells 2009;27(1):175-182. (27) Singh M, Singh P, Gaudray G et al. An improved protocol for efficient engraftment in NOD/LTSZ-SCIDIL-2Rgammanull mice allows HIV replication and development of anti-HIV immune responses. PLoS One 2012;7(6):e38491. (28) Perez LE, Rinder HM, Wang C, Tracey JB, Maun N, Krause DS. Xenotransplantation of immunodeficient mice with mobilized human blood CD34+ cells provides an in vivo model for human megakaryocytopoiesis and platelet production. Blood 2001;97(6):1635-1643. (29) Suzuki K, Hiramatsu H, Fukushima-Shintani M, Heike T, Nakahata T. Efficient assay for evaluating human thrombopoiesis using NOD/SCID mice transplanted with cord blood CD34+ cells. Eur J Haematol 2007;78(2):123-130. (30) Bruno S, Gunetti M, Gammaitoni L et al. Fast but durable megakaryocyte repopulation and platelet production in NOD/SCID mice transplanted with exvivo expanded human cord blood CD34+ cells. Stem Cells 2004;22(2):135-143. (31) Salles II, Thijs T, Brunaud C et al. Human platelets produced in nonobese diabetic/severe combined immunodeficient (NOD/SCID) mice upon transplantation of human cord blood CD34(+) cells are functionally active in an ex vivo flow model of thrombosis. Blood 2009;114(24):5044-5051. (32) Hu Z, Yang YG. Full reconstitution of human platelets in humanized mice after macrophage depletion. Blood 2012;120(8):1713-1716. (33) Makino S, Kunimoto K, Muraoka Y, Mizushima Y, Katagiri K, Tochino Y. Breeding of a non-obese, diabetic strain of mice. Jikken Dobutsu 1980;29(1):1-13. (34) Serreze DV, Chapman HD, Varnum DS, Gerling I, Leiter EH, Shultz LD. Initiation of autoimmune diabetes in NOD/Lt mice is MHC class I-dependent. J Immunol 1997;158(8):3978-3986. (35) Takenaka K, Prasolava TK, Wang JC et al. Polymorphism in Sirpa modulates engraftment of human hematopoietic stem cells. Nat Immunol 2007;8(12):13131323. (36) Bosma M, Schuler W, Bosma G. The scid mouse mutant. Curr Top Microbiol Immunol 1988;137:197-202. (37) Nonoyama S, Smith FO, Bernstein ID, Ochs HD. Strain-dependent leakiness of mice with severe combined immune deficiency. J Immunol 1993;150(9):38173824. 24 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. (38) Hioki K, Kuramochi T, Endoh S, Terada E, Ueyama Y, Ito M. Lack of B cell leakiness in BALB/cA-nu, scid double mutant mice. Exp Anim 2001;50(1):67-72. (39) Brimnes J, Reimann J, Claesson MH. Immunoglobulin leakiness in scid mice with CD4(+) T-cell-induced chronic colitis. Clin Immunol 2000;96(3):222-229. (40) Bosma MJ. B and T cell leakiness in the scid mouse mutant. Immunodefic Rev 1992;3(4):261-276. (41) Harrison P. Platelet function analysis. Blood Rev 2005;19(2):111-123. (42) Bath PM, Carney C, Markandu ND, MacGregor GA. Platelet volume is not increased in essential hypertension. J Hum Hypertens 1994;8(6):457-459. (43) Ricks DM, Kutner R, Zhang XY, Welsh DA, Reiser J. Optimized lentiviral transduction of mouse bone marrow-derived mesenchymal stem cells. Stem Cells Dev 2008;17(3):441-450. (44) Siapati EK, Bigger BW, Miskin J et al. Comparison of HIV- and EIAV-based vectors on their efficiency in transducing murine and human hematopoietic repopulating cells. Mol Ther 2005;12(3):537-546. (45) Davis BM, Humeau L, Slepushkin V et al. ABC transporter inhibitors that are substrates enhance lentiviral vector transduction into primitive hematopoietic progenitor cells. Blood 2004;104(2):364-373. (46) Shi Q, Fahs SA, Kuether EL, Cooley BC, Weiler H, Montgomery RR. Targeting FVIII expression to endothelial cells regenerates a releasable pool of FVIII and restores hemostasis in a mouse model of hemophilia A. Blood 2010;116(16):3049-3057. 25 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Table 1. Non-restrictive linear amplification-mediated PCR (nrLAM-PCR) determined insertion sites of 2bF8 transgene # Chr. 1 2 3 4 5 1 1 1 3 6 Dir. of T.g. ↑ ↑ ↑ ↑ ↑ 6 7 7 7 ↓ ↑ 8 9 7 8 ↓ ↑ 10 11 12 13 11 11 11 11 ↓ ↓ ↓ ↓ 14 12 ↓ 15 16 17 18 19 14 14 16 16 17 ↑ ↓ ↑ ↑ ↓ Gene ↓ DDI2 (DNA Damage Inducible Protein 2) Intron 1 ↑ ZSWIM5 (Zinc Finger, SWIM Domain Containing 5) Intron 1 ↓ AHS1L (Probable histone-lysine N-methyltransferase ASH1L) Intron 14 ↓ CLASP2 (Cytoplasmic Linker Associated Protein 2) Intron 12 Intergenic [Located downstream of Hypothetical Protein LOC100288169 and upstream of DUSP22 (Dual Specificity Phosphatase 22)] ↑ DGKB (Diacylglyceral kinase beta isoform 1, 2) Intron 2 ↑ GTF2IRD2 (General transcription factor II-I repeat domain-containing protein 2A) Intron 2 ↓ PILRB (Paired immunoglobulin-like type 2 receptor beta precursor) Intron 3 Intergenic [Located downstream of UNC5D (Netrin receptor UNC5D precursor) and upstream of KCNUI (potassium channel subfamily U member)] ↓ RBM14-RBM41 (RBM14-RBM4 protein isoform 1, 2) Intron 1 ↑ ARRB1 (beta-arrestin-1 isoform A and B) Intron 1 ↑ BSCL2 (Bernadinelli-Seip Congenital Lipidystrophy 2 [Seipin]) Intron 5 Intergenic [Located downstream of LTBP3 (latent transforming growth factor beta binding protein 3) and upstream of SSSCA1 (Sjogren syndrome / scleroderma autoantigen 1)] Intergenic [Located downstream of ATP2B1 (plasma membrane calcium-transporting ATPase 1isoform 1a, 1b) and upstream of C12orf12 (hypothetical protein LOC196477)] ↓ SOCS4 (Suppressor of Cytokine Signaling 4) Exon 4 in 3’UT ↑ GPR132 (G protein-coupled receptor 132) Intron 1 ↑ CREBBP (Creb Binding Protein) Exon 30 ↑ KIAA0430 (Limkain b-1 isoform 1, 2, 3) Intron 6 ↓ SUZ12 (Supressor of Zeste 12 Homolog [Drosophila]) Intron 15 “Chr.” means chromosome; Arrows indicate the directions of the transgene (Tg) or gene relative to the standard chromosomeal numbering. The insertion sites listed in this table were from 5 NSG recipients. 26 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Figure legends Figure 1. Human cell chimerism in NSG and NSGF8KO mice that received 2bF8 LV-transduced hCB CD34+ cells. Blood samples were collected from tail bleeds. Human platelets were identified by a mouse anti-human GPIbα antibody, AP1. Mouse platelets were stained with a rat anti-mouse CD41 (GPIIb) antibody. Human leukocytes were identified by mouse anti-human CD45 antibody and mouse leukocytes were identified by rat anti-mouse CD45 antibody. (A) Representative flow cytometric analysis of platelets in peripheral blood of NSG and NSGF8KO recipients at 3 weeks after transplantation. (B) Representative flow cytometric analysis of leukocytes in peripheral blood of NSG and NSGF8KO recipients between 5-7 weeks after transplantation. (C) Human platelet chimerism in NSG and NSGF8KO recipients. Samples from NSG mice and normal human individuals were used as controls. Data are summarized from 10 trials of xeno-transplants. D) Human leukocyte chimerism in NSG and NSGF8KO recipients. Samples from NSG mice and normal human individuals were used as controls. Data are summarized from 10 trials of xeno-transplants. (E) Time course of human platelet production in NSG recipients. Data are summarized from 2 trials of xeno-transplants. n = 4 to 9 mice for each group. (F) Time course of human leukocyte production in NSG recipients. Data are summarized from 2 trials of xeno-transplants. n= 4 to 7 mice for each group. Figure 2. Genetic analysis of 2bF8 LV-transduced hCB transplantation recipients. (A) PCR analysis of the 2bF8 transgene. Genomic DNA was purified from peripheral leukocytes. Leukocytes from NSG mice and a normal human individual were used as controls. 2bF8 transgenic mouse DNA was used as a positive control for the 2bF8 27 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. transgene. Shown is one representative experiment that was performed three times. Wildtype (WT) mouse FVIII and human GAPDH were used as internal controls. (B) Realtime quantitative PCR quantified the average copy number of the 2bF8 transgene per cell in 2bF8 LV-transduced hCB transplantation recipients. Peripheral leukocyte-derived genomic DNA was used to analyze 2bF8 proviral DNA with normalization to human GAPDH. Bars represent mean ± SD. For individual mice analyzed more than once over the course of study, the average copy number was calculated. Genomic DNA from NSG mice was used as a control. Data are summarized from 2 trials of xeno-transplants. Figure 3. Analysis of platelet FVIII expression in 2bF8 LV-transduced xenotransplanted recipients. Platelets were lysed in 0.5% CHAPS. The levels of plateletderived FVIII expression were determined by chromogenic FVIII activity (FVIII:C) assay of platelet lysates from recipients at least three weeks after transplantation. Bars represent mean ± SD. For individual mice analyzed more than once over the course of study, the average FVIII level was calculated. Platelets from NSG mice, C57BL6 mice, and normal human individuals were used as controls. (A) Quantitative evaluation of FVIII activity levels in platelet lysates from 2bF8 LV-transduced hCBT recipients (humanized) NSG or NSGF8KO recipients. (B) Quantitative evaluation of FVIII activity levels in hCB-derived human platelets based on human platelet numbers as determined by flow cytometry. Data are summarized from 10 trials of xeno-transplants. Figure 4. Electron microscopy determines cellular location of 2bF8 transgene protein. Isolated platelets from 2bF8LV-transduced xeno-transplanted NSG recipients (A-C), normal human individuals (D-F), and untransplanted NSG control mice (G-I) 28 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. were immunostained for hFVIII and either endogenous human or mouse VWF. hFVIII was stained with mouse anti-human FVIII monoclonal antibody 103.3 and probed with goat anti-mouse colloidal gold probe (6 nm, some indicated by arrow). VWF was stained with rabbit anti-human VWF polyclonal antibody (Dako), which cross reacts with mouse VWF, and probed with goat anti-rabbit colloidal gold probe (10 nm, some indicated by arrowhead). The results show hFVIII is stored together with human VWF in platelet αgranules of 2bF8LV-transduced human platelets (A-C). Scale bar is 100 nm. Figure 5. Phenotypic correction assessment. (A) Tail clip survival test assessing the phenotypic correction in 2bF8LV-transduced humanized NSGF8KO mice. The tail clipping test was performed at least 4 weeks after transplantation. Mice surviving beyond 24 hours were considered to have achieved phenotypic correction. Untransduced hCBtransplanted and untransplanted NSGF8KO recipients were tested as controls. Data are summarized from 10 trials of xeno-transplants. (B) ROTEM analysis. The ROTEM analysis was performed between 6 and 8 weeks after transplantation when human platelet chimerism was 0.4-5.5%. Representative TEMograms from untransplanted NSG, untransplanted NSGF8KO, and 2bF8LV-transduced humanized NSGF8KO (with 2% human platelet chimerism) mice are shown. (C) Whole blood clotting time determined by ROTEM analysis. Untransplanted NSG and NSGF8KO mice were used as controls. Data are summarized from 5 trials of xeno-transplants. 29 Figure 1 Platelets Human control NSG/2bF8LV NSG/2bF8LVhCBT recipient 0.00 0.00 98.11 0.05 56.49 0.78 0.95 99.05 1.28 0.56 15.67 27.07 NSGF8KO/2bF8LV- NSGF8KO/2bF8LVNSGF8KO/2bF8LV NSGF8KO/2bF8LV hCBT recipient hCBT recipient 58.9 0.49 18.4 0.44 10.8 29.8 6.48 74.4 Mouse GPIIb Leukocytes y Huma an CD45 NSG control Human control NSG/2bF8LVNSG/2bF8LV hCBT recipient 0.03 0.25 98.4 1.03 81.4 0.56 0.72 99.0 0.53 0.002 8.12 9.93 Mouse CD45 NSGF8KO/2bF8LV- NSGF8KO/2bF8LVNSGF8KO/2bF8LV NSGF8KO/2bF8LV hCBT recipient hCBT recipient Hum man CD45 B Mouse GPIIb 91.1 0.39 86.1 0.16 2.48 6.02 7.56 6.18 Mouse CD45 From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Human n GPIb NSG control Human GPIb A P < .01 60 P < .01 40 20 n=10 n=6 n=14 n=12 n=10 n=9 5w NSG Human 40 20 0 n=6 n=6 n=14 n=12 n=9 3w 5w NSG Human 3w 5w n=9 80 1400 1200 1000 60 800 40 600 400 20 200 0 0 0 1 2 3 4 5 7 10 12 Time after transplantation (weeks) ytes (x 103/μL) Human leukocy F Human platelets Murine platelets Mouse plateletts (x 103/μL) Human plateletts (x103/μL) E 100 60 NSG/ N hCBT 2bF8LV-h recipients 3w Co ontrols 5w NSG/ V-hCBT 2bF8LV reciipients 3w GF8KO/ NSG 2bF8LV V-hCBT reciipients 0 80 1.75 1.50 Human leukocytes Murine leukocytes 1.75 1.50 1.25 1.25 1 00 1.00 1 00 1.00 0.75 0.75 0.50 0.50 0.25 0.25 0.00 0.00 0 1 2 3 4 5 7 10 12 Time after transplantation (weeks) ytes (x103/μL) Mouse leukocy 80 P = .15 100 Controls Human leu ukocyte chimerrism (%) 100 P = .12 D From www.bloodjournal.org by guest on June 15, 2017. For personal use only. platelet chimeris sm (%) Human p C NSGF8 8KO/ hCBT 2bF8LV-h recipients Figure 1 2bF8tg+ contrrol dH2O 4 3 2 1 DNA Marker NSG/2bF8LVhCBT recipients 2bF8 (650 bp) WT mFVIII (233 bp) Average c copy number of 2 2bF8 transgene e per human cell hGAPDH (100 bp) B 4 3 2 1 0 n=7 n=3 NSG/2bF8LVNSG hCBT recipients control From www.bloodjournal.org by guest on June 15, 2017. For personal use only. NSG control A Human control Figure 2 Figure 3 n=4 n=4 C 57 B L6 H um an n=4 SG n = 14 N 0.00 n=6 2bF8LV-hCBT 2bF8LV hCBT Untransduced and untransplanted recipients controls 25 20 15 10 5 0 n=6 n = 14 NA n=4 NA n = 12 n=4 C 57 B L6 H um an 0.25 P = 0.99 N SG 0.50 30 N SG N SG F8 K O 0.75 Platelet FVIII:C elets (mU//108 human plate 1.00 1 00 N SG N SG F8 K O Platelet FVIII:C U/108 total platele ets) (mU 1.25 B P = .33 Untransduced t d d and d 2bF8LV-hCBT U untransplanted recipients controls From www.bloodjournal.org by guest on June 15, 2017. For personal use only. A uman platelets s Hu untransduced)) (u 2bF F8LV-transduc ced hu uman platelets s A B C D E F G H I 100 nm From www.bloodjournal.org by guest on June 15, 2017. For personal use only. ouse platelets s Mo (untransduced ) Figure 4 Figure 5 B C mm NSG 60 40 20 20 40 60 CT % Surrvival 80 60 40 10 20 N/A n=3 mm 60 40 20 20 40 60 20 30 40 50 min NSGF8KO control CT 10 NSGF8KO control n=4 Un ntransduced recipients 20 - 60 0.3 3-2 2.1 - 15 n=7 2bF8 LVtransduced recipients 0 Human platelet chimerism (%) (at tail clipping) n=7 mm 60 40 20 20 40 60 control 20 30 40 50 min NSGF8KO/2bF8LVhCBT recipient CT 10 20 30 40 50 min P < .001 3600 3000 2400 1800 1200 600 0 NSG control NSGF8KO control NSGF8KO/ 2bF8LV-hCBT recipients From www.bloodjournal.org by guest on June 15, 2017. For personal use only. 100 CT (time to 2 mm m in second) A From www.bloodjournal.org by guest on June 15, 2017. For personal use only. Prepublished online November 22, 2013; doi:10.1182/blood-2013-08-520478 Platelet gene therapy corrects the hemophilic phenotype in immunocompromized hemophilia A mice transplanted with genetically manipulated human cord blood stem cells Qizhen Shi, Erin L. Kuether, Yingyu Chen, Jocelyn A. Schroeder, Scot A. Fahs and Robert R. Montgomery Information about reproducing this article in parts or in its entirety may be found online at: http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requests Information about ordering reprints may be found online at: http://www.bloodjournal.org/site/misc/rights.xhtml#reprints Information about subscriptions and ASH membership may be found online at: http://www.bloodjournal.org/site/subscriptions/index.xhtml Advance online articles have been peer reviewed and accepted for publication but have not yet appeared in the paper journal (edited, typeset versions may be posted when available prior to final publication). Advance online articles are citable and establish publication priority; they are indexed by PubMed from initial publication. Citations to Advance online articles must include digital object identifier (DOIs) and date of initial publication. Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036. Copyright 2011 by The American Society of Hematology; all rights reserved.