Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Transmission (medicine) wikipedia , lookup
Introduction to viruses wikipedia , lookup
Sociality and disease transmission wikipedia , lookup
Bacterial cell structure wikipedia , lookup
Gastroenteritis wikipedia , lookup
Infection control wikipedia , lookup
Anaerobic infection wikipedia , lookup
Marine microorganism wikipedia , lookup
Traveler's diarrhea wikipedia , lookup
Human microbiota wikipedia , lookup
Urinary tract infection wikipedia , lookup
Neonatal infection wikipedia , lookup
Triclocarban wikipedia , lookup
Carbapenem-resistant enterobacteriaceae wikipedia , lookup
Bacterial morphological plasticity wikipedia , lookup
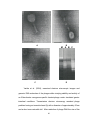
REVIEW OF LITERATURE 17 REVIEW OF LITERATURE 2.1 BACTERIOPHAGE THERAPY Prior to the discovery and widespread use of antibiotics, it was suggested that bacterial infections could be prevented and/or treated by the administration of bacteriophages (viruses attacking bacteria). Although the early clinical studies with bacteriophages were not rigorously pursued in the United States and Western Europe, phages continued to be utilized in the former Soviet Union and Eastern Europe. Ernest Hankin, a British bacteriologist, reported in 1896 on the presence of marked antibacterial activity (against Vibrio cholerae) which he observed in the waters of the Ganges and Jamuna rivers in India, and he suggested that an unidentified substance (which passed through fine porcelain filters and was heat labile) was responsible for this phenomenon and for limiting the spread of cholera epidemics. Two years later, the Russian bacteriologist Gamaleya observed a similar phenomenon while working with Bacillus subtilis, and the observations of several other investigators are also thought to have been related to the bacteriophage phenomenon. However, non of these investigators further explored their findings until Frederick Twort, a medically trained bacteriologist from England and Felix d’Herella, a French Canadian bacteriologist reintroduced the subject almost 20 years after Hankin’s observation by reporting a similar phenomenon and advancing the hypothesis that it may have been due to a virus (Sulakvelidze et al., 2001). 18 Biomedical technology today is very different from what it was in the early days of phage therapy research, and our understanding of biological properties of phages and the basic mechanisms of phage-bacterial host interaction has improved dramatically since the days of early therapeutic uses of bacteriophages (Sulkvelidze and Kutter, 2004). The concept of phage therapy to treat bacterial infections was born with the discovery of the bacteriophage almost a century ago. After a chequired history, its current renaissance is fueled by the dangerous appearance of antibiotic-resistant bacteria on a global scale. As a mark of this renewed interest, the unanswered problems of phage therapy are now being addressed, especially for human use (Duckworth and Gulig, 2002). 2.1.1 Development From the time Felix d’Herelle discovered bacteriophage he was interested in their relationship agents. He systematically investigated the nature of bacteriophages and explored their ability to function as therapeutic agents d’Herellel (1917). The studies on treatment of bacterial dysentery by using phages were conducted by Felix d’Herelle at Paris hospital in the summers of 1919 (Summers, 1999). The major milestones in the development of phage therapy and important studies on human phage therapy conducted by various researchers throughout the world are recorded in brief as under. Spence and McKinley (1924), Bacterial dysentery: Phages were administrated orally to 20 patients infected with Shiga (9 patients) and Flexner (11 patients) bacillus. Twelve patients not treated with bacteriophage, but 19 treated with the conventional methods, served as control group during the study. 30% less mortality was reported in phage-treated patients compared to phage-untreated patients. d’Herelle (1925), Plague: 4 plague patients recovered dramatically after injecting bacteriophages into their bubos. This is the first published report of the use of phage therapy against plague. The success outcome was largely responsible for the subsequent initiation of large-scale studies of the efficacy of phages against plague and cholera in India. d’Herelle et al. (1928), Enteric infections: Orogastric administration of phages dramatically reduced the cholera-associated fatality rate from 27-30% to Zero, in the Campbell hospital in Calcutta. During field trials, the prophylactic/therapeutic administration of phages to 74 patients in Punjabi villages reduced the mortality rate to 8%, compared to 63% among the 124 patients not treated with phages. Burnet et al. (1930), Dysentery: Anti-dysentery phages were used for prophylaxis/treatment of dysentery. Babalova et al. (1968), Dysentery: 17,044 children were treated with phage preparations vx. 13,725 children in control groups. Based on clinical diagnosis, the incidence of dysentery was 3.8–fold less than that occurring in the control, phage untreated group. Isoliani et al. (1980), Lung infections: Phages were successfully used, together with antibiotics, to treat lung and pleural infections in patients. 20 45 Meladze et al. (1982), Lung infections: Phages were used to treat 223 patients with infections of the lung parenchyma and pleura, and the results were compared to those of 117 patients treated with antibiotics. Full recovery was observed in 82% of the patients in the phage-treated group, as opposed to 64% of the patients in the antibiotic treated group. El-Tahan et al. (1983), Burnt infections: Reported the successful bacteriophage therapy in burn wound sepsis caused by P. aeruginosa and the recovery in almost all the patients was observed. Martynova et al. (1984), Opportunistic infections: S. aureus and P. aeruginosa specific phages were used prophylactically (in forms of mouth rinses) 2 times/day for 3-5 days in 27 patients. Control group consisted of 10 healthy individuals. Phage treatment was associated with normalization of microflora in infected sites, and it also stimulated the production of secretory immunoglobulins (IgA in particular). Cislo et al. (1987), Skin Ulcers: 31 patients with chronically infected skin ulcers were treated orally and locally with phages. The success rate was 74%. Minor side effects (eczema, pain, or vomiting) were observed in 6 patients. Kochetkova et al. (1989), Post-operative infections: 131 cancer patients with post-surgical wound infections participated in the study. Of these, 65 patients received phages, alone or in combination with antibiotics, and the rest received antibiotics. Phage treatment was successful in 82% of the cases, and antibiotic treatment was successful in 61% of the cases. 21 Abul-Hassan et al. (1990), Burnt infections: Phage therapy was used to control P. aeruginosa infection in 30 cases of resistant infections in patients with burn areas ranging from 10-25%. Post-phage cultures showed absence of organisms in 12 patients and their presence in 18. Satisfactory graft take was present in 18 cases only and inferior take in 12 cases. Sakandelidze (1991), Infectious allergoses: 1,380 patients with infectious allergoses were treated with phages (360 patients), antibiotics (404 patients), or a combination of phages and antibiotics (576 patients). Clinical improvement was observed in 86, 48 and 83% of the cases, respectively. Miliutina and Vorotyntseva (1993), Salmonellosis: The efficacy of treating salmonellosis using phages and a combination of phages and antibiotics was examined. The combination of phages and antibiotics was reported to be effective in treating cases where antibiotics alone were ineffective. Kwarcinski et al. (1994), E. coli infections: Recurrent subphrenic abscess (After stomach resection) caused by an antibiotic resistant strain of E. coli was successfully treated with phages. Perepanova et al. (1995), Urogenital infections: Adapted phages were used to treat acute and chronic urogenital inflammation in 46 patients. The efficacy of phage treatment was 92% (marked clinical improvements) and 84% (bacteriological clearance). Weber-Dabrowska et al. (2001), Cancer: 20 Cancer patients refractory to treatment with commonly available antibiotics were treated by oral 22 administration of specific phages (3 doses/day, for 2 to 9 weeks). Some patients were also treated by local administration of phages. Complete healing of the local lesions and termination of the supurative process was reported for all phage-treated patients. Side effects were not observed in any of the patients subjected to phage treatment. Markoishvili et al. (2002), Wound infections: The wounds healed completely in 67 (70%) of 96 patients whose wounds were covered with a phage-containing biodegradable matrix. In 22 cases in which microbiological data were available, healing was associated with the concomitant elimination of, or a reduction in, specific pathogenic bacteria in the wounds. Paisano et al. (2004), Dental Infection: In vitro antimicrobial effect of bacteriophages on human infected dentin was studied. Results revealed high efficacy of bacteriophages on human dentin infected with Enterococcus faecalis ATCC 29219. 2.1.2 Prospects Infectious disease experts have warned that there is now a compelling need to develop totally new classes of antibacterial agents, ones that cannot be resisted by the same genes that render bacteria resistant to antibiotics. Phage therapy represents such a new class. The impediments like bacterial debris in the preparations, rapid clearance in the body, etc. can be overcome, frezing up the phages so that their attributes such as, exponential growth, and the ability to mutate against resistant bacteria can be used to great advantage. 23 The three additional attributes of phages as their future prospects according to Carlton (1999) are as mentioned below. Host specificity: While the host specificity is somewhat of a drawback (requiring a match up of phage to bacterial target, and/or the development of highly multivalent phages), it also offers the great advantage that the phages will not kill other species of bacteria. Thus, e.g., phage therapy is not likely to kill off the healthy flora of the intestines, lungs or urogential tract, and it is therefore unlikely to provoke the illnesses and deaths seen when antibiotics cause overgrowth of pathogens (such as Clostridia difficile and Candida albincans). Genetic engineering: It is possible to genetically engineer phages to express new traits of potential value. In so doing, scientists will have to deal with the legitimate concerns of regulatory agencies concerning recombinant organisms. The regulatory obstacles may be well worth the price, given the powerful engineering tools that are currently available. Ideal candidates for co-therapy with antibiotics: If a given bacterium acquires resistance to a phage (e.g., by a mutation in the receptor site or in the endonucelase enzymes), the mutation is likely to “teach” the bacterium to resist the antibiotics (which do not target those structures). Similarly, if a given bacterium acquires resistance to an antibiotic (e.g., by a mutation in the reflux pump or in the ribosomal subunits), that mutation is not likely to “teach” the bacterium to resist the phage (which does not target those structures). Thus, if the bacterium is exposed to both agents, the odds are remote that 24 any resistance genes it starts to express (or acquires anew) will enable it to survive. There are reports that bacteria tend to mutate against antibiotics once in every 106 divisions, while they tend to mutate against phages once in every 107 divisions. Therefore the odds of a given bacterium mutating against a phage and an antibiotic at the same time would be the product of 106x107, meaning it would likely take 1013 bacterial divisions for such a double mutation to occur. Given that low probability, the co-administration of phages and antibiotics may help prevent the emergence of bacterial resistance to antibiotics, thereby greatly prolonging their clinical usefulness (and vice versa). Just as multiple classes of anti-HIV medications are administered to AIDS patients, to prevent the emergence of resistant strains of that virus, so it is that co-therapy with phages and antibiotics may also prove to be of great clinical value. From a clinical standpoint, phages appear to be very safe. This is not surprising, that humans are exposed to phages from birth (and, possibly, even in utero). The abundance of phages in the environment – and the continuous exposure of humans to them – explains the extremely good tolerance of the human organisms to phages. These observations strongly suggest that phage therapy may provide one of the safest as well as most environmentally friendly methods currently available for prophylaxis and treatment of bacterial infections. However, in order not to compromise the safe use of therapeutic phage preparations, they should be produced, purified, characterized, and tested using current, state of the art biotechnology. This approach will help ensure the consistency and safety of therapeutic phage 25 preparations, and it is also likely to increase their therapeutic and prophylactic potential. The important advantages of using phages over antibiotics attributing to the major prospects of phage therapy according to Ahmad (2002) are as follows. Phages Antibiotics Highly specific to the targeted infection Less specific hence all sensitive bacteria including normal flora destroyed Application intervals may be as long as Required once every 7 days usually at short intervals (multiple applications per day) Once applied it can continue working as Have long as the host (infection) persists a limited life hence regular administration required Grow and destroy at the site of infections Usually metabolized and distributed all hence available where most needed over the body Exist for virtually all known bacteria Some bacterial species are notoriously resistant to several antibiotics Can be dispersed in any media Some may be insoluble in water Low production cost Relatively more expensive Exponential growth leading to continuous No such increase fight against infection Natural abundance Limited supply Multiple resistance highly unlikely More common due to MDR plasmids Minimal resistance (no plasmid-borne Can be high due to plasmid-borne resistance is known) resistance Shelf life can be long if stored appropriately Some have short shelf life No disruption to normal flora Disturbance to normal flora leading to side effects such as secondary infection by yeast Side effects including serious reactions No serious side effects known known Phage-resistance bacteria remain sensitive Resistance is not limited to target bacteria to other phages with different target sites 26 2.1.3 Limitations The discovery of the bacteriophage or phage is somewhat controversial. The consensus is that 20 years after initial observations of unfilterable, heat labile agents with activity against – Vibrio cholerae (Ernest, 1986), it was discovered by Twort in 1915, who made similar observations and who hypothesized this to be due to a virus, and independently by d’Herelle in 1917 (d’Herelle, 1917; Topley and Wilson, 1936). Since it was realized that these bacterial viruses destroy their bacterial host while remaining harmless to humans, it has been the dream of researchers to use phages to treat bacterial infections. One of the most important factors that contributed to the decline of interest in phage therapy in the western world was a credibility problem. A paucity of appropriately conducted studies and the lack of well-established and standardized testing protocols interfered with rigorously documenting the value of phage therapy. In addition, several companies started producing phages commercially and, in their eagerness to boost profits, some made exaggerated claims concerning the effectiveness of their products. Many productions – related problems also complicated early phage therapy research. Before the availability of spray-dryers, freeze-dryers, refrigerators and freezers, the viability of phage preparations was a concern; phage titers could rapidly decline and render the preparations ineffective. Thus, several “stabilizers” and “preservatives” were used in an attempt to increase the viability of phages. However, in the absence of a good understanding of the biological nature of phages and their stability to various physical and chemical agents, many of the ingredients added to prolong 27 phage shelf-life (e.g., phenol, which was included in some of the early therapeutic phage preparations) actually had a detrimental effect on phage viability and were toxic for humans. 2.2 PSEUDOMONAS AERUGINOSA Pseudomonas aeruginosa was first detected by Gessard in 1982 from a blue pus. Until recently, only P. aeruginosa is considered as important human pathogen, from among numerous species of Pseudomonads (Greenwood, 1998). P. aeruginosa is an aerobic, motile, Gram –ve rod. The production of colourful water-soluble pigments, particularly bluish green pycocynanin and yellowish green pyoverdin are the outstanding bacteriological features of this human pathogen. It is the epitome of an opportunistic pathogen of humans and also demonstrates the most consistent resistance to antimicrobials of all the human pathogens (Ahmad, 2002). It is being considered as dangerous and dreaded human pathogen because of its notorious resistance to wide range of antibiotics. 2.2.1 Pathogenicity P. aeruginosa is an opportunistic human pathogen, it is one of particular virulence. The pathogen generally requires a significant break in first line defenses (such as wound) or a route past them (such as a contaminated solution or intratracheal tube) to initiate infection (Kenneth and Stanley, 2002). The major infections of P. aeruginosa are both invasive and toxigenic. However, the ultimate infection may be seen under three distinct phases: colonization, invasion and dissemination. The disease process may stop at any stage. Particular bacterial determinants of virulence mediate each 28 of these stages and are ultimately responsible for the clinical symptoms that accompany the disease. Colonization: The fimbriae of Pseudomonas will adhere to the epithelial cells of the upper respiratory tract and, by inference, to other epithelial cells as well. These adhesions appear to bind to specific galactose or mannose or sialic acid receptors on epithelial cells. Colonization of the respiratory tract by Pseudomonas requires fimbrial adherence and may be aided by production of a protease enzyme that degrades fibronectin in order to expose the underlying fibrial receptors on the epithelial cell surface. Tissue injury may also play a role in colonization of the respiratory tract since P. aeruginosa will adhere to tracheal epithelial cells of mice infected with influenza virus but not to normal tracheal epithelium. Besides pili and the mucoid polysaccharide, there are possibly two other cell surface adhesins utilized by Pseudomonas to colonize the respiratory epithelium or mucin. Invasion: The ability of P. aeruginosa to invade tissue depends upon its resistance to phagocytosis and the host immune defenses, and the extracellular enzymes and toxins that break down physical barriers and otherwise contribute to bacterial invasion. The bacterial capsule or slim layer effectively protects cells from opsonization by antibodies, complement deposition, and phagocyte engulfment. Two extracellular proteases have been associated with virulence that exerts their activity at the invasive stage. Dissemination: Blood stream invasion and dissemination of Pseudomonas from local sites of infection is probably mediated by the same cell associated 29 and extracellular products responsible for the localized disease, although it is not entirely clear how the bacterium produces systemic illness. P. aeruginosa is resistant to phagocytosis and the serum bactericidal response due to its mucoid capsule and possibly lipo-polysaccharides. The lipid A moiety of Pseudomonas mediates lipo-polysaccharides (endotoxin) pathologic aspects of Gram-negative septicemia. the usual It is also reasonable to assume that Pseudomonas Exotoxin A exerts some pathologic activity during the dissemination stage. P. aeruginosa produces two extracellular protein toxins, Exoenzyme S and Exotoxin A. Exoenzyme S is probably an exotoxin. It has the characteristic subunit structure of the A-component of a bacterial toxin, and it has Adinosine di-phosphate ribosylating activity (for a variety of eukaryotic proteins) characteristic of exotoxins. Exoenzyme S is produced by bacteria growing in burned tissue and may be detected in the blood. It has been suggested that exoenzyme S may act to impair the function of phagocytic cells in the bloodstream and internal organs to prepare for invasion by P. aeruginosa. 2.2 .2 Pathogenesis P. aeruginosa is a physiologically versatile organism, widely distributed in soil, water, sewage, the mammalian gut and plants. P. aeruginosa is susceptible to drying but will survive for many months in water at ambient temperature. Simple nutritional requirements and the ability to metabolize a variety of organic substances may enable P. aeruginosa to survive and multiply in fluids and moist environments found in hospital wards. Infection 30 with P. aeruginosa is generally restricted to hospitalized patients. Predisposing factors for nosocomial infections include metabolic, hematologic, and malignant diseases. Hospital-acquired infections occur in patients who have had prior instrumentation or manipulative procedures such as urethral catheterizations, tracheotomies, lumbar punctures, and intravenous infusions of medications and fluids. Patients become susceptible to P.aeruginosa infections after prolonged treatment with immunosuppressive agents, corticosteroids, antimetabolities, antibiotics, and radiation. Hospitalized patients may acquire the bacteria from common environmental sources by contact with human or inanimate vectors. The major infections caused by P. aeruginosa are as follows (Keneeth, 2002). Respiratory infections: Respiratory infections caused by P. aeruginosa occur almost exclusively in individuals with a compromised lower respiratory tract or a compromised systemic defense mechanism. Primary pneumonia occurs in patients with chronic lung disease and congestive heart failure. Bacteremic pneumonia commonly occurs in neutropenic cancer patients undergoing chemotherapy. Lower respiratory tract colonization of cystic fibrosis patients by mucoid strains of P. aeruginosa is common and difficult, if not impossible to treat. Urinary tract infections: Urinary tract infections (UTI) caused by P. aeurginosa are usually hospital-acquired and related to urinary tract catheterization, instrumentation or surgery. P. aeruginosa is the third leading cause of hospital-acquired UTIs, accounting for about 12 percent of all 31 infections of this type. The bacterium appears to be among the most adherent of common urinary pathogens to the bladder uroepithelium. The urinary tract infection can occur via an ascending or descending route. In addition, Pseudomonas can invade the bloodstream from the urinary tract, and this is the source of nearly 40 percent of Pseudomonas bacteremias. Wound and burnt infections: P. aeruginosa can cause a variety of skin infections, both localized and diffuse. The common predisposing factors are breakdown of the integument which may result from burns, trauma or dermatitis; high moisture conditions such as those found in the ear of swimmers and the toe webs of athletes and combat troops, in the perineal region and under diapers of infants, and on the skin of whirlpool and hot tub users; neutropenia; and AIDS, Pseudomonas has also been implicated in folliculitis and unmanageable forms of acne vulgaris. Ear infections including external otitis: P. aeruginosa is the predominant bacterial pathogen in some cases of external otitis including “swimmer’s ear”. The bacterium is infrequently found in the normal ear, but often inhabits the external auditory canal in association with injury, maceration, inflammation, or simply wet and humid conditions. Bacteremia: P. aeruginosa causes bacteremia primarily in immunocompromised patients. Predisposing conditions include hematologic malignancies, immunodeficiency relating to AIDS, neutropenia, diabetes mellitus and severe burns. Most Pseudomonas bacteremia is acquired in hospitals and nursing homes. Pseudomonas accounts for about 25 percent of all hospital acquired Gram–negative bacteremias. 32 Gastrointestinal infections: P. aeruginosa can produce disease in any part of the gastrointestinal tract from the oropharynx to the rectum. As in other forms of Pseudomonas disease, those involving the gastrointestinal tract occur primarily in immunocompromised individuals. The organism has been implicated in perirectal infections, pediatric diarrhea, typical gastroenteritis, and necrotizing enterocolitis. The GI tract is also an important portal of entry in Pseudomonas septicemia. 2.2.3 Drug resistance Clinically significant infections with P. aeruginosa should not be treated with single drug therapy, because the success rate is low with such therapy and because the bacteria can rapidly develop resistance when single drugs are employed. A penicillin active against P. aeruginosa- ticarcillin, mezlocillin, and piperacillin – is used in combination with an aminoglycoside, usually gentamicin, tobramycin, or amikacin. Other drugs active against P.aeruginosa include aztreonam, imipenem and the newer quinolones, including ciprofloxacin. Of the newer quinolones, including ciprofloxacin, ceftazidime and cefoperazone are active against P. aeruginosa; ceftazidime is used in primary therapy of P. aeruginosa infections (Norrby et al., 1993). To a considerable extent, the intrinsic resistance of P. aeruginosa is due to outer membrane porins that restrict the entry of antimicrobials to the periplasmic space. The bacterium is naturally resistant to many antibiotics due to the permeability lipopolysaccharide. barrier afforded by its outer membrane Since its natural habitat is the soil, living in association with the bacilli, actinomycetes and molds, it has developed resistance to a 33 variety of naturally occurring antibiotics. Moreover, Pseudomonas maintains antibiotic resistance plasmids and it is able to transfer these genes by means of transduction and conjugation (Farjadian et al., 1996 and Kenneth, 2002). P. aeruginosa strains are regularly resistant to pencillin, ampicillin, cephalothin, tetracycline, chloramphenicol, sulfonamides and the earlier aminoglycoisdes (streptomycin, kanamycin). Much effort has been directed towards the development of antimicrobic with anti-Pseudomonas activity. The aminoglycosides, gentamicin, tobramycin, and amikacin, are all active against most strains despite the presence of mutational and plasmid mediated resistance. Carbenicillin and ticarcillin are active and can be given in high doses, but plasmid-mediated resistance and permeability mutations occur more frequently than with the aminoglycoides. A primary feature of the third generation cephalosporins (ceftazidime), carbapenems (imipenem), or monobactams (azthreonam) is their activity against Pseudomonas. In general, more serious systemic P. aeruginosa infections are usually treated with a combination of an anti-Pseudomonas beta-lactam antimicrobic and an aminoglycoside, particularly in neutropenic patients. Ciprofloxacin is also used in treatment of such cases. In all instances susceptibility must be confirmed by in vitro susceptibility tests. 2.3 BACTERIOPHAGE Bacterial viruses or bacteriophages, frequently called “phages”, were discovered twice at the beginning of the 20th century. Frederick William Twort, a British pathologist in London, described in 1915 the glassy transformation of “Micrococcus” colonies by a transmissible agent. 34 He proposed several explanations, one of which was that the agent was viral in nature. Felix Hubert d’Herelle, a French Canadian then working at the Pasture Institute of Paris, observed the lysis of Shigella cultures in broth and described it in 1917. Twort did not pursue his discovery, but attempted for decades to propagate vertebrate viruses on inert media. d’Herelle, on the contrary, clearly recognized the viral nature of his agent and devoted the rest of his scientific life to it. He coined the term “Bacteriophage”, devised several techniques still in use, postulated the intracellular multiplication of viruses, and introduced phage therapy of infectious diseases (d’Herelle, 1917). The viral nature of bacteriophages, contested for a time, was definitely recognized in 1940 after the advent of the electron microscope (Pfankuch and Kausche, 1940). 2.3.1 Biology The study of phages begins with the work of Max Delbruck in the late 1930’s, Nobel Laureate Delbruck, originally a physicist, began studying phages as genetic and biochemical experimental systems. This work led others to focus their studies on the structure biology and assembly of phages. Life cycle: Bacteriophages undergo two possible life cycles. These are the lytic (or virulent) and lysogenic. Lytic phage multiply vegetatively and kill the host cell at the end of the growth cycle. Temperate phages which undergo the lysogenic cycle as well as multiplying vegetatively can also persist in a lysogenic state, whereby the phage genotype can exist indefinitely by being inserted in the bacterial chromosome (known as the prophage state) (Karl Thiel, 2004). 35 Phages undergoing lytic cycles only are virulent. Lytic cycles consist of several steps and show considerable variations according to the type of phage. The general diagrammatic representation of the lytic cycle is as shown under. Phages encounter bacteria by chance and adsorb to specific receptors, generally located on the cell wall, but also on flagella, pilli, capsules, or the plasma membrane. The phage nucleic acid enters the host and the shell remains outside. In phages with contractile tails, the cell wall is degraded by phage enzymes located on the tail tip. The sheath then contracts and the tail core is brought in contact with the plasma membrane. It depends largely on the physiological state of the host and varies between 20 min and 30–40 hrs. Phage nucleic acid is transcribed to mRNA using host and/or phage RNA polymerases. The assembly of new phages is called maturation. Phage constituents assemble spontaneously or with the help of specific enzymes (Ackermann and DuBow, 1987). 36 The lysogenic cycle on the other hand comprises replication of phage, nucleic acid together with the host genes for several generations with major metabolic consequences for the cell. This is a latent mode of infection and it occurs at a very low frequency. The phage gene in this state may occasionally revert to lytic cycle. Leading to release prophages particles. This property is known as lysogens and phage that can develop both lytically and lysogenically are said to be phages (Beneett and Howe, 1998). 2.3.2 Structure and Classification Structure: All phages have single, linear double stranded DNA (dsDNA) chromosomes stored in a protein coat shell called the capsid or head. The capsid is built by protein molecules along icosahedrally symmetric arrays to form the distinctive shape of phages. The tail extends from one corner of the capsid and interacts with a single host cell. During infection, the distal end of the tail adsorbs to the exterior of the host cell as phage DNA travels through the tail into the cells (Ackermann and DuBow, 1989). 37 Antje et al. (1998) assigned the phages to different virus families, species and strains based on morphology, DNA homology and host range. The phenotypic diversity of the 22 bacteriophages was examined by electron microscopy and phages were identified by following morphological criteria out lined as per the International Committee of Taxonomy of Viruses. Morphological studies of 22 phages detected by them revealed that all of the phages had tails and thus belong to the order Caudovirales. The icosahedral heads of the phages had diameters between 50.2 and 99.3 nm. The phages could be assigned to three virus families. Eleven of the phages belonged to the family Myoviridae, which contains phages that have icosahedral heads and long contractile tails; seven phages were assigned to the family Siphoviridae which contains phages that have icosahedral heads and long flexible tails and four phages, which had icosahedral heads and short tails, belonged to the family Podoviridae. Phages belonging to 3 different families and their micrographs illustrated by them are as under. A. Myoviridae B. Siphoviridae 38 C. Podoviridae Electron microscopic images of the somatic coli phages (faecal coli forms) isolated from environmental samples by Duran et al. (2002) were as shown below. The structure (A) of bacteriophage with isometric head and short tail indicates Myoviridae and another structure (B) of phage with long straight flexible tail indicate the family Siphoviridae. A. Myoviridae B. Siphoviridae Sandra et al. (2004) tested stool samples from pediatric diarrhoea patients and environment water samples in Dhaka, Bangladesh and sewage from Switzerland, for E. coli bacteriophages, yielded nearly exclusively phages with a contractile tail belonging to family Myoviridae. The electron microscopic structure and genomic DNA of the A, B and C are main three selected phages based on their potential lytic activity on pathogenic E. coli were as shown under. All three phages showed the typical morphology of T4 like phages with 170 kb genome (D) upon pulsed field electrophoresis. 39 A B M C A B C D Verthe et al. (2004), examined electron microscopic images and genomic DNA molecules of the phages while studying stability and activity of an Enterobacter aerogenes–specific bacteriophage under simulated gastrointestinal conditions. Transmission electron microscopy revealed phage particles having an isometric head (A) with a diameter of approximately 65 nm and a short non-contractile tail. After restriction of phage DNA the size of the 40 major bands (B) were approximately 23,000 bp, 7000 bp and 4000 bp. These morphological properties and the estimated genome size of at least 34 kb correspond to the T7 like phages of the genus Podoviridae, family Podoviridea and order Caudovirales. A B Classification: The first global attempt to systematically classify viruses took place at the International Congress of Microbiology held in Moscow in 1966. This meeting established the International Committee of Taxonomy of Viruses (ICTV), whose mission was to develop a universal taxonomic system for all viruses infecting animals, plants, fungi, bacteria, and later, archaea. Beginning with its first report in 1971, the International Committee of Taxonomy of Viruses has met regularly to update virus definitions and taxonomy guidelines, with the seventh and most recent report published in 2000 (Regenmortel, et al., 2000). Viruses are grouped together by shared characteristics, with subgroups having smaller clusters of shared attributes. For example, the tailed order of phage (Caudovirales) is broken down into 41 three families; phage with long, contractile tails (Myoviridae), phage with long, noncontractile tails (Siphoviridae), and phage with short tails (Podoviridae). The families are further broken down into genus and subgenus by criteria such as genome configuration (linear, circular, super coiled), host range, and genome size. The inadequacy of the International Committee of Taxonomy of Viruses classification is evident when one looks closely at the numbers. Of the completed phage genomes currently deposited in GenBank (Benson, et al., 2004), 40% (92 of 228) are unclassified beyond the level of family according to International Committee of Taxonomy of Viruses conventions. Furthermore, an additional 10% (23 of 228) are not even assigned an order and are simply listed as “unclassified bacteriophage”. It is generally agreed that future phage classifications must reflect genomic data as a primary component. In bacteria, this is easily accomplished by examining the conserved 16S ribosomal genes. However, phage lack ribosomal DNA and there are no conserved gene or protein sequences common to all phage on which to base a classification (Schuch et al., 2002). Furthermore, any immediate attempt to reclassify phage based exclusively on comparative genomics may be biased toward the lambdoid phages or those involved in industrial fermentation, since these phages dominate the current assemblage of sequenced genomes. Nonetheless, in the fall of 2002, three separate phage research groups proposed alternative classification schemes as outlined below. 42 An option proposed by Rohwer and Edwards is based on a “Phage proteome tree”, which is constructed by grouping phage both relative to their rear neighbours and in the context of all other phage (Rohwer and Edwards, 2002). This method analyzed the entire predicted proteome for a given phage and the results were then transformed into a distance matrix. A tree was constructed based on relationships between phage proteins. The author list several anomalies in the International Committee of Taxonomy of Viruses system that could be resolved by the proteome tree. Additionally, the proteome tree moves, which International Committee of Taxonomy of Viruses classifies as Tectiviridae due to the presence of a lipid membrane below the capsid, to a subgenus of the family Podoviridae. A second approach, put forth by a group at the Pittsburgh Bacteriophage Institute, suggests that it may be impossible to have a strictly hierarchical taxonomic system given the genetic mosaicism arising from horizontal gene transfer among phage (Lawarence et al., 2002). In this paradigm, the top levels of taxonomy would still follow the hierarchical Linnean approach, i.e., viruses would be divided into “domains” according to genome type (double stranded DNA, single stranded DNA, Single–stranded RNA, and double–Stranded RNA), with a further partition known as “divisions” to separate defining characteristics such as tailed phage from filamentous phage. However, from this point on, three basic tenets would guide the remainder of the classification. First, members of a group should exhibit similarity in one or more loosely defined cohension mechanisms. Second, significant sequence data, preferably from whole genomes, should be 43 available for evolutionary assignment to a taxonomic cluster. Third, the groups may be reticulate, i.e., phage may simultaneously belong to several groups based on multiple and/or differing criteria from the first two tenets. This web like design has an inherent flexibility that is not afforded by either phenetic or genetic hierarchical approaches. Finally, a third classification scheme, based on comparative genomics of a structural gene module, has been proposed (Proux et al., 2002). Since it is believed that the structural genes are the oldest and most conserved module in dairy phage, dot plots of temperate lactococcal phage were used to observe graded relatedness between DNA and protein sequences as well as similarity in the organization of the structural genes in the absence of sequence relatedness. Whereas, comparative genomics of non-structural genes tended to lump all of the lactococcal phages studied into one species, comparative genomics of the structural genes delineated four phage species and distinguished two genera based on head morphology. It may not be such a bad idea that we agree to disagree (Daniel, 2004) with the exception of a few remarkable outliners, some of which are noted above. 2.3.3 Sources and Distribution Bacteriophages or “Phages” are viruses of prokaryotes including eubacteria and archaebacteria. They were discovered and described twice, first in 1915 by the British pathologist Frederick William Twort and then in 1917 by the Canadian bacteriologist Felix Hubert d’Herella working at the Pasteur Institute of Paris. With about 3500 isolates of known morphology, phages constitute the largest of all virus groups. Phages are tailed, cubic, 44 filamentous, or pleomorphic. Tailed phages are far more numerous than other types, are enormously diversified, and must be very old in geological terms (Richard Sharp, 2001). Phages have been found in over 100 bacterial genera distributed all over the bacterial world; in aerobes and anaerobes, actinomycetes, archaebacteria, cyanobacteria and other phototrophs, endospore formers, appendaged, budding, gliding, mycoplasmas and chlamydias. and sheathed bacteria, spirochetes, Phage like particles of the podovirus type have even been found in endosymbionts of paramecia. However, tailed phages reported in cultures of green algae and filamentous fungi are probably contaminants. Most phages have been found in a few bacterial groups: enterobacteria (over 650 phages), bacilli, pseudomonads, staphylococci, and streptococci. clostridia, lactococci, This largely reflects the availability and ease of cultivation of these bacteria and the amount of work invested. About half of phages have been found in cultures of lysogenic bacteria. Tailed phages predominate everywhere except in mycoplasmas. In archaebacteria, they have been found in the genus Halobacterium only and not yet in methonotrophs and extreme thermophiles. Siphoviridae are particularly streptococci. frequent in Myoviruses enterobacteria, actinomycetes, and pseudomonads, coryneforms, podoviruses bacilli, and are lactococcis relatively clostridia. frequent This and in particular distribution must have phylogeneic reasons. Except for phages from extreme environments, phage species generally seem to be distributed over the whole earth. This is suggested by 45 electron microscopic observations of rare and characteristical phage morphotypes in different countries and global occurrence of certain lactococcal phage species in dairy plants and of RNA coliphages in sewage. Unfortunately, most data are from developed countries. Sizes of phage populations are difficult to estimate because plaque assays and enrichment and (most) concentration techniques depend on bacterial host; they therefore only detect phages for specific bacteria and environmental conditions. Consequently, phage titers vary considerably – for example, for coliphages 109/g in human feces, 107/ml in domestic sewage and 105/g of actinophages in soil. Several sources have been used by different researchers for the isolation of bacteriophages and to understand their prevalence. Etwert (1980) studied the distribution of phages in raw sewage and treated effluents. Bitton (1987) also studied the distribution of phages raw domestic waste water. Antije et al. (1998) and Yoon (1999) reported distribution or phages in sea water and marine environment respectively. Richard Sharp (2001) recorded the distribution of phages in sea water, sewage and also soil. Keven et al. (2003) and Ghanaat et al. (2003) were also recorded the prevalence of bacteriophages in different soil and sewage. Bachrach et al. (2003) have isolated bacteriophages from human saliva. Phages for Enterococcus faecalis were found in saliva samples. The presence and stability of the E. faecalis bacteriophages in human saliva suggests a possible role of these bacteriophages in a oral ecosystem and phage therapy as a way to control oral bacteria might be considered. 46 Hitch et al. (2004) have isolated bacteriophages from the oral cavity. The composition of the oral cavity does not appear to be heavily influenced by interactions between bacteriophages and their hosts and reported bacteriophage for control of oral infections may need to be obtained from other sources. 2.4 IN VITRO ACTIVITY OF BACTERIOPHAGES Smith and Huggins (1982) while studying efficacy of phages against pathogenic E. coli, isolated 15 phages from specimens of sewage, out of which 9 were identified as specific phages based on the degree of the lytic activity (diameter of plaques). The 9 anti E. coli phages were much more virulent than the others based on in vitro lytic activity. 106–109 viable particles of the phages were required to lyse broth of E. coli seeded with 3x108 viable organisms. 50–120 resistant colonies were also observed within the zone produced by 1 drop of undiluted preparations of few anti E. coli phages spotted on a lawn of pathogenic E. coli, determining the difference in degree of in vitro lytic activity of phages. Prior observations of phage host systems in vitro have led to the conclusion that, susceptible host cell population must reach a critical density before phage replication can occur. Such a replication threshold or “proliferation threshold” density would have broad implications for the therapeutic use of phage. Kasman et al. (2002) were demonstrated experimentally that, such replication threshold exists and support the threshold in terms of a classical model for the kinetics of colloidal particle interactions in solution. 47 An in vitro study was performed by Steven Hagens et al., (2004) to develop genetically stable, efficient safe therapeutic phage for the infections caused by P. aeruginosa. The use of technique minimized endotoxin release and hence significantly reducing the side effects. Flynn et al. (2004) have studied the exploitation of bacteriophage as biocontrol agents to eliminate the pathogen E. coli. Two distinct lytic phages isolated against a human strain of E. coli and a cocktail of phages were evaluated for their ability to lyse the bacterium in vivo and in vitro. However, bacteriophage – insensitive mutants emerged following the challenge and commonly reverted to phage sensitivity within 50 generations. Tanji et al. (2004) screened from various sources, 26 phages infected with E. coli. Among them 9 caused visible lysis of E. coli cells in Luria Bertani liquid medium. However, prolonged incubation of E. coli cells and phage allowed the emergence of phage resistant cells. A bacteriophage showing high in vitro lytic activity against a clinically important strain of Enterobacter aeruginosa was isolated from hospital sewage by Verthi et al. (2004). The stability and lytic activity of phage against the clinical isolate under simulated gastro–intestinal in vitro conditions was also evaluated. Sandra et al. (2004) studied in vitro bacteriolytic activities of E. coli phages to understand implications of phage therapy. 40 phages specific to pathogenic strain of E. coli were isolated from stool samples of pediatric diarrhea patients and environmental water samples in Dhaka, Bangladesh and sewage from Switzerland. 4 selected phages based on prominent plaque 48 features were rescreened for their lytic potential on the E. coli by a tube lysis test. This test is more labour intensive than the spot test but, offers a more rigorous assessment of bacterial lysis in vitro activities of the test phages. 2.5 FACTORS INFLUENCING PHAGE ACTIVITY Eric et al. (1996) studied effects of sunlight on the viability and structure of bacteriophage in marine aquatic systems. Destruction of virus particles is concluded to be a process separate from loss of infectivity. It is also concluded that strong sunlight affects the viability of bacteriophages in surface waters, with the result that direct counts of virus like particles over the estimate of bacteriophage capable of both infection and replication. However, in deeper waters, where solar radiation is not a significant factor, direct counts should more accurately estimate numbers of viable bacteriophages. Yoon et al (1999) were suggest that, a novel bacteriophage, designated as VPP97 which infects the of Vibrio para haemolytics (hallophilic, Gram-negative bacteria) isolated most commonly from marine environments. In this studies characterized phages were almost totally inactivated at 70oC and at pH below 5 or over 10, and the phage treatment appears effective to the infection by V. parahaemolytics. Feng et al. (2003) studied effects of pH and temperature in the survival of coli phages. MS2 phage survived better in acidic conditions than in an alkaline environment in contrast Q phage had a better survival rate in alkaline conditions; than in an acidic environment. The inactivation rates of both coli phages were lowest within the pH range 6-8 and the temperature range 5-350C. The inactivation rates of both coli phages increased when the 49 pH was decreased to below 6 or increased to above 8. The inactivation rates of both coli phages increased with increasing temperature. Substances or conditions that denature proteins or react chemically with proteins or nucleic acids will in activate phages. The inactivation of MS2 and Q observed in their study could be attributed to reactive radicals and levels of heat stability. Temperature has a major effect on the effectiveness and/or the rate of kill or a given microorganism because it controls the rate of chemical reactions. Thus, as temperature increases, the rate of kill induced by a chemical will also increase. In addition, pH can affect the ionization of chemicals. At extreme pH values, the high concentrations of hydrogen ion and hydroxyl ion present in water are considered to be far greater than the concentration of free reactive radicals and therefore dominate viral inactivation mechanism. Verthi et al. (2004) studied on effect of pH, bile salts and pancreatin on bacteriophage in simulated gastro-intestinal in vitro conditions. After 1 hour incubation at 34oC at different pH levels, there were no significant differences between the initial phage titer of 6.2 0.3x105 pfu/ml and the phage concentrations at pH 9, 7, 6 and 4. However, at pH2, the concentration of bacteriophage dropped below the detection limit (1.0x101 pfu/ml) immediately after the addition of phage. 2.6 IN VIVO EFFICACY OF BACTERIOPHAGE Smith and Huggins (1982) studied successful treatment of experimental E. coli infections in mice using phages and their general superiority over antibiotics. The minimum lethal dose (LD50) of pathogen E. coli was 3x107/ml and phage was 3x108/ml. 50 The phage administered intramuscularly 3-5 days before challenge with a potentially intramuscularly induced infection of MW was protective. lethal However they observed variations in protective effect of phages propagated on different pathogenic strains of E. coli. A single intramuscular dose of anti E. coli phage was more effective than multiple intramuscular doses of tetracycline, ampicillin, chloramphenicol, or trimethoprim plus sulfphfurozole in curing mice of a critically induced infection of pathogen E. coli. The few phage resistant mutants of E. coli were also observed during the study. According to them 3x103 to 3x104 phage particles were sufficient to cure mice given a potentially lethal dose of pathogenic E. coli. Soothill (1992) examined bacteriophage for P. aeruginosa in experimental infections of mice to investigate their potential for the treatment of infection of man. A Pseudomonas phage protected mice against LD50–(8x107 cfu/ml) of a virulent strain of a P. aeruginosa with a LD50 of 1.2x107 particles. This studies support the view that bacteriophages could be useful in the treatment of human infections caused by antibiotic resistant strains of bacteria. Gowri Sankar et al. (1998) evaluated phage therapy to treat experimental infections in mice due to a clinical isolate of P. aeruginosa, which was isolated from patients admitted into a rural hospital. Bacteriophage specific for this pathogen was isolated from a domestic sewage. Test animals (mice) were injected with the bacterial pathogen (0.1 ml suspension with 1.75x104 cells/ml) through the intraperitoneal route. Their phage patterns were also administered 0.5-1.0 ml suspension with 51 (5.5x104 pfu/ml). Subsequently through the same route after a time lap of 6 hours. Animals protected with phage ingestion exhibited no sign of illness when compared to the unprotected ones. This protection was attributed to the marked reduction (from 6.3x108/g to 4.0x105/g of viscera of mice) of the pathogen load in the tissues of the animals in response to the presence and persistence of phage. Karen et al. (2002) studied phage therapy of local and systemic disease caused by Vibrio vulnificus in mice. In this study, they examined the potential use of bacteriophages as therapeutic agents against V. vulnificus in an iron-dextran-treated mouse model of V. vulnificus infection. Mice were injected subcutaneously with 10 times the lethal dose V. vulnificus and injected intravenously, either simultaneously or at various times after infection, with phages. Treatment of mice with phages could prevent death; systemic disease, as measured by cfu/g of liver and body temperature; and local disease, as measured by cfu/g of lesion material and histopathologic analysis were carried out. Samples of the infected blood, liver and peritoneal lavage fluid were sampled at various intervals, after injection of the phage, and pfu/ml or g were enumerated. Data was expressed in means and standard deviations. Biswajit et al. (2002) reported that bacteriophage therapy rescues mice bacteremic from a clinical isolate of vancomycin resistant Enterococcus faecium. Ghanaat et al. (2003) have investigated the potential of phages for the treatment of P. aeruginosa infection in mice. A strain of P. aeruginosa was 52 used which was resistant to many or routinely used antibiotics. Bacteriophage was originally isolated from sewage by standard methods using mentioned strain of P. aeruginosa as host (Slopek et al., 1987). Pathogen free mice were chosen at 3rd wks. of age. First approximate lethal dose 50% of this strain of P. aeruginosa were surveyed (around 105 cfu). In their experiment, a much higher dose (107 cfu) could kill 100% of non-treated mice. Two groups of 10 mice (case and control) were inoculated intraperitoneally with 107 cfu of P. aeruginosa. In the case group, 109 pfu of phage particles was inoculated intraperitoneally cavity, at the meantime and every 12 hr upto 4 doses. After inoculation of mice with 107 cfu of P. aeruginosa, they became ill and eventually, collapsed within 24 hr of inoculation. When both P. aeruginosa and phage (107 cfu of bacteria and 109 pfu of phage) were administered simultaneously followed by 4 other doses of 109 pfu of phage every 12 hr, no mortality was observed at all. The numbers of dead mice in the control and treated groups were 10 and 0, respectively. They concluded that in mice, which inoculated with P. aeruginosa, bacteriophage administration showed significant protection against mortality. Studying the kinetics of bacterium and phage multiplication in these mice showed that in the absence of phage the P. aeruginosa multiplied in mice almost as fast as in broth culture. When phage was given to these mice intraperitoneally, the phage multiplied rapidly on the bacteria and prevented massive multiplication, resulting in a decline in bacterial numbers. 53 Naturally occurring, antibiotics resistance bacteria were used to measure the growth of phage in vivo by Richard et al. (2004). E. coli RRU and the pathogenic K-1-capsualted E. coli ATCC 23503 were used as host for phages T4 and K1-5. 107 log-phase bacteria were employed. Newly weaned Rattus rattus rats were used as experimental animals oral doses were administered observations were made in the digestive system. For phage and bacterial samples, rats were killed, dissected, the guts sectioned and the contents removed by squeezing. Four T4–like coli phages with broad host ranges for diarrhoea associated E. coli serotypes were isolated by Sandra et al. (2004) from stool specimens of pediatric diarrhoea patients and from environmental water samples. All four phages showed a highly efficient gastrointestinal passage in adult mice when added to drinking water. Viable phages were recorded from the faeces in a dose dependent way. The minimal oral dose for consistent faecal recovery was as low as 103 pfu of phage/ml of drinking water. In conventional mice, the orally applied phage removed restricted to the gut lumen and as expected for a non-invasive phage, no histopathological changes of gut mucosa were detected in the phage-exposed animals. E. coli strains introduced into the intestines of conventional mice and traced as ampicillin – resistant colonies were efficiently lysed in vivo by phage added to the drinking water. 54