Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
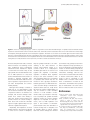
58 j Journal of Molecular Cell Biology (2009), 1, 58 –60 doi:10.1093/jmcb/mjp014 Published online August 14, 2009 Research Highlight Cohesin: A Multi-purpose Chromatin Glue Laura A. Dı´az-Martı´nez and Hongtao Yu* Department of Pharmacology, Howard Hughes Medical Institute, University of Texas Southwestern Medical Center, Dallas, TX 75390, USA * Correspondence to: Hongtao Yu, E-mail: [email protected] Long thought to be the glue responsible for holding sister chromatids together, cohesin has been found to be stickier than previously thought. Recent discoveries point to cohesin having a role in transcription regulation by mediating long-distance intra-chromosomal interactions. In higher eukaryotes, cohesin is known to bind to chromatin during interphase. As cells advance into mitosis, most cohesin is removed from chromosomes and only a tiny amount remains bound to the centromeres (Figure 1). This remaining cohesin, which contributes to sister-chromatid cohesion, is removed at the moment of anaphase onset. Almost immediately after that, in telophase, cohesin binds again to the chromatin. For a complex whose main function is thought to be in mitosis, this cycle of cohesin binding peaking in interphase and being removed in mitosis seems counterintuitive. If the main function of the cohesin complex is to hold sister chromatids together, then why would a cell want so much cohesin bound to the chromatin in G1, when the chromosomes have not been duplicated? And why is most cohesin removed from chromosomes in mitosis, when the definitive test of sister chromatid cohesion occurs? The answer to these questions, thanks to new discoveries, seems to be that cohesin also plays important roles in interphase through a contribution to chromatin structure and spatial organization. The first hints of a role for cohesin in interphase were the gene silencing defects observed in yeasts and flies containing mutations in cohesin subunits and cohesin loading factors (reviewed by Dorsett, 2009). These defects in gene expression involved the malfunction of enhancers, insulators or other transcription regulatory elements found kilobases away from the affected genes. These regulatory elements are thought to influence gene expression via formation of chromatin loops which bring them in close proximity to their targets. Central to this idea is the 11-zinc finger protein CTCF, which binds to both the nuclear matrix and the chromatin and has therefore been proposed to act as a tether that brings together specific chromatin sites. Although CTCF binding to DNA is sequence specific, there are an estimated 14 000 – 20 000 CTCF binding sites in the human genome, which happen to lie in gene-rich regions, hinting at an important role of CTCF in the regulation of gene expression. More importantly, CTCF has been shown to be involved in the formation of chromatin loops at several loci including the imprinted region Igf2/H19 and has therefore been considered the corner stone of chromatin organization in interphase (reviewed in Zlatanova and Caiafa, 2009). Interestingly, a link between CTCF and the cohesin complex has been discovered indicating that cohesin contributes to the formation of chromatin loops. The initial evidence for a role of cohesin in CTCF-related control of gene expression came from ChIP studies that mapped cohesin-binding sites in the human genome (Parelho et al., 2008; Wendt et al., 2008). These studies showed that the binding sites of cohesin overlapped with CTCF-binding sites, with 71% of the identified sites binding both cohesin and CTCF while 19% and 10% bind cohesin or CTCF alone, respectively (Parelho et al., 2008). The association of CTCF and cohesin at CTCF-binding sites was confirmed independently by an in vitro assay that examined proteins bound to a 169 bp DNA probe containing a CTCF-binding site. This assay identified the cohesin component Scc3/SA1 as a protein enriched in the fraction bound to the CTCF site, but not to a mutated site (Rubio et al., 2008). Using siRNAs to knock-down Rad21 or CTCF (Parelho et al., 2008; Wendt et al., 2008) or a mutant CTCF-binding site (Rubio et al., 2008), these studies showed that cohesin enrichment at CTCF sites was drastically diminished in the absence of CTCF, while cohesin inactivation did not significantly affect CTCF binding (Parelho et al., 2008; Wendt et al., 2008). More importantly, experiments using reporter plasmids containing enhancer/insulator elements demonstrated that binding of both CTCF and cohesin is necessary for the proper function of these elements (Parelho et al., 2008; Wendt et al., 2008). But, how does cohesin influence gene expression? The beautiful work of Hadjur et al. (2009) has now provided us with important clues. By studying the CTCF and cohesin association at the IFNG locus, they showed that cohesin and CTCF association at certain CTCF sites is increased in TH1 effector memory cells, which express higher amounts of IFNG compared with their precursors (nonpolarized CD4 T cells). That is, gene expression correlated with CTCF/cohesin binding at the endogenous locus. Using chromosome conformation capture (3C) techniques, they demonstrated that the increase in CTCF/cohesin binding at these regulatory sites, which are tens of # The Author (2009). Published by Oxford University Press on behalf of Journal of Molecular Cell Biology, IBCB, SIBS, CAS. All rights reserved. Journal of Molecular Cell Biology j 59 Figure 1 Cohesin as a multi-purpose glue. Cohesin is long known to form inter-chromatid bridges on replicated sister chromatids that are required for sister-chromatid cohesion. Recent studies have shown that cohesin collaborates with CTCF to form intra-chromosomal bridges in interphase, which play important roles in transcription regulation. This active role of cohesin as a chromatin glue involved in nuclear architecture is consistent with the large amounts of chromatin-bound cohesin observed in interphase. However, chromatin structures such as intraand inter-chromosomal interactions are incompatible with chromosome individualization and condensation and therefore have to be removed during mitosis. A small amount of cohesin (and CTCF) is known to remain at centromeres in mitosis. kilobases away from each other, correlates with an increase in crosslinking of such sites. Furthermore, these long-distance intra-chromosomal interactions depend on both CTCF and cohesin. They are cell type specific and correlate with the induction of IFNG expression. In sum, these studies indicate that CTCF and cohesin collaborate to establish long-range intrachromosomal interactions that are necessary for regulation of gene expression and can have developmental and environmental plasticity. Although these findings constitute a giant leap in our understanding of the role of cohesins inside the cells, a myriad of questions remain to be answered. For example, how are these linkages established? Does cohesin bind directly to CTCF? Are known cohesin-loading components involved in cohesin binding to these sites? How are the two chromatin regions brought together? Why are loops needed to regulate transcription? Importantly, the implications of these findings extend to more than just finding a novel role for cohesin in interphase, they also greatly contribute to our understanding of the other functions of cohesin. These findings compel us to consider cohesin as a multi-purpose chromatin glue that can be used to hold together two segments of chromatin, regardless of whether these segments belong to the same chromatid or two sister chromatids (Figure 1). It is tempting to speculate that cohesin may also be involved in the long-range interactions between genomic loci on different chromosomes observed during coordinated transcriptional events (Hu et al., 2008). Under this view, the cohesin complex might not only be involved in sister chromatid cohesion or in the formation of intrachromosomal loops involved in transcription, but perhaps the specific chromatin structures formed by cohesin linkages might also be important for other processes, such as DNA replication and repair or the formation of chromatinprotein nuclear bodies. The abundance of cohesin binding sites in interphase and its dramatic decline in mitosis suggest that most of these linkages are removed upon mitotic entry, perhaps to allow chromatin compaction and proper mitotic progression (Yanagida, 2009). Although most cohesin is removed during early mitosis, a small amount of cohesin remains at the centromeres. Strikingly, CTCF has also been shown to localize at the centromeres of mitotic chromosomes (Rubio et al., 2008). It will be interesting to test whether CTCF is involved in the recruitment or maintenance of cohesin at the centromeres and whether these proteins play an architectural role at the centromeres in addition to their role in cohesion. References Dorsett, D. (2009). Cohesin, gene expression and development: lessons from Drosophila. Chromosome Res. 17, 185 –200. Hadjur, S., Williams, L.M., Ryan, N.K., Cobb, B.S., Sexton, T., Fraser, P., Fisher, A.G., and Merkenschlager, M. (2009). Cohesins form chromosomal cis-interactions at the developmentally regulated IFNG locus. Nature 460, 410 – 413. Hu, Q., Kwon, Y.S., Nunez, E., Cardamone, M.D., Hutt, K.R., Ohgi, K.A., Garcia-Bassets, I., Rose, D.W., Glass, C.K., Rosenfeld, M.G., et al. (2008). Enhancing nuclear receptor-induced transcription 60 j Journal of Molecular Cell Biology requires nuclear motor and LSD1-dependent gene networking in interchromatin granules. Proc. Natl Acad. Sci. USA 105, 19199–19204. Parelho, V., Hadjur, S., Spivakov, M., Leleu, M., Sauer, S., Gregson, H.C., Jarmuz, A., Canzonetta, C., Webster, Z., Nesterova, T., et al. (2008). Cohesins functionally associate with CTCF on mammalian chromosome arms. Cell 132, 422–433. Dı´az-Martı´nez and Yu Rubio, E.D., Reiss, D.J., Welcsh, P.L., Disteche, C.M., Filippova, G.N., Baliga, N.S., Aebersold, R., Ranish, J.A., and Krumm, A. (2008). CTCF physically links cohesin to chromatin. Proc. Natl Acad. Sci. USA 105, 8309– 8314. Wendt, K.S., Yoshida, K., Itoh, T., Bando, M., Koch, B., Schirghuber, E., Tsutsumi, S., Nagae, G., Ishihara, K., Mishiro, T., et al. (2008). Cohesin mediates transcriptional insulation by CCCTC-binding factor. Nature 451, 796 –801. Yanagida, M. (2009). Clearing the way for mitosis: is cohesin a target? Nat. Rev. Mol. Cell Biol. 10, 489 –496. Zlatanova, J., and Caiafa, P. (2009). CTCF and its protein partners: divide and rule? J. Cell Sci. 122, 1275 – 1284.