Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
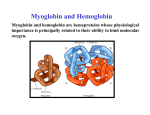
Protein Structure Related to Function How the structure of a given protein determines its function? Why does haemoglobin carry oxygen or Insulin take part in Glucogenesis and not vice versa?? Introduction: The ability of a protein to perform its function in the cell depends in part upon its ability to assume and retain its proper functional conformation. The proper conformation is achieved by regulated folding during synthesis, aided by chaperone proteins. Mutations and other changes that divert proteins from their normal folding pathways or that destabilize their native state may underlie several human diseases. For example, alterations of the thermodynamic stability in folding of the native state and critical folding intermediates may prevent folding into a functional conformation on a biological scale. Oxygen Binding Proteins: Hemoglobin and Myoglobin: The oxygen-carrying proteins in vertebrates are hemoglobin and myoglobin. Hemoglobin is found in red blood cells, and serves to carry oxygen in the blood and transports carbon dioxide and hydrogen ion. Myoglobin, which is located in muscle cells, provides a reserve supply of oxygen and facilitates movement of oxygen within muscles. Myogobin is a single polypeptide chain, containing one heme prostetic group. Hemoglobin, however, is a tetramer consisting of two alpha chains, and two beta chains an _2_2 subunit structure. The protein portion of myoglobin and each of the hemoglobin subunits are extremely similar and typical of the globins. Myoglobin, for example, is extremely compact. Approximately 75% of the main chain of myoglobin is in an alphahexical conformation. There are eight major helical segments (labeled A, B, C, ....), five nonhelical segments between the helical segments, plus two residues at the aminoterminal end (labeled NA1 and NA2) and five residues at the carboxyl-terminal end (labeled HC1 - HC5). The interior of the protein is composed almost entirely of nonpolar residues (e.g., leucine, valine, methionine, phenylalanine). Essentially identical structure is seen in each of the hemoglobin subunits. The ability of myoglobin or hemoglobin to bind oxygen depends on the presence of a heme prosthetic group. It is the heme that gives these proteins their characterisitc red color. All four chains of hemoglbin contain one identical heme prostetic groups. All of the heme in both myoglobin and hemoglobin are protoporphyrin IX structures, made up of four pyrrole rings. The pyrrole rings are joined together by methene bridges to form a tetrapyrrole ring. Attached to the edges of the ring are four methyl groups, two vinyl groups and two propionate groups. Located at the center of the protoporphyrin ring is an iron(II) ion. The iron(II) is bound to the four nitrogens of the protoporphyrin ring in a position slighly below the plane of the tetrapyrrole ring. The iron(II) can also form bonds at axial positions above and below the plane of the ring (fifth and sixth coordinate positions). It is the iron(II) that binds oxygen. Iron(III) has no oxygen-binding affinity. One axial position of the iron(II) is occupied by a histidine nitrogen from the protein chain (called the proximal histidine; histidine F8 in both myoglobin and hemoglobin). The second axial position is partially obscurred by another histidine (called the distal histidine; histidine E7 in both myogobin and hemoglobin). This partial obstruction of the axial position prevents the heme from binding oxygen too tightly. Oxygen is bond at an angle, weakening the iron(II)-oxygen bond. The binding of oxygen pulls the iron(II) into the plane of the heme. Allosteric interactions between the heme subunits enable hemoglobin (not myoglobin) to coordinately bind and transport oxygen, carbon dioxide and hydrogen ion. The binding of an oxygen to a hemoglobin heme "pulls" the F helix of the heme subunit, initiating conformational changes that enhance the binding of addtional oxygen to the remaining three hemes (i.e., cooperative binding). Further, the affinity of hemoglobin for oxygen is carbon dioxide- and pH-dependent. Both hydogen ions and carbon dioxide promote the release of oxygen in peripheral tissues. In the lungs oxygen promotes the release of bound hydrogen ions and carbon dioxide. The purpose of this tutorial is to provide programmed tutorials and 3-dimensional images of hemoglobin and myoglobin that can be manipulated to investigate the structures of these proteins. Hemoglobin Allostery: Hemoglobin is the oxygen carrier in the blood. Hemoglobin is a protein made up of four chains - two alpha and two beta chains. The arrangement of the chains is such that hemoglobin may be viewed as a dimer of dimers, each dimer being made up of one alpha and one beta chain. Each one of the chains carries a non-covalently bound heme group that binds oxygen. The heme group is a macrocyclic molecule - called a porphyrin – which has a bound iron atom. Each heme group can bind one oxygen molecule. Thus, the hemoglobin tetramer can bind a total of four oxygen molecules. Hemoglobin also serves the purpose of transporting carbon dioxide. Oxygen is bound by hemoglobin at the lungs, carried through the blood stream to various tissues, where it gives up the oxygen, and binds carbon dioxide and transports it back to the lungs. Carbon dioxide is bound as the bicarbonate ion, HCO3-, at the dimer interface and not to the heme. The binding of oxygen to hemoglobin is cooperative. Thus, binding of one oxygen causes the next oxygen to bind more strongly, the third oxygen even more strongly and so on. In fact, hemoglobin is usually observed in only two states - all four chains carrying oxygen, or completely oxygen free. This is shown by the oxygen binding curve in fig. Figure: Oxygen binding curves for myoglobin and hemoglobin. In contrast to myoglobin, the single chain oxygen storage molecule found in tissues, which displays normal saturation binding of oxygen, the curve for hemoglobin is sigmoidal, indicating cooperative binding. The modulation of the affinity of a site for a ligand by ligand binding at another site is called Allostery. Hemoglobinopathies: Sickle Cell Disease (SCD) A Hemoglobinopathy Disorder Sickle Cell Disease (SCD) is caused by the malfunction of the red blood cells in affected individuals causing a very severe form of anemia. Inheritance and Frequency The gene defect for sickle cell disease is an autosomal recessive genetic trait and is unknowingly passed down from generation to generation. This faulty gene only emerges when two carriers have children together and pass it to their offspring. For each pregnancy of two such carriers, there is a 25% chance that the child will be born with the disease and a 50% chance the child will be a carrier for the gene defect. It is estimated that on average, sickle cell disease effects 1 of every 1,300 infants in the general population and approximately 1 of every 400 of African descent. It is seemingly more common in individuals of African and Mediterranean ancestry; however, it is not limited to these groups. Ancestor groups should not be a reason to avoid screening for this disorder. It can be found in children of all backgrounds. Signs & Symptoms Untreated newborns often develop septicemia, an infection of the blood, and die within a few weeks of birth. Long Term Effects: There is an extremely high mortality for sufferers under the age of five, although, better treatments are becoming available to help cope with this disease.