Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
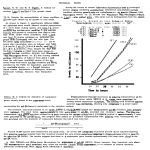
Mammalian Genome 7, 138–142 (1996). © Springer-Verlag New York Inc. 1996 Microsatellite mapping of the bovine roan locus: a major determinant of White Heifer Disease C. Charlier,1 B. Denys,1 J.I. Belanche,1 W. Coppieters,1 L. Grobet,1 M. Mni,1 J. Womack,2 R. Hanset,1 M. Georges1 1 2 Département de Génétique, Faculté de Médecine Vétérinaire, Université de Liège, 20 Boulevard de Colonster (B43), B4000-Liège, Belgium Department of Veterinary Pathobiology, Texas A&M, College Station, Texas, USA Received: 30 May 1995 / Accepted: 6 September 1995 Abstract. In the Belgian Blue Cattle breed, coat color variation is mainly under the influence of a single autosomal locus, the roan locus, characterized by a pair of codominant alleles: r+ (black) and R (white). Heterozygous r+R animals have intermingled black and white hairs, yielding the ‘‘blue’’ phenotype typical of the breed. Major interest for the roan locus stems from its pleiotropic effect on fertility, owing to the critical role of the R allele in the determinism of White Heifer Disease. We describe the linkage mapping of the roan locus to bovine Chromosome (Chr) 5, in the interval between microsatellite markers BPI and AGLA293, with an associated lodscore of 11.2. Moreover, we map a candidate gene, the Steel locus coding for the mast cell growth factor, to bovine Chr 5. Introduction Despite its name, the Belgian Blue cattle breed is characterized by a coat color polymorphism dominated by three distinct phenotypes: black spotted, blue spotted, and white animals (Fig. 1). Closer scrutiny of the pigmented sectors in blue animals reveals a mixture of black and white hairs. This trimorphism is reminiscent of the well-known coat color pattern found in the Shorthorn breed with its red spotted, roan (red and white hairs intermingled) spotted, and white individuals. As early as 1905, Barrington and Pearson (1906) used segregation data of these Shorthorn phenotypes to test Mendel’s theory of particulate inheritance. A model assuming a single diallelic locus with two codominant alleles fitted the observed segregation ratios reasonably well. However, the occurence of a nonneglectable proportion of cases violating Mendel’s rules led Barrington and Pearson to question the relevance of Mendel’s theory. Subsequently several authors proposed more elaborate models to account for the occasional exceptions (for example, Evvard et al. 1930), but it was Wright (1917) who concluded that if the more complicated models explained the exceptions, the original simple model proposed, though questioned by Barrington and Pearson themselves, explained the overall data best by far, and that it was, therefore, likely that one autosomal locus with a pair of codominant alleles played a major role in the determination of the observed trimorphism. More recently, Hanset (1959, 1985) pointed towards the close resemblance between the segregation ratios observed in the Belgian Blue cattle breed for the black, blue, and white phenotypes, and in the Shorthorn breed for the red, roan, and white phenotypes. As Wright did earlier in Shorthorn, Hanset concluded from these studies that a single diallelic locus–the roan locus–, with codominant alleles r+ (black) and R (white), was superior to any other Correspondence to: C. Charlier model in explaining the overall observations. Exceptions could virtually all be accounted for by assuming incomplete penetrance for the Rr+ or blue genotype, a proportion of Rr+ animals being recorded as white (.9%) or black (.9%). This is actually not unexpected in light of the documented variation in the ratio of white to black hairs in blue sectors, yielding very light to dark blue animals. Circumstantial evidence indicates that this varying blackto-white-hair ratio may be under genetic control, but to the best of our knowledge this has not been studied in detail. Moreover, while in the Belgian Blue cattle breed the recessive s allele at the Self locus [responsible for piebald spotting, (e.g., Ibsen 1993)] is virtually fixed, there is substantial variation in the expressivity of the piebald trait, with some extreme animals being nearly completely devoid of pigmented sectors. Finally, some poorly characterized phenotypes such as the ‘‘colorsided’’ spotting attributed to the segregation of a dominant Sc factor at the same Self locus (Wriedt, 1925), are segregating in the Belgian Blue population at low frequency, potentially complicating the classification of animals into discrete phenotypic classes. The segregation of the same roan locus in both the Shorthorn and Belgian Blue cattle breed would not be surprising given the important migration of Shorthorn genes that occurred into the Belgian cattle population during the 19th century. The fact that the pigmented hairs are red in Shorthorn while black in the Belgian Blue reflects the high frequency of the dominant E allele at the Extension locus (Charlier et al. 1995b), probably imported through Dutch black and white animals. Occasionally, however, red spotted or red-roan spotted animals are still encountered in the Belgian Blue breed, pointing towards the segregation of the e allele or ‘‘red factor’’ in this population albeit at low frequency. Besides the fact that it attracted the attention of famous geneticists in the beginning of this century, the roan locus has intrigued scientists because of its predominant role in the determinism of White Heifer Disease. This disorder is characterized by a range of anomalies of the female genital tract resulting from the aberrant development of the Müllerian ducts. Lesions include the occurrence of a posterior vaginal occlusion, absence or incompletely developed vagina, cervix, uterine body, and one or both uterine horns; these different lesions occur either isolated or in conjunction. Ovaries are invariably found not to be affected in White Heifer Disease (Hanset and Ansay 1961). As its name implies, a striking feature of this disorder is its high frequency in association with the white coat color phenotype in cattle populations segregating for the roan locus. These include in particular the Shorthorn and Belgian Blue cattle breeds. In the Belgian Blue cattle breed, more than 90% of affected animals have the white phenotype, the remainder being more often blue than black. Studies in this breed (Hanset 1969a) led to the conclusion that White Heifer Disease has a multifactorial basis, including genetic and environmental components. The roan locus would be C. Charlier et al.: Bovine roan locus and White Heifer Disease 139 Fig. 1. Illustration of the coat-color polymorphism segregating in the Belgian Blue cattle population, characterized by the three genotypes: white, black, and blue. the major genetic determinant, acting in conjunction with one or, more probably, several auxiliary genes. The heritability of the defect within the white phenotype has been estimated at 0.2–0.25. Evidence was given that the involvement of the roan locus is probably due to a direct effect of the R allele, rather than to a closely linked gene, or separate allele at the roan locus itself. In the Belgian Blue cattle breed, the incidence of this disorder has been as high as 5–10% of the cow population in the 1950s. Because of systematic selection against the auxiliary genes, through the elimination of affected females and progeny testing of sires, the incidence of the disease has been reduced to 1.5% (Hanset 1969b). In this paper, we describe the localization of the bovine roan locus to Chromosome (Chr) 5, in the interval between microsatellite markers BP1 and AGLA293. Since the linkage analysis was based on the hypothesis of a simple monogenic determinism of the coat color trimorphism with two codominant alleles r+ (black) and R (white), this study supports the validity of the corresponding model. Moreover, using somatic cell hybrids, we map a candidate gene, the Steel locus coding for the ligand of the c-kit tyrosine kinase receptor, to the same bovine chromosome. Materials and methods Pedigree material: the ‘‘Sart-Tilman’’ pedigree. To map the roan locus, we used a pedigree composed of six white Belgian Blue sires mated to 37 black spotted Friesian cows. All six sires have been extensively progeny tested in the Belgian Blue cattle population, and their proportions of white, blue, and black offspring indicate that they are all of the RR genotype. Conversely, all Friesian cows are assumed to be of r+r+ genotype, as the roan phenotype is not segregating in this population. The resulting 37 ‘‘F1’’ cows were all phenotypically blue, as predicted from the monogenic model with a pair of codominant alleles. The F1 r+R cows were backcrossed to three progeny-tested white Belgian Blue sires (RR), yielding 105 backcross (BC) offspring segregating at the roan locus. ‘‘F1’’ and ‘‘BC’’ animals were held at the Sart-Tilman experimental station and phenotyped for coat color by repeated visual examination; 59 blue spotted and 46 white BC offspring were obtained (x21 = 1.6, p ≈ 0.2). The described animals are part of a larger pedigree (the Sart-Tilman pedigree) initially constructed to map the mh locus (Charlier et al. 1995a). One branch of this pedigree was ignored in this analysis as it segregates for the ‘‘colorsided’’ or Sc factor (Wriedt, 1925) that interacts with the expression of the roan locus. The IBRP panel of international bovine reference families was used for map construction (Barendse et al. 1994). Marker genotyping. A battery of 213 microsatellite markers distributed over the 29 bovine autosomes and compiled from three partially overlap- ping and recently published bovine maps (Barendse et al. 1994; Bishop et al. 1994; Georges et al. 1995), was chosen to allow for a systematic scanning of the bovine genome. The primer sequences used for the amplification of the Chr 5 microsatellites are reported in Table 1. Microsatellite genotyping was performed essentially as previously reported (Georges et al. 1995). Linkage analysis. Linkage analyses were performed on a Sun Sparc Classic workstation with version 5.0 of the LINKAGE programs (Lathrop and Lalouel 1984). To deal with the large number of marriage loops, the ‘‘roan’’ pedigree was split into 37 nuclear families, each corresponding to a single F1 cow, with respective Belgian Blue sire, Friesian dam, Belgian Table 1. Primer sequences used for the amplification of the seven chromosome 5-specific microsatellite markers. BM6026.UP1 BM6026.DN1 BP1.UP1 BP1.DN1 AGLA293.UP1 AGLA293.DN1 AGLA254.UP1 AGLA254.DN1 IGFL.UP1 IGFI.DN1 BM315.UP1 BM315.DN1 GCAAGTAAGACCCAACCAAC ACTGATGTGCTCAGGTATGACG AAAATCCCTTCATAACAGTGCC CATCGTGAATTCCAGGGTTC GAAACTCAACCCAAGACAACTCAAG ATGACTTTATTCTCCACCTAGCAGA GCTGCTTGGCACAGGCAAA GGATTAATTTCTGGACTCTG GGGTATTGCTAGCCAGCTGGT CATATTTTTCTGCATAACTTGAACCT TGGTTTAGCAGAGAGCACATG GCTCCTAGCCCTGCACAC Fig. 2. Sequence alignment of the fifth exon of the bovine (Zhou et al. 1994), human (Martin et al. 1990), and murine (Brannan et al. 1992) mast cell growth factor gene. The location of the primers used to generate a bovine-specific 106-bp STS are underlined. 140 C. Charlier et al.: Bovine roan locus and White Heifer Disease Table 2. Lodscore tables obtained by pair-wise linkage analysis between the roan locus and the corresponding Chr 5 microsatellite marker. The IGFI microsatellite proved monomorphic in our pedigree material, therefore yielding no information (NI). u Marker 0 0.01 0.05 0.1 0.2 0.3 0.40 BM6026 BP1 AGLA293 AGLA254 IGFI BM315 −` −` −` −` NI −` −0.93 11.10 0.83 −24.55 NI −51.67 3.85 11.47 3.14 −9.40 NI −27.10 5.20 10.75 3.70 −3.67 NI −17.03 5.32 8.56 3.53 0.70 NI −7.86 4.20 5.91 2.73 1.92 NI −3.39 2.39 2.97 1.54 1.57 NI −1.02 Fig. 3. Lodscore curve obtained with the LINKMAP program (Lathrop and Lalouel 1984), by sliding the roan locus through the fixed Chr 5 marker map. Recombination rates between adjacent markers were converted to centiMorgan with Haldane’s mapping function. Blue mate(s), and resulting BC offspring. Coat color phenotypes were encoded as a ‘‘numbered alleles’’ system (11: white; 12: blue; 22: black), or as an ‘‘affection status’’ (1: white; 2: blue or black) to test the effects of misclassification by varying the penetrance for the three different genotypes. Synteny mapping of the Steel locus. Primers that should amplify part of exon 5 of the bovine Steel gene were designed from the published bovine stem cell factor cDNA sequence (Zhou et al. 1994), assuming conservation of intron position between bovine, human, and mice. The primers were targeted towards poorly conserved segments of the exon to specifically amplify bovine product, while avoiding cross-amplification of rodent product in particular (Fig. 2). These primers were shown to amplify the expected 106-bp fragment from bovine genomic DNA, while yielding no product from genomic DNA of the rodent species used to construct the somatic cell hybrid panel (data not shown). A previously described panel of somatic cell hybrids (Dietz et al. 1992) was tested for the segregation of this Steel STS. The resulting segregation patterns were analyzed according to Chevalet and Carpet (1986). Results To find a marker for the roan locus, we undertook a systematic chromosome-by-chromosome genome scan, using a selected panel of 213 microsatellite markers (see Materials and methods) spread across the bovine autosomes with average spacing of 15 cM. Sequential pair-wise linkage analysis was performed between the genotyped microsatellite markers and the roan locus, with the MLINK program. When a set of seven markers covering bovine Chr 5 was screened, four of the tested markers yielded positive pair-wise lodscores with roan: BM6026, BP1, AGLA293, and AGLA254 (Table 2). According to previously published maps (Bishop et al. 1994; Georges et al. 1995), these correspond to the microsatellites located on the centromeric side of the Chr 5 linkage group. To more precisely determine the relative map positions of these four microsatellites, we genotyped the IBRP pedigrees with the corresponding markers. Genotypic data from the roan and IBRP pedigrees were combined to construct male and female linkage maps using the ILINK program. Maximum likelihood order and associated male and female recombination rates between adjacent markers, for the centromeric side of the Chr 5 microsatellite map, are respectively: Cen–BM6026(14.1-8.2)–BP1(23.3-10.3)– AGLA293(22.8-40.6)–AGLA254. The most likely locus order given our data set is in agreement with a previously reported map of bovine Chr 5 involving the same markers (Bishop et al. 1994). The odds versus the next most likely order (positioning AGLA254 on the centromeric side of the linkage group) was 69:1 given our data. To position the roan locus on the established map, its location was varied with respect to the microsatellite markers held at fixed positions, and corresponding lodscores were computed with LINKMAP (Fig. 3). As all informative meioses for the segregation of the roan locus were female, the position of this locus is given with respect to the female marker map. This analysis points towards the most likely position of the roan locus in the interval between BP1 and AGLA293, at 3.7 cM from the former and with an associated lodscore of 11.2. Positions yielding a lodscore one unit below the maximum of 11.2 roughly define a 95% confidence interval of 11.8 cM. As deviations from a strictly Mendelian inheritance pattern have repeatedly been documented for the studied coat color tri- C. Charlier et al.: Bovine roan locus and White Heifer Disease 141 Table 3. Effect of misclassification on the positioning of the roan locus with respect to microsatellite marker BP1, and on the likelihood of the pedigree data. % Misclassification Distance from BP1 (cM) Odds 0 2.5 5 10 3.7 2.8 1 0 1 2.6 3.3 1.3 morphism, we looked for evidence of phenotypic misclassifications in the analyzed data set. Frequent misclassification would manifest itself by a high incidence of double recombinants. Analysis of the individual genotypes revealed only one such offspring in the BP1–roan–AGLA293 chromosome segment assuming that order, which is not unexpected given the number of analyzed backcross offspring (108) and the estimated distance between BP1 and AGLA293 (10.3%). Moreover, a high incidence of double recombinants would confuse the multipoint analysis, generating discrepancies with the two-point analysis. It can be seen that the closest microsatellite marker as determined by two-point analysis (BP1) also appears as the closest marker when performing multipoint analysis. To more formally address the misclassification issue, lodscore curves were regenerated with LINKMAP as described, but allowing for a varying percentage of misclassification in the backcross generation, that is, 2.5%, 5%, or 10% of genetically white (RR) offspring would be classified as blue (r+R), and vice versa. The genotypes of the parental and F1 generations were considered unambiguous, as all used Belgian Blue sires (RR) have been progeny tested for coat color genotype in the commercial population, and the Black and White Friesian dams come from a monomorphic black (r+r+) population. As shown in Table 3, allowing for misclassification yielded for a marginal increase in likelihood at best, while shifting the roan locus closer to BP1. Altogether, these results point towards absence of significant phenotypic misclassification in our data set. As microsatellite marker BP1 was shown to be very closely linked to the myogenic factor-5 (Myf5) gene (Bishop et al. 1994), the present linkage study positions the roan locus to a chromosome segment bounded by two anchored reference loci: Myf5 and IGFI. These two type I markers are also syntenic in human as well as mice, where they map to Chr 12 and 10 respectively (Taylor et al. 1993). Intriguingly, the corresponding chromosome segment is know to harbor the Sl or Steel locus in mice, positioned at approximately 8.5 cM from Myf5 and 11 cM from IGFI (Taylor et al. 1993). The Sl locus codes for the mast cell growth factor (Brannan et al. 1992), and mutations at this locus invariably affect coat color pigmentation (Green 1990). Although the phenotypic expression of the murine Sl mutations differs in several points from the bovine roan phenotype and associated White Heifer Disease (see Discus- sion), it would nevertheless qualify as a reasonable candidate gene if it mapped to the corresponding chromosomal region. To start answering that question, we typed a bovine somatic cell hybrid panel for a bovine-specific Sequence Tagged Site developed from the bovine stem cell factor cDNA sequences (see Materials and methods). Table 4 summarizes the obtained results, clearly demonstrating that the homolog of the murine steel locus indeed maps to bovine Chr 5. Discussion The observation of an entirely blue F1 generation obtained by crossing white Belgian Blue sires with black Friesian dams, and the subsequent 50%–50% segregation of blue and white offspring in a backcross to white Belgian Blue sires, clearly supports a monogenic hypothesis with a pair of codominant alleles to explain the coat color trimorphism observed in the Belgian Blue Cattle breed. This assumption is dramatically strengthened by the identification in this study of a genetic marker on bovine Chr 5 that cosegregates with the postulated roan locus. Given the similarities between the coat color polymorphism in the Belgian Blue and Shorthorn populations, as well as the documented links between both populations, it is reasonable to extend our conclusions to the Shorthorn population and to assume that the same roan locus with the same r+ and R alleles determines the coat color patterns observed in both breeds. The black versus red color of pigmented hairs in Belgian Blue and Shorthorn respectively is assumed to reflect the near fixation in both populations of different alleles at the unlinked Extension locus. As the Extension locus has recently been identified in cattle (Charlier, in preparation), this hypothesis becomes easily testable. Comparative mapping data allow us to predict that the bovine roan locus maps to an evolutionary conserved chromosome segment bounded by the Myf5 and IGF-I loci. Interestingly, a possible candidate for the bovine roan gene has previously been assigned to this conserved chromosome fragment: the steel or Sl locus mapping to Chr 10 in mice. Mutations at the Sl locus in mice (coding for the mast cell growth factor, now known to be the ligand of the c-kit tyrosinase kinase receptor), like mutations at the W locus (coding for the c-kit tyrosinase kinase receptor), cause a reduction in pigmentation, sterility, and macrocytic anemia (for example, Green 1990; Jackson 1994). Generally speaking, the reduction in pigmentation manifests itself as black-eyed white animals in homozygotes, and as a diluted coat color with some white spotting in heterozygotes. The coat color manifestations, however, are quite heterogeneous, and features of some Sl or W mutations can be considered reminiscent of the studied bovine coat color trimorphism. Indeed, black-eyed white animals homozygous for a number of Sl and W mutations Table 4. Synt. Chr. Mark. Conc. Corr Synt. Chr. Mark. Conc. Corr U1 U2 U3 U4 U5 U6 U7 U8 U9 U10 U11 U12 U13 U14 U15 16 9 5 21 10 3 25 29 18 1 13 22 4 27 6 RDB23 ESR IFNG RF131 TCRa AMY1 IGF2 COL1A UPK1 RDB21 OT GPX PGY3 PLAT CASK 59% 45% 94% 54% 50% 50% 54% 60% 43% 45% 76% 73% 54% 53% 28% .04 −.12 .89 −.11 −.05 −.18 −.04 .13 −.39 −.12 .45 .38 −.04 −.01 −.48 U16 U17 U18 U19 U20 U21 U22 U23 U24 U25 U26 U27 U28 U29 X 11 2 8 15 23, 20 19 7 17 14 27 26 12 24 28 X POMC FUCA1 IFNO CD3e PL GMP INSR ALDH2 TG ANT OAT F10 MBP ACTA DMD 53% 59% 73% 48% 52% 43% 45% 50% 45% 45% 59% 73% 65% 50% 57% −.07 .09 .42 −.22 −.16 −.28 −.35 −.05 −.18 −.17 −.16 .42 .48 .00 −.08 142 exhibit pigmentation of the ears and/or snout as observed in some white individuals from the Belgian Blue Cattle breed. Moreover, mice heterozygous for some mutations at the same loci exhibit white speckling or interspersed white hairs, yielding a phenotype referred to as roan. The sterility characterizing most Sl and W homozygous mutants results from the absence of primordial germ cells from the gonads. These symptoms, therefore, differ dramatically from the lesions characterizing White Heifer Disease, which result from the aberrant development of the Müllerian ducts. Interestingly, however, a neutron irradiation-induced mutation—the cloud-gray mutation—has been described in mice (Kelly 1974), for which 17 out of 53 analyzed homozygous females and one out of 66 heterozygous females showed imperforate vaginas, reminiscent of the symptoms observed in White Heifer Disease. As its name implies, this mutation also has a diluting effect on coat color. As this mutation did not recombine with the Sl mutations in 98 offspring from double heterozygous mice, it was assumed to be a new Sl allele (Slcg), although it could as well be a mutation in a very closely linked gene. While mice homozygous for Sl or W mutations exhibit severe anemia, in cattle to the best of our knowledge no association has been reported between coat color phenotype and haematopoietic parameters. The Steel factor is the ligand for the c-kit tyrosine kinase receptor and is thought to be involved in the migration and/or differentiation of the melanoblasts (reviewed in Jackson 1994). The phenotypes associated with the three genotypes at the bovine roan locus are in reasonable agreement with the expected effects of a mutation in the Steel gene. Melanoblast migration could be completely impaired in RR animals, resulting in unpigmented hairs. In r+R animals, a proportion of the hair bulbs would fail to be colonized by melanoblasts. This would imply that the number of melanoblasts originally colonizing the hair bulb is very low, either generally or more specifically in r+R individuals. Colonization by a single melanoblast, however, would be sufficient to generate the number of melanocytes required for full pigmentation of the hair, explaining why hairs from heterozygous r+R animals are apparently either fully pigmented or completely white. The availability of the bovine Steel cDNA sequence should allow us to rapidly test whether this gene cosegregates with the roan locus and, if this eventually proves to be the case, to look for causal mutation(s). Mapping the roan gene is the first step towards its isolation and characterization, which should shed some light on how a gene affecting coat color exercises a pleiotropic effect on the development of the Müllerian ducts. Acknowledgments. The financial support of the Institut pour l’Encouragement de la Recherche Scientifique dans l’Industrie et l’Agriculture (IRSIA) is greatly acknowledged. We thank Dr. Leif Andersson for stimulating discussions. We are indepted to Dr. Jay Hetzel, Dr. Alan Teale, and the BOVMAP consortium for providing us with DNA from the IBRP reference pedigrees. We are indepted to Prof. J.M. Bienfait, Prof. L. Istasse, and Dr. I. Dufrasne for their help in generating the Sart-Tilman pedigree. References Barendse, W., Armitage, S.M., Kossarek, L.M., Shalom, A., Kirkpatrick, B.W., Ryan, A.M., Clayton, D., Li, L., Neibergs, H.L., Zhang, N., Grosse, W.M., Weiss, J., Creighton, P., McCarthy, R., Ron, M., Teale, A.J., Fries, R., McGraw, R.A., Moore, S.S., Georges, M., Soller, M., Womack, J.E., Hetzel, D.J.S. (1994). A genetic linkage map of the bovine genome. Nature Genet. 6, 227–235. Barrington, A., Pearson, K. (1906). On the inheritance of coat color in cattle. Biometrika 4, 427–464. C. Charlier et al.: Bovine roan locus and White Heifer Disease Bishop, M.D., Kappes, S.M., Keele, J.W., Stone, R.T., Senden, S.L.F., Hawkins, G.A., Solinas Toldo, S., Fries, R., Grosz, M.D., Yoo, J., Beattie, C.S. (1994). A genetic linkage map for cattle. R. (1990). Brannan, C.I., Bedell, M.A., Resnick, J.L., Eppig, J.J., Handel, M.A., Williams, D.E., Lyman, S.D., Donovan, P.J., Jenkins, N.A., Copeland, N.G. (1992). Developmental abnormalities in Steel (17H) mice result from a splicing defect in the Steel factor cytoplasmic tail. Genes Dev. 6, 1832–1842. Charlier, C., Coppieters, W., Farnir, F., Grobet, L., Leroy, P., Michaux, C., Mni, M., Schwers, A., Vanmanshoven, P., Hanset, R., Georges, M. (1995a). The mh gene causing double-muscling in cattle maps to bovine chromosome 2. Mamm. Genome, in press. Charlier, C., Belanche, I., Brouwers, B., Coppieter, W., Karim, L., Vanmanshoven, P., Womack, J., Georges, M. (1995b). Mutations in the MSH receptor underly coat color polymorphism in cattle. In preparation. Chevalet, C., Carpet, F. (1986). Statistical decision rules concerning synteny or independence between markers. Cytogenet. Cell Genet. 43, 132– 139. Dietz, A.B., Georges, M., Threadgill, D.W., Womack, J.E., Schuler, L.A. (1992). Somatic cell mapping, polymorphism, and linkage analysis of bovine prolactin-related proteins and placental lactogen. Genomics 14, 137–143. Evvard, J.M., Shearer, P.S., Lindstrom, E.W., Smith, A.D.B. (1930). The inheritance of color and horns in blue gray cattle. Iowa Agric. Exp. Stn. Res. Bull. 133, 1–16. Georges, M., Nielsen, D., Mackinnon, M., Mishra, A., Okimoto, R., Pasquino, A.T., Sargeant, L.S., Sorensen, A., Steele, MR.R., Zhao, X., Womack, J.E., Hoeschele, I. (1995). Mapping quantitative trait loci controlling milk production by exploiting progeny testing. Genetics 139, 907–920. Green, M.C. (1990). Catalog of mutant genes and polymorphic loci. In Genetic Variants and Strains of the Laboratory Mouse, M.F. Lyon, A.G. Searle, eds. (Oxford: Oxford University Press), pp. 333–335. Hanset, R. (1959). L’hérédité des robes dans la race bleu-blanc de Moyenne et Haute Belgique. Ann. Méd. Vét. 3, 161–188. Hanset, R. (1969a). Croisements expérimentaux avec des génisses atteintes de la White Heifer Disease. Ann. Méd. Vét. 113, 3–11. Hanset, R. (1969b). La White Heifer Disease dans la race bovine de Moyenne et Haute Belgique: un bilan de dix années. Ann. Méd. Vét. 113, 12–21. Hanset, R. (1985). Coat colour inheritance in the Belgian White and Blue cattle breed. Génét. Sél. Evol. 17, 443–458. Hanset, R. Ansay, M. (1961). La White Heifer Disease: nouvelle description et essai de classification rationnelle de ses différentes formes. Ann. Méd. Vét. 105, 133–146. Ibsen, H.L. (1933). Cattle inheritance. Genetics 18, 441–482. Jackson, I.J. (1994). Molecular and developmental genetics of mouse coat color. Annu. Rev. Genet. 28, 189–217. Kelly, E.M. (1974). Mouse News Lett. 50, 52. Lathrop, M., Lalouel, J.M. (1984). Easy calculations of lodscores and genetic risk on small computers. Am. J. Hum. Genet. 36, 460–465. Martin, F.H., Suggs, S.V., Langley, K.E., Lu, H.S., Ting, J., Okino, K.H., Morris, C.F., McNiece, I.K., Jacobsen, F.W., Mendiaz, E.A., Birkett, N.C., Smith, K.A., Johnson, M., Parker, V.P., Flores, J.C., Patel, A.C., Fisher, E.F., Erjavec, H.O., Herrera, C.J., Wypych, J., Sachdev, R.K., Pope, J.A., Leslie, I., Wen, D., Lin, C.H., Cupples, R.L., Zsebo, K.M. (1990). Primary structure and functional expression of rat and human stem cell factor DNAs. Cell 63, 203–211. Taylor, B.A., Frankel, W.N., Burmeister, M., Bryda, E. (1993). Mouse Chromosome 10. Mamm. Genome 4 (Suppl.), S154–S163. Wriedt, C. (1925). Color-sided cattle. J. Hered. 16, 51–56. Wright, S. (1971). Color inheritance in mammals: cattle. J. Hered. 8, 521– 527. Zhou, J.-H., Hikono, H., Ohtaki, M., Kubota, T., Sakurai, M. (1994). Cloning and characterization of cDNAs encoding two normal isoforms of bovine stem cell factor. Biochim. Biophys. Acta 1223, 148–150.