Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
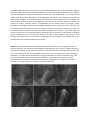
Dynamic analysis of early steps of midgut and hindgut morphogenesis during blastula extension in chicken embryos. Vincent Fleury, Hans Bodart and Nicolas Chevalier Laboratoire Matière et Systèmes Complexes/UMR7057 Université Paris Diderot 10 rue Alice Domon et L2onie Duquet 75013 Paris, France Abstract Considering the importance of the digestive function, the formation of the digestive tract has for long received scientific attention. However, little work was so far dedicated to dynamic studies of the morphogenesis of the gut at the earliest stages. This is readily explained by the fact that the gut is positioned “underneath” during early stages of development on the endoerm side, and even “inside” the embryo at later stages. In this article we use a recently developped protocol for observing chicken development on embryos which are turned “upside down”, to analyze the pattern of movemetns during fomation of the gut, and their reconnection at the caecum. The main result of this article is that there exists a biomechanical explanation to the presence of wider and thicker entities (the hindgut, the stomach) at the extremity of a thinner and less dense tube (the midgut and foregut). This is due to the structure of the posterior extension of the blastula, at the end of gastrulation. Introduction. Most embryologic studies in vertebrate embryology show embryos from a dorsal point of view. This is because vertebrate embryos lie naturally on their ventral side, either on the egg for the anamniotes, or on a mass of yolk for the megalolecithal eggs of birds. This is probably why the studies of the endoderm development, and especially of the digestive tract, lag somewhat behind somites or limb development studies. However, the digestive tract is arguably one of the msot important group of organs of the vertebrate body, and it deserves being understood in detail. This is all the more important as several developmental pathologies, called generically Hirschprung syndrom[00], are related to defects in the neural wiring of the gut. Recent work suggest that these defects are themselves related to the existence of a barrier at the hindgut/midgut frontier which is anatomically identified as the caecum[00]. One may expect that different genetic pathways are responsible for the existence of such a dichotomy between a distal thick and wide organ, as the hindgut, and a thinner, softer and narrower proximal gut,=. It is actually true that the gut has evolved many very specialized functions, as shown for example in the differences in rabbit gut morphology and physiology[00], vs the avian gut [00]. In man, conversely, the caecum has no function, and remains as a vestigial useless organ in the form of the appendix[00]. However, the existence of a dichotomy between a thin gut and a thick gut suggests that there might be a more fundamental explanation to this observation. We present here a series of dynamic observations of embryo development as pbserved “from underneath”, by turning embryos upside-down during their development, and imaging the ventral side. The biomechanical analysis of these movies suggests an correlation between the formation of the hindgut and of the midgut, and even of more anterior portions of the digestive tract such as the stomach. Indeed, as shown the process of embryo extension causes an elongatipon of the median portion of the gut, finally indetified as the midgut. Conversely, the hindgut undergoes a more compressive stress, by which the area of the presumtive hindgut and caecum, thicken, instead of thinning dn,as does the gut. Thjis can be illustrated by the stretch of a rubber strip held at some point of the strip : the portion ahead of the point where the trip is held will sytretch, while the portion posterior to the position of holding will be compressed. By this mechanism, having a thin gut, comes together with having a thick hindgut, and the geometry of the caecum is dependent on the topology of the median axis, at the position where the endoderm folds over the chord. Results. We have generated movies of embryo development at most if not all stages from the “as received” embryo, until formation of the abdominal fold. Movie 1, Fig. 1 shows a complete sequence of movement starting by the rotatory movements on the blastula and continuing by gastrulation, and up to until formation of the initial body folds of the embryo. It is an underneath movie, showing the endoderm. It is observed that the blastula is strongly stretched in the posterior direction. Fig. 2A,B shows a mechanical analysis of the strain in the blastula, showing that the blastula is actually streteched in both the anterior and posterior direction, although much strongly in the posterior direction. The visual impression of Movie 1 is shows it obviously. Figure 1. Figure 2. When the patern of stretch is analyzed during time (Fig. 2(C)), it is observed that the stretch is at first global, but progressively, th stretch is concentrated in the middle segment, while both ends of the median axis move “en bloc”, or even slightly in compression. It is a classical fact in elasticity, that, when a piece of tissue is stretched in one direction, it thins down in the other. This is indeed the case during blastula extension, as the tissue along the median axis narrows in the let-right direction, as it is stretched (Fig. 3). However, not all the tissue thins down. Carefull inspection shows that the area of the node remains thicker as it is pulled posteriorly (Fig. 4). Careful dissection of the hypoblast at an early developmental stage (primitive streak stage Hamilton and Hamburger stage 4) shows that the hypoblast is actually in contact, and even attached to the mesoderm which spreads underneath the ectoderm in the area of the primitive streak. It is likely that, since the cellular condensation along the primitive streak thickens it, cells reach out towards the hypoblast and adhere to it. Recent analysis of the pattern of pull of the mesoderm shows that the mesoderm, after ingressing through the primitive streak, pulls in an antero-posterior pattern (Supp. Fig. 1, reprinted from Ref. XX). Since the mesoderm pulls as it spreads in between the ectoderm and endoderm, a similar stretch is observed on both tissues, namely, there is an elongation of the gastrulating embryo along the median axis simultaneously on the endoderm and the ectoderm. This generates a tyical pattern of strain which is qaudrupolar (Fig. 5) both on the ectoderm, and the endoderm. If we follow the extremity of the median axis as it is pulled posteriorly, we observe that a series of folds are eventually formed which are perpendicular to the pulling direction (Fig. 6). These folds are : the amniotic fold (Fig. 6(A), the body contour fold (Fig. 6(B)), and the gut fold (Fig. 6(C)). These folds grow and form the walls of these structures (fig. 7). We managed to follow the initial steps of formation of the hindgut wall. Indeed, there exists a furrow that follows exactly the contours of the compressed and thicker node area, located just above the future anal pit (Fig. 8). The global pattern of the hindgut and caecum are actualy already recognizable in the pattern of the tissues at this stage. Conclusion We were able to follow in vivo the development of chicken embryos as observed “from underneath”. This allows one to observe directly the ventral side and especially the endoderm of embryos. The movements which are observeed suggest that the schematic structure of the gut, which is composed of a long and thin middle segment, and of wider segments at both ends, is not at all a matter of chance, but a genuine biomechanical consequence of a simple fact : when stretched by the edges, a piece of tissue thins down in its middle part. The transition from a flat structure to a 3D tube of the hindgut occurs by folding at the extremity of the of the median axis which “collide” against the peripheral tissue. The cecum structure arises from progressive closure of the reconnection of the elongated part with the thicker part, and is already recognizable on the tissue pattern when they are still flat (Fig. 9). The body of the embryo is formed by a system of encased folds, which roll up close up one inside the other : the gut rolls up to form a tube, the body contour rolls up to wrap the gut, and the amnios closes up to wrap the entire animal.