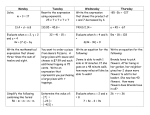
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Effect of leaf herbivory by spider mite Tetranychus urticae on flower characteristics of different cucurbits Marit A.M. Rutten (910125718060) Dr. ir. Iris F. Kappers, Laboratory of Plant Physiology, Wageningen March – December 2014 Master Biology Thesis (PPH-80430) Effect of leaf herbivory by spider mite Tetranychus urticae on flower characteristics of different cucurbits Marit A.M. Rutten Abstract Cucumber, melon, watermelon, courgette, pumpkin and bitter gourd are some economically very important (cultivated) fruits that belong to the Cucurbitaceae family, which is troubled by the spider mite Tetranychus urticae. This herbivore causes chlorotic lesions in the leaves, which can have a tremendous effect on the plant’s performance, for example by changes in flower morphology and metabolism. Most cucurbits depend on pollination for fruit and seed production. In this study the variation among different cucurbit species with respect to flower characteristics that may be influenced by herbivory on leaves by spider mites is explored. With LC-MS and GC-MS the production of endogenous metabolites and volatiles are investigated, respectively. For the non-bitter cucumber and the bitter Chinese cucumber the RNA of flowers and leaves will be isolated to test which terpene synthase family (TPS) genes (TPS1, TPS9 and TPS19) are expressed in control and infested samples, and therefore may be responsible for the volatile emission identified with GC-MS. This study demonstrates a large range of qualitative and quantitative variation in volatile emission induced by leaf herbivory of spider mites (Tetranychus urticae) in leaves of different cucurbit. But there was no significant effect on flower volatile emission. The TPS genes investigated in this study were presumably not (always) the ones that were responsible for the emission of certain volatiles. Some flowers were most probably not able to produce plant-defence related volatiles, because of the tremendous drop in JA-Ile/JA ratio in flowers when plants were infested with spider mites. 1 Introduction Some economically very important cultivated fruits, which are produced worldwide, belong to the Cucurbitaceae family. Examples of such fruits are cucumber, melon, watermelon, courgette, pumpkin and bitter gourd. In 2012 the world produced about 65,134,078 tonnes of cucumbers and gherkins of which 86.5% was produced in Asia, The Netherlands accounts for 0.6% of total cucumber production (FAO, 2014). The Cucurbitaceae family, like other plant families, is troubled by various herbivores eating plant parts, especially the leaves (Barber et al., 2011; Park and Lee, 2005). Fortunately, the plant evolved some ways to protect itself from such harm. However, as Herms and Mattson (1992) already stated, a plant must find a balance between investing in growth and development on the one hand and defence on the other hand, because both are essential for life. Searching for this balance often results in a dilemma for the plant: to compete with neighbouring plants it has to grow fast enough, but on the other hand it has to maintain its defence mechanisms to survive in an environment with herbivores and pathogens (Herms and Mattson, 1992). Spider mite (Tetranychus urticae) Plants are attacked by herbivores and pathogens that belong to various taxons (e.g. arthropods, insects and mammals). Arthropods and insects represent the largest group of phytophagous organisms, approximately 1-3 million species (Schoonhoven et al., 2005). In this research a member of this large herbivore-group is taken into account, the two-spotted spider mite, Tetranychus urticae Koch (Fig. 1). An important characteristic of this mite is a dark spot at both sides of the body. This spider mite is a generalist herbivore and a serious pest in many crops, for instance cucumber (Park and Lee, 2005), but also in other cucurbits. Spider mites belong to the group of sucking feeders. They feed on the mesophyll in leaves by piercing cells with their stylet. The cells are sucked out which results in chlorotic lesions (reductions in chlorophyll concentration) on the leaves of the plant. Because of this, plant development will be less successful and when the damage caused by the spider mites is severe enough, it can be devastating for the plant. Spider mites prefer to live in colonies on the downside of leaves where they spin silk webs, which protect both mites and eggs against predators. A female spider mite can lay hundreds of eggs during her life. The small, spherical eggs hatch after 2-3 days. The time needed to complete the whole life cycle (Fig. 1) depends on the temperature of the climate the spider mites live in. A natural enemy of this spider mite species is the predatory mite Phytoseiulus persimilis (Fig. 1), which can eliminate whole spider mite populations (Fasulo and Denmark, 2009). Figure 1 Left up: Spider mite Tetranychus urticae Right: The life cycle of spider mites Left down: Predatory mite Phytoseiulus persimilis 2 Plant defence mechanisms Plants have evolved a multitude of defence mechanisms to prevent serious damage caused by herbivores, which can be divided in the main groups direct and indirect defence. Plant traits that affect the performance (survival and growth) and behaviour of attacking herbivores themselves are direct defences, those that attract the natural enemies of herbivores are indirect defences. Both strategies can be either constitutive (always expressed/present) or inducible (activated by herbivory) (Arimura et al., 2005; Kessler and Baldwin, 2002). Constitutive direct defences can be physical and/or chemical. An example of a physical one are trichomes (plant hairs), which are grown from the epidermal cells on numerous plant species. Trichomes are a good defence mechanism against piercing-sucking arthropods, like spider mites. When hairs are dense, they form a physical barrier that makes it very difficult for this kind of herbivores to reach the plant’s epidermis. To make it even more difficult, some plant hairs (glandular trichomes) can produce chemicals which are sticky (Wagner, 1991; Werker, 2000). As a response to herbivore attack (induced response), plants can change their phenotype (phenotypic plasticity) by making changes in their physical characteristics, like increased trichome density, which is an induced direct defence mechanism (Traw and Dawson, 2002). However, this also has an influence on the natural enemies (e.g. predatory mites) of piercing-sucking arthropods (e.g. spider mites). Those predators often walk large distances over the leaf surface to find their prey/host and trichomes are tough obstacles (Krips et al., 1999). Plants can also change their phenotype by making changes in their chemical characteristics, for example the production of secondary metabolites (Kessler and Baldwin, 2002; Mumm and Dicke, 2010). Secondary metabolites Secondary metabolites are organic compounds with a low molecular mass which have no function in plant development, growth or reproduction but in plant defence (Kessler and Baldwin, 2002; Mumm and Dicke, 2010). Some secondary metabolites will directly decrease the number of herbivores on the plant. Therefore, they are called direct defences. Examples of such secondary metabolites are toxins and alkaloids which are toxic to herbivores, and phenolic acids, tannins and lignin which reduce the digestibility and/or palatability of the plant material. These compounds are called endogenous metabolites, because they stay in the plant tissue (Dicke, 2009; Kessler and Baldwin, 2002; Lambers et al., 2008). Not all secondary metabolites produced in direct defence mechanisms are endogenous, some are volatile (De Moraes et al., 2001). Other secondary metabolites will not have a direct negative effect on the number of herbivores, but attract the natural enemies of those herbivores. Therefore, they are called indirect defences where specific volatile organic compounds (VOCs) are involved (Kessler and Baldwin, 2002; Dicke and Baldwin, 2010). VOCs, signal transduction pathways and HIPVs During their life cycle, plants communicate with organisms in their surroundings through volatile organic compounds (VOCs). These compounds are produced in every part of the plant, from roots underneath the soil till flowers at its top. VOCs determine the smell of the plant and can diffuse easily through the air or the soil. Therefore, they can be detected by organisms from a distance. Which kinds of organisms are attracted by a volatile depends on the plant part that emits the volatile. The volatile-profile of a leaf blend differs from one of a flower blend. This is because of the different functions of the plant parts. The flowers of most plants are meant to attract pollinators with their visual, tactile and olfactory signals. The scent of a flower can be a complex blend containing many compounds from different chemical 3 classes. The seven main classes are: aliphatics, benzenoids/phenyl propanoids, C5-branched compounds, terpenoids, nitrogen-containing compounds, sulphur-containing compounds and a class of various cyclic compounds. However, little is known about which individual compound attracts a specific pollinator (Knudsen et al., 1993; Knudsen et al., 2006; LucasBarbosa et al., 2011; Pichersky and Gershenzon, 2002). Herbivores can detect specific volatiles emitted by leaves and because of that locate their (host) plant. When herbivorous arthropods eat plants’ tissue, their oral secretions (regurgitant) contacts the plant. This regurgitant consists of elicitors that are species specific components that can be recognised by the plant’s membrane receptors (e.g. Hilker and Meiners, 2010). As a result three endogenous interconnecting signal transduction pathways in the plant are activated: the octadecanoid pathway with jasmonic acid (JA) as a key compound, the shikimic acid pathway with salicylic acid (SA) as a key compound, and the ethylene pathway. Jasmonic acid (JA), salicylic acid (SA) and ethylene (ET) are phytochemicals which have protective characteristics: they activate plant defence genes (e.g. Pieterse and Dicke, 2007). These plant defence genes are activated by transcription factors. In the JA signalling pathway, members of the jasmonate-ZIM domain (JAZ) protein family repress these transcription factors. It is known that jasmonic acid conjugated with the amino acid isoleucine (jasmonoyl-isoleucine or JA-Ile) is able to degrade the JAZ proteins, which results in free transcription factors and eventually the expression of plant defence genes (Thines et al., 2007). Some of those genes will induce the production of VOCs. As a consequence, the VOC composition of herbivore-damaged plants differs from the VOC composition of intact plants. This change in volatile mixture can be either quantitative (different emission level of components also present in non-infested plants) and/or qualitative (release of other components). Herbivore-damaged plants emit VOCs that can attract the natural enemies (arthropod predators and parasitoids) of the herbivorous arthropods. Those VOCs are also called HIPVs (herbivore-induced plant volatiles) and they are used by natural enemies to locate their prey (Dicke and Baldwin, 2010; Mumm and Dicke, 2010; Dicke et al., 2003). HIPVs are emitted locally at the site of damage (often a leaf), but can also be emitted systemically from undamaged parts of attacked plants, so there is a connection between plant parts. The aim of this connection is to prevent the attacker from spreading through the whole plant (Heil and Ton, 2008). Carnivores foraging for prey or hosts perceive HIPVs with their peripheral olfactory system that can be located for instance on antennae or palpi. The peripheral olfactory system consists of olfactory sensilla containing receptor neurons that are sensitive to specific HIPVs within a HIPV blend (Schoonhoven et al., 2005). Those specific HIPVs provide carnivores, for example, with information about the plant species or cultivar that emits the odour (Takabayashi and Dicke, 1996) and which herbivore species is damaging the plant (e.g. De Moraes et al., 1998). HIPV blends are often complex mixtures with multiple different compounds that are also from the same seven chemical classes as compounds in the scent of a flower. However, most compounds belong to one of the four main groups: fatty acid derivatives, phenylpropanoids, benzenoids and terpenoids (Dudareva et al., 2004; Mumm and Dicke, 2010). Terpenoids Terpenoids, also called isoprenoids or terpenes, are a large group of various compounds that are produced by many plant species. A relatively small number of terpenoids are involved in primary metabolism. Most terpenoids are secondary metabolites, which have a function in communication and defence (Aharoni et al., 2005; Trapp and Croteau, 2001). The terpene synthase family (TPS) is a set of related genes, approximately 20 to 150 (Chen et al., 2011), that encodes for enzymes which synthesize different terpenes (Bohlmann et al., 1998). All 4 terpenoids, primary metabolites as well as secondary metabolites, are derived from isopentenyl diphosphate. Via different pathways (mevalonate (MVA) pathway or methylerythritol 4-phosphate (MEP) pathway) consisting of terpene synthases monoterpenes, sesquiterpenes, diterpenes, triterpenes and tetraterpenes are produced. Monoterpenes and sesquiterpenes are volatile under ambient temperature and atmospheric pressure (Aharoni et al., 2005; Trapp and Croteau, 2001). Many plant species infested with the spider mite Tetranychus urticae emit volatile blends containing terpenoids (see Van den Boom et al., 2002). It is known that some terpenes are attractive to predatory mites. For example (E)-β-ocimene, linalool, (E)-4,8-dimethyl1,3,7-nonatriene [(E)-DMNT] and (E,E)-4,8,12-trimethyl-1,3,7,11-tridecatetraene [(E,E)TMTT] (Dicke et al., 1990; De Boer et al., 2004). Some examples of plants emitting terpenoids as a response to spider mite attack are lima bean (Phaseolus lunatus L.) and the cultivated cucumber (Cucumis sativus L.) (Bouwmeester et al., 1999; Dicke et al., 1990; Kappers et al., 2010; Mercke et al., 2004; Takabayashi et al., 1994). It is likely that other cucurbit-species will also produce terpenoids as a response to herbivory by the spider mite Tetranychus urticae. Cucurbits For the cultivated cucumber already a lot is known about its interaction with spider mites. However, the non-bitter cucumber (C. sativus var corona) does not need to be pollinated to produce fruits (seedless fruits) while other cucurbits depend on pollination (often by insects) for fruit and seed production. Previous research has shown that leaf herbivory does lead to changes in flower morphology and metabolism (e.g. Kessler and Halitschke, 2009; Theis et al., 2009; Thomson et al., 2004). As already mentioned, HIPVs are produced in leaves as a response to herbivory and they can be emitted systemically because plant parts are connected. As a consequence, HIPVs may also be expressed in the flowers, which will result in a different flower volatile blend. This may have an influence on the behaviour of flower visitors like pollinators (e.g. Adler et al., 2001), which may result in less or no pollinated flowers and thus less or no fruit production. For growers this is devastating. Studying volatile emission of vegetative and flower tissues of a plant may give a more complete picture of what the effect of plant defence against herbivores is and eventually could be on pollination (Lucas-Barbosa et al., 2011). The cucurbits cucumber (Cucumis sativus var corona, Cucumis sativus var Chinese long, 9930 and Cucumis sativus var hardwickii), melon (Cucumis melo), watermelon (Citrullus lanatus), courgette (Cucurbita pepo), pumpkin (Cucurbita maxima) and bitter gourd (Momordica charantia) will be studied to look at variation in interaction with spider mites within the Cucurbitaceae family (Fig. 2). Van Poecke and colleagues (2001) had shown that the model plant Arabidopsis thaliana is capable of emitting volatiles, as a response to herbivory, to attract carnivorous arthropods. Examples of emitted volatiles by A. thaliana are methyl salicylate and terpenoids (van Poecke et al., 2001). Both are known to be induced by herbivory in many plant species (Dicke et al., 1990). The entire genome of Arabidopsis thaliana is known. It also contains known terpenoid synthase genes (Aubourg et al., 2002; Chen et al., 2004; Tholl et al., 2005). Therefore Arabidopsis will also been grown and infested with spider mites. So there is strong evidence that flower morphology and metabolism change when there are herbivores attacking the plant. This leads to the following research question: Does herbivory by spider mites (Tetranychus urticae) on the leaves of different cucurbits have influence on flower/leaf characteristics? 5 First of all it must be confirmed that the spider mites really cause damage to the plant they are feeding on. Damage can consist of the chlorotic lesions the spider mites create on the leaves. Therefore, changes in the amount of chlorophyll will be measured on leaves of all cucurbit species. The objective of this study is to explore the variation among members of the plant family Cucurbitaceae with respect to flower characteristics that may be influenced by herbivory on leaves by spider mites (Tetranychus urticae). The characteristics that will be investigated are the production of endogenous metabolites and volatiles (especially terpenes) in flowers as well as leaves of the different cucurbits with and without leaf herbivory. With the methods liquid chromatography–mass spectrometry (LC-MS), and headspace collection in combination with gas chromatography–mass spectrometry (GC-MS) it is determined which kind of endogenous metabolites and volatiles are produced, respectively. For cucumber (Cucumis sativus var corona and Cucumis sativus var Chinese long, 9930) the RNA of flowers and leaves will be isolated to test which terpene synthase family (TPS) genes (TPS1, TPS9 and TPS19) are expressed in control and infested samples, and therefore may be responsible for the volatile emission identified with GC-MS. The genome of cucumber (Cucumis sativus var. sativus L.) is recently discovered (Huang et al., 2009) as well as a substantial part of the genome of melon (Cucumis melo L.) (GarciaMas et al., 2012). Some terpene synthase genes are already characterised in cucumber (Mercke et al., 2004), for other terpenoids it is currently investigated by which terpene synthase genes they are produced. Until now 26 TPS are known for cucumber (personal comment dr. ir. Iris F. Kappers). The known genomes and terpene synthase genes in Arabidopsis and cucumber can be helpful in studying the signal transduction pathways that induce direct and indirect defence mechanisms in the other plant species. It can be expected that there will be an overlap between cucumber and the other Cucurbitaceae members, because they are genetically closely related. Members of the family Cucurbitaceae are able to produce cucurbitacins. Cucurbitacins are bitter triterpenoid compounds that are responsible for the bitter taste of the foliage, the fruits are normally not bitter. These triterpenoid compounds are toxic to most organisms (can also be toxic to mammals, including man) and therefore are part of the defence of cucurbits against herbivores (Miró, 1995). Cucurbitacins are present constitutively in the plant, but can also be induced by herbivores (Agrawal et al., 1999). Plants that taste bitter contain the gene Bi, non-bitter plants contain the recessive allele bi (bitterfree) (Pierce and Wehner, 1990). The fruits of cucumber plants can become bitter as a result of stress. The bi gene in non-bitter plants prevents this from happening (Balkema-Boomstra et al., 2003). Cucurbitacins occur widely in wild and cultivated Cucurbitaceae (Miró, 1995). It is speculated that cucurbitacins are involved in the resistance towards the cucurbit-pest: the two-spotted spider mite, Tetranychus urticae Koch. Much research about this resistance is done in cucumber (Cucumis sativus). The research of Da Costa and Jones (1971) demonstrated that non-bitter cucumber plants suffered much more from feeding by the two-spotted spider mite than did bitter plants. The spider mite-mortality on bitter cucumber varieties was higher because mites did not evolve mechanisms to detoxify the bitter cucurbitacins in the leaves (Da Costa and Jones, 1971; Pierce and Wehner, 1990). Gould (1978) also found results supporting the hypothesis that cucurbitacins in the family Cucurbitaceae are involved in resistance against the twospotted spider mite. Even did Balkema-Boomstra and colleagues (2003) with cucurbitacin C. In the cultivated non-bitter cucumber Cucumis sativus (cultivar Eversweet) so far, only one cucurbitacin-form is identified. This cucurbitacin-form is similar to cucurbitacin C (Rice et al., 1981). However, spider mite resistance in cultivated non-bitter cucumbers (and other cucurbit species) is not (yet) reality (Balkema-Boomstra et al., 2003). 6 Figure 2 Phylogenetic tree of the family Cucurbitaceae In this research the tribes Momordiceae, Cucurbiteae and Benincaseae are important. To the Momordiceae-tribe belongs the plant Momordica charantia L. (bitter gourd). Cucurbita pepo L. (courgette/zucchini) and Cucurbita maxima Duchene (pumpkin) are in the Cucurbiteae-tribe. To the Benincaceae-tribe belongs Citrullus lanatus Thunb. (watermelon) and the genus Cucumis with the species Cucumis melo L. (melon) and Cucumis sativus L. with the varieties hardwickii, corona and Chinese long, 9930. 7 Material and methods Plant and mite material Seeds of six different cucurbit species were sown and grown in 1 L pots with potting compost at 22°C during 12 hours of light, 18°C at night and 70% relative humidity in a climate chamber at Wageningen University, The Netherlands (Table 1). TL lamps were used which relatively burned with 55% at an intensity of 175 µmoll. The seeds of Momordica charantia L. were sown and grown in 1 L pots with potting compost in a box made of glass in a greenhouse to create a humid, warm environment which they prefer (Njoroge and Van Luijk, 2004). Here they grew at 18-24°C, 20-85% relative humidity under a natural daylight photoperiod (March-April) for 2.5 weeks before being transferred to the same climate chamber as the other cucurbits. The dormancy in the seeds of Cucumis sativus L. var. hardwickii had to be broken by putting them in an Eppendorf tube with sterile water and warm them till 55°C in a warm water bath for 2 hours. They were germinated on water agar (2%) with 30 mg/L MS (Murashige & Skoog medium) in an oven of 28°C. After germination the hardwickii seedlings were grown further in a climate chamber with a temperature of 24°C till they had grown about 4 leaves. Then they were transferred to the same climate chamber as the other cucurbits. Before the Arabidopsis thaliana L. seeds were able to germinate on Rockwool, they needed a pre-treatment. The seeds were sterilized by washing them with 70% EtOH, 10% bleach and three times with sterile water to wash off the remaining EtOH and bleach. After sterilization, the seeds were germinated on Rockwool blocks in the same climate chamber as the other plant species. The phytophagous two-spotted spider mite Tetranychus urticae Koch was reared on lima bean plants (Phaseolus vulgaris L.) in a greenhouse at Wageningen University, The Netherlands. Table 1 Cucurbitaceae members (cucurbits) used in this research Bitter/non-bitter leaves Flowers ♀/♂/ Scientific name Trivial name Cultivated/wild Cucumis sativus L. var. corona cucumber cultivated (hybrid) non-bitter ♀ Cucumis sativus L. var. Chinese long, 9930 cucumber cultivated bitter (cucurbitacin C) ♀+♂ Cucumis sativus L. var. hardwickii cucumber wild bitter (cucurbitacin C) ♀+♂ Cucumis melo L. melon cultivated Citrullus lanatus Thunb. watermelon cultivated ♀+♂ Cucurbita pepo L. courgette cultivated ♀+♂ Cucurbita maxima Duchesne pumpkin cultivated ♀+♂ Momordica charantia L. bitter gourd bitter ♀+♂ (pentanorcucurbitacin A & B, momordicine II & IV) 8 Plant treatments The cucurbit plants used for experiments were 3-5 weeks old and had four or five fully expanded leaves (C. sativus var hardwickii did not germinate). Arabidopsis thaliana L. plants used for experiments had a rosette that was not fully grown and did not form a peduncle yet. Of each species (about 10 plants per species) the plants were split into two separate, but comparable compartments (Table 2). One half stayed in the climate chamber as control plants and the other half was put in tents in a greenhouse at Wageningen University, The Netherlands (Fig. 3). These tents were suitable for infestations with spider mites, which was done by laying a piece of lima bean leaf, infested with spider mites, on top of each plant. The Figure 3 Tent containing plants mites occupied the leaves themselves. The spider mites were on infested with spider mites the plants for approximately 10 days before further experiments were carried out. Table 2 Number of plants per treatment for all plant species Plant species C. sativus var corona C. sativus var Chinese long, 9930 C. melo (melon) C. lanatus (watermelon) C. pepo (courgette) C. maxima (pumpkin) M. charantia (bitter gourd) A. thaliana Plant number per treatment (biological replicates) Control Infested 5 5 5 5 5 5 5 4 5 5 5 5 4 3 3 3 Harvest leaf and flower material About 15 days after the start of the plant treatment, leaf number 4 or 5 (or some leaves of the rosette of A. thaliana) of each plant was harvested and ground with a mortar and pestle in liquid nitrogen. Material of each leaf was divided over Eppendorf tubes, around 100 mg leaf material per tube, for different experiments: chlorophyll measurement, RNA isolation and liquid chromatography–mass spectrometry (LC-MS). Another Eppendorf tube was filled with leftover material as back-up. Of each plant species flowers were harvested and ground with a mortar and pestle in liquid nitrogen. If applicable, male and female flowers were harvested separately. The time points at which flowers were harvested varied between plant species because of the difference in plant development rate. Material of each flower was divided over Eppendorf tubes, around 100 mg flower material per tube, for different experiments: RNA isolation and liquid chromatography–mass spectrometry (LC-MS). Another Eppendorf tube was filled with leftover material as back-up. 9 Chlorophyll analysis The amount of chlorophyll in leaves of control and infested plants was measured as an indication of leaf damage caused by spider mites. Spider mites suck out the cells of a leaf, which results in chlorotic lesions (Fasulo and Denmark, 2009). The amount of chlorophyll in the samples was determined with a spectrophotometer, which measured the absorbance of chlorophyll at various wavelengths. For each plant species there were about 5 control plants and 5 infested plants, which function as biological replicates, from which leaves were investigated (see Table 2). Per sample around 100 mg ground leaf material was put in an Eppendorf tube. 1 mL of 96% EtOH (around 4°C) was added as a solvent to each sample, which was homogenized on a vortex till the leaf material thawed. All samples were stored in the dark in a fridge (around 4°C) overnight. After homogenizing on a vortex again, the samples were centrifuged at low speed. For each sample a part of the supernatant was diluted 10 times with 96% EtOH, the other part stayed undiluted. Of both the diluted and undiluted extractions 200 µL was added in wells of a 96-well plate. As a control, some wells were filled with 200 µL 96% EtOH. This 96-well plate was read by a spectrophotometer at wavelengths 665 nm and 649 nm which gave information about the light absorbance by chlorophyll A and B and the controls. With the formulas provided by Rowan (1989), 𝐶ℎ𝑙 𝐴 (𝜇𝑔 𝑚𝐿−1 ) ≈ 13.70 × 𝐴665 − 5.76 × 𝐴649 𝐶ℎ𝑙 𝐵 (𝜇𝑔 𝑚𝐿−1 ) ≈ −7.60 × 𝐴665 + 25.8 × 𝐴649 the concentration of chlorophyll A and B in each sample was calculated (Raymond, 2006). Those concentrations were corrected for the solvent by subtracting the concentration of 96% EtOH. To get to know the amount of chlorophyll per fresh leaf weight (µg/mg FW), the concentrations for chlorophyll A and B were divided by the weight of the ground leaf material. For each plant species the average amount of chlorophyll per fresh leaf weight from the biological replicates was calculated. These average amounts of chlorophyll in the control and infested leaves were compared with each other by setting the amount of the controls at 100%. Therefore, the chlorophyll percentage in the infested leaves was calculated which gave an indication for the chlorophyll reduction caused by spider mites sucking out the contents of the cells in leaves. Dynamic headspace collection Dynamic headspace (airspace) collection was applied in the climate chamber as well as in the greenhouse to analyse the headspace profile and its quantity of herbivory-induced volatiles in the different cucurbits. In this research, the headspace collection was done in different ways depending on the size of the plant parts from which the scents were collected. Ideally, the investigated leaves and flowers stayed attached to the plant, because mechanical damage by cutting the leaf or flower from the plant could influence the volatile emission of the plant part. This was done by developing a new headspace set-up using a PET (polyethylene terephthalate) bag (Toppits® Melitta) instead of a glass jar (Fig. 5). This bag was placed around the leaf or flower on a living cucurbit plant and was loosely attached to the stem of the leaf or flower to close it a bit. There were two holes in the bag that functioned as an inlet and an outlet, the same as in a glass jar. In both the inlet and outlet a stainless steel cartridge was connected, which contained 200 mg of the adsorbent Tenax TA (20/35 mesh; GraceAlltech, Deerfield, MI, USA). Using a pump air was blown into the bag with a constant flow of 150±5 mL min-1 through the inlet cartridge which filtered the air. On the outlet cartridge a portable battery-operated air sampler was placed that drew the air through the cartridge for 1 hour with a constant flow of 100±5 mL min-1 (Ametek/du Pont de Nemours & Co., type Alpha-2, 10 DEHA International, Huizen, The Netherlands). Here the volatiles emitted by the plant part were trapped. The difference in air flow strength was necessary to prevent dirty air leaking out from the bag from the hole around the stem of the leaf or flower where the bag did not close well enough. However, not all plant parts of all investigated plant species were big enough to cover with the PET bag. Therefore, the plant part needed to be harvested and the new headspace collection set-up recently developed by The Laboratory of Plant Physiology at Wageningen University & Research Centre was used (Fig. 5). This set-up does not make use of the portable battery-operated air samplers anymore and it has 12 positions to collect the headspace at the same time. In principle it works the same as with the PET bag. A plant or plant part was covered with a glass jar (2.5 L). The inner space was connected with an inlet and outlet with cartridges which contained 200 mg Tenax TA. A pump blew air into the jar with a constant flow of 150±5 mL min-1 through the inlet cartridge which filtered the air. Another pump was connected to the outlet cartridge that drew air from the inside of the jar through the outlet cartridge with a constant flow of 100±5 mL min-1 for 1 hour. Here the volatiles emitted by the plant part were trapped. Also in this set-up the difference in flow strength was necessary to prevent dirty air leaking from the outside of the jar into the system. A computer was connected to both pumps to control the air flow and other environmental conditions. Figure 5 Headspace collection set-up Left: Headspace collection set-up with a PET bag. The bag was placed around the leaf or flower. With a pump air was blown into the bag through the inlet Tenax cartridge which filtered the air. The portable batteryoperated air sampler drew the air through the outlet Tenax cartridge to trap the emitted volatiles. A higher inlet airflow than outlet airflow was necessary to prevent dirty air from leaking into the PET bag. Right: New headspace collection set-up developed by The Laboratory of Plant Physiology at Wageningen University & Research Centre. The glass jar was placed on the flower. With a pump air was blown into the jar through the inlet Tenax cartridge which filtered the air. Another pump drew the air through the outlet Tenax cartridge to trap the emitted volatiles. A higher inlet airflow than outlet airflow was necessary to prevent dirty air from leaking into the glass jar. 11 When the new headspace set-up was not available, the headspace was collected in the old fashioned way by putting the plant part of interest in a glass jar (0.5 L). The jar was closed with a lid equipped with an inlet and an outlet. In both the inlet and outlet a stainless steel cartridge was put, which contained 200 mg Tenax TA. Air was drawn through the cartridges for 1 hour using a portable battery-operated air sampler with a constant flow of 100±5 mL min-1. Gas chromatography–mass spectrometry Headspace samples of cucurbit leaves and flowers were analysed with a gas chromatography– mass spectrometry (GC-MS) machine to identify the volatiles that were trapped (analyte). This machine consisted of Thermo Trace GC Ultra (Thermo Fisher Scientific, Waltham, MA, USA) connected to a Thermo Trace DSQ (Thermo Fisher Scientific, Waltham, MA, USA) quadrupole mass spectrometer. Before being analysed, the Tenax cartridges were dry-purged with nitrogen (pressure 20 PSI) for 10 minutes at room temperature to remove any water. The cartridges were heated at 250°C by a thermal desorption system with a helium flow of 30 mL min-1 to elute the volatiles from the adsorbent Tenax. To concentrate the analytes, they were cooled down by an electronically-cooled sorbent trap at 3°C and brought splitless onto the analytical column by rapidly increasing the temperature in the cold trap to 250°C. This column started with a temperature of 40°C and increased with 10°C min-1 until 280°C (Kappers et al., 2011). Gas chromatography is based on a mobile phase which carries the analyte with it through a column containing stationary phase materials as a film at its wall. The analyte consists of molecules with specific chemical and physical characteristics which determine the rate at which they are transported through the column by the mobile phase, and their interaction (adsorption) by the stationary phase materials in the column. So, the column retains the molecules, elutes them eventually when the temperature of the column got high enough and therefore the different molecules reach the end of the column at specific time points (retention time). In this way the compounds are separated from each other and are further analysed by the mass spectrometer (Tholl et al., 2006). In the mass spectrometer the compounds that exit the column were ionized by electron impact (EI) at 70 eV, which resulted in charged molecule fragments. They were separated by the quadrupole according to their mass-to-charge ratio of 45-400 m/z. With its specific mass spectrum on a certain retention time it was clear how abundant the molecule (fragment) was present in the headspace collection sample. To align the peaks in the chromatograms of all headspace collection samples, Metalign software (PRI-Rikilt, Wageningen, The Netherlands) was used which resulted in two excel files: a noise and an amplitude file. The noise was subtracted from the amplitude with the IF-formula: IF (amplitude value > 5000, amplitude value, 10 (a random number)). Eventually the peaks with mass 93 (important terpene-mass) were selected for, the average and standard deviation was calculated and a Students t-test was used to see at which retention times the control and infested samples were significantly different (p<0.05). For those significantly different samples the fold change was calculated. If the amplitude of infested plant material was higher than that of control plant material, than there was an increase in volatile emission level by the infested plant (shown in tables 4-7 as upward arrows). When not, there was a decrease in volatile emission level by the infested plant (shown in tables 4-7 as downward arrows). A fold change higher than 10 did indicate an important peak in the volatile blend. Peaks were visualized in the chromatograms (shown in figures 8-12 and in Appendix 3), which were also consulted for the determination of volatile emission level by comparing the relative abundances of the control and infested peak with each other. For peaks with a fold change higher than 10, the putative identity of the volatiles was searched for by using Xcalibur and by comparing their retention times and mass spectra with those in NIST and Wiley spectral libraries (Kappers et al., 2011). Some other peaks 12 without a fold change higher than 10 could also be interesting to know the volatile identity of. For example when at a specific retention time the peaks of both treatments differ (a lot) in their relative abundance. Liquid chromatography–mass spectrometry Leaf and flower material of C. sativus var Chinese long, 9930 and C. sativus var corona was examined for endogenous stress-related phytohormones by a liquid chromatography–tandem mass spectrometry (LC-MS/MS). The metabolites of interest were jasmonic acid (JA), its bioactive amino acid conjugate (JA-Ile), salicylic acid (SA) and abscisic acid (ABA). The leaf or flower sample was homogenized in a mortar using liquid nitrogen and divided over three 2ml microcentrifuge tubes (technical replicates). About 15 mg of fresh plant weight was extracted with 1 mL 10% ice cold methanol/water. To control the extraction recovery and validate the quantification of the selected metabolites, stable isotope-labelled compounds were used as internal standards: [2H6]-JA and [2H2]-JA-Ile (Olchemim), [2H4]-SA and [2H6]ABA (Cambridge Isotope Laboratories). Tungsten carbide balls were added to each sample. The samples were homogenized in a mixer mill MM400 at a frequency of 27 Hz for 3 minutes and further sonicated in an ultrasonic bath for 2 minutes. Samples were extracted 25 min/4°C using a rotator shaker. To separate the plant material from the extraction solvent, the tubes were centrifuged for 4 minutes at 14,000xg at 4°C. The supernatant was transferred to a new tube. The plant material was re-extracted by adding 1 mL 10% ice cold methanol/water, quickly spin stirring by vortex and centrifuging it again. Both supernatants were combined and further purified by solid phase extraction. The Oasis® HLB (1 cc/30 mg cartridge, Waters Co., Milford, MA, USA) sorbent was conditioned with 1 mL of 100% methanol (organic solvent) and equilibrated with 1 mL of MiliQ water. These liquids were used to activate functional groups of the sorbent. The supernatant was loaded onto the column and the sorbent was subsequently washed with 1 ml of extraction solvent (10% methanol/water). The compounds were eluted off the cartridge by adding 3 mL 80% methanol. Samples were evaporated under the stream of nitrogen till dryness and reconstructed with 40 µL of 15% ACN: 15 mM HCOOH, v/v (initial conditions of LC gradient) (Flokova et al., 2014). Samples were further analysed by the ultra-high performance liquid chromatography– tandem mass spectrometry, consisting of the ACQUITY UPLC System (Waters) and Xevo TQ S (Waters) triple quadrupole mass spectrometer. The solvent used as a mobile phase was composed of 15mM HCOOH (A, aqueous part) and ACN (B, organic part). The compounds were separated by ACQUITY UPLC CSH C18 column (100x2.1 mm, 1.7 μm; Waters) at a flow rate 0.4 ml/min and temperature 40°C with binary gradient as follows: isocratic elution at 15% B (from 0 to 1 min), linear increase to 60% B (1-7 min), linear increase to 80% B (7-9 min), logarithmic increase to 100% B (9-10 min). Finally the column was washed with 100% B for 1 min and equilibrated for initial conditions for 2 min. The effluent was introduced into the electrospray ion source of the mass spectrometer. The capillary voltage was set on 3kV. The source block/desolvation temperature was 120°C/ 550°C with desolvation gas flow 650 l.h-1. Compounds were quantified by multiple ion monitoring mode (MRM) with settings listed in Flokova et al., 2014. Gene expression of TPS1, TPS9 and TPS19 genes The RNA in leaf as well as flower material of the plant species C. sativus var corona and C. sativus var Chinese long, 9930 was isolated to test which terpene synthase family (TPS) genes (TPS1, TPS9 and TPS19) were expressed in control and infested samples. Per sample around 100 mg ground leaf or flower material was put in an Eppendorf tube to which 1 mL TriPure was added. This mixture was homogenized on a vortex and was allowed to stand at room temperature for 5 minutes before 200 µL of chloroform was added. This mixture was again 13 homogenized on a vortex and allowed to stand at room temperature for 5 minutes. TriPure and chloroform form a phenol-chloroform extraction which is a liquid-liquid extraction technique for purifying RNA and eliminating plant proteins. After centrifugation at 12,000xg at 4°C for 15 minutes the supernatant containing the RNA was pipetted into a new Eppendorf tube, while the pellet containing proteins and other plant cell debris was thrown away. To the new Eppendorf tube 0.5 times the supernatant volume of 96% ethanol (EtOH) was added and mixed by inverting. This mixture was applied to an RNeasy spin column placed in a 2 mL collection tube and centrifuged at 8000xg for 15 seconds. The flow-through was discarded. From this point onwards the protocol “Purification of Total RNA from Plant Cells and Tissues and Filamentous Fungi” was used starting with step 7 on page 53 (RNeasy Mini Handbook 06/2012, QIAGEN). During this protocol the RNA on the RNeasy spin column was washed with RW1 and RPE buffer. Every time a specific buffer was applied to the column, a waiting step of 5 minutes was introduced to give the buffer time to work, instead of centrifuging immediately. At step 11 of the protocol 100-200 µL RNase-free water was used to elute the RNA from the RNeasy spin column, it depends on if a back-up was preferred. With NanoDrop (Nanodrop 1000 3.7.1) the RNA concentration and the A260/A280 and A260/A230 values were measured. The A260/A280 and A260/A230 values should be around 2.0. When these values were sufficient the RNA was used for the DNase treatment. 89 µL RNA was applied to an Eppendorf tube together with 10 µL DNase buffer and 1 µL DNase. This was allowed to stand at room temperature for 15 minutes. From this point onwards the protocol “RNA Cleanup” was used starting with step 1, second line on page 54 (RNeasy Mini Handbook 06/2012, QIAGEN). During this protocol the RNA was washed with RLT and RPE buffer and with 96% ethanol (EtOH). Also within these washing steps, a waiting step of 5 minutes was introduced to give the buffer or the ethanol time to work, instead of centrifuging immediately. At step 7 of the protocol 30-100 µL RNase-free water was used to elute the RNA from the RNeasy spin column. Again with NanoDrop the RNA concentration and the A260/A280 and A260/A230 values were measured. The A260/A280 and A260/A230 values should again be around 2.0 (see Appendix 1). With reverse transcription polymerase chain reaction (RT-PCR) complementary DNA (cDNA) transcripts from RNA were created, which were eventually used to detect gene expression. This cDNA was made by using 1 µg of RNA. A total volume of 20 µL, of which 4 µL 5x iScript reaction mix and 1 µL iScript Reverse Transcriptase, was added to Eppendorf PCR tubes, supplemented with Nuclease free water. The PCR machine (GeneAmp PCR System 9700, PE Applied Biosystems) followed the reverse transcription programme: 5 minutes at 25°C, 30 minutes at 42°C, 5 minutes at 82°C and hold at 10°C (see Appendix 1). With NanoDrop the concentration of created cDNA was measured, which should be around 1000 ng/µL and the difference in concentration among the cucurbit samples should be less than 100 ng/µL (see Appendix 2, step 7). The cDNA was diluted to a volume of 200 µL. The samples were run on the qPCR machine (Bio-RAD MyiQTM Single colour Real-Time PCR Detection System) in duplicate (see Appendix 2). Forward and backward primers of TPS1, TPS19 and TPS9 and actin were used in combination with the cDNA of the flowers of C. sativus var Chinese long, 9930 and C. sativus var corona. TPS1, TPS19 and TPS9 encode for enzymes which synthesize the terpenes linalool, α-farnesene and β-ocimene, respectively (personal comment dr. ir. Iris F. Kappers). Actin was used as a reference gene, because it is a housekeeping gene that is expressed in each plant cell under normal conditions, and its expression is quite stable under different conditions. The threshold cycles of the genes of interest (TPS1, TPS19 and TPS9) were compared to the threshold cycle of the housekeeping gene actin to see how relative transcripts of each of the TPS genes were. 14 Results Chlorophyll analysis Figure 6 Left: control plants 15 days after germination. Right: plants infested with spider mites for 20 days. On both pictures C. pepo (courgette) plants are shown. Figure 6 showed what the treated plants of C. pepo (courgette) looked like after a certain period. The control plants of this species were greener than the plants infested with spider mites. In Table 12 (see Appendix 4) the amount of chlorophyll per fresh leaf weight (µg/mg FW) is shown. To check if the amount of chlorophyll A and B per fresh leaf weight (µg/mg FW) for the diluted and undiluted samples was sufficient, the values for the diluted samples (diluted 10 times), when multiplied with 10, should be the same as for the undiluted samples. However, for chlorophyll A this was not the case, the values were often much higher. Therefore, the values of the diluted samples were used for further calculations. The amount of chlorophyll per fresh leaf weight (µg/mg FW) for the diluted samples of all control plants were seen as the maximum amounts of chlorophyll that could be present in leaves without damage caused by herbivores. Therefore, these amounts were set at 100%. For the infested plants it was calculated what percentage their amount of chlorophyll per fresh leaf weight was, compared to the control plants. So for example for chlorophyll A in the nonbitter cucumber C. sativus var corona, 0.045 is set at 100%, and 0.011 is then (0.011 ∗ 100) = 25% 0.045 So the amount of chlorophyll A per fresh leaf weight in infested non-bitter cucumber plants was 25%. This calculation was done for all plant species and showed that for all species, plants infested with spider mites had a lower chlorophyll A and chlorophyll B percentage than control plants (Table 3). The percentages of chlorophyll A and B in infested leaves of A. thaliana and different cucurbits from table 3 were shown in figure 7. The chlorophyll percentage of the control plants was always 100% and therefore not shown. For all plant species a decrease in chlorophyll percentage can be seen when the plant was infested with spider mites. Most of the time there was less chlorophyll A left in the leaves than chlorophyll B. How large the effect of herbivory was on the amount of chlorophyll left in the plant varied between the plant species. 15 Table 3 Amount of chlorophyll per fresh leaf weight (µg/mg FW) in percentages for control and infested leaves in A. thaliana and different cucurbits in diluted samples. Plant species C. sativus var corona C. sativus var Chinese long, 9930 C. melo (melon) C. lanatus (watermelon) C. pepo (courgette) C. maxima (pumpkin) M. charantia (bitter gourd) A. thaliana Treatment Diluted Amount of chlorophyll per fresh leaf weight (µg/mg FW) in percentages Chlorophyll A Chlorophyll B Average ± SD Average ± SD (%) (%) (%) (%) control infested control infested diluted diluted diluted diluted 100 25 100 71 10 5 18 6 100 25 100 80 10 5 21 4 control infested control infested control infested control infested control infested control infested diluted diluted diluted diluted diluted diluted diluted diluted diluted diluted diluted diluted 100 55 100 68 100 28 100 66 100 62 100 52 35 5 4 4 4 7 23 13 23 22 6 18 100 57 100 73 100 28 100 76 100 71 100 63 28 8 3 5 5 6 25 16 26 25 6 13 compared to Figure 7 Chlorophyll A and B percentage in leaves of A. thaliana and the different cucurbits that were infested with spider mites for about 15 days. The chlorophyll percentage of the control plants was always 100% and therefore not shown. 16 Gas chromatography–mass spectrometry The headspace of leaves and flowers of different cucurbits, induced by spider mites feeding on the leaves, consisted of 25 volatile compounds that did represent most of the volatile blend emitted by leaves and flowers (Table 4, 5, 6 and 7). The composition of the volatile blend varies between the plant species. Most of these compounds can be found in the infested as well as the control plants, especially in the flowers. In leaves, some volatiles were only emitted by infested plants: (-)-β-pinene, ((E)-β-)ocimene, linalool, (E)-4,8-dimethyl-1,3,7nonatriene [(E)-DMNT], methyl salicylate (MeSA), α-farnesene and (E,E)-4,8,12-trimethyl1,3,7,11-tridecatetraene [(E,E)-TMTT] (Table 4). The majority of the volatile compounds in both leaves and flowers were terpenes: about 75% of the whole blend, consisting of eight monoterpenes (α-pinene, (-)-β-pinene, (E)-β-ocimene, ocimene, linalool, linalool tetrahydride, nerol and (E)-2-decanal), nine sesquiterpenes (α-bourbonene, cedrene, α-cubebene, caryophyllene, α-farnesene, β-farnesene, β-ylangene, germacrene and nerolidol) and two homoterpenes (DMNT and TMTT). The remaining compounds were a phenylpropanoid (MeSA) and two aldehydes (nonanal and decanal). In flowers an aromatic alcohol (benzyl alcohol) and another aldehyde (benzaldehyde) were found, however, these compounds and the two aldehydes (nonanal and decanal) did not contain mass 93 like the other volatile compounds, which are terpenoids. In figures 8 and 9 the chromatograms of the volatile blends of the infested and control leaves of the cucumber species C. sativus var Chinese long, 9930 and C. sativus var corona are shown. For the Chinese bitter cucumber the volatile profile of the control and the infested leaves was quite different (Fig. 8). Mite feeding on leaves induced the emission of five volatiles not seen in control plants: (E)-β-ocimene, (E)-DMNT, methyl salicylate (MeSA), αfarnesene and (E,E)-TMTT. For compounds which are not in the biochemical class of terpenes like the aldehydes nonanal and decanal, the relative abundance was decreased when comparing the blend of infested to control leaves. Geranylacetone, on the other hand, had an increased relative abundance. 17 Table 4 Important volatiles in headspace of cucurbit leaves according to Metalign results and chromatograms. Within each plant species the headspace of infested leaves is compared with the headspace of control leaves. The production of a specific volatile in infested plants can be increased, decreased or is about the same level as in control plants. This is determined with the values given by Metalign and with comparing the relative abundance of the control and infested peaks in the chromatogram. Leaves Plant species C. sativus var corona Code Compound Biochemical class f g h o u v F G M N S y U A α-pinene (-)-β-pinene β-ocimene monoterpene monoterpene monoterpene monoterpene monoterpene monoterpene sesquiterpene sesquiterpene sesquiterpene sesquiterpene sesquiterpene homoterpene homoterpene phenylpropanoid (E)-β-ocimene x B I P linalool tetrahydride linalool α-bourbonene α-cubebene cedrene caryophyllene α-farnesene (E)-DMNT (E,E)-TMTT methyl salicylate (MeSA) nonanal decanal methyl perillate geranylacetone ↑ ↓ − ? * increase decrease about the same level this or other compound only in infested plant aldehyde aldehyde − C. sativus var Chinese long, 9930 − C. melo (melon) − ↑* − ↑* − ↑* ↑* ↑* ↑* ↑* ↑* ↑* ↑* ↑* ↑* ? ↑* C. lanatus (watermelon) ↑ ↑ and ↑* ↑* ↑* ↑* C. pepo (courgette) C. maxima (pumpkin) M. charantia ↑* − ? ↑* ? ↑* ? ↑* ? ↑* ↑* ↑* ↑ ?↑ ?↑ − ↑* ↑* ↑* ↑* ? ↑* ↑* ↑* ↓ ↓ ↓ ↓ ↓ ↓ − ↓ ↑ ↑ ↑ ↑ − ↑ − ↑ ↓ ↑ ↓ − ↑* − ■ Significant difference infested and control plant, fold change > 10 ■ Significant difference infested and control plant, fold change > 10, not always clear peak ■ Significant difference infested and control plant, fold change < 10 ■ No significant difference infested and control plant compared to 18 Table 5 Important volatiles in headspace of cucurbit male flowers according to Metalign results and chromatograms. Within each plant species the headspace of male flowers grown on infested plants is compared with the headspace of male flowers grown on control plants. The production of a specific terpene can be increased or decreased in flowers of infested plants or is about the same level as in flowers of control plants. This is determined with the values given by Metalign and with comparing the relative abundance of the control and infested peaks in the chromatogram. ♂ flowers Biochemical class Plant species C. sativus var Chinese long, 9930 Code Compound C. pepo (courgette) C. maxima (pumpkin) g m n v O Q R S T x B P β-pinene-(1S)-(-) nerol (E)-2-decanal linalool β-ylangene β-farnesene germacrene α-farnesene nerolidol nonanal decanal geranylacetone monoterpene monoterpene monoterpene monoterpene sesquiterpene sesquiterpene sesquiterpene sesquiterpene sesquiterpene aldehyde aldehyde ↑ ↓ increase decrease ■ Significant difference infested and control plant, fold change > 10 ■ Significant difference infested and control plant, fold change > 10, not always clear peak − about the same level ■ Significant difference infested and control plant, fold change < 10 ? * this or other compound only in infested plant ■ No significant difference infested and control plant M. charantia ?↓ ?↓ ↓ − ↑ ↑ − − ↑ ?↓ ↑ ↓ ↑ ↑ − ↑ ↑ ↑ ↑ ↓ − compared to 19 Table 6 Important volatiles in headspace of cucurbit female flowers according to Metalign results and chromatograms. Within each plant species the headspace of female flowers grown on infested plants is compared with the headspace of female flowers grown on control plants. The production of a specific terpene can be increased or decreased in flowers of infested plants or is about the same level as in flowers of control plants. This is determined with the values given by Metalign and with comparing the relative abundance of the control and infested peaks in the chromatogram. Code ♀ flowers Compound f g h n v Q S T A α-pinene β-pinene-(1S)-(-) β-ocimene (E)-2-decanal linalool β-farnesene α-farnesene nerolidol methyl salicylate (MeSA) x B P nonanal decanal geranylacetone ↑ increase ■ Significant difference infested and control plant, fold change > 10 ↓ decrease ■ Significant difference infested and control plant, fold change > 10, not always clear peak − about the same level ■ Significant difference infested and control plant, fold change < 10 Biochemical class monoterpene monoterpene monoterpene monoterpene monoterpene sesquiterpene sesquiterpene sesquiterpene phenylpropanoid aldehyde aldehyde Plant species C. sativus var corona C. sativus var Chinese long, 9930 M. charantia ?− ?− ↑ − ?− ↑ ↑ ↑ ↑ ↑* ↓ ↓ − ↑ ↑ ↑ this or other compound ■ No significant difference infested and control plant ? only in infested plant * No results of female flowers of the plant species C. pepo and C. maxima because no adult female flowers were grown on infested plants. ↓ ↓ ↓ compared to 20 Table 7 Important volatiles in headspace of cucurbit bisexual flowers according to Metalign results and chromatograms. Within each plant species the headspace of bisexual flowers grown on infested plants is compared with the headspace of bisexual flowers grown on control plants. The production of a specific terpene can be increased or decreased in flowers of infested plants or is about the same level as in flowers of control plants. This is determined with the values given by Metalign and with comparing the relative abundance of the control and infested peaks in the chromatogram. Code flowers Compound g u x B P β-pinene-(1S)-(-) linalool tetrahydride nonanal decanal geranylacetone Biochemical class Plant species C. melo (melon) monoterpene monoterpene aldehyde aldehyde − − ↑ ↑ ↓ ■ Significant difference infested and control plant, fold change > 10 ■ Significant difference infested and control plant, fold change > 10, not always clear peak ■ Significant difference infested and control plant, fold change < 10 ■ No significant difference infested and control plant ↑ increase ↓ decrease − about the same level compared to ? this or other compound * only in infested plant In the headspace of the non-bitter cucumber C. sativus var corona there was not much difference between the blend of control and infested leaves (Fig. 9). Mite feeding on leaves induced the emission of the volatile (E)-DMNT, which was not seen in control plants. For compounds which are not in the biochemical class of terpenes like the aldehydes nonanal and decanal, the relative abundance was decreased when comparing the blend of infested to control leaves. For geranylacetone the relative abundance stayed at about the same level. In figures 10, 11 and 12 the chromatograms of the volatile blends of the infested and control male and female flowers of the cucumber species C. sativus var Chinese long, 9930 and C. sativus var corona are shown. Two important volatiles which not belong to the terpenoid class were only seen in chromatograms of flowers: benzaldehyde and benzyl alcohol. For the Chinese bitter cucumber the volatile profile of the male flowers grown on control and infested plants was the same (Fig. 10). The relative abundance of the volatiles linalool, nonanal1 and geranylacetone was decreased when comparing the blend of male flowers on infested plants to male flowers on control plants. The compounds decanal, βfarnesene and nerolidol, on the other hand, had an increased abundance and for α-farnesene it stayed at about the same level. 1 Not sure nonanal is the right identity name for this volatile compound 21 Figure 8 Chromatogram of control and infested leaves of C. sativus var Chinese long, 9930 with the important volatile compounds (Table 4). Figure 9 Chromatogram of control and infested leaves of C. sativus var corona with the important volatile compounds (Table 4). 22 Figure 10 Chromatogram with the important volatile compounds emitted by male flowers grown on infested and control plants of the plant species C. sativus var Chinese long, 9930 (Table 5). And two important volatiles only emitted by the flowers: i = benzaldehyde and q = benzyl alcohol. (?x? not sure about identity). Figure 11 Chromatogram with the important terpene volatiles emitted by male flowers grown on infested and control plants of the plant species C. sativus var Chinese long, 9930 (Table 6). And two important volatiles only emitted by the flowers: i = benzaldehyde and q = benzyl alcohol. 23 Figure 12 Chromatogram with the important terpene volatiles emitted by female flowers grown on infested and control plants of the plant species C. sativus var corona (Table 6). And two important volatiles only emitted by the flowers: i = benzaldehyde and q = benzyl alcohol. (?S? not sure about identity). The female flowers of C. sativus var Chinese long, 9930 had the same volatile profile as the male flowers except for the compound linalool (Fig. 11). Linalool was not emitted and the infested female flowers produced methyl salicylate (MeSA) instead. The relative abundance of the volatiles β-farnesene, α-farnesene, nerolidol, nonanal, decanal and geranylacetone was increased when comparing the blend of female flowers on infested plants to female flowers on control plants. The volatile profile of female flowers of the non-bitter cucumber C. sativus var corona was the same for flowers grown on control and on infested plants (Fig. 12). It contained some of the compounds also found in female flowers of the Chinese bitter cucumber: nonanal, decanal, α-farnesene2 and geranylacetone. And it contained volatiles not seen in the volatile profile of the bitter cucumber: (E)-2-decenal and α-pinene or ocimene. For compounds which are not in the biochemical class of terpenes like the aldehydes nonanal and decanal, the relative abundance was decreased when comparing the blend of female flowers on infested plants to female flowers on control plants. The volatile (E)-2-decenal, on the other hand, had an increased abundance and for α-farnesene, geranylacetone and α-pinene or ocimene it stayed at about the same level. The chromatograms of leaves and flowers of the other cucurbits can be found in Appendix 3. The volatile profiles of leaves and flowers of A. thaliana are not shown, because something went wrong with the GC-MS machine while analysing the Tenax tubes containing the captured headspaces of this plant species. 2 Not sure α-farnesene is the right identity name for this volatile compound 24 Liquid chromatography–mass spectrometry Table 8 Amount of the endogenous secondary metabolites jasmonic acid (JA), jasmonoyl-isoleucine (JA-Ile), salicylic acid (SA) and abscisic acid (ABA) in pmol/g fresh sample weight in control and infested leaves and flowers of the cucumbers C. sativus var corona and C. sativus var Chinese long, 9930. JA JA-Ile SA Amount of endogenous secondary metabolite (pmol/g fresh weight) Plant species C. sativus var corona Treatment control infested control infested Plant material leaf leaf ♀ flower ♀ flower C. sativus var Chinese long, 9930 control infested control infested control infested leaf leaf ♂ flower ♂ flower ♀ flower ♀ flower ABA Average ± SD Average ± SD Average ± SD Average ± SD 0.939 6.127 0.461 0.603 0.072 0.578 0.067 0.043 ND* 1.326 2.844 0.483 ND* 0.103 0.314 0.060 13994 4077 10809 13904 5240 1576 1014 1037 5751 19531 58396 70677 589 1476 4283 4750 1.192 3.100 0.793 0.516 0.677 0.496 0.110 0.302 0.087 0.041 0.055 0.068 0.052 0.597 6.050 0.390 0.475 0.657 0.004 0.063 0.983 0.056 0.050 0.039 ND* 7920 23165 12874 13891 7155 ND* 5935 1448 1705 3039 752 52784 12451 76385 84540 53342 79562 3662 797 4765 5331 5170 11743 * ND = not detected 25 Figure 13 The amount of the endogenous secondary metabolites jasmonic acid (JA), jasmonoyl-isoleucine (JA-Ile), salicylic acid (SA) and abscisic acid (ABA) in control and infested leaves and female flowers of C. sativus var corona. 26 Figure 14 The amount of the endogenous secondary metabolites jasmonic acid (JA), jasmonoyl-isoleucine (JA-Ile), salicylic acid (SA) and abscisic acid (ABA) in control and infested leaves, male and female flowers of C. sativus var Chinese long, 9930. 27 The amount (pmol/g fresh weight) of the endogenous stress-related phytohormones jasmonic acid (JA), its bioactive amino acid conjugate (JA-Ile), salicylic acid (SA) and abscisic acid (ABA) in leaf and flower material of C. sativus var Chinese long, 9930 and C. sativus var corona, was shown in table 8. For each plant species plant material of three control and three infested plants was harvested and every sample was divided over three tubes, which served as technical replicates. For those technical replicates the average amount of each metabolite was taken, which did result in an amount of each metabolite within every plant. Eventually the average amount of each metabolite was calculated within the three control and the three infested plants of each plant species, which was shown in table 8. Figures 13 and 14 showed the content of table 8. The amount of the endogenous secondary metabolites JA and ABA in leaves and female flowers of C. sativus var corona increased when plants were infested with spider mites. The amount of salicylic acid in female flowers grown on infested non-bitter cucumber plants increased; however, in the leaves it decreased (Fig. 13). The amount of the metabolites JA, and SA in both male and female flowers of C. sativus var Chinese long, 9930 decreased when plants were infested with spider mites. In the leaves of those plants the amount of JA increased. Furthermore, the amount of ABA in male and female flowers grown on bitter cucumber plants increased; however, in the leaves it decreased (Fig. 14). The ratio of jasmonoyl-isoleucine (JA-Ile) and jasmonic acid (JA) in all plants parts of the plant species C. sativus var corona and C. sativus var Chinese long, 9930 is shown in table 9. The female flowers grown on infested plants of the non-bitter cucumber had a lower JA-Ile/JA ratio than the flowers on the control plants. The same applied to male flowers of the bitter cucumber. Female flowers grown on infested plants of the bitter cucumber and the leaves of these infested plants had a higher JA-Ile/JA ratio than the controls. Table 9 Ratio JA-Ile/JA in leaves, male and female flowers of C. sativus var corona and C. sativus var Chinese long, 9930. Plant species Treatment Plant material JA-Ile/JA ratio C. sativus var corona control infested control infested leaf leaf ♀ flower ♀ flower NP* 0.216 6.163 0.801 C. sativus var Chinese long, 9930 control leaf 0.043 infested leaf 0.192 control ♂ flower 7.625 infested ♂ flower 0.755 control ♀ flower 0.701 infested ♀ flower 1.326 * NP = not possible, because JA-Ile not detected for control leaves of C. sativus var corona. 28 Gene expression of TPS1, TPS9 and TPS19 genes The RNA concentration and the A260/A280 and A260/A230 values in control and infested leaf and flower material of the plant species C. sativus var corona and C. sativus var Chinese long, 9930 was checked with NanoDrop. For the leaf material of both species the RNA concentration and the A260/A280 and A260/A230 values were not sufficient. Therefore, leaves were not used for further experiments. The values for flowers on the other hand, were sufficient. For the flowers of each plant species and treatment, the threshold cycles of the genes of interest (TPS1, TPS19 and TPS9) were compared to the threshold cycle of the housekeeping gene actin to see how relative transcripts of each of the TPS genes were. The difference in number of threshold cycles was used to calculate the relative gene expression (RGE) and eventually the log2 ratio (Table 10). When the gene of interest needed a lower number of cycles to cross the threshold than actin, there was a higher amount of cDNA in the sample coding for that particular gene. Therefore, the RGE value was high. A gene of interest had a low RGE value when it needed more cycles to cross the threshold than actin, because of its low amount of cDNA present in the sample. The log2 ratio of a specific gene compared the RGEs of that gene between an infested sample and a control sample. TPS1 and TPS9 in the female flowers of the non-bitter cucumber had a positive log2 ratio while TPS19 had a negative log2 ratio (Fig. 15). The TPS1 and TPS19 genes had a negative log2 ratio in both male and female flowers of the Chinese cucumber (Fig. 16). Table 10 Average relative gene expression (RGE) of TPS genes TPS1, TPS19 and TPS9 in male and female flowers grown on control and infested plants of the plant species C. sativus var corona and C. sativus var Chinese long, 9930. And the log2 ratio calculated with the formula: log (RGE infested/RGE control;2). TPS gene Average RGE (relative gene expression) Plant species Treatment Plant material C. sativus var corona control infested control infested control infested ♀ flower ♀ flower ♀ flower ♀ flower ♀ flower ♀ flower 1 1 19 19 9 9 0.482 0.911 0.117 0.035 0.002 0.006 C. sativus var Chinese control long, 9930 infested control infested control infested control infested ♂ flower ♂ flower ♂ flower ♂ flower ♀ flower ♀ flower ♀ flower ♀ flower 1 1 19 19 1 1 19 19 23.772 2.428 0.087 0.041 27.617 0.816 0.367 0.024 log2 ratio 0.917 -1.756 1.834 -3.292 -1.106 -5.081 -3.933 29 compared to Figure 15 The log2 ratio of TPS genes TPS1, TPS19 and TPS9 in female flowers of the plant species C. sativus var corona. Figure 16 The log2 ratio of TPS genes TPS1and TPS19 in female flowers of the plant species C. sativus var Chinese long, 9930. No TPS 9 measurement for this cucumber species, because primers were not ready. 30 Discussion The objective of this study was to explore the variation among members of the plant family Cucurbitaceae with respect to flower characteristics that may be influenced by herbivory on leaves by spider mites (Tetranychus urticae). First of all it had to be confirmed that spider mites really caused damage to the plant they were feeding on. Damage did consist of chlorotic lesions the spider mites created on the leaves, which resulted in loss of chlorophyll content. Park and Lee (2002) show a reduction in chlorophyll content of leaves of C. sativus infested with spider mites. This loss of chlorophyll was also the case for all cucurbit species and A. thaliana included in this study. In most species, the reduction in chlorophyll A was larger than that of chlorophyll B. This is because in most green plants the total chlorophyll content has a chlorophyll A/B ratio of 3:1 (Raven et al., 2005). Therefore, in proportion to this ratio more chlorophyll A will be gone in infested leaves than chlorophyll B. How large the effect of herbivory was on the total amount of chlorophyll left in the plant varied between the plant species. Most probably, this is because of differences in the plant defence mechanisms that try to avoid that plants are eaten by herbivores. For C. sativus var Chinese long, 9930 it is known it contains cucurbitacins that are responsible for the bitter taste of the leaves. This will repel most individual herbivores (Miró, 1995). C. sativus var corona, on the other hand, does not contain these cucurbitacins and is therefore not bitter. This difference in bitterness most probably leads to the higher and lower amount of chlorophyll left in infested leaves of the bitter Chinese cucumber and the non-bitter cucumber, respectively. Leaves of the cultivated plant species C. melo (melon), C. lanatus (watermelon), C. pepo (courgette) and C. maxima (pumpkin) also showed a reduction in chlorophyll content. According to Miró (1995) melon, courgette and pumpkin contain cucurbitacins in different compositions (watermelon was not encountered). Courgette only contains one type of cucurbitacin (cucurbitacin E), while pumpkin and melon contain 3 (cucurbitacin B, D and E) and 4 types (cucurbitacin B, D, E and I), respectively. The amount of chlorophyll left in infested courgette leaves was lower than in infested leaves of pumpkin and melon, most probably because the latter have a better defence mechanism against herbivory. Cucurbitacins may play a role in plant defence, and the number of cucurbitacin types may have an effect on the effectiveness of plant defence against herbivores. There are also other plant components that can help protect the plant against attackers: M. charantia contains for example momordicines which make the leaves of the plant very bitter (Raman and Lau, 1996). Nevertheless, also this plant species showed a reduction in chlorophyll content when infested with spider mites. It can be concluded that spider mites really caused damage to all the plant species they were feeding on in this study. In a previous study about Cucumis sativus by Thomson and colleagues (2004) it is found that leaf herbivory by Helix aspersa during the flowering phase leads to a reduction in male flowers. In the study of Sigrid Dassen (2012) on the effect of leaf herbivory by spider mites on flower number on Cucumis sativus, it seemed there were a slight reduction and a slight increase in female flower and male flower number, respectively. In Cucurbita pepo (courgette), leaf herbivory resulted in a more male-biased sex ratio (Krupnick et al., 2000). So, it can be suggested that leaf herbivory could have an effect on flower number in different cucurbits. Therefore, flower number could be seen as another way to determine plant damage caused by herbivores. It would be interesting to investigate in a future study what the effect of leaf herbivory by spider mites is on flower number on the different cucurbits studied in this study. The non-bitter cucumber C. sativus var corona does not need to be pollinated to produce fruits (seedless fruits), however, other cucurbits depend on pollination (often by insects) to produce fruits (and seeds). If the flower sex ratio of those latter cucurbits is changed because of leaf herbivory, it could eventually have an effect on fruit production on the plant. 31 One of the characteristics that were investigated was the volatile emission by flowers and leaves grown on plants with and without leaf herbivory for the different cucurbits. The headspace of the flowers and leaves was analysed using TD-GCMS and a total of 25 volatile compounds were identified that did represent most of the volatile blend emitted by flowers and leaves. The majority of the volatile compounds in both leaves and flowers consisted of terpenes. The question is if spider-mite infestation changes the volatile blend emitted by flowers and leaves. And if so, is this change qualitative (release of new components) and/or quantitative (different emission level of components also present in non-infested plants)? With respect to the leaves there was quite some difference in the volatile profile of control and infested plants. Some volatiles were only emitted by infested plants, so induced by mite feeding: (-)-β-pinene, ((E)-β-)ocimene, linalool, (E)-4,8-dimethyl-1,3,7-nonatriene [(E)-DMNT], methyl salicylate (MeSA), α-farnesene and (E,E)-4,8,12-trimethyl-1,3,7,11tridecatetraene [(E,E)-TMTT]. This indicates that the change in leaf volatile profile when a plant is attacked by herbivores is qualitative (release of new components). There is also a quantitative change in leaf volatile profile: some volatiles were emitted by control as well as infested leaves and for some of those volatiles the emission level did decrease, while for most others it did increase. All cucurbits except M. charantia did emit some of those spider miteinduced volatile compounds. Most of these volatiles are known to attract the predatory mite Phytoseiulus persimilis, among them (E)-β-ocimene, linalool (E)-DMNT and methyl salicylate (Dicke et al., 1990) and it is reported that the presence of (E,E)-TMTT in a volatile blend does improve the attraction (De Boer et al., 2004). The results of this study did indicate that herbivore-damaged cucurbits emit herbivore-induced plant volatiles (HIPVs) in their leaves that can be used by natural enemies to locate their prey. Behavioural bioassays with predatory mites can provide significant evidence that the changed volatile profile of infested leaves of the different cucurbits really help attracting natural enemies. It can also be used to compare infested plants between the cucurbit species to see what kind of volatile blend, with their specific volatile profile, is the most effective to attract predatory mites. In this way there will maybe become more understanding about which compounds are used by natural enemies to locate their prey. In flowers on the other hand, there was almost no difference in the volatile profile of flowers grown on control compared to those grown on infested plants. Female flowers of mite-infested Chinese bitter cucumber plants did emit methyl salicylate while this was not emitted from these flowers when leaves were not infested. However, there was no significant difference between the amplitudes of this volatile in flowers grown on control and infested plants (Students t-test, P=0.49). This suggests that in flowers the change in volatile profile is not qualitative when a plant is attacked by herbivores (release of new components). Furthermore, there was not a convincing quantitative (different emission level of components also present in non-infested plants) change in volatile emission levels in flowers. Although spider mite-damaged vegetative tissue emitted herbivore-induced plant volatiles (HIPVs), there was no effect of herbivore damage on flower volatile emission in these cucurbits. Lucas-Barbosa and colleagues (2011) summarise that in several plantherbivore-combination studies plants respond to herbivory by making changes in flower volatile emission. In one of these studies it was found that leaf damage induces a significant increase of volatiles in male flowers, but not in female flowers of C. pepo (Theis et al., 2009). However, leaf damage was not caused by herbivores, but by making holes in the leaves that mimicked beetle damage. The flower volatile blend of tomato plants whose leaves where damaged by the caterpillar species (Manduca sexta) differed significantly form the blend of non-damaged plants (Solanum peruvianum) (Kessler and Halitschke, 2009). However, Effmert and colleagues (2008) found no effect of chewing damage by caterpillars (Manduca sexta) on flower volatiles of Nicotiana suaveolens. They did find an increase in leaf volatiles 32 like in this study. However, the study is about chewing insects and the timing of damage is much later (just before anthesis) than in this study. Mite-infestation already started when plants had developed about four or five expanded leaves, flower buds where just starting developing in the axils of the leaves. Headspace collection of male and female flowers of the plant species was about 20-30 days after first spider mite infestation. In the study of Sigrid Dassen (2012) on the effect of leaf herbivory by spider mites on volatile emission in flowers on Cucumis sativus, the infestation duration was about the same. This most probably indicates that infestation duration may not be the reason why there is almost no difference in the volatile profile of flowers grown on control compared to those grown on infested plants. However, in that study it was found that flowers of plants suffering from spider-mite herbivory released different volatile compounds and also at a different intensity (Sigrid Dassen, 2012). In a future study about the effect of leaf herbivory by spider mites on flower volatile emission in different cucurbits, it would be interesting to follow the volatile emission of leaves and flowers in time, because the total amount of volatile compounds emitted by a plant increases during spider-mite infestation and when herbivory progresses, other volatile compounds are released (Kappers et al., 2010). This indicates that during spider-mite infestation the emission of volatiles changes quantitatively and qualitatively. There could also be the possibility that the flowers grown on plants infested with spider mites emit the right volatile profile, but the control flowers do not. The control plants were grown in a clean climate chamber. However, there is always a chance herbivores manage to enter the room with accidental help by people, or when new plant material enters the room that might not be totally free of herbivores. There was no herbivore-damage visible on the leaves of the cucurbits, but thrips, for instance, are herbivores that feed on (developing) flowers, not on leaves. This florivory may induce an herbivore-induced volatile profile. Another characteristic that was investigated was the production of endogenous stress-related phytohormones examined with liquid chromatography–mass spectrometry. Leaf and flower material of the plant species C. sativus var Chinese long, 9930 and C. sativus var corona was examined for: jasmonic acid (JA), its bioactive amino acid conjugate (JA-Ile), salicylic acid (SA) and abscisic acid (ABA). These are phytohormones related to stress and their signalling pathways are responsible for plant resistance against: water and salt stress (ABA), pathogen attack (SA) and herbivore attack (JA), nevertheless the pathways can interact (Thaler and Bostock, 2004). Spider mites are herbivores and therefore induce specifically JA. However, these cell piercing herbivores may also induce SA-mediated defence genes (Zhang et al., 2009). The amount of the endogenous secondary metabolites JA and ABA in leaves and female flowers of C. sativus var corona was increased when plants were infested with spider mites. This increase in JA is to be expected when plants are attacked by herbivores. The amount of SA in infested leaves was decreased compared to control leaves. These results correspond to the interaction of the signalling pathways shown in figure 17: JA and SA act antagonistically (increase in JA, than decrease in SA); ABA has a positive relationship with JA (increase in ABA, than increase in JA); ABA has a negative relationship with SA (increase in ABA, than decrease in SA) (Thaler and Bostock, 2004). In contrast, the amount of SA in female flowers grown on infested non-bitter cucumber plants was increased compared to flowers on control plants. So, spider mites induced the production of JA in leaves of the nonbitter cucumber, while in the flowers both JA and SA are induced. There was a tremendous drop in JA-Ile/JA ratio in flowers when plants were infested. JA-Ile is the active form of JA and can make sure that plant defence genes are expressed (Thines et al., 2007). When there is less JA-Ile produced in infested flowers, less plant defence genes are expressed. This may 33 support the not-changed volatile profile of flowers grown on infested plants: infested flowers on non-bitter cucumber were most probably not able to produce plant-defence related volatiles. The volatile profile of infested leaves changed compared to the profile of control leaves: infested leaves on the non-bitter cucumber were able to produce herbivore-induced plant volatiles (HIPVs) that are able to attract natural enemies of the herbivore. This production of plant defence volatiles might suggests that in infested plants, JA turns into its active form JA-Ile, to make sure that defence genes come to expression (Thines et al., 2007). This supports an increase in JA-Ile/JA ratio when plants are infested by spider mites, and therefore a lower JA-Ile/JA ratio in control leaves (despite it was not possible to measure JAIle in leaves). Figure 17 Diagram of proposed relationship between abscisic acid, salicylic acid and jasmonic acid signalling pathways. Solid arrows indicate a positive regulating relationship between stresses and hormones. Dashed arrows indicate a positive (+) or negative (-) relationship between the hormonal signalling pathways (Thaler and Bostock, 2004). The amount of the endogenous secondary metabolite JA in infested leaves of C. sativus var Chinese long, 9930 was increased when compared to control leaves. This is also seen in the results of Sigrid Dassen, 2012. This increase in JA is to be expected when plants are attacked by herbivores. However, in both male and female flowers grown on infested plants the amount of JA was decreased. The ABA level in the infested plant parts was the opposite of the JA level in those plant parts: ABA was decreased in leaves and increased in flowers when infested with spider mites. These results do not correspond to the interaction of the signalling pathways shown in figure 17: ABA should have a positive relationship with JA (increase in ABA, than increase in JA) (Thaler and Bostock, 2004). This decrease of ABA in infested leaves is also in contradiction with the increased ABA level in infested leaves found by Sigrid Dassen, 2012. The amount of the metabolite SA in both the male and female flowers was decreased when plants were infested with spider mites. This increase in ABA and the decrease in SA in flowers correspond to the interaction of the signalling pathways shown in figure 17: ABA has a negative relationship with SA (increase in ABA, than decrease in SA) (Thaler and Bostock, 2004). The JA-Ile/JA ratio in infested leaves was increased compared to control leaves, which gives the indication that more JA turns into its active form to make sure that defence genes come to expression (Thines et al., 2007). This may support the change in volatile profile of infested leaves: infested leaves on the Chinese bitter cucumber were able to produce herbivore-induced plant volatiles (HIPVs) that are able to attract natural enemies of the herbivore. The JA-Ile/JA ratio in female flowers grown on infested plants was increased compared to those on control plants. In male flowers, however, there was a tremendous drop in JA-Ile/JA ratio when plants were infested. When there is less JA-Ile produced in infested male flowers, less plant defence genes are expressed. This may support the not-changed 34 volatile profile of male flowers grown on infested plants: infested male flowers on the Chinese bitter cucumber were most probably not able to produce plant-defence related volatiles. The last characteristic that was investigated was the gene expression of the terpene synthase family (TPS) genes TPS1, TPS9 and TPS19 in control and infested flower material of the plant species C. sativus var corona and C. sativus var Chinese long, 9930. TPS1, TPS9 and TPS19 encode for enzymes which synthesize the terpenes linalool, β-ocimene and αfarnesene, respectively (personal comment dr. ir. Iris F. Kappers). The log2 ratios of TPS1 and TPS9 in female flowers of C. sativus var corona were positive, indicating higher relative gene expressions of TPS1 and TPS9 in infested flowers. In theory, this would result in an induced emission of linalool and β-ocimene. TPS19 on the other hand, had a negative log2 ratio, which indicates a relative decrease of the transcription of the gene so in theory the emission of α-farnesene by infested flowers would be reduced. The log2 ratios of TPS1 and TPS19 in both male and female flowers of C. sativus var Chinese long, 9930 were negative, indicating lower relative gene expressions of TPS1 and TPS19 in infested flowers. In theory, this would result in a reduced emission of linalool and α-farnesene. In the study of Sigrid Dassen (2012) on the effects of spider-mite infestation of leaves on flower characteristics it was found that the expression of TPS19 was increased in female flowers, but reduced in male flowers. When comparing gene expression with volatile emission, it is shown that some results confirm each other while others are contradictory (Table 11). The volatile emission results showed that the male flowers of C. sativus var Chinese long, 9930 did emit less linalool when spider mites attacked the plant. On gene level, this observation is confirmed by a relative decrease in TPS1 transcripts in infested male flowers. This could indicate that this particular TPS gene could be responsible for the emission of linalool in male flowers of the bitter Chinese cucumber. However, the difference between infested and control male flowers for linalool in the volatile emission results was not significant (Students t-test, P=0.076). The female flowers of C. sativus var Chinese long, 9930 did emit more linalool and αfarnesene when spider mites attacked the plant. However, the difference between infested and control female flowers for linalool and α-farnesene in the volatile emission results was not significant (Students t-test, linalool P=0.438, α-farnesene P=0.526). On gene level, this observation of more linalool and α-farnesene emission is not confirmed, because of the relative decreases in TPS1 and TPS19 transcripts in infested female flowers. So the gene expression and volatile emission for female flowers of the bitter Chinese cucumber are contradictory. The terpene synthase family (TPS) is a set of related genes that encodes for enzymes which synthesize different terpenes (Bohlmann et al., 1998). Until now 26 TPS genes are known for cucumber. However, it is not yet known for all genes which terpene(s) they can synthesize. TPS1, TPS9 and TPS19 encode for enzymes which synthesize the terpenes linalool, β-ocimene and α-farnesene, respectively. Nevertheless, it can be that one TPS gene encodes for various terpenes and that the production of a specific terpene can be activated by more than one individual TPS gene. The latter is true for linalool (personal comment dr. ir. Iris F. Kappers). Therefore, the increase in linalool and α-farnesene emission in infested female flowers of the bitter Chinese cucumber could maybe be confirmed on gene level when the gene expression of other TPS genes is investigated. 35 Table 11 Comparison of the results of the gene expression and the volatile emission experiments. Flowers grown on infested plants are compared with flowers grown on control plants. Plant species Plant material TPS gene Compound Experiment Gene Volatile expression emission C. sativus var corona ♀ flower 1 19 9 1 linalool α-farnesene β-ocimene linalool ↑ ↓ ↑ ↓ − ?− ?− ↓ 19 1 19 α-farnesene linalool α-farnesene ↓ ↓ ↓ − ↑ ↑ C. sativus var Chinese long, ♂ flower 9930 ♀ flower ↑ ↓ − ? increase decrease about the same level this or other compound ■ results of experiments are the same ■ results of experiments are opposite In conclusion, this study demonstrates a large range of qualitative and quantitative variation in volatile emission induced by leaf herbivory of spider mites (Tetranychus urticae) in leaves of different cucurbit species. Furthermore, this study found no significant effects of herbivore damage on flower volatile emission in these cucurbits. The production of endogenous stressrelated phytohormones varied between the plant species C. sativus var corona and C. sativus var Chinese long, 9930 and between the plant parts. The female flowers of C. sativus var corona and the male flowers of C. sativus var Chinese long, 9930 grown on infested plants were most probably not able to produce plant-defence related volatiles, because of the tremendous drop in JA-Ile/JA ratio in flowers when plants were infested. When comparing the gene expression of TPS1, TPS9 and TPS19 with the volatile emission of linalool, βocimene and α-farnesene in flowers of C. sativus var corona and C. sativus var Chinese long, 9930, it was shown that spider mites reduced the emission of linalool as well as the gene expression responsible for the production of linalool in male flowers of the Chinese bitter cucumber. However, for the other compound-flower combinations the comparison ended in contradictions. 36 Acknowledgements I would like to thank Harro for allowing me to do this research at the Plant Physiology group. It is a big group with people from all kinds of places having knowledge on various topics. Still, it is one group and I met nice people and had very nice experiences during lunch time and coffee breaks, and especially during the barbeque and lab trip. Iris, thank you very much for introducing me to this interesting topic with cucumber and its relatives in the spotlights. I learned many new things about cucurbits, which I could not have learned during my part-time job at the greengrocery De Komkommerin on Saturdays. I did appreciate it that you just let me figure out things by myself, because that is what research is all about: make mistakes, learn from them and hopefully do things right the next time. I did also appreciate it when you pushed me in the right direction in case I was stuck or too focused on one thing and almost forgetting the rest. I would also like to thank Kristyna, Yuanyuan and Jun for helping me with experiments and advising or teaching me when I had questions. This study stimulates me to get to know more about plants and the interaction with their environment (especially insects). The next challenge will be a consultancy project about plant metabolites in sweet pepper plants. 37 Appendix 1 RNA isolation from Cucumber and cDNA synthesis for qPCR Grind ± 100 mg leaf material in liquid nitrogen to powder In the fumehood: Add 1 mL TriPure & homogenize on the vortex, Let stand for 5 min at RT (Tripure contains phenol!) Add 200 µl Chloroform, vortex, let stand for 5 min at RT Centrifuge for 15 min, 12.000xg, 4 °C Pipet the supernatant in a fresh tube Add 0.5 Volume of 96 % EtOH, Mix by inverting Apply to the column of the RNA box (red box) Wash with the wash buffer (Follow the protocol in the Kit!) Step 1: RW1 (1x) Step 2: RP (2x) Elute RNA with 200 µl water from the kit DNAse treatment: 89 µl RNA 10 µl DNAse buffer 1 µl DNAse 15 min at 25 °C Apply onto a RNA column to purify (Follow the protocol in the Kit!) Wash and elute as described above Measure concentration on the nanodrop Use 1 µg of RNA to synthesize iScript DNA: 4 µl 5X iScript reaction Mix 1 µl iScript Reverse Transcriptase X µl Nuclease free water X µl RNA-template (100 fg-1µg total RNA) 5 min 25 °C 30 min 42 °C 5 min 85 °C 38 Appendix 2 Protocol for Quantitative Real-Time PCR 1. Isolate RNA following the protocol from Ralph “RNA isolation Tripure-small sample”. For Research Use Only. Ralph Litjens © All rights reserved _ 1 _ R--11 RNA isolations ((TriPure) – small sample) 1) Harvest 2 leave disks, (diameter 1.5 cm – weight about 80 mg) 2) and freeze in liquid nitrogen (store -80ºC) 3) Grind leave disks in 1.5 ml tube, using the “blue potter” 4) Add 500 ul TriPure (Roche) 5) Homogenize using the homogenizer with the “steel white tip potter”. 6) Add 500 ul TriPure and vortex 7) Centrifuge 1 min microcentrifuge max speed at room temperature 8) Transfer supernatant into a fresh tube (2 ml) (Then, prepare chloroform, isopropanol and 75% ethanol in fresh 50 ml blue-cap tubes.) 9) Add 0.2 ml chloroform and vortex for 15 seconds 10) Incubate at room temperature for 2-15 minutes 11) Centrifuge 15 min microcentrifuge 12,000 g at 4ºC 12) Transfer the supernatant to a fresh tube (1.5 ml) 13) Add 500 ul isopropanol and vortex 14) Incubate at room temperature for 10 minutes 15) Centrifuge 10 min microcentrifuge max speed at 4ºC 16) Remove supernatant and wash the pellet with 800 ul 75% EtOH 17) Centrifuge 5 min microcentrifuge max speed at 4ºC 18) Remove supernatant and dry the pellet briefly (less than 3 min) 19) (not too dry because it will be hard to dissolve the RNA pellet) 20) Dissolve pellet in 95 ul MQ (keep on ice from this moment) (Normally, dissolve pellet in 30-50 µl MQ. However, 90 µl perfectly fit the following steps.) 21) Incubate 5 minutes at 60ºC to dissolve the RNA completely 22) Centrifuge 10 min microcentrifuge max speed at 4ºC 23) Transfer the supernatant to a fresh tube (1.5 ml). 24) Store the RNA at -70ºC For tobacco leaves………….. up to 100 µg total RNA could be isolated 2. Check the RNA quality by running 5 µl of RNA samples on gel to see whether there are sharp bands. (The RNA loading buffer is in the general stock in -20°C. The gel and running buffer are preferably to be freshly prepared.) 3. Treated the RNA samples with DNAse. - RNA 90 µl - DNAse buffer 10 µl - DNAse I 1 µl Then incubate 15 min at 25 °C Then add EDTA (25mM) 1 µl, incubate 10 min at 50 °C to stop the reaction. (if immediately clean RNA, EDTA treatment is not necessary! ) 39 4. Purify the RNA by “RNA easy Mini Handbox” following the protocol in the box. 5. Measure the concentration of RNA after purification by Nanodrop. If the values of 260/280 and 260/230 are above 1.8, the purity and quality of RNA is good for real time PCR. (If the quality is not good enough, such as the value of 260/230 is quite low, purify that RNA sample with the kit again. ) 6. Reverse transcription on PCR machine. a. Calculate the amount of RNA solution needed for 1 µg RNA. The system was optimized for 1 µg RNA. (The maximum amount of RNA solution is 15 µl in the following reaction system. If the RNA concentration is too low and even 15 µl RNA solution can not reach 1 µg, this RNA sample should be concentrated by “Speed Vacuum”.) - iScript cDNA synthese kit from Biorad (20µl) - 5×iScript reaction Mix 4 µl - iScript reverse transcriptase 1 µl - RNA sample+ MQ water 15µl b. Reverse transcription programme: 25 °C 5min (annealing of primer) 42 °C 30 min (extension) 85 °C 5 min (kill the RTase) Hold at 10 °C 7. Measure the concentration of cDNA by Nanodrop. The concentrations are around 1000 ng/µl. The differences of concentration among all samples should be less than 100 ng/µl. 8. Dilute cDNA to the volume of 200ul, or 400 (Depending) with fresh MQ water. 9. Prepare RT-PCR mixture. a. Get fresh MQ water in fresh 50 ml blue-cap tube. (First turn on the tap of MQ machine, and let the MQ water run for a few seconds. Then load the MQ water in the tube.) b. Prepare RT-PCR mixture in a flow cabinet, but do not turn on the flow cabinet. triplicates duplicates - Fresh MQ water 13 µl 11.5µl - RT-PCR Buffer (2×) 32.5 µl 22.5µl - Forward primer (3 µM) 6.5 µl 4.5µl - Reverse primer (3 µM) 6.5 µl 4.5µl - cDNA template 6.5 µl 2µl - ------------------------------------------------------------------------+) 65 µl +) 45 µl Prepare this mixture in a 0.5 ml tube. Note: i. “RT-PCR Buffer” mentioned above was “BIO-RAD iQTM SYBR@ Green Supermix 2× Mix for Real-Time PCR 100 × 50 µl reactions, Catalog 170-8880”. This buffer should be stored at -20 40 °C. But once it’s opened, it should be stored in 4 °C. It will be stable at 4 °C for at least 6 months. ii. The primers were dissolved in MQ water. The primers were designed by the software “Beacon Designer” which is especially for designing primers for RT-PCR. For more details, please see the “Protocol for designing primers for RT-PCR”. iii. The tips used in this and following steps are special tips which have filters inside. iv. The pipettes used in this and following steps are preferably those pipettes only used for RT-PCR work. (One set of these pipettes can be found in the drawer near qPCR machine.) v. Always prepare a mixture of “no template control (NTC)” by adding MQ water instead of cDNA template and a mixture of “RNA template control” by adding RNA instead of cDNA template. 10. Then divide the 65 µl mixture into 3 wells (20 µl/ well) in the RT-PCR plate as triplicates. When dividing the mixture, cover the other wells you are not working with! 11. Carefully seal the RT-PCR plate with special film (Microseal, adhesive sealer). When taking out the film, only hold its edge which is not transparent. Do not touch the transparent part of the film! After the film sticking to the RP-PCR plate, use a small wooden board (can be found in the top drawer near RT-PCR machine) to scrape the film several times. 12. Centrifuge the RT-PCR plate in Taqman centrifuge gently in the shaker room for several minutes to spin down the droplets attaching to the wall of the wells. 13. Run RT-PCR machine. a. Turn on the RT-PCR machine at least 10 min in advance to preheat it. Then turn on the computer connected to the machine. b. Put your RT-PCR plate in the machine. c. The short cut of the software “Bio-red iQ5” can be found on the desktop of the computer. Double click it. d. Click “Protocol”. Then choose the file ”C:\ Program Files\ Bio-Rad\ iQ5\ users\ Jos (or Liping)\ 2stepAmp+MelJos.tmo” e. Click “Plate”. i. Then choose the file “C:\ Program Files\ Bio-Rad\ iQ5\ users\ Jos (or Liping)\ JosEVSol5.pls” ii. Click “Create New”. a) “Sample volume” is 20 µl. b) “Seal Type” is film. c) “Vessel Type” is plates. d) Click button . Then choose the well positions of NTC. (Left click mouse, then drag the mouse through those 3 wells. Do not 41 click each well one by one. ) e) Click button . Then choose the well positions of samples. (Left click mouse, then drag the mouse through 3 wells which are triplicates of one sample. Then drag the mouse through another 3 wells which belong to another sample.) f) If there are some wrongly chosen wells, just click to erase them. g) Then click “Save & Exit Plate Editing”. Save it in your own file folder. f. Click “Run”. Choose “Collect well…” g. Click “Begin Run”. 14. View the RT-PCR results about one and a half hour later. a. Click “Data File”. Then choose the data file you saved. b. Click “Data analysis”. Now you are in the “PCR Quant” window. c. Click “Results”. Then copy the result table and paste it in an Excel file for further data analysis. Note: If you would like to know whether the primers are good, click “Melt Curve/Peak”. In “Melt Peak Chart”, if there is only one peak for each sample, that means there is an unique product and there is not unspecific amplification. 15. Analyze data in Excel file. a. Copy the raw data to another Excel sheet, and name it “RGE” (relative gene expression). Do not work on the raw data directly. b. Fill in the “Identifier” column with sample name and primer name (such as “WT- actin”). Then it’s easy to see which well is which. c. Keep “Identified”, “Threshold”, “Replicate #” and “Ct Std. Dev” columns, and remove the other columns which are not necessary for data analysis. d. Calculate the “δ Ct”. Ct value (threshold cycle) of gene of interest subtracts the Ct value of house-keeping gene for the same sample. e. Calculate the “RGE” by using the function of “f(x)=POWER(2,- δ Ct)”. 42 f. Calculate the “mean RGE”. Since there are 3 replicates for each sample, so there are 3 “RGE” value for each sample. The “mean RGE” is the average of those 3 “RGE” value. g. Calculate the “RGE Sta. Dev.” by using the function of “f(x)= =STDEV(RGE1, RGE2, RGE3)” h. Make a chart with type of “Clustered column”. The χ-axis is the sample name and the у-axis is the “mean RGE” value. The error bar is the “RGE Sta. Dev.” value. Do log scale for у-axis. (Right click on у-axis→ click “Format Axis” → click “Scale” → choose “Logarithmic scale” .) For example (fig 1) i. The data of REG value can also be *100 (or 1000 times, or more), then do the Log(X, 2) to make the columns better to show, the last data show the fold number of different genes. j. The REG of the transgenic can also be divided to WT to show the fold difference. For example: (fig 2) 100.00 10.00 1.00 w t-OC 1-OC 2-OC 3-OC 4-OC 5-OC 6-OC 7 OC1 8-OC 9-OC 10 OC1 11 OC1 12-OC 13-OC 14-OC 15-OC 0.10 0.01 0.00 chry FDS, CDS 16 REG fold 14 12 10 1581JA-leaf 1581JA+leaf 8 1581JA-ovary 6 1581JA+ovary 4 2 0 chrCDS chrFDS1 chrFDS2 For more details about analyzing data, please refer to “Kenneth J. Livak and Thomas D. Schmittgen . Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2- δCt CT Method. METHODS 25, 402–408 (2001) ” Written by: Ting Modified by Jing 43 Appendix 3 Chromatograms of leaves and flowers of other cucurbit plant species. Figure 18 Chromatogram of control and infested leaves of C. melo (melon) with the important volatile compounds (Table 4). Figure 19 Chromatogram of control and infested leaves of C. lanatus (watermelon) with the important volatile compounds (Table 4). 44 Figure 20 Chromatogram of control and infested leaves of C. pepo (courgette) with the important volatile compounds (Table 4). Figure 21 Chromatogram of control and infested leaves of C. maxima (pumpkin) with the important volatile compounds (Table 4). 45 Figure 22 Chromatogram of control and infested leaves of M. charantia (bitter gourd) with the important volatile compounds (Table 4). Figure 23 Chromatogram with the important volatile compounds emitted by male flowers grown on control and infested plants of the plant species C. pepo (courgette) (Table 5). 46 Figure 24 Chromatogram with the important volatile compounds emitted by male flowers grown on control and infested plants of the plant species C. maxima (pumpkin) (Table 5). Figure 25 Chromatogram with the important volatile compounds emitted by male flowers grown on control and infested plants of the plant species M. charantia (Table 5). 47 Figure 26 Chromatogram with the important volatile compounds emitted by female flowers grown on control and infested plants of the plant species M. charantia (Table 6). Figure 27 Chromatogram with the important volatile compounds emitted by bisexual flowers grown on control and infested plants of the plant species C. melo (melon) (Table 7). 48 Appendix 4 Table 12 Amount of chlorophyll per fresh leaf weight (µg/mg FW) for control and infested leaves in A. thaliana and different cucurbits in undiluted and diluted samples Amount of chlorophyll per fresh leaf (µg/mg FW) Plant Chlorophyll A Chlorophyll B ± SD Average species Treatment (Un)diluted Average C. sativus control undiluted 0.188 0.018 0.318 * var corona diluted 0.045 0.005 0.020 infested undiluted 0.105 0.020 0.055 * diluted 0.011 0.002 0.005 C. sativus control undiluted 0.192 0.035 0.231 * var Chinese diluted 0.035 0.006 0.014 long, 9930 infested undiluted 0.183 0.017 0.163 * diluted 0.025 0.002 0.011 C. melo control undiluted 0.185 0.028 0.263 (melon) diluted 0.038* 0.013 0.018 infested undiluted 0.188 0.017 0.120 diluted 0.021* 0.002 0.010 C. lanatus control undiluted 0.174 0.021 0.408 (watermelon) diluted 0.054* 0.002 0.025 infested undiluted 0.217 0.018 0.262 * diluted 0.037 0.002 0.019 C. pepo control undiluted 0.149 0.011 0.240 * (courgette) diluted 0.034 0.002 0.014 infested undiluted 0.095 0.022 0.047 * diluted 0.010 0.002 0.004 C. maxima control undiluted 0.213 0.032 0.157 * (pumpkin) diluted 0.029 0.007 0.012 infested undiluted 0.168 0.025 0.105 diluted 0.019* 0.004 0.009 M. charantia control undiluted 0.199 0.012 0.464 (bitter diluted 0.063* 0.015 0.029 gourd) infested undiluted 0.211 0.011 0.304 diluted 0.039* 0.014 0.020 A. thaliana control undiluted 0.212 0.035 0.294 * diluted 0.040 0.002 0.019 infested undiluted 0.169 0.040 0.155 * diluted 0.021 0.007 0.012 weight ± SD 0.029 0.002 0.011 0.001 0.063 0.003 0.014 0.001 0.101 0.005 0.016 0.001 0.020 0.001 0.029 0.001 0.017 0.001 0.013 0.001 0.046 0.003 0.027 0.002 0.097 0.007 0.140 0.007 0.033 0.001 0.048 0.003 * Values of diluted samples multiplied with 10 should give the same values as for the undiluted samples, however, they did not. 49 References Adler LS, Karban R, Strauss SY (2001) Direct and indirect effects of alkaloids on plant fitness via herbivory and pollination. Ecology 82: 2032-2044 Agrawal AA, Gorski PM, Tallamy DW (1999) Polymorphism in plant defense against herbivory: constitutive and induced resistance in Cucumis sativus. Journal of Chemical Ecology 25: 2285-2304 Aharoni A, Jongsma MA, Bouwmeester HJ (2005) Volatile science? Metabolic engineering of terpenoids in plants. Trends in Plant Science 10: 594-602 Arimura G, Kost C, Boland W (2005) Herbivore-induced, indirect plant defences. Biochimica et Biophysica Acta 1734: 91-111 Aubourg S, Lecharny A, Bohlmann J (2002) Genomic analysis of the terpenoid synthase (AtTPS) gene family of Arabidopsis thaliana. Molecular Genetics and Genomics 267: 730745 Balkema-Boomstra AG, Zijlstra S, Verstappen FWA, Inggamer H, Mercke PE et al. (2003) Role of cucurbitacin in resistance to spider mite (Tetranychus urticae) in cucumber (Cucumis sativus L.). Journal of Chemical Ecology 29: 225–235 Barber NA, Adler LS, Bernardo HL (2011) Effects of above- and belowground herbivory on growth, pollination, and reproduction in cucumber. Oecologia 165: 377-386 Bohlmann J, Meyer-Gauen G, Croteau R (1998) Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proceedings of the National Academy of Sciences of the United State of America 95: 4126-4133 Bouwmeester HJ, Verstappen FWA, Posthumus MA, Dicke M (1999) Spider mite-induced (3S)-(E)-nerolidol synthase activity in cucumber and lima bean – the first dedicated step in acyclic C11-homoterpene biosynthesis. Plant Physiology 121: 173-180 Chen F, Ro D-K, Petri J, Gershenzon J, Bohlmann J et al. (2004) Charaterization of rootspecific Arabidopsis terpene synthase responsible for the formation of the volatile monoterpene 1,8-cineole. Plant Physiology 135: 1956-1966 Chen F, Tholl D, Bohlmann J, Pickersky E (2011) The family of terpene synthases in plants: a mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. The Plant Journal 66: 212-229 Da Costa CP, Jones CM (1971) Cucumber beetle resistance and mite susceptibility controlled by the bitter gene in Cucumis sativus L. Science 172: 1145-1146 De Boer JG, Posthumus MA, Dicke M (2004) Identification of volatiles that are used in discrimination between plants infested with prey or nonprey herbivores by a predatory mite. Journal of Chemical Ecology 30: 2215-2230 De Moraes CM, Lewis WJ, Pare PW, Alborn HT, Tumlinson JH (1998) Herbivore-infested plants selectively attract parasitoids. Nature 393: 570-573 50 De Moraes CM, Mescher MC, Tumlinson JH (2001) Caterpillar-induced nocturnal plant volatiles repel nonspecific females. Nature 410: 577-580 De Vos RC, Schipper B, Hall RD (2012) High-performance liquid chromatography-mass spectrometry analysis of plant metabolites in Brassicaceae. Methods in Molecular Biology 860:111-28. Dicke M (2009) Behavioural and community ecology of plants that cry for help. Plant, Cell and Environment 32: 654-665 Dicke M, Baldwin IT (2010) The evolutionary context for herbivore-induced plant volatiles: beyond the ‘cry for help’. Trends in Plant Science 15: 167–175 Dicke M, van Beek TA, Posthumus MA, Dom NB, van Bokhoven H et al. (1990) Isolation and identification of volatile kairomone that affects acarine predator-prey interactions involvement of host plant in its production. Journal of Chemical Ecology 16: 381-396 Dicke M, van Poecke RMP, de Boer JG (2003) Inducible indirect defence of plants: from mechanisms to ecological functions. Basic and Applied Ecology 4: 27-42 Dudareva N, Pickersky E, Gershenzon J (2004) Biochemistry of plant volatiles. Plant Physiology 135: 1893-1902 Effmert U, Dinse C, Piechulla B (2008) Influence of green leaf herbivory by Manduca sexta on floral volatile emission by Nicotiana suaveolens. Plant Physiology 146: 1996-2007 Fasulo TR, Denmark HA (2009). Featured creatures, twospotted spider mite. Available: http://entomology.ifas.ufl.edu/creatures/orn/twospotted_mite.htm. Consulted in March 2014. Flokova K, Tarkowska D, Miersch O, Strnad M, Wasternack C et al. (2014) UHPLC-MS/MS based target profiling of stress-induced phytohormones. Phytochemistry 105: 147-157 Garcia-Mas J, Benjak A, Sanseverino W, Bourgeois M, Mir G et al. (2012) The genome of melon (Cucumis melo L.). Proceedings of the National Academy of Sciences of the United State of America 109: 11872-11877 Gould F (1978) Resistance of cucumber varieties to Tetranychus urticae: Genetic and environmental determinants. Journal of Economic Entomology 71: 680-683 Heil M, Ton J (2008) Long-distance signalling in plant defence. Trends in Plant Science 13: 264-272 Herms DA, Mattson WJ (1992) The dilemma of plants: to grow or to defend. The Quarterly Review of Biology 67: 283–335 Hilker M, Meiners T (2010) How do plants “noticce” attack by herbivorous arthropods? Biological Reviews 85: 267-280 Huang S, Li R, Zhang Z, Li L, Gu X et al. (2009) The genome of the cucumber, Cucumis sativus L. Nature Genetics 41: 1275-U29 51 Kappers IF, Hoogerbrugge H, Bouwmeester HJ, Dicke M (2011) Variation in herbivoryinduced volatiles among cucumber (Cucumis sativus L.) varieties has consequences for the attraction of carnivorous natural enemies. Journal of Chemical Ecology 37: 150-160 Kappers IF, Verstappen FWA, Luckerhoff LLP, Bouwmeester HJ, Dicke M (2010) Genetic variation in jasmonic acid- and spider mite-induced plant volatile emission of cucumber accessions and attraction of the predator Phytoseiulus persimilis. Journal of Chemical Ecology 36: 500-512 Kessler A, Baldwin IT (2002) Plant responses to insect herbivory: the emerging molecular analysis. Annual Review of Plant Biology 53: 299-328 Kessler A, Halitschke R (2009) Testing the potential for conflicting selection on floral chemical traits by pollinators and herbivores: predictions and case study. Functional Ecology 23: 901-912 Knudsen JT, Eriksson R, Gershenzon J, Ståhl B (2006) Diversity and distribution of floral scent. Botanical Review 72: 1-120 Knudsen JT, Tollsten L, Bergström G (1993) Floral scents – a checklist of volatile compounds isolated by head-space techniques. Phytochemistry 33: 253-280 Krips OE, Kleijn PW, Willems PEL, Gols GJZ, Dicke M (1999) Leaf hairs influence searching efficiency and predation rate of the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). Experimental and Applied Acarology 23: 119-131 Krupnick GA, Avila G, Brown KM, Stephenson AG (2000) Effects of herbivory on internal ethylene production and sex expression in Cucurbita texana. Functional Ecology 14: 215-225 Lambers H, Chapin III FS, Pons TL (2008) Plant physiological ecology, 2nd edn. Springer Science+Business Media, New York, USA. pp. 445-477. Lucas-Barbosa D, van Loon JJA, Dicke M (2011) The effects of herbivore-induced plant volatiles on interactions between plants and flower-visiting insects. Phytochemistry 72: 16471654 Mercke P, Kappers IF, Verstappen FWA, Vorst O, Dicke M et al. (2004) Combined transcript and metabolite analysis reveals genes involved in spider mite induced volatile formation in cucumber plants. Plant Physiology 135: 2012-2024 Miró M (1995) Cucurbitacins and their pharmacological effects. Phytotherapy Research 9: 159-168 Mothershead K, Marquis RJ (2000) Fitness impacts of herbivory through indirect effects on plant-pollinator interactions in Oenothera macrocarpa. Ecology 81: 30-40 Mumm R, Dicke M (2010) Variation in natural plant products and the attraction of bodyguards involved in indirect plant defense. Canadian Journal of Zoology 88: 628–667 52 Njoroge GN, Van Luijk MN (2004) Momordica charantia L. Available: http://www.prota4u.org/protav8.asp?h=M4&t=Momordica,charantia&p=Momordica+charanti a#Synonyms. Consulted in March 2014. Park YL, Lee JH (2002) Leaf cell and tissue damage of cucumber caused by twospotted spider mite (Acari: Tetranychidae). Journal of Economic Entomology 95: 952-957 Park YL, Lee JH (2005) Impact of twospotted spider mite (Acari: Tetranychidae) on growth and productivity of glasshouse cucumbers. Journal of Economic Entomology 98: 457–463 Pichersky E, Gershenzon J (2002) The formation and function of plant volatiles: perfumes for pollinator attraction and defence. Current Opinion in Plant Biology 5: 237-243 Pierce LK, Wehner TC (1990) Review of genes and linkage groups in cucumber. Hortscience 25: 605-615 Pieterse CMJ, Dicke M (2007) Plant interactions with microbes and insects: from molecular mechanisms to ecology. Trends in Plant Science 12: 564-569 Raman A, Lau C (1996) Anti-diabetic properties and phytochemistry of Momordica charantia L. (Cucurbitaceae). Phytomedicine 2: 349-362 Raven PH, Evert RF, Eichhorn SE (2005) Biology of plants, 7th edn. W.H. Freeman and Company, New York, USA, pp 119-121. Rice CA, Rymal KS, Chambliss OL, Johnson FA (1981) Chromatographic and mass-spectral analysis of cucurbitacins of 3 Cucumis sativus cultivars. Journal of Agricultural and Food Chemistry 29: 194-196 Schoonhoven LM, van Loon JJA, Dicke M (2005) Insect-plant biology. Oxford University Press, Oxford Sigrid Dassen (2012) Volatiles of male and female flowers of Cucumis sativus are differentially affected by Tetranychus urticae leaf herbivory. Thesis. Takabayashi J, Dicke M (1996) Plant-carnivore mutualism through herbivore-induced carnivore attractants. Trends in Plant Science 1: 109–113 Takabayashi J, Dicke M, Takahashi S, Posthumus MA, van Beek TA (1994) Leaf age affects composition of herbivore-induced synomones and attraction of predatory mites. Journal of Chemical Ecology 20: 373-386 Thaler JS, Bostock RM (2004) Interactions between abscisic-acid-mediated responses and plant resistance to pathogens and insects. Ecology 85: 48-58 Theis N, Kesler K, Adler LS (2009) Leaf herbivory increase floral fragrance in male but not female Cucurbita pepo subsp. texana (Cucurbitaceae) flowers. American Journal of Botany 96: 897-903 53 Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A et al. (2007) JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signalling. Nature 448: 661–666 Tholl D, Boland W, Hansel A, Loreto F, Röse USR et al. (2006) Practical approaches to plant volatile analysis. The Plant Journal 45: 540-560 Tholl D, Chen F, Petri J, Gershenzon J, Pickersky E (2005) Two sesquiterpene synthases are responsible for the complex mixture of sesquiterpenes emitted from Arabidopsis flowers. The Plant Journal 42: 757-771 Thomson VP, Nicotra AB and Cunningham SA (2004) Herbivory differentially affects male and female reproductive traits of Cucumis sativus. Plant Biology 6: 621-628 Trapp SC, Croteau RB (2001) Genomic organization of plant terpene synthases and molecular evolutionary implications. Genetics 158: 811-832 Traw MB, Dawson TE (2002) Differential induction of trichomes by three herbivores of black mustard. Oecologia 131: 526-532 Van den Boom CEM, van Beek TA, Dicke M (2002) Attraction of Phytoseiulus persimilis (Acari: Phytoseiidae) towards volatiles from various Tetranychus urticae-infested plant species. Bulletin of Entomoligical Research 92: 539-546 Van Poecke RMP, Posthumus MA, Dicke M (2001) Herbivore-induced volatile production by Arabidopsis thaliana leads to attraction of the parasitoid Cotesia rubecula: chemical, behavioral, and gene-expression analysis. Journal of Chemical Ecology 27: 1911-1928 Wagner GJ (1991) Secreting glandular trichomes: More than just hairs. Plant Physiology 96: 675-679 Werker E (2000) Trichome diversity and development. Advances in Botanical Research 31: 1-35 Zhang PJ, Zheng SJ, Van Loon JJA, Boland W, David A et al. (2009) Whiteflies interfere with indirect plant defense against spider mites in Lima bean. Proceedings of the National Academy of Sciences USA 106: 21202-21207 http://faostat.fao.org/ 54