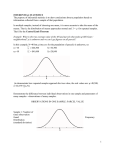
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Biological Conservation 108 (2002) 335–359 www.elsevier.com/locate/biocon The red fox in Australia—an exotic predator turned biocontrol agent J.E. Kinneara,*, N.R. Sumnerb, M.L. Onusa a Department of Conservation and Land Management (CALM), CALMScience Division, Woodvale Research Centre, PO Box 51, Wanneroo, 6065, Western Australia b Western Australian Marine Research Laboratories, Fisheries Department, PO Box 20, North Beach, 6020, Western Australia Received 5 June 2001; received in revised form 5 February 2002; accepted 15 March 2002 Abstract The most important problem regarding mammal conservation in mainland Australia is the low abundance and limited distributions of many species, a legacy of an unprecedented collapse of the mammal fauna on a continental scale that unfolded following European colonisation. Two major hypotheses (not necessarily always mutually exclusive) have been proposed to account for the collapse (1) niche loss–damage due to a variety of causes and (2) predation by exotics, in particular by the red fox. This paper provides evidence the supporting the latter cause as a major factor. Five case studies in Western Australia demonstrate that the fox is an efficient predator that restricts medium-sized marsupials to refugia at low densities. Removal of the fox by baiting typically produces two prey responses (1) significant population recoveries and (2) the colonisation and exploitation of habitats outside of refugia. To date, 11 medium-sized marsupial species, representing seven families, have responded in a like manner. The impact of the fox on its known marsupial prey mimics a biocontrol agent as it severely limits prey distribution and abundance. Niche denial and population suppression characterise it’s actions on a suite of vulnerable species not yet fully documented. # 2002 Elsevier Science Ltd. All rights reserved. Keywords: Marsupial conservation; Fox (Vulpes vulpes); Predation; Predator removal; Biocontrol; Meta-analyses 1. Introduction Essentially, after an invading species becomes established the ecological ‘rules’ or processes which operated in a given system change. . . . the ‘rules’ or processes permitting the coexistence of a species complex are drastically altered leading to extinction of native species’ (Mooney and Drake, 1989, p. 492). During the two centuries that followed the European invasion of Australia, something happened to the ecological processes or ‘rules’ governing Australia’s ecosystems. This conclusion became inevitable after the publication of a paper by Burbidge and McKenzie (1989) on the current status of a major species complex—the mammals. In their pioneering paper, they demonstrated that Australia had suffered more mammal * Corresponding author. E-mail address: [email protected] (J.E. Kinnear). extinctions, more range contractions, and more population declines than any other biogeographical realm. In retrospect therefore, it is difficult to avoid this unfortunate conclusion—the European colonisation of Australia resulted in an unprecedented wildlife catastrophe on a continental scale. It is our view that the cause of the mammal decline will never be universally attributed to any particular factor as causality in ecology is typically complex (see Hilborn and Stearns, 1982), but unquestionably, some factors will be identified as being more significant (proximate) than others. This of course raises the question: what were/are the major factors that made survival impossible for so many mammal species over such vast areas of the Australian landscape? The ensuing search for answers has failed to achieve a consensus as two competing views have emerged, one wide ranging in scope, the other much more specific. Advocates of the former have focused on a list of environmental and biotic factors such as climate changes; habitat loss; habitat damage and fragmentation; 0006-3207/02/$ - see front matter # 2002 Elsevier Science Ltd. All rights reserved. PII: S0006-3207(02)00116-7 336 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 the impact of competitors, for example, rabbits, livestock and feral species; wildfires; disease and predation—the last has been generally judged to be a secondary factor or dismissed entirely (Burbidge and McKenzie, 1989; Johnson et al., 1989; Morton, 1990; Hone, 1999). Other workers have placed a greater emphasis on depredations by introduced species, in particular, by the European red fox (Vulpes vulpes; Hoy, 1923; Wood Jones, 1923–1925; Finlayson, 1961; Christensen, 1980; King et al., 1981; Kinnear et al., 1988, 1998; Friend, 1990; Friend and Thomas, 1994; Short et al., 1994, 2002; Augee et al., 1996; Hobbs and Mooney, 1998). The debate continues (Caughley and Gunn, 1996; Hone, 1999). As a first step, we propose that the primary causes of the mammal decline, both past and present, be viewed within the context of two working hypotheses not necessarily mutually exclusive in every case. The hypotheses are: H1. Niche loss–damage (NLD). H2. Predation by introduced species in particular, by the fox. In terms of niche theory (sensu Hutchinson, 1957), the focus of the NLD hypothesis is primarily on the fundamental niche, while the predation hypothesis is concerned with the realised niche. These distinctions are profound, and indeed, in many cases, the fate of the surviving mammal fauna may be dependent upon their resolution. 1.1. Fundamental vs. realised niches: implications for conservation other species; we show that niche denial exists for a suite of marsupial species surviving in a variety of habitats over a wide geographic area. 1.2. A proposed model We submit that currently, one model broadly applies, at least for a range of medium-sized marsupial species. We submit that the European fox in Australia mimics the role of a multi-specific biocontrol agent affecting a range of marsupial species. Evidence supporting this hypothesis is presented in the following sections— although, we have strong reservations about the longterm stability of many existing relationships. 1.3. Scope of this paper In the following sections, we describe five case studies, undertaken over the period 1980–1996, as a result of a fox control experiment initiated in 1978 on rockwallabies (Kinnear et al., 1988, 1998). The results of this study motivated us to carry out a series of fox removal trials on sites supporting populations of other rare and endangered marsupial species. Each trial is presented as a separate case history with short sections describing Methods, Results and Discussion. Meta-analysis was used to collectively analyse the trials and the paper concludes with a General Discussion that addresses many debated issues pertaining to the predation hypothesis. 2. Methods 2.1. Case studies Factors affecting the fundamental niche of a species are commonly associated with habitat change due to alteration and manipulation of the indigenous landscape, with the result that the landscape may no longer meet the requirements of affected species. In essence, the NLD hypothesis asserts that colonisation has produced a landscape greatly diminished in carrying capacity (zero in many instances), and thus is no longer capable of supporting much of its indigenous mammalian wildlife. The predation hypothesis provides an alternative explanation—niche denial. It asserts that despite the changes to the indigenous landscape, a major cause of the mammal decline has been due to predation. Thus, for many Australian mammals, niches still exist that are capable of supporting flourishing populations, but occupancy has been denied because of depredations by an exotic predator in particular, the red fox. Niche denial by the fox, which mimics actual loss of carrying capacity, first became evident in a study involving rare and endangered rock-wallabies (Kinnear et al., 1988, 1998). In this paper we extend the concept to 2.1.1. Basic design Unless otherwise noted later, the basic design of the case studies consisted of the following procedures. Abundance indices of medium-sized marsupials were collected before the implementation of fox control, and again, after an extended period of fox control. Where circumstances permitted, an unreplicated control site (no fox control) was used. Indices of abundance were obtained by trapping and by repeated spotlight counts using long range, 100-W lights along defined routes. Wire cage traps, typically set at intervals of 100 m, were baited with bread and peanut paste, or a combination of rolled oats, peanut paste and sultanas. Traps were set at dusk and cleared each morning. Trapped animals were fitted with numbered tags in both ears and released at the capture site. On smaller experimental sites ( < 12,000 ha) foxes were controlled by the monthly application of 1080poison meat baits (fresh or dried) or intact fowl eggs using proven methods (sodium monofluoroacetate, 4.5 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 mg; Kinnear et al., 1988, 1998; Wong et al., 1995). A larger mainland site was systematically baited aerially following the methods devised and evaluated by Thompson and Algar (2000) and Thompson et al. (2000). Additional details are described, where relevant, in the appropriate case study. 2.2. Case 1. The Dampier Archipelago: Rothschild’s rock-wallaby The Dampier Archipelago was separated from the mainland ca. 8000 years ago by rising sea levels. It now comprises 42 islands of varying sizes lying within a 45 km radius of the town of Dampier on the northwest coast of Western Australia (Fig. 1). The climate is maritime arid. Rainfall is seasonal (summer), but unreliable with a mean annual fall of 276 mm, and evaporation exceeds rainfall by a factor of nine. The mean maximum temperature is 35 C (Morris, 1990). Rothschild’s rockwallaby (Petrogale rothschildi) is found on three islands, Rosemary (1062 ha), Enderby (3290 ha) and Dolphin (3203 ha; Fig. 1). The first recorded fox bounty payments were paid in 1930 at Roebourne on the nearby mainland (Ride et al., 1964). Foxes were prevalent on the adjacent mainland and they were also present on Dolphin, Angel and 337 Gidley Islands when this study was launched. Feral cats (Felis catus) were present on Dolphin Island and the nearby mainland; the dingo (Canis dingo) is absent from archipelago. 2.2.1. Methods Beginning in 1979–1980, standardised spotlight counts of rock-wallabies on selected areas of Enderby Island (fox free) and Dolphin Island (fox infested) were carried out (Fig. 1). Traverses on foot were made at using a 100-W long-range spotlight powered by a 12 V automotive battery mounted on a modified backpack. Wallabies are readily sighted in these landscapes as the islands largely consist of low hummock grasslands and scree. An aerial baiting program for foxes was initiated (1984) following some exploratory trials (1980–1981) to determine the level of bait acceptance and the risk to non-target species (Table 1). The northern region of Burrup Peninsula was baited to create a buffer zone or sink for foxes. This step was based on the assumption that the dispersal rate of foxes to Dolphin Island would be reduced. Foxes inhabiting the nearby islands, Angel and Gidley (Fig. 1) were poisoned in order to eliminate potential immigrants from these sources. In 1990, the spotlight traverses on Enderby and Dolphin Islands were repeated together with traverses Fig. 1. The Dampier Archipelago: Rothschilds’ rock-wallaby is found on the mainland (at low densities) and on three islands in the archipelago— Enderby, Rosemary and Dolphin. Foxes arrived in the region during the 1930s and subsequently colonised Dolphin, Angel and Gidley Islands. The upward projecting bars next to Enderby (fox free) and Dolphin Islands (fox infested initially, and subsequently, after the implementation of fox control) represent the number of sightings per three hours of transect (see also, Table 2). 338 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Table 1 Details of baiting events (1980–1989) carried out in the Dampier Archipelago (see Fig. 1) Date Method Areas baited Comments October 1980 Manual Dolphin I. 38 beach sectors baited May 1981 Manual Dolphin I.; Burrup Peninsula Most southern beaches on Dolphin; north Burrup beaches September 1984 Aerial Dolphin, Angel, and Gidley Islands; Burrup 2530 baits (1.5 mg 1080) dropped in 200 m grid pattern; 1230 baits laid along tracks on Burrup. October 1987 Aerial Dolphin, Angel, and Gidley Islands; Burrup 2010 baits dropped (4.5 mg 1080) October 1989 Aerial Dolphin, Angel, and Gidley Islands; Burrup 1500 baits on beaches, valleys, mangroves and fresh water pools of selected baited and unbaited areas of the adjacent Burrup Peninsula. 2.2.2. Results: relative abundance: fox free vs. fox infested There were striking differences in rock-wallaby sightings between the fox-free and fox-infested islands (Table 2; Fig. 1). During the 10+ hours of traverses made on each island for the period 1979–1980, 195 rockwallabies were sighted (18.7/h 1.73 SE) on Enderby, while only three wallabies were observed (0.30/h) on Dolphin Island—an average rock-wallaby sighting ratio of 62:1 (fox-free/fox-infested). 2.2.3. Relative abundance: under fox control After a 6-year period of fox control, less intensive (ca. 3 h) spotlight traverses produced these results: on foxfree Enderby, 19.2 1.35 rock-wallaby sightings/h were recorded while on fox-controlled Dolphin, 8.8 0.84/ h—a sighting ratio of approximately 2:1—nearly a 26 fold sighting increase for Dolphin Island. The 1990 Burrup traverses (8 h) produced one wallaby sighting (Table 2, Fig. 1). 2.2.4. Discussion 2.2.4.1. Refugia and niche extensions. On fox-free Enderby and Rosemary Islands, footprints and pads of rock-wallabies were conspicuous on sand-plain areas where they prefer to graze. In contrast on Dolphin Island before fox control, the converse prevailed; signs were non-existent apart from an occasional sighting on rock-piles. Under fox control, not only did the sighting rate increase, but some wallabies were encountered foraging a considerable distance from their rocky shelter. Kinnear et al. (1988, 1998) observed a similar increase in foraging range (i.e. realised niche extension) in response to fox control in the case of the rock-wallaby Petrogale lateralis, which lives in an agricultural setting. One could argue that the Dolphin Island rock-wallaby population was being limited by transient abiotic factors that ceased to operate during the latter course of the study. This argument is not supported by the sightings on nearby Enderby Island, which did not vary. Significantly, there was also no change in rock-wallaby sightings on the adjacent Burrup peninsula in the absence of repeated baitings, which has been shown to be necessary on the mainland in order to sustain the required level of fox control (Fig. 1; Table 2; Kinnear et al., 1988; Thompson et al., 2000). Butler (1983) recorded zero sightings during a comprehensive biological survey of the Burrup Peninsula. The long fox-baiting interval (24–36 months) on Dolphin Island was sufficient to produce a rock-wallaby population recovery because of the presence of a water barrier that presumably served as a deterrent to potential invaders, and the creation of a buffer zone on the adjacent Burrup Peninsula. Buffer zones have since been shown to retard recolonisation by foxes (Thompson et al., 2000). 2.3. Case 2: Tutanning: a wheatbelt nature reserve remnant In the Western Australia (WA) wheatbelt, more than 90% of the original landscape (ca. 15 million ha) has been cleared for agriculture. The region previously supported a diverse mammal fauna with 46 species occurring in the region (Shortridge, 1909). During the 1960s six species of medium sized mammals were present in the Tutanning remnant (2200 ha), and three of these species were shown to be moderately abundant in a study by Sampson (1971). However, during the early 1970s all populations crashed, and by 1978 three species—numbat or banded-anteater (Myrmecobius fasciatus), quenda (Isoodon obesulus, southern brown bandicoot) and western ringtail possum (Pseudocheirus occidentalis)—were no longer resident. Sightings of the woylie (a bettong or rat kangaroo, Bettongia penicillata) ceased and two other species, the tammar wallaby (Macropus eugenii) and the brushtail possum 339 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Table 2 Relative abundance of rock-wallabies on islands in the Dampier Archipelago under the following conditions (1) fox free (2) fox infested (3) fox control (see Fig. 1) Date Traverse time (min) Dolphin Island (fox infested) 1979–1980 census (no fox control) 6 June 1979 170 5 May 1980 82 6 May 1980 85 7 May 1980 135 9 May 1980 130 Totals Mean 602 231 Enderby Island (fox free) 1979–1980 census 4 June 1979 103 5 June 1979 70 6 June 1979 60 7 June 1979 70 5 May 1980 50 12 May 1980 85 13 May 1980 90 14 May 1980 100 628 3 9 12 13 7.2 9.2 10.0 34 1 0 <1 0 27 25 27 23 10 19 28 36 15.7 21.4 27.0 19.7 12.0 13.0 18.7 21.6 195 18.61.73 Enderby Island (fox free) 1990 census 6 June 1990 96 7 June 1990 67 8 June 1990 68 Totals MeanSE 0 0 0.71 0.89 0 8.80.84 Burrup Peninsula (Fox infested) 1990 census 9–11 September 1990 183 (unbaited area) 24–26 October 1990 305 (baited 12 months ago) Totals MeanSE 0 0 1 2 0 Number sighted per hour 0.30 Dolphin Island 1990 census (fox control) 23 June 1990 75 24 June 1990 78 25 June 1990 78 Totals MeanSE Rock-wallabies sighted 231 (Trichosurus vulpecula) were infrequently sighted. This was the situation in 1984 when we initiated a fox control program within Tutanning Nature Reserve. 2.3.1. Methods Prior to the implementation of fox baiting, two trapping sessions for woylies were carried out. The first session (January 1979) was limited to a southeast area previously trapped by Sampson (1971). The second session took place in October–November 1984 prior to the first baiting. On this occasion, we set up trap-lines at sites over a wider area of the reserve. Linear sets of 10 traps (1 km long) placed along tracks at 100 m intervals sampled a range of plant associations throughout the reserve (Fig. 2). Beginning in November 1984 and following the procedures developed by Kinnear et al. 31 24 19 19.4 21.5 16.8 74 19.21.35 (1988, 1998), poison baits were laid monthly at 100 m intervals along all internal tracks and the boundaries of the reserve. In September 1989 after approximately 5 years of fox baiting, trap lines were set up as previously described. When it soon became evident that the woylies had increased substantially, we extended our coverage to other areas of the reserve that we had not previously trapped. The reserve was trapped again in 1992 (Fig. 2). 2.3.2. Results: trapping A summary of the trapping results for woylies is presented in Table 3. Fig. 2 illustrates the trap success rate (percentage) for individual trap lines for the years 1984, 1989 and 1992. 340 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 2.3.3. Spotlight counts Fig. 3 illustrates the trends (1979–1992) and more recent (1998) increases for tammars are also shown (from CALM’s Western Shield database, courtesy of P. Orell). The number of sightings prior to fox control are similar to those recorded during an extensive series of spotlight surveys carried out by Arnold and Steven (1988, personal communication). They recorded all mammal sightings while spotlighting for kangaroos along the reserve boundary. During 50 circuits of the boundary carried out between June 1979 and December 1984, they recorded on average < 2 tammars, and < 1 brushtail possums. Woylies were not recorded. 2.3.4. Discussion The initial trapping effort undertaken in January 1979 covered Sampson’s study area. During his study (1967– 1970), he achieved trap success rates ranging from 10 to 21% for woylie, quenda and brushtail possum. Our effort in 1979 (380 trap nights) yielded a single male woylie, a success rate of 0.26% (Table 3). The period, 1950s–early 1970s, during which a suite of medium-sized marsupials was moderately abundant within Tutanning was a local phenomenon, and moreover, it occurred in the absence of overt fox control. One would be hard pressed therefore, to interpret Sampson’s data as evidence that the fox might be a biocontrol Fig. 2. Trap success rates obtained for woylies at different sites within Tutanning Nature Reserve before the implementation of fox control (1984), and after 5 and 8 years of control. Prior to fox control, woylies were captured at only two sites that were largely comprised of dense thickets of Gastrolobium spp., which served as predation refugia. Trap success rate averaged 4.5% within the two refuges and zero for all other trap lines (overall average 2.17%). During subsequent trapping periods (1989 and 1992), woylies were captured over a very much wider area of the reserve. Table 3 A summary of trapping results (excluding repeats) for Tutanning woylies before fox control, and after the implementation of controla Year Total trap nights No fox control January 1979 October–November 1984 380 322 1 7 0.26 2.17 Fox control commenced November 1984 September 1989 July–August 1992 320 266 64 76 20.00 28.57 a Total captures Trap success (%) During January 1979 trapping was limited to Sampson’s (1971) study area. In 1984, a wider area was trapped in particular, Gastrolobium thickets (see text), which proved to be refuge sites. In 1989 and 1992, an even wider area was sampled (Fig. 2). J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 341 Fig. 3. Spotlighting counts obtained before and after fox control for three marsupials species persisting in Tutanning Nature reserve. (a) Woylies sightings, which prior to fox control, were zero because of a restricted distribution within dense cover. (b) Brush-tail possums were sighted mainly in trees, but following fox control, sightings increased and individuals were frequently sighted on ground with young. (c) Tammar wallaby sightings, which gradually increased from <2 to 40 h1. (The 1998 data courtesy of P. Orell, Wildlife Branch.) agent. However in retrospect, there are some compelling reasons to believe that his study may have been confounded; four reasons are discussed as follows. 1. During the 1950s, a fox bounty payment system was in force (Whitehouse, 1976). Apart from the financial incentive to control foxes, foxes were actively hunted for recreation, and because they were perceived to be a menace to young lambs. 2. The period of fauna abundance coincided with a fire succession stage when the cover was dense, and thus provided some protection from predators (A.R. Main, personal communication). 3. Tutanning carries extensive stands of toxic plants (Gastrolobium spp.). This genus is noted for its ability to synthesise monofluoroacetate (1080). Woylies and brushtail possums are strikingly tolerant to 1080 (King et al., 1981; Twigg and King, 1991) and by inference, this suggests that they consume 1080 plant materials. Indeed, woylies are known to cache toxic seeds of Gastrolobium, and there are anecdotal accounts from early settlers of domestic pigs dying through secondary poisoning after being fed woylies. Thus it is possible that depredations by foxes on toxic victims were contributing to their control. 4. Christensen (1980) has linked certain instances of mammal abundance with broad-scale rabbit control by the WA Agriculture Protection Board (APB). The toxin 1080 was used and it is noted for its impact on non-target species higher in the food chain (secondary poisoning). Algar and Kinnear (1996) described an incident when radio-collared foxes died following rabbit control by the ABP on nearby farmland. McIlroy and Gifford (1991) have also recorded similar findings. During the 1960s, the use of 1080 for rabbit control peaked and fox numbers were low (King et al., 1981). An APB policy change in the early 1970s led to the abandonment of broadscale rabbit control. 342 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 It is likely therefore that the earlier circumstances negated to some extent the impact of the fox within Tutanning. Whatever the cause, this period of abundance was a local, transient phenomenon. By the mid1970s, all species were in decline with three species declining to extinction and moreover, the decline coincided with the cessation of broad-scale rabbit control. This situation has since been reversed by the implementation of fox control; the survivors—woylies, tammars and brushtail possums—have recovered and are now common. The quenda and the numbat have been successfully re-introduced (Friend and Thomas, 1994; personal communication). 2.3.5. The Tutanning Annex The Annex (114 ha) is a remnant block that is separated from the SE sector of the main Tutanning reserve by a farm paddock. Its vegetation is dominated by Allocasuarina spp. commonly known by early settlers as ‘tamma or tammar bush’. 2.3.6. Methods A spotlight survey was carried out in December 1987. Fox baiting was extended to the Annex in November 1989. Follow-up spotlight surveys were made in November 1992 and May 1998 (the latter by P. Orell, Wildlife Branch). 2.3.7. Results and discussion The results (Fig. 4) illustrate a 12–20 fold increase in tammar sightings, most of which were observed feeding nearby in adjacent paddocks. Brushtail possums, previously unsighted, were recorded in significant numbers (Fig. 4). In summary, the Annex results, except for the absence of the woylie, mirrored the results obtained from the nearby main reserve. Tammars were conspicuously more abundant when compared to the main reserve. A public road bisects the main reserve and poaching has been noted. This may have accounted for the difference, as the Annex has no public access. After steps were taken to curtail poaching (L. Silvester, District Wildlife Officer, personal communication), recent (1998) surveys of tammars (by P. Orell, personal communication) have recorded a large increase for the main reserve (Fig. 3). 2.4. Case 3: Boyagin Nature Reserve The 4780 ha Boyagin Nature Reserve (ca. 24 km NW of Pingelly WA) is comprised of two parts (west and east) separated by farmland. Unlike Tutanning’s recent history, there are no records, official or otherwise, indicating that Boyagin enjoyed a flourishing period of mammal abundance. Naturalist H. Butler surveyed the area in 1972 and sighted one tammar; 10 years later, Reserves officer D. Hart trapped intensively and Fig. 4. The Tutanning Annex remnant is separated from the main reserve by a farm paddock. (a) The large increase by tammars following fox control has elevated the species from rarity to pest status locally, as most were observed grazing on farmland. Data for 1998 (P. Orell) were collected at seeding time and hence sightings were largely restricted to the annex where tammars were less visible. (b) Brushtail possum sightings. captured one brushtail possum (CALM Departmental records). In 1985, numbats were released into East Boyagin without fox control, but the translocation failed. However under fox control, a second translocation was successful (Friend, 1990). We initiated a monitoring program on both sections for other medium-sized mammals in 1989 for the purpose of comparing their abundance. When this objective was achieved, baiting was extended to W. Boyagin in late 1989 and we continued to monitor abundance on both sides. 2.4.1. Methods Trapping and spotlight methodology were as described previously. Despite an intensive trapping effort, and after 7 years of fox control on E. Boyagin, and 3 years on W. Boyagin, no woylies were captured. We therefore concluded that woylies were no longer resident on Boyagin. In 1992, 40 woylies, sourced from Dryandra 343 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Woodland, were released on East and West Boyagin (14 females, six males per side). 2.4.2. Results and discussion 2.4.2.1. Trapping results: possums. Fig. 5 displays the trapping results for both East and West Boyagin. There is a marked difference in capture rates between E. Boyagin (baited from 1985) and W. Boyagin (not baited until November 1989). West Boyagin essentially served as a control population for four years (no fox control, 1985–1989). When the trapping results for both sides are compared for this period, the benefits conferred by baiting are clearly evident, as there was a nine-fold difference by 1989 (Fig. 5). 2.4.2.2. Trapping results: woylies. Woylies, which were re-introduced to Boyagin in 1992, were firmly established by 1996 on both sides, but at substantially different densities and this possibly confounded the trapping results for both woylies and possums species (Table 4). Woylie captures predominated on W. Boyagin and possum captures were relatively few, but this was reversed for E. Boyagin. On W. Boyagin 88 woylies were captured over 150 trap nights (59% success rate) while on E. Boyagin 11 woylies and 52 possums were captured. To allow for this competition for traps, we have also elected to calculate the trap success rate for possums by subtracting the woylie captures from the total number of traps set (Table 4). We cannot account for these differences; it could be due to the baiting history, differences in carrying capacity, competition for traps or a combination of these factors. Spotlighting results (1989–1992) for possums and tammars are shown in Fig. 6. With fox control, an increasing trend is evident for possums, as is the case for tammars although the sighting rate is lower. Visibility affects spotlight counts and generally, it is the density of cover that is the major determinant, but other factors may be important. 2.5. Case 4: Dryandra Woodland Dryandra Woodland is a collection of 17 remnants comprising a total area of ca. 28,000 ha (Friend et al., 1995). The main block (ca. 12,000 ha—the largest remnant) was selected as the study site. In 1970–1971, A.A. Burbidge (unpublished report) carried out an intensive trapping and spotlight fauna survey in the main block over a 12 month period. His survey coincided with Sampson’s study at Tutanning, but unlike Sampson’s results, Burbidge’s trapsuccess rates for medium-sized mammals were low namely, brushtail possum 4.0%; woylie 2.7%; quenda 0.35%—the latter not having been trapped since then. Table 4 Woylie trapping data (1996) 4 years after translocation to Boyagin under fox control. Brushtail possum data also listed Trap nights Woylie including repeats Woylie no repeats Possum including repeats Possum no repeats East Boyagin 50 5 50 1 50 3 5 0 3 13 22 17 13 22 14 Totals 9 8 52 49 West Boyagin 50 30 50 29 50 29 30 17 12 6 6 6 6 3 2 Totals 59 18 11 88 Fig. 5. A comparison of trap success rates for brushtail possums on East and West Boyagin under differing fox control regimes. Data for 1996* corrected to allow for trap interference by woylies (see text). 344 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Fig. 6. Spotlight counts for Boyagin Nature Reserve. (a) Brushtail possums. (b) Tammar wallaby. The next trapping exercise made in 1975 failed to capture a single specimen of any of the above species (Christensen, unpublished, departmental files). In September 1982, J.A. Friend commenced a fox baiting trial on a section of the main block of Dryandra (Fig. 7). The purpose of the program was to test the response of the numbat population to fox control (Friend, 1990; Friend and Thomas, 1994). Friend’s baited area encompassed some sites previously trapped by Burbidge and Christensen. This area was trapped again in 1984 along with additional trap lines that were established outside of Friend’s baited area (Fig. 7, Departmental files). At this stage, after 2 years of fox control, it was evident that woylie trap success rate were substantially greater in Friend’s baited area compared to outside of the baited area. We trapped the same areas again in 1989 and shortly afterwards, the whole of the main Dryandra block was baited. Three years later (1992) we carried out a trapping census that sampled the entire main block. 2.5.1. Results and discussion 2.5.1.1. Trapping results: woylies. Fig. 8 illustrates the results for 1984 and 1989, after 2 and 7 years, respectively of localised fox control (trap-lines 1–4, within Friend’s baited area, trap-lines 5–13, outside of the baited area, Fig. 7), and for 1992, after 3 years fox control throughout the entire main block. Woylies responded to fox control in a manner comparable to Boyagin and Tutanning. In the absence of fox control, it was necessary to trap intensively for extended periods in order to detect their presence, and in many areas none were captured. For example, within the area that became Friend’s baiting area, Burbidge captured 21 woylies during 822 trap-nights of effort (2.55% success), while Christensen captured zero woylies during 158 trap-nights. By 1984 after 2 years of fox control, a 50% increase in effort by Christensen produced 36 woylies (15.3% success). In 1989, we caught 75 woylies in Friend’s baited area at a rate comparable with the 1984 effort. Judging from 1989 results, it would appear from our trapping effort that woylie densities had stabilised within Friend’s baited area (ca. 16% in 1984 and 1989). However, by 1992, after the whole of the main block had been baited for 3 years, the trap success rate (Fig. 8) increased to 56%—presumably due to a higher level of fox control achieved by the baiting of the entire main block. Kinnear et al. (1988) found that foxes rapidly reinvaded small baited areas. The results from trap-lines 5–13 (area not baited until 1989; Figs. 7 and 8) situated at varying distances from the perimeter of the baited zone, indicates a zero to low survival rate for woylies not directly afforded the protection provided by baiting. Some protection was indirectly provided for woylies living adjacent to the southern boundary of Friend’s baited area (trap-lines 5– 8, Figs. 7 and 8). In general, the trap success rate for woylies varied inversely with distance from the baited area (exception, trap line 11, Figs. 7 and 8). After 3 years of fox control throughout the main block, woylies extended their realised niche to SE sections of Dryandra where none had been trapped or sighted previously (Fig. 8). The SE section yielded trap success rates in excess of 70% for some trap lines. In 1987, a spotlight traverse through this area (Coelac Road, Fig. 7) produced zero sightings for all species. 2.5.1.2. Spotlighting results: no fox control. During 1971–1972 in addition to trapping, A.A. Burbidge carried out an extensive series of spotlight transects (19 random and 20 fixed routes). An inspection of Burbidge’s random and standardised spotlight data does not reveal any major differences for all species. Thus, a traverse along any route only produced < 1 sighting on average (Table 5). In 1987, our results for unbaited areas of Dryandra were similar (Table 6). Woylie and possum sightings were typically low prior to the baiting of the entire block, but they both increased sharply after baiting of J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 345 Fig. 7. The main block of Dryandra Woodlands is a large (ca. 12,000 ha) wheatbelt native remnant. This figure displays the location of 13 woylie trap-lines, and ‘Friend’s baited area’ (the enclosed area; Friend, 1990), which represents an area selectively baited from 1982 to 1989. The whole of Dryandra has been baited from 1989 onwards. the entire block (Table 6). Tammar sightings were infrequent and variable and to date have remained so (P. Mawson, personal communication). 2.6. Case 5: Fitzgerald River National Park Fitzgerald River National Park (FRNP) is located on the south coast of WA between Bremer Bay and Hopetoun (Fig. 9). The park (329,000 ha) is approximately 27 times larger than Dryandra’s main block (Chapman and Newbey, 1995). Apart from wildfires, it has a history that has been relatively free of disturbance. Chapman and Newbey (1995, p. 93) list 26 mammal species that were known to be present in the Fitzgerald area prior to European settlement. Many of these species were also part of a now greatly fragmented wheatbelt fauna, and both regions have experienced a similar level of extinctions (> 40%). Survival of medium sized mammals has been typically poor, as only four spp of a known 11 spp (> 150 g) were recently recorded by Chapman and Newbey. Moreover, the surviving species were not recorded as being abundant or widely distributed. For example, Chapman and Newbey trapped only one quenda, sighted only two tammars and eleven brush wallabies (Macropus irma; uncommon and restricted). Brushtail possums were recorded as being present along watercourses (numbers not stated). 2.6.1. Objectives The objectives of the FRNP study were threefold: 1. To extend fox control to a much larger unfragmented area in a more mesic biogeographical region of the state. 2. To determine if two aerial baitings per year were sufficient to significantly lower and maintain fox numbers over a large unfragmented area of conservation estate. 3. To assess if this level of baiting produced a positive population response by resident mediumsized mammals. 2.6.2. Methods Trap lines extending for 4 km (40 traps set at 100-m intervals) were set out along vehicle tracks so as to sample the range of plant associations namely heath, woodland, riparian and mallee within the study areas. Spotlight traverses likewise sampled the earlier associations but covered a larger area. 346 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Fig. 8. Dryandra Woodlands showing the trapping results for woylies obtained for the years 1984, 1989, and 1992. Initially, woylie increases were largely confined to Friend’s baited area (baited from 1982) and some nearby trap sites (Fig. 7). Trap success rates declined to zero at the more distant sites. The whole of Dryandra was baited in 1989 and by 1992, woylies had colonised most of the reserve at high densities. Table 5 Counts (numbers h1) of marsupial species observed along fixed spotlight transects before fox control, within Friend’s baited area (Fig. 7), and after fox control throughout the main block Possum sightings (h1) Tammar sightings (h1) 1.68 2.32 2.0 0.57 0.50 0.54 Localized fox control from 1982 (Friend’s baited area) December 1987 (5.03) 2.58 December 1989 (8.12) 13.3 1.19 2.71 0 0.37 Fox control (1989) extended to whole of Dryandra main block September 1990 (4.5) 15.7 November 1992 (5.74) 23 6.67 9.9 4.67 1.23 Year (total hours) No fox controla 1970 Random routes (31.6) 1971 Fixed routes (43.8) Means/h a Woylie sightings (h1) 0.60 0.27 0.41 Data provided by A.A. Burbidge. 2.6.2.1. Baiting protocol. The FRNP was partitioned on a north–south axis into approximately two equal parts (Fig. 9). Beginning in February 1991 and ending September 1995, the western part was aerially baited twice yearly during spring and autumn. Baits consisted of dried meat containing 4.5 mg, 1080 and were distributed at a density of approximately six baits per km2. This protocol has been shown to be effective method for reducing fox densities (Thompson and Algar, 2000; Thompson et al., 2000). The timing of the baitings was designed to minimise recruitment by exposing foxes to poison baits during the reproductive and dispersal phases of their life cycle (spring/autumn, respectively). Supplementary baitings J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 (December 1995 and February–March 1996) were made from a vehicle at the study sites on the western side after aerial baitings ceased in September 1995. 2.6.2.2. Assessing the level of fox control. We used the cyanide technique to estimate the effectiveness of the baiting regime (Algar and Kinnear, 1992). This Table 6 Counts (numbers h1) of marsupial species observed along fixed spotlight transects excluding Friend’s baited area (Fig. 7) Year (total hours) Possum Tammar Woylie (numbers h1) (numbers h1) (numbers h1) No fox control except for Friend’s baited area December 1987 (2.43 h) 1.65 2.88 December 1987 (1.88 h) 0.53 2.66 Mean 1.16 2.78 0.00 0.00 0.00 Whole of Dryandra main block baited 1989 September 1989 (2.55 h) 5.49 3.92 September 1990 (1.10 h) 2.73 6.36 September 1990 (1.08 h) 2.78 6.48 September 1990 (1.02 h) 4.72 5.88 0.00 0.00 0.00 0.00 After 3 years of fox control of entire main block November 1992 (2.42 h) 27.69 22.31 0.00 347 technique generates an abundance index—the number of foxes sampled per 20 km transects. During the first year of fox control, cyanide transects were carried out on both the baited and unbaited sides of the park 2–4 weeks after each baiting (February 1991, November 1991). Cyanide transects were repeated on both sides 5– 6 months after the March 1993 baiting, but just prior to the next baiting (September). 2.6.3. Results and discussion 2.6.3.1. Effectiveness of the baiting protocol. The relative impact of the aerial baitings on fox numbers is indicated in the sample sizes of the cyanide transects for the unbaited and baited areas (Fig. 10). On the baited side, the cyanide index declined by an average of 80% for the 1991 trials, and by 75% for the 1993 trial. The latter outcome signified that recruitment by foxes was negligible between spring and autumn baiting events. These results are comparable with more comprehensive studies on bait uptake and fox mortality (Thompson and Algar, 2000; Thompson et al., 2000). In a trial involving 45 radio-tagged foxes, Thompson et al. (2000; Fig. 2) showed that following an aerial baiting, more than 70% of the tagged foxes were dead within 10–15 days and all were dead after 44 days. Fig. 9. Fitzgerald River National Park: The park (ca. 329,000 ha) was divided approximately into two parts and the western side was aerially baited twice yearly beginning in February 1991. Two similar study sites were selected on each side of the park, and medium-sized mammals were monitored until 1996. 348 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Fig. 10. The western side of the park (Fig. 9) was baited at approximately 6-month intervals. The effectiveness of the baitings was assessed by cyanide baitings (Algar and Kinnear, 1992), a sampling technique that generates an index of fox abundance—number of foxes sampled by cyanide baits/20 km. In 1991, we sampled the fox populations on both sides 2–4 weeks after each baiting. Fig. 10 indicates that the baitings reduced the fox population by approximately 75–80%. In 1993, we carried out an autumn baiting, and we sampled the fox populations again approximately 6 months later, just prior to the next (spring) baiting. The purpose of this sampling was to determine if the fox population had recovered during the 6-month interval between baitings. The results indicate that there was no net recruitment, whereas on the unbaited side, foxes had recovered to their original density. 2.6.3.2. Possum population responses: trapping. Possums were present on the unbaited eastern side, but 604 trap nights over a 2-year period yielded only a single possum in woodland habitat. In contrast, 600 trap nights yielded 48 possums on the baited side, and significantly, the capture rate increased exponentially (Fig. 11; 1998 data courtesy P. Collins). 2.6.3.3. Spotlighting. Spotlighting produced 123 possum sightings within the baited area, but only 12 sightings within the unbaited area (Table 7). An upward trend in tammar sightings from the baited study sites is also evident (Table 7). No tammars were sighted in the unbaited area. 2.6.4. Postscript: an update Since the conclusion of this study in 1996, fox control has continued at a higher level (4 times year1) under the auspices of Western Shield a statewide exotic predator control program (Bailey, 1996). CALM regional staff have continued to monitor mammals within our western study area. A brief summary of their trapping results follows (information provided by P. Collins). Possums have continued to increase (Fig. 11) to the extent that they are considered to be a hindrance as they dominate the capture statistics to the exclusion of other potential subjects. Quenda (bandicoots) are being captured routinely. Brush wallabies are now commonly Fig. 11. A comparison of brushtail possum trap success rates in Fitzgerald River National Park—unbaited versus baited (see text for more recent results). sighted over wide areas. The carnivorous marsupial, Dasyurus geoffroii, (a quoll) has been captured for the first time, and there has been a spate of captures of the very rare and endangered Dibbler (Parantechinus apicalis). 3. Statistical methodology: meta-analysis Meta-analysis was first used for studies in the social sciences (see Hedges and Olkin, 1985; Cooper and Hedges, 1994, among others). It has also been applied in ecology (Gurevitch and Hedges, 1993; Gurevitch et al., 1992). This type of analysis is suited to a situation where a researcher(s) wishes to combine the results of several, possibly contradicting, separate studies in order to reach a general overall conclusion or summarise the results of past research. The data from the foregoing trapping and spotlight trials will be analysed separately using meta-analysis to test whether fox control, as achieved by using poison baits, increases the abundance of medium sized native marsupials. It maybe one of the first times this technique has been used on wildlife survey data. Additionally, bootstrapping will be used to determine the distribution of the effect sizes from different studies. 3.1. Methods Small numbers of observations from separate studies were combined to enable a meaningful analysis. The data for each study consisted of one or more measures directly related to the abundance of medium-sized native marsupials both before and after the commencement of a fox control program. For each study all survey data before fox control commenced were combined to obtain the best indicator of abundance. However, after the commencement of the fox control program at a 349 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 site a gradual increase in abundance was expected. For this reason only data from surveys conducted more than 2 years after baiting commenced were combined to allow time for a response to occur. All studied sites in Western Australia where adequate data for both with and without fox control (treatment and control) was available were used for the analysis. However, two species of native marsupials had become locally extinct at certain sites: the tammar at Dryandra (along the selected spotlight routes) and woylie at Boyagin. Since no response was possible, these species at these sites were excluded from the analysis. 3.1.2. Spotlight surveys For the spotlight transects (Table 9), the number of animals spotted per hour was used. When data from two or more surveys for a treatment were available, they were combined by adding the counts from each survey and re-computing the rate of sightings (equivalent to the mean weighted by the duration of the survey in hours). The standardised difference (di) was used to represent the effect size 3.1.1. Trapping surveys For the trapping surveys (Table 8), the proportion of successes expressed as the number of animals caught divided by the number of traps set was used. When data from two or more surveys were available, the total number of animals trapped divided by the total number of traps set (trap nights) was used. For these data the variance was estimated from the binomial distribution. where X ti is the weighted mean of the treatment group (with fox control) for the ith study, X ci is the weighted mean of the control group (prior to fox control) and si is the pooled standard deviation for the two groups. The conditional variance of di was estimated as di ¼ vi ¼ X ti X ci si ð1Þ nti þ nci di2 þ t c t n i ni 2 ni þ nci ð2Þ where nti is the within-study sample size in the treatment group (survey duration in hours) and nci the withinstudy sample size in the control group of the ith study. Table 7 Fitzgerald River National Park: a comparison of spotlight counts (numbers h1) of brush-tail possums and tammars along fixed routes in the unbaited eastern section and the baited western section Year (total hours) Eastern section—no fox control December 1994 (2.9) November 1995 (6.5) Possums sightings (h1) Tammars sightings (h1) 1.72 1.08 0.00 0.00 Western section—fox control from 1991 April–November 1993 (5.2) 2.13 December 1994 (6.1) 4.41 November 1995 (2.38) 19.3 February–May 1996 (3.4) 11.5 1.37 1.63 5.47 10.6 3.2. Meta-analysis A homogeneity test statistic (Shadish and Haddock, 1994) was used to test whether the studies shared a common population effect size. If test statistic Q exceeds the upper-tail critical value of the chi-square distribution at k1 degrees of freedom, where k is the number of studies, the observed variance in the effect sizes is significantly greater than expected for studies that share a common effect size. When Q is rejected the heterogeneity may be accounted for by using a random effects model rather than fixed effects model. Under a random effects model the effect size is random with its own distribution. Table 8 Meta-analysis trapping data: random effects models—proportions Site Dryandra 1971–1972 Dryandra Trapline 7 Dryandra Trapline 8 Dryandra Trapline 9 Dryandra Trapline 10 Dryandra Trapline 11 Dryandra Trapline 12 Dryandra Trapline 13 Tutanning Boyagin FRNP Species Woylie Woylie Woylie Woylie Woylie Woylie Woylie Woylie Woylie Possum Possum No fox control Fox control Nights1 Captures1 Nights2 Captures2 980 120 40 39 40 40 40 40 702 696 604 21 10 3 2 1 0 0 0 8 4 1 772 16 20 20 20 24 16 16 586 844 600 146 4 6 7 9 6 4 3 140 149 48 350 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Table 9 Meta-analysis spotlight data: random effects models—mean differences Site Dryandra Route 2 Dryandra Route 2 Tutanning Tutanning Tutanning Tutanning Annex Tutanning Annex Boyagin Boyagin Dolphin Island FRNP FRNP Species Woylie Possum Woylie Possum Tammar Possum Tammar Possum Tammar Rock-wallaby Possum Tammar Control Treatment Si Hours1 Rate1 Hours2 Rate2 4.98 4.98 23.34 23.34 23.34 1.38 1.38 11.18 11.18 10.02 9.37 9.37 3.61 3.41 0 0.39 0.47 0 5.07 0.27 0.09 0.33 1.28 0 2.42 2.42 34.94 34.94 34.94 2.91 2.91 19.00 19.00 3.85 7.41 7.41 27.69 22.31 1.00 4.75 5.01 7.90 77.66 5.95 1.32 8.83 5.05 2.66 The variance component for the random effects model was estimated in two ways (Shadish and Haddock, 1994). Firstly, the ordinary (unweighted) sample estimate of the variance of the effect sizes was used to estimate the variance component. Secondly, the weighted sample estimate of the variance of the effect sizes was used to estimate the variance component. The remaining calculations for both the trapping and spotlight surveys follow those described by Shadish and Haddock (1994). A Mathcad worksheet (Mathsoft, 1995), available from the authors, was set up to perform the computations. A one-tailed t-test was used to test the null hypothesis that the population effect size is zero (i.e. fox control does not increase the abundance of medium sized native marsupials) against the alternative hypothesis that, the mean effect size is greater than zero (fox control increases the abundance of medium sized native marsupials). The test statistic was calculated using both the ordinary (unweighted) sample estimate of the variance of the effect sizes, and the weighted sample estimate of the variance of the effect sizes to estimate the variance component. 3.3. Bootstrapping The weighted effect sizes were bootstrapped to test the same null hypothesis against the alternative hypothesis. The bootstrapping approach does not require the same assumptions of normality as the t-test and will perform better when the data are skewed. When plotted, the bootstrapped values provide valuable information on the distribution of the effect sizes. 3.4. Results 3.4.1. Trapping surveys The homogeneity test statistic for the fixed effects model Q=73.3 exceeded the upper tail critical value of chi-square at 10 degrees of freedom (P< 0.001) indicating 2.15 0.58 0.21 1.43 2.18 2.54 25.74 2.06 0.33 0.86 1.52 1.73 the heterogeneity could not be explained by sampling error alone, if studies all shared a common population effect size. A random effects model was required to take proper account of the heterogeneity between studies. Both test statistics calculated using the ordinary (unweighted) sample estimate of the variance of the effect sizes and the weighted sample estimate of the variance of the effect sizes to estimate the variance component were highly significant (P < 0.001). The bootstrapping result was also highly significant since the bootstrapped values did not include zero. The null hypothesis was rejected in favour of the alternative hypothesis: fox control increases the abundance of medium-sized native marsupials. 3.4.2. Spotlight surveys The homogeneity test statistic for the fixed effects model Q=60.0 exceeded the upper tail critical value of chi-square at 11 degrees of freedom (P < 0.001) indicating the heterogeneity could not be explained by sampling error alone if studies all shared a common population effect size. A random effects model was required to take proper account of the heterogeneity between studies. The test statistic calculated using the ordinary (unweighted) sample estimate of the variance of the effect sizes was significant (P < 0.05), whereas, the test statistic calculated using the weighted sample estimate of the variance was highly significant (P < 0.001). The bootstrapping result was also highly significant since the bootstrapped values did not include zero. The null hypothesis was rejected in favour of the alternative hypothesis: fox control increases the abundance of medium sized native marsupials. 3.5. Discussion It was not surprising that random effects models were required to take proper account of the heterogeneity J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 between studies. The variation in the response between studies was large due to different species, sites and differing number of years between the commencement of fox control and when the surveys were conducted. Random effects models were required to properly account for this variation. For many of the spotlight surveys there was barely sufficient observations to estimate the pooled standard deviation, at least two observations both before and after fox control were needed. However, for Dryandra it was only possible to estimate the standard deviation before fox control commenced since there was only one subsequent observation. It is likely that the standard deviation for this site was underestimated, and in turn responsible for a large value for the standardised mean difference. The standard deviation for Tutanning Annex could only be estimated for the observations after fox control commenced for the same reason. 351 4. In the absence of fox control, prey species persist at low densities within sites that serve as predation refugia. However, because the population densities are very low within refugia, it is unlikely that such populations will be stable over the long-term (see later). 5. Removal of foxes from refuge sites not only increases native prey populations, but also enables prey species to occupy and utilise habitats not exploited in the presence of foxes—an example of niche denial followed by niche expansion. 6. In general, at sites still carrying multiple species of medium-sized mammals, fox control produced a positive population response by all species. 7. To date, the suppression of prey species by the fox has not yet affected reproductive fitness (due to bottlenecking) as indicated by the positive population responses of endangered prey. 3.6. Conclusions When all studies are considered together, baiting to control foxes has a large positive effect on the abundance of medium sized native marsupials. For the spotlight studies the effect sizes ranged from 1.5 to 32.6, these are considered to be very large (Cohen, 1969) indicating a dramatic response in the abundance of native marsupials to fox control. The tammar population within Dryandra Woodland and Boyagin Nature Reserve did not respond positively to fox control. Possible reasons are: visibility restrictions due to dense cover; carrying capacity intrinsically low for tammar; NLD; and poaching. Tammars have responded positively elsewhere (i.e. Tutanning and FRNP; see also Tables 10 and 11) and in some cases, this species has locally achieved ‘pest’ status. 4.1. An update from Western Shield 4. General discussion To have generality, results should be consistent under a wide variety of circumstances (Johnson, 1999). The rationale for the case studies described in this paper originated from fox removal experiments from sites that were replicated along with controls (Kinnear et al., 1988, 1998)—a hypothetical-deductive (H-D) design (Beck, 1997). It is significant that all of the case studies produced results similar to the rock-wallaby study, i.e. fox control produces significant population increases of rare and endangered marsupials. Additionally, these outcomes have extended our understanding of the impact of fox predation in the following ways: 1. The fox is a very efficient predator of mediumsized marsupials (see later for further discussion regarding predator efficiency). 2. Suppression of marsupial populations by the fox affects an array of species over a wide geographic range and in a variety of habitats. 3. Suppression of native prey by foxes was common to both small (2200 ha) and large areas (329,000 ha) of conservation estate. Under Western Shield, a wide area management response to the fox threat, the number of sites subject fox control has increased, and likewise for species translocations. A central database exists and is being maintained under the auspices of CALM’s Wildlife Branch (P. Orell, personal communication). Some results as of 1999 are as follows: baiting has produced positive population responses at 25 sites comprising a total of 11 species and representing seven marsupial families. In addition, seven species belonging to six families have been successfully translocated to 15 sites where they formerly occurred (Tables 10 and 11). Clearly, these data indicate that the spectrum of Western Australian marsupial prey species suppressed by the fox is broad, and on this basis we make this following generalisation namely that: the European fox should be recognised as a multi-species biological control agent affecting a subset of Australian mammals—the full range yet to be documented. This generalisation is based on an initial H-D experiment and a series of case histories (this paper and other cases; see Table 10), the latter methodology being one that has been championed by Shrader-Frechette and McCoy (1993). In their critique of ecological 352 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Table 10 A list of marsupial species that have persisted in predation refuges at low densities, and have since increased and expanded their use of the habitat following ongoing fox baiting programs Species Number of sites Reference source Notes and comments Family Macropodidae—Wallabies Black-flanked rock-wallaby (Petrogale lateralis) Six sites Kinnear et al. (1998) and unpublished works Population increases at five wheatbelt sites. Population increase within Cape Range National Park (Kinnear, 1995) Rothschild’s rock-wallaby (Petrogale rothschildi) One site This paper Dolphin Island, Dampier Archipelago (See text) Yellow-footed rock-wallaby (Petrogale xanthopus) Numerous sites P. Alexander, and P. Copley, Dept. Environ . . . S. Australia Operation ‘‘Bounceback’’—a wide-area feral animal control program in the Flinders and Olary Ranges involving fox control Tammar wallaby (Macropus eugenii) Five sites This paper; WSa database Tutanning NR; Tutanning annex; FRNP; Perup and Kingston forest (near Manjimup) This paper Wheatbelt remnants: Dryandra and Tutanning; Perup and Kingston forests (Manjimup) Sinclair et al. (1996) Last recorded in the 1870s and believed to be extinct. Nil captures during survey of Two Peoples Bay Nature Reserve in 1970s; rediscovered in 1994 after the implementation of fox control program Family Pseudocheiridae—Ringtail Possums Western ringtail possum Two sites (Pseudocheirus occidentalis) WS database Perup and Kingston forests (near Manjimup) Family Phalangeraidae—Brushtail Possums Western brushtail possum Seven sites (Trichosurus vulpecula) This paper; WS database Wheatbelt remnants: Tutanning, Boyagin, Lake Magenta; FRNP, South Coast; Batalling (near Collie), Perup and Kingston forests; (Manjimup) Family Perameloidae—Bandicoots and Bilbies Quenda or Southern brown Six sites bandicoot (Isoodon obesulus) C. Pentland (Hon. Thesis); WS database Twin Swamps and Ellenbrook (Perth suburb); L. Magenta (wheatbelt); Perup and Kingston (Manjimup); Waychinnicup NR (near Albany) Family Myrmecobiidae—The numbat or banded ant-eater Numbat (Myrmecobius fasciatus) Two sites Friend (1990) Wheatbelt; Dryandra; Perup (Manjimup forest site) Family Potoroidae—Potoroos and Bettongs (Rat Kangaroos) Woylie or Brush-tail bettong, Four sites or rat kangaroo (Bettongia penicillata) Gilberts’ potoroo (Potorous gilbertii) One site. Two peoples Bay, south coast of WA Family Dasyuridae—Quolls or native cats and other carnivorous marsupials Red-tail phascogale (Phascogale calura) Three sites Friend (unpublished) Chuditch or Western quoll (Dasyurus geoffroii) a Four sites Morris et al. (1995) ‘Western Shield’ database (courtesy of P. Orell, Wildlife Branch). Wheatbelt remnants. Fox removal produced increases on baited sites, but not on unbaited sites Batalling forrest; Hills forest; Noggerup; St. John forest J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 353 Table 11 A list of marsupial species that have been successfully translocated to sites where the species formerly occurreda Species Number of sites Notes and comments Family Macropodidae—Wallabies Black-flanked rock-wallaby (Petrogale lateralis) One site Querekin rock population extinction occurred in late 1980s in the absence of fox control (Kinnear et al., 1998). Five animals sourced from Nangeen Hill translocated to Querekin in 1990 after implementation of fox control. In 1998, 45 individuals trapped Tammar wallaby (Macropus eugenii) One site Batalling forest. Translocations to five other sites, but too early to make an assessment Family Potoroidae—Potoroos and Bettongs (Rat Kangaroos) Woylie or Brush-tail rat kangaroo (Bettongia penicillata) Seven sites Most sites within Jarrah forest: Batalling, St. John forest, Hills Forest, Julimar. Wheatbelt: Boyagin. Recently to the semi-arid sites Lake Magenta and Francois Peron National Park; and both appear to be successful. Source populations; Dryandra, Perup Boodie (Bettongia lesueur) One site Heirisson Prong (Shark Bay, a CSIRO Project, Short, 1999). Animals sourced from fox-free island Family Pseudocheiridae—Ringtail Possums Western ringtail possum (Pseudocheirus occidentalis) One site Leschenault Peninsula: animals sourced from Busselton area either from wildlife carers or rescued from residential development sites Family Perameloidae—Bandicoots and Bilbies Western Barred Bandicoot (Perameles bougainville) One site Heirisson Prong (Shark Bay, a CSIRO Project) animals from fox-free island source Quenda or Southern brown bandicoot (Isoodon obesulus) Four sites Tutanning, Dongolocking, Boyagin Nature Reserves and Mt. Barker. Animals sourced mainly from sites due for development Family Myrmecobiidae—The numbat or banded ant-eater Numbat (Myrmecobius fasciatus) Three sites Dragon Rocks, Boyagin and Tutanning Nature Reserves. Population recoveries of numbats, a diurnally active species, can be limited by feral cats and native predators eg, Chuditch (a quoll or native cat) and raptors as is the case in Batalling forest and Karroun Hill Family Dasyuridae—Quolls or native cats and other carnivorous marsupials Chuditch or Western quoll (Dasyurus geoffroii) Four sites a Julimar forest; L. Magenta; Cape Arid; Mt Lindesay. An initial translocation to a forest site (Lane Poole, 1987) failed in the absence of fox control All sites subject to ongoing fox baiting programs. Data from Western Shield database (courtesy of P. Orell, Wildlife Branch). methodology, they argue for a case history approach, as they believe that an excessive reliance on H-D experimentation is responsible for the dearth of valid generalisations in ecology. 4.2. Biological control agents: essential propertiesattributes In theory, the ideal biocontrol system (as envisaged for economic pests) is a system where (1) the pest population is driven to extinction or if not, it is maintained at below threshold densities at which damage is tolerable (2) the controlling agent is host specific (3) the agent can increase rapidly in response to host increases and (4) the controlling agent persists even though prey densities are maintained at low densities (Murdoch, 1990, 1992, 1994; Murdoch and Briggs, 1996). Parasitoids tend to meet these specifications. Generalist predators can be very effective, but they have undesirable properties, as they are typically not targetspecific—a caveat confirmed by the list of marsupial victims suppressed by the fox (Table 10). Indeed, the fox would be an example of biological control gone utterly wrong, a disastrous worst-case scenario. 354 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 4.2.1. The fox as a biocontrol agent As a biocontrol agent, the fox is not lacking in appropriate attributes: it has a high searching ability; it is a mobile, broad-niche species as indicated by its rapid spread and current distribution (King and Smith, 1985; Coman, 1995; Saunders et al., 1995). Furthermore, it is not tightly coupled to any particular prey, as it is a dietary generalist par excellence because of its carnivore–omnivore–scavenger tendencies—the latter making it vulnerable to control by baiting. It is a highly skilled predator and indeed, it is even claimed to be capable of regulating rabbit populations (Newsome et al., 1989; Pech et al., 1992), but see Boutin (1995). It has been documented as a surplus killer of marsupials (Kinnear et al., 1998; Short et al. 2002). In drought affected habitats it may become locally extinct, but because of its mobility it can readily re-invade. All of these characteristics make the fox an effective biocontrol agent that affords little respite to vulnerable fauna lacking a history of co-evolution (Short et al., 2002). 4.2.2. Models and biocontrol agents In a series of papers modelling predator–prey systems, Sinclair, Pech and colleagues (1998 and references therein) have produced a number of models describing a range of possible outcomes. A predator cum biocontrol agent would fall within the ambit of their models. We suggest that two models would apply namely: (1) the predator limits prey populations to low densities and maintains them as such in refuges, and (2) the predator drives prey populations to extinction. Model 1 fits the observed refuge populations described in this paper, but in our view, the question of stability would remain suspect as the system may well revert to a model 2 in the long term (see Sections 4.9, 4.10 for examples). Translocations of prey to non-refuge sites without fox control would fit model 2; such outcomes have occurred (e.g. numbats at Boyagin; woylies within the jarrah forest, Christensen, unpublished). 4.2.3. Missing Australian marsupial pests Environmental disturbances can affect different species in different ways; some species may decline as a result whereas others may prosper because of new favourable circumstances and thus achieve pest status. In parts of Australia, kangaroos have become so, but medium-sized marsupial pests are practically unknown over wide areas of mainland Australia. Currently, in Australia, one has to look to fox-free Tasmania and Kangaroo Island to find examples. Both of these habitats have historically experienced an environmental impact comparable to the mainland following settlement. On Kangaroo island tammars are culled by shooting; in fox-free Tasmania, 1080 is used to control medium-sized marsupials. In contrast, on mainland WA, apart from kangaroos, one needs to use 1080 to control foxes in order to produce a marsupial pest. Tutanning tammar wallabies and wheatbelt rock-wallabies are examples (P. Orell, personal communication). This outcome is consistent with the view that the fox is acting as a biocontrol agent. 4.3. Possible confounding of the results due to feral cats and the dingo The impact of the dingo can be discounted because it was absent from all of the study sites (see also Short et al., 2002 for more detailed discussion regarding the dingo). The feral cat was present at all sites. Its predation ecology has been reviewed by Dickman (1996a, b), but the available evidence about its impact on medium-sized mammals presents a confusing picture. Christensen and Burrows (1994) failed in an attempt to re-introduce two marsupials species at a desert site because of cat predation. The fox and the dingo were effectively controlled, but control of the feral cat was not successful. Conversely, feral cats are present in fox/dingo-free Tasmania yet the fauna, including the barred bandicoot (Perameles gunnii), is largely intact. This species, a likely prey item, is still relatively widespread and abundant in Tasmania (Mallick et al., 1997, 1998), but critically endangered on the Australian mainland (Seebeck, 1995). Cats have been present on Garden and Rottnest—two small islands near Perth, WA—dating from the colonial period, but despite this long association, two wallabies, the tammar and the quokka (Setonix brachyurus), continue to thrive despite their presence. However this pattern does not hold for other island situations, as the cat is linked to the extinction of a hare wallaby, a bettong and a bandicoot (Burbidge, 1971). It can be argued that, given the results of the case studies, the data may have been confounded by the removal of feral cats as well as foxes. However this is highly unlikely, as numerous trials have revealed that meat baits are ineffective in controlling feral cats (Christensen and Burrows, 1994; unpublished trials). Thus we can reasonably conclude that the treatment was selective for foxes. 4.4. Other possible limiting factors The five case studies consistently produced results (as has CALM’s Western Shield) that conformed to the following pattern namely: fox control, as achieved by baiting, typically results in significant population increases and niche expansions by medium-sized marsupials (see Section 3.2). These results eliminate NLD as a factor—that is, lack of carrying capacity—and likewise, reproductive fitness. Alternative explanations and interpretations are nonetheless possible. Any other explanation needs to J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 satisfy these questions. (1) Given the low population densities of every experimental population prior to fox control, why so few marsupials? (2) Why did they all increase after the implementation of baiting programs? Some possible biotic factors previously listed will be considered (see Section 1). These include factors such as pathogens (including parasites) and competitors, for example, rabbits, domestic stock and feral species. Pathogens will be considered first. On considering the diversity of sites, the diversity of species, and the geographic scale of the experiments, then on epidemiological grounds it seems improbable that every species, at every site, were limited by a pathogen(s). If pathogens were involved, then it seems equally as improbable that all pathogens abruptly ceased to be limiting following baiting. Nor it is likely that the applied treatment (meat baits) provided a cure. None of the sites carried domestic stock or alien feral species. All sites carried rabbits apart from the Dampier Archipelago sites, which are rabbit-free. No attempts were made to control rabbits at any site. Given that foxes are alleged to limit rabbit populations (Pech et al., 1992), then it is possible that baiting could have enabled rabbits to increase. No pattern is discernable, as marsupials increased in the presence of rabbits and also, when they were absent. Abiotic environmental factors include climatic factors such as drought, extreme temperatures or events that adversely affect survival and reproduction. Again, given the geographic scale of this study, it also seems unlikely that, prior to fox control, these limiting factors were operative in every case over such a wide geographic range—such is the nature of the WA climate. It also seems improbable that a simultaneous relaxation of these factors, leading to conditions favouring population growth, would have coincided with the commencement of baiting at every site. Furthermore, four of the five sites—Dryandra, Boyagin, Tutanning and the FRNP—are situated in a productive agricultural region, which does not experience extended periods of adverse climatic conditions. Population crashes of prey species can occur in this environment, but the environmental stress would be of short duration, hence population recoveries would follow relatively quickly. This was not the case, because all of these sites had a long history of low abundance preceding the trials—apart from Tutanning, which has been already discussed (see descriptions of these sites and their historical backgrounds). Finally, purists of the H-D school will object to the experimental design (Beck, 1997, but see critique of HD by Shrader-Frechette and McCoy, 1993). Nevertheless, it should be recalled that the baiting trials were not devoid of control sites. With the exception of Tutanning, every site had some areas that served as untreated controls—i.e. no baiting. It is noteworthy 355 that, no population recoveries occurred at any of these untreated sites. In summary, when the evidence is considered in its entirety, the weight of the evidence consistently supports a strong case for excluding many other potentially limiting factors. 4.5. The fox as an efficient predator of marsupials Biocontrol agents/predators will extinguish their prey when the predation rate exceeds the prey growth rate over all prey densities. This outcome may be avoided if prey have access to a refuge (Rosenzweig and MacArthur, 1963), or if the predator switches to an alternative prey when prey become rare. Significantly however, Short et al. (2002) found that the expected switch did not occur in a case involving predation by foxes on a population of rat-kangaroos (Bettongia lesueur). After reducing the bettong population to a low density, foxes continued to depredate bettongs in the presence a very high density rabbit population (Oryctolagus cuniculus). A predator or biocontrol agent is judged to be efficient (and therefore, its prey vulnerable) if it is capable of limiting its prey populations to low densities (Rosenzweig and MacArthur, 1963; Rosenzweig, 1969). Predator efficiency and prey vulnerability are therefore, relative attributes. A predator may be efficient in relation to a particular prey species but not so in relation to other species. As prey species, many Western Australian marsupials are vulnerable (e.g. in WA to date, 11 spp), partly because they are not particularly adapted to cope with high depredation rates due to (1) their intrinsically low fecundity (especially macropodines e.g. wallabies) (2) low population growth rates, and (3) inappropriate anti-predator defenses and life histories (relative to the fox) due to their Gondwana heritage. 4.6. Gondawana anti-predator defenses: transferable and sufficient? Co-evolved anti-predator defenses (and lifestyles) by indigenous prey in response to their native predators do not necessarily protect prey from the depredations of a recently introduced exotic species. Hanski et al. (2001) argue that the impact of predators has been under-rated because of a failure to acknowledge ecological differences amongst predators. Krefft’s (1866, p. 20) description’s of a hare-wallaby’s shelter (Lagochestes sp., prior to the colonisation by the fox) is indicative. It reads as follows to quote . . . ‘‘(It) is generally found asleep under some saltbush or any other sheltered locality.’’ Apparently, most any shady site would do even in the presence of its native predators, which is in sharp contrast to existing habitats—for example, rock piles, dense cover, trees and burrows—in which surviving marsupial species are found today. 356 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 Hare-wallabies like most medium-sized marsupials are now either extinct or rare over much of their former ranges, as are most of the arid zone mediumsized marsupials. Rock-wallabies are another case. Numerous species have managed to thrive in fragmented rocky outcrops in the presence of their natural predators for thousands of years—a fragmented lifestyle that allegedly, is suppose to make them more vulnerable to predators (see Section 4.7). However, following the arrival of the fox in WA, widespread extinctions have occurred, and extinctions are still being recorded (Pearson, 1992; Kinnear et al., 1998). From these observations, two inferences emerge concerning fragmentation (1) a fragmented lifestyle did not prevent rock-wallabies from establishing thriving populations in the presence of their natural predators, and more significantly (2) their anti-predator defences, while sufficient to cope with native predators, have since proven to be hopelessly inadequate in relation to the fox (Pearson, 1992; Kinnear et al., 1998). A review of the evidence regarding the vulnerability of marsupials to fox depredations can be found in Short et al. (2002). 4.7. Predation and habitat fragmentation It is generally believed that populations surviving in small remnants (high perimeter-to-area ratio) are more vulnerable to predation than those persisting in large tracts of suitable habitat. The evidence supporting this argument is largely based on studies of avian nest predation (Reese and Ratti, 1988). Brown and Litvaitis (1995) failed to demonstrate increases in foraging efficiencies by foxes and coyotes on cottontail rabbits (Sylvilagus transitionalis) in a fragmented landscape. Reese and Ratti (1988) concluded that the evidence is limited, and that the situation is more complex than a simple area effect. It is also noteworthy that the mammals in the large Fitzgerald River National Park (329,000 ha) fared no better than those in the extensively fragmented areas of the WA wheatbelt (Chapman and Newbey, 1995). 4.8. Habitat disturbance and predation The argument linking of enhanced prey vulnerability with habitat fragmentation has a parallel in regards to habitat disturbance. It has been argued that closed forest habitats in their relatively undisturbed state exclude foxes (Jarman, 1986). Road and track construction are alleged to diminish the effectiveness of such habitats as these constructs are suppose to facilitate access by foxes thus making native species more vulnerable. Meek and Saunders (2000) believe this perception needs to reassessed, because they radio-tracked foxes living and foraging within dense forest and heaths in the absence of roads. Using radio-tagged foxes, Algar (unpublished) recorded a similar finding in Kalbarri National Park in WA where the loss of medium-sized mammals has been almost total. Dolphin Island has remained essentially ‘‘pristine’’ (no history of permanent human settlement, agriculture, mining etc.); the FRNP has a similar history, yet medium-sized mammals of these sites have been impacted just as severely as highly disturbed areas. Clearly these perceptions need to be re-assessed. As shown in the case studies, the fox appears to be a very efficient predator of many medium-sized marsupials under a wide variety of environmental circumstances. 4.9. NLD and the significance of translocations More evidence about the efficiency of the fox and NLD can be deduced from successful translocation experiments. The woylie translocation to Boyagin was successful as was the Numbat translocation (Friend, 1990; see list in Table 11). Kinnear et al. (1998) recorded the extinction of a small rock-wallaby population at a site not subject to fox control (Querekin). Subsequently five wallabies were translocated to the site from a nearby colony, and the founders under fox control have since increased to 45 (Eldridge et al., 2001). Four points arise from these translocation examples: (1) existing former habitats still retain the capacity to meet the requirements of the earlier species (niches intact, little or no NLD) (2) in the absence of fox control, the sites were unable to maintain populations of the earlier species (carrying capacity zero in the presence of the fox) (3) the founding populations were not lacking in reproductive fitness despite a history of bootlenecks and (4) one should not assume that sites serving as refugia are secure (Pearson, 1992; Kinnear et al., 1998). 4.10. Marsupial refugia: are they safe havens? Rock-piles, trees, and dense vegetative cover (often containing 1080 poison plants in WA; Christensen, 1980; King et al., 1981) are associated with marsupial survival and thus, serve as predation refugia. Refugia become evident whenever remnant populations expand and establish viable populations in the surrounding habitat following the implementation of fox control. The observed habitat colonisations outside of refugia (realised niche expansions) by the woylie in Tutanning and Dryandra illustrate this point (Figs. 2 and 8; see also Kinnear et al., 1998). In WA, there are reasons to suspect that, in the absence of fox control, the available refugia are not particularly secure principally because prey densities are so low. Furthermore, Pearson (1992) has provided evidence of instability in the case of rock-wallaby populations living on arid zone refuge sites. During 1988–1990, he visited 80 sites historically known to have carried wallabies (as recently as 1940), and he found that 83% J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 of these populations were now extinct. Kinnear et al. (1998) recorded a rock-wallaby population extinction in a more mesic environment. These findings are not encouraging for it is evidence that refugia are not necessarily a guarantee of population stability. Wildlife agencies should take note; a risk-averse policy is advisable, for it seems unwise to assume that known refuge populations are secure. 4.11. Relevance of predation models The utility of predation models is limited because of the difficulties encountered in testing them. In the cases involving cryptic predator–prey relationships, as for example, secretive nocturnal species (the Australian situation), the testing of would be a formidable task fraught with difficulties. In essence, the task consists of determining who is killing whom at any given prey density. One needs to be able to record every individual predation event (who and whom) over a range of prey densities. Technological advances are needed. One model, of considerable interest, involves multiple prey population states. The model predicts that under predation, prey may exist in either of two stable states, a low density (predator pit) state or a higher density state. The predator is supposed to regulate its prey at low densities, but at high densities it becomes inefficient. The latter high density state is of interest, because fox control would be not required if prey were sufficiently numerous (see also, Pech et al., 1992). This model could be crudely tested by creating a high density prey population followed by the withdrawal of fox control. It would be a test of the efficiency of the fox as a predator over a range of marsupial prey densities. We suspect that the prey would crash for reasons described in Short et al. (2002). The Tutanning history is an example. Nevertheless, it should be done because of the implications for fox management, and also because it would advance our understanding of the ecology of the fox in Australia. 5. Concluding remarks The most important problem regarding mammal conservation in Australia is the low abundance and limited distributions of many species (Burbidge and McKenzie, 1989; Smith and Quin, 1996). Advocates of the nicheloss-damage hypothesis have compiled a list of abiotic and biotic interactions allegedly responsible for the presumed loss of carrying capacity of the landscape. Accordingly therefore, the restoration of the landscape to a more pristine state is one of the keys to mammal conservation. As advocates of the view that predation by the fox is also a major threatening process, we do not dispute the 357 fact that the carrying capacity of the Australian landscape in relation to mammals (and other spp) has been affected in an absolute sense. What we seek to emphasise is that predation by the fox can mimic loss of carrying capacity through niche denial. When a species is classified as rare and endangered, then the task facing conservation biologists is to identify the limiting factors (sensu Krebs, 1995) affecting such populations. Predation in WA is such a factor, and we see no reason why the fox factor should not be widespread. It does raise a matter that may also be commonplace, but has not yet been investigated. NLD and fox predation may be jointly proximate—that is, both habitat restoration and predator control (including feral cats in some circumstances), may be needed to produce a positive population response by an endangered species. In some situations niche denial may well confound and distort perceptions about species habitats, as it confines surviving species to refugia, a situation that can create the belief that refugia represent preferred or essential habitats, when in fact, they are survival niches. It seems likely that these ecological distortions caused by exotic predators have confounded attempts to achieve an understanding of the factors threatening survival of endangered species. Moreover, it is also likely that, this confusion has influenced research, policy-making, and wildlife management. In view of these conclusions, how this invader is perceived is vitally important—indeed, critical for the survival and future of many threatened species. To argue that the fox in Australia is a typical predator coexisting with its prey in a dynamic equilibrium is to belie its impact (Hone, 1999). One might reasonably classify it as a keystone species that qualifies as a ‘diversity-reducing’ predator (sensu Schoener and Spiller, 1996), which it certainly appears to be. It is our proposition that the impact of the fox would be more fully appreciated, if it were recognised for what it really is—an exotic predator, pre-adapted to assume the role of a biocontrol agent. Both of these interpretations emphasise the need to identify and to the assess risk to all species of wildlife posed by the fox. Additionally, there is an urgent need to devise a long-term method of fox control that ideally, would be self-sustaining, because 1080-baiting, effective as it is, is at best an essential holding action. The current search for a biocontrol method (Tyndale-Biscoe, 1994) must be supported and maintained. In the meantime the use of 1080 must be safeguarded until a biological solution is found. Acknowledgements We are especially grateful to P. Orell and P. Mawson of CALM’s Wildlife Branch, for their assistance in compiling Tables 10 and 11. P. Orell and P. Collins 358 J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 kindly provided recent census data that enabled us to update the situation in Tutanning Nature Reserve and FRNP respectively. A.A. Burbidge and P. Christensen generously provided important historical data for Dryandra Woodland. I. Abbott reviewed an earlier draft and made numerous helpful comments and suggestions. Professors A.R. Main and A. Kinnear provided much thoughtful advice and encouragement. An anonymous referee provided much constructive criticism. References Algar, D., Kinnear, J.E., 1992. Cyanide baiting to sample fox populations and measure changes in relative abundance. In: O’Brien, P., Berry, G. (Eds.), Wildlife Rabies Contingency Planning in Australia. Bureau of Rural Resources Proceedings No. 11, AGPS, Canberra, pp. 135–138. Algar, D., Kinnear, J.E., 1996. Secondary poisoning of foxes following a routine 1080 rabbit-baiting campaign in the Western Australian wheatbelt. CALMScience 2, 149–152. Arnold, G.W., Steven, D.E., 1988. Variations in distribution of western grey kangaroos, Macropus fuliginosus ocydromus, in Tutanning Nature Reserve and their impact of adjacent farmland. Australian Wildlife Research 15, 119–128. Augee, M.L., Smith, B., Rose, S., 1996. Survival of wild and handreared Ringtail possums (Pseudocheirus peregrinus) in bushland near Sydney. Wildlife Research 23, 99–108. Bailey, C., 1996. Western Shield: bringing wildlife back from the brink of extinction. Landscope 11 (4), 41–48. Beck, M.W., 1997. Inference and generality in ecology: current problems and an experimental solution. Oikos 78, 265–273. Boutin, S., 1995. Testing predator–prey theory by studying fluctuating populations of small mammals. Wildlife Research 22, 89–100. Brown, A.L., Lituaitis, J.A., 1995. Habitat features associated with predation of New England cottontails: what scale is appropiate? Canadian Journal of Zoology 73, 1005–1011. Burbidge, A.A., 1971. The Flora and Fauna of the Monte Bello Islands (Department of Fisheries and Fauna Report 9, pp. 1–19). Perth, Western Australia. Burbidge, A.A., McKenzie, N.L., 1989. Patterns in the modern decline of Western Australia’s vertebrate fauna: causes and conservation implications. Biological Conservation 50, 143–198. Butler, W.H., 1983. Burrup Peninsula Fauna Survey. Woodside Offshore Petroleum Pty., Ltd, Perth. Caughley, G., Gunn, P., 1996. Conservation Biology in Theory and Practice. Blackwell Science, Cambridge, MA. Chapman, A., Newbey, K.R., 1995. A biological survey of the Fitzgerald Area, Western Australia. CALMScience Supplement 3, 1–258. Christensen, P.E.S., 1980. A sad day for native fauna. Forest Focus 23, 3–12. Christensen, P., Burrows, N., 1994. Project desert dreaming: experimental reintroduction of mammals to the Gibson Desert, Western Australia. In: Serena, M. (Ed.), Reintroduction Biology of Australian and New Zealand Fauna. Surrey Beatty & Sons, Chipping Norton, pp. 199–207. Cohen, J.E., 1969. Statistical Power Analysis for the Behavioral Sciences. Acedemic Press, New York. Coman, B.J., 1995. Fox (Vulpes vulpes). In: Strahan, R. (Ed.), The Mammals of Australia. Reed Books, Chatswood, NSW, pp. 698–699. Cooper, H., Hedges, L.V., 1994. The Handbook of Research Synthesis. Russell Sage Foundation, New York. Dickman, C.R., 1996a. Impact of exotic generalist predators on the native fauna of Australia. Wildlife Biology 2, 185–195. Dickman, C.R., 1996b. Overview of the impacts of feral cats on Australian native fauna. Australian Nature Conservation Agency, Canberra. Eldridge, M.D.B., Kinnear, J.E., Onus, M.L., 2001. Source population of dispersing rock-wallabies (Petrogale lateralis) identified by assignment tests on multilocus genotypic data. Molecular Ecology 10, 2867–2876. Finlayson, H.H., 1961. On Central Australian mammals. Part IV. Records of the South Australian Museum 14, 141–191. Friend, J.A., 1990. The numbat Myrmecobius fasciatus (Myrmecobidae): history of decline and potential for recovery. Proceedings of the Ecological Society Australia 16, 369–377. Friend, J.A., Thomas, N.D., 1994. Reintroduction and the numbat recovery programme. In: Serena, M. (Ed.), Reintroduction Biology of Australian and New Zealand Fauna. Surrey Beatty & Sons, Chipping Norton, pp. 189–198. Friend, T., Bowra, T., Gorton, S., Hilder, D., Mitchell, D., Moncrieff, D., Sutton, A., 1995. Dryandra Woodland Management Plan 1995– 2005. Management Plan No. 30. Department of Conservation and Land Management, Crawley, Western Australia. Gurevitch, J., Hedges, L.V., 1993. Meta-analysis: combining the results of independent studies in experimental ecology. In: Scheiner, S., Gurevitch, J. (Eds.), Design and Analysis of Ecological Experiments. Chapman and Hall, New York, pp. 378– 398. Gurevitch, J., Morrow, L.L., Wallace, A., Walsh, J.S., 1992. A metaanalysis of competition in field experiments. American Naturalist 140, 539–572. Hanski, I., Henttonen, H., Korpimaki, E., Oksanen, L., Turchin, P., 2001. Small-rodent dynamics and predation. Ecology 82, 1505– 1520. Hedges, L.V., Olkin, I., 1985. Statistical Methods for Meta-Analysis. Academic Press, New York. Hilborn, R., Stearns, S.C., 1982. On inference in ecology and evolutionary biology: the problem of multiple causes. Acta Biotheoretica 31, 145–164. Hobbs, R.J., Mooney, H.A., 1998. Broadening the extinction debate: population deletions and additions in California and Western Australia. Conservation Biology 12, 271–283. Hone, J., 1999. Fox control and rock-wallaby population dynamics— assumptions and hypotheses. Wildlife Research 26, 671–673. Hoy, C.M., 1923. The present status of the Australian mammal fauna. Journal of Mammalogy 4, 164–166. Hutchinson, G.E., 1957. Concluding remarks. Cold Spring Harbour Symposium for Quantitative Biology 22, 415–427. Jarman, P., 1986. The red fox—an exotic large predator. In: Kitching, R.L. (Ed.), The Ecology of Exotic Animals and Plants. John Wiley and Sons, Brisbane, pp. 43–61. Johnson, D.H., 1999. The insignificance of statistical significance testing. Journal of Wildlife Management 63, 763–772. Johnson, K.A., Burbidge, A.A., McKenzie, N.L., 1989. Australian Macropodoidea: status, causes of decline and future research and management. In: Grigg, G., Jarman, P., Hume, I. (Eds.), Kangaroos, Wallabies and Rat-Kangaroos. Surrey Beatty & Sons, NSW, pp. 641–657. King, D.R., Oliver, A.J., Mead, R.J., 1981. Bettongia and fluoroacetate: a role for 1080 in fauna management. Australian Wildlife Research 8, 529–536. King, D.R., Smith, L.A., 1985. The distribution of the European red fox (Vulpes vulpes) in Western Australia. Records of the Western Australian Museum 12, 197–205. Kinnear, J.E., 1995. Rock wallabies of Yardie Creek. Landscope 11 (2), 36–37. Kinnear, J.E., Onus, M.L., Bromilow, R.N., 1988. Fox control and rock-wallaby population dynamics. Australian Wildlife Research 15, 435–450. Kinnear, J.E., Onus, M.L., Sumner, N.R., 1998. Fox control and J.E. Kinnear et al. / Biological Conservation 108 (2002) 335–359 rock-wallaby population dynamics—II. An update. Wildlife Research 25, 81–88. Krebs, C.J., 1995. Two paradigms of population regulation. Wildlife Research 22, 1–10. Krefft, G., 1866. On the vertebrate animals of the Lower Murray and Darling, their habits, economy, and geographical distribution. Transactions of Philosophic Society of New South Wales 1862- 65, 1–33. Mallick, S.A., Haseler, M., Hocking, G.J., Driessen, M.M., 1997. Past and present distribution of the eastern barred bandicoot Perameles gunnii in the midlands, Tasmania. Pacific Conservation Biology 3, 397–402. Mallick, S.A., Hocking, G.J., Driessen, M.M., 1998. Road-kills of the eastern barred bandicoot (Perameles gunnii) in Tasmania: an index of abundance. Wildlife Research 25, 139–145. Mathsoft, 1995. Mathcad User’s Manual Mathcad Plus 6.0. Mathsoft Inc, Cambridge, MA, USA. McIlroy, J.C., Gifford, E.J., 1991. Effects on non-target animal populations of a rabbit trial-baiting campaign with 1080 poison. Wildlife Research 18, 315–325. Meek, P.D., Saunders, G., 2000. Home range and movement of foxes (Vulpes vulpes) in coastal New South Wales, Australia. Wildlife Research 27, 663–668. Mooney, H.A., Drake, J.A., 1989. Biological invasions: a scope program overview. In: Drake, J.A., Mooney, H.A. (Eds.), Biological Invasions: a Global Perspective. John Wiley & Sons, Chichester, pp. 491–506. Morris, K., 1990. Dampier Archipelago Nature Reserves: Management Plan No. 18. Department of Conservation and Land Management, Crawley, Western Australia. Morris, K., Orell, P., Brazell, R., 1995. The effect of fox control on native mammals in the jarrah forest, Western Australia. In: Proceedings 10th Australian Vertebrate Pest Control Conference (Hobart), Tasmanian Department of Primary Industry and Fisheries, PO Box 46, King Meadows, 7249. Morton, S.R., 1990. The impact of European settlement on the vertebrate animals of arid Australia: a conceptual model. Proceedings of the Ecological Society of Australia 16, 201–213. Murdoch, W.W., 1990. The relevance of pest-enemy models to biological control. In: Mackauer, M., Ehler, L., Roland, J. (Eds.), Critical Issues in Biological Control. Intercept Ltd, Andover, Hants, UK, pp. 1–24. Murdoch, W.W., 1992. Ecological theory and biological control. In: Jain, S.K., Botsford, L.W. (Eds.), Applied Population Biology. Kluwer Academic Publishers, Boston, MA, pp. 197–221. Murdoch, W.W., 1994. Population regulation in theory and practice. Ecology 75, 271–287. Murdoch, W.W., Briggs, C.J., 1996. Theory for biological control: recent developments. Ecology 77, 2001–2013. Newsome, A.E., Parer, I., Catling, P.C., 1989. Prolonged prey suppression by carnivores—predator-removal experiments. Oecologia 78, 458–467. Pearson, D.J., 1992. Past and present distribution and abundance of the black-footed rock-wallaby in the Warburton region of Western Australia. Wildlife Research 19, 605–622. Pech, R.P., Sinclair, A.R.E., Newsome, A.E., Catling, P.C., 1992. Limits to predator regulation of rabbits in Australia: evidence from predator-removal experiments. Oecologia 89, 102–112. Reese, K.P., Ratti, J.T., 1988. Edge effect: a concept under scrutiny. Transactions of the 53rd North American Wildlife and Natural Resources Conference 53, 127–135. Ride, W.D.L., Crawford, I.M., Storr, G.M., Berndt, R.M., Royce, R.D., 1964. Report on the aboriginal engravings and flora and fauna of Depuch Island. Special Publication of the Western Australian Museum 2, 11–88. 359 Rosenzweig, M.L., 1969. Why the prey curve has a hump. American Naturalist 103, 81–87. Rosenzweig, M.L., MacArthur, R.H., 1963. Graphical representation and stability conditions of predator–prey interactions. American Naturalist 97, 209–223. Sampson, J.C., 1971. The Biology of Bettongia penicillata Gray, 1837. PhD Thesis, The University of Western Australia. Saunders, G., Coman, B.J., Kinnear, J., Braysher, M., 1995. Managing vertebrate pests: foxes. Australian Government Publishing Service, Canberra. Seebeck, J.H., 1995. Eastern Barred Bandicoot Perameles gunnii. In: Strahan, R. (Ed.), The Mammals of Australia. Reed Books, Chatswood, NSW, pp. 182–183. Schoener, T.W., Spiller, D.A., 1996. Devastation of prey diversity by experimentally introduced predators in the field. Nature 381, 691–694. Shadish, W.R., Haddock, C.K., 1994. Combining estimates of effect size. In: Cooper, H., Hedges, L.V. (Eds.), The Handbook of Research Synthesis. Russell Sage Foundation, New York, pp. 261–281. Short, J., 1999. Decline and Recovery of Australian Mammals with Particular Emphasis on the Burrowing Bettong Bettongia lesueur. PhD Thesis, Murdoch University, Perth, Western Australia. Short, J., Kinnear, J.E., Robley, A., 2002. Surplus killing by introduced predators in Australia—evidence for ineffective anti-predator adaptations in native prey species. Biological Conservation 103 (3), 283–301. Short, J., Turner, B., Parker, S., Twiss, J., 1994. Reintroduction of endangered mammals to mainland Shark Bay: a progress report. In: Serena, M. (Ed.), Reintroduction Biology of Australian and New Zealand Fauna. Surrey Beatty & Sons, Chipping, pp. 183–188. Shortridge, G.C., 1909. An account of the geographical distribution of the marsupials and monotremes of S.W. Australia, having special reference to the specimens collected during the Balston Expedition 1904–1907. Proceedings of the Zoological Society London 1909, 803–848. Shrader-Frechette, K.S., McCoy, E.D., 1993. Method in Ecology: Strategies for Conservation. Cambridge University Press, Cambridge. Sinclair, E.A., Danks, A., Wayne, A.F., 1996. Rediscovery of Gilbert’s Potoroo Potorous tridactylus in Western Australia. Australian Mammalogy 19, 69–72. Sinclair, A.R.E., Pech, R.P., Dickman, C.R., Hik, D., Mahon, P., Newsome, A.E., 1998. Predicting effects of predation on conservation of endangered prey. Conservation Biology 12, 564–575. Smith, A.P., Quin, D.G., 1996. Patterns and causes of extinction and decline in Australian conilurine rodents. Biological Conservation 77, 243–267. Thompson, P.C., Algar, D., 2000. The uptake of dried meat baits by foxes and investigations of baiting rates in Western Australia. Wildlife Research 27, 451–456. Thompson, P.C., Marlow, N.J., Rose, K., Kok, N.E., 2000. The effectiveness of a large-scale baiting campaign and an evaluation of a buffer zone strategy for fox control in Western Australia. Wildlife Research 27, 465–472. Twigg, L.E., King, D.R., 1991. The impact of fluoroacetate-bearing vegetation on Native Australian fauna: a review. Oikos 61, 412–430. Tyndale-Biscoe, C.H., 1994. Virus-vectored immunocontraception of feral mammals. Reproduction, Fertility and Development 6, 281–287. Whitehouse, S., 1976. Bounty systems in vermin control. Journal of Agriculture of Western Australia 17 (3), 1–5. Wong, D.H., Kinnear, J.E., Runham, C.F., 1995. A simple rapid bioassay for compound 1080 (sodium flouroacetate) in bait materials and soil—its technique and applications. Australian Wildlife Research 22, 561–568. Wood Jones, F., 1923–1925. The mammals of South Australia parts I– III. Government Printer, Adelaide.