Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
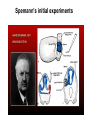
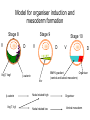
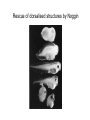
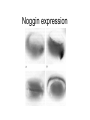
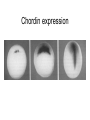
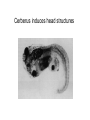
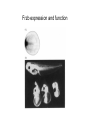
Axis determination and early development in amphibians We are standing and walking with parts of our body which could have been used for thinking had they developed in another part of the body (Hans Spemann 1943) Spemann’s initial experiments Spemann’s organiser • The term "Organizer" was coined to emphasize the ability of this dorsal blastopore lip tissue to direct the development of the host tissue and to give these redirected cells a coherent, unified, organization. "A piece of the upper blastoporal lip of an amphibian embryo undergoing gastrulation exerts an organizing effect on its environment in such a way that, if transplanted to an indifferent region of another embryo, it causes there the formation of a secondary embryonic anlage. Such a piece can therefore be designated as an organizer." This tissue had the ability to invaginate and differentiate autonomously, to induce the neural plate and, by assimilative induction, to organize somites from the lateral plate mesoderm of the host. The functions of the organiser • The ability to self-differentiate dorasl mesoderm • The ability to dorsalise the surrounding mesoderm into paraxial (somite-forming) mesoderm (when it would otherwise form ventral mesoderm) • The ability to dorsalise the ectoderm, inducing the formation of the neural tube • The ability to initiate the movements of gastrulation Molecular events forming the Organiser sperm Dsh Dsh V D No -catenin in nuclei Cortical rotation V GSK Dsh D GSK -catenin degraded -catenin stable -catenin in nuclei WNT pathway Transcriptional events V D -catenin degraded Tcf3 proteins -catenin stabilised (nuclear) Siamois gene Repressed Input from TGF- (Vg1, Nodal) signalling pathway Tcf3 proteins+ -catenin Activated Siamois protein goosecoid gene Activated Dorsal axis Ability of goosecoid mRNA to induce a new D-V axis • An embryo (16-cell stage) whose ventral vegital pole was injected with goosecoid mRNA induced a second axis (with two complete sets of head structures precursors). This did not happen when embryos were injected with a mutant form of the DNA-binding homeodomain (Niehrs et al, 1993 Cell 72: 491-503) Model for organiser induction and mesoderm formation Stage 8 V Stage 9 D VegT, Veg1 V D -catenin Xnr -catenin VegT, Vg1 Stage 10 Nodal related high Nodal related low V D BMP4 gradient Organiser (ventral and lateral mesoderm) Organiser Ventral mesoderm The Diffusible proteins of the Organiser I: BMP inhibitors • Noggin: diffusible protein accomplishing two of the major functions of the organiser: induction of dorsal ectoderm to form neural tube and formation of dorsal mesoderm. Binds to BMP4 and BMP2 and inhibits their binding to receptors (Zimmerman et al 1996 Cell 86, 599-606). • • • Chordin: diffusible protein directing central nervous system development. Binds to and inhibits BMP4 (Sasai et al 1994 Cell 79, 779-790). Nodal-related protein 3 (Xnr-3): diffusible protein synthesized by the superficial cells of the organiser and is also able to block BMP4 (Smith et al 1995 Cell 82, 37-46). Follistatin: secreted glycoprotein that induces epidermal cell fate (skin) overriding the default neural fate (Hemmati-Brinvalou and Melton 1994 Cell 88, 273-281). Rescue of dorsalised structures by Noggin Noggin expression Chordin expression Bone Morphogenetic Factor 4 (BMP4): an Organiser antagonist • The Xenopus BMP4 is a member of the TGF family of ligands. It induces ventral mesoderm and (ventral) epidermal fates and suppresses neural ones. Blood cells kidneys muscle BMP4 relative concentration The Diffusible proteins of the Organiser II: Wnt inhibitors • • Cerberus responsible for the anterior-most head structures; cytokine of the cysteine knot superfamily binds Xwnt8 (member of the Wnt family) as well as BMPs and Nodal-related (Bouwmeester et al 1996 Nature 382, 595-601). FRZB and Dickkopf Wnt receptor antagonists prevent signalling from Wnt receptors. Frzb is able to bind Wnt agonists and by binding to Frizzled proteins prevent the interaction between ligand and receptor (Wang et al, 1997 Cell 88, 757-766; Glinka et al, 1998 Nature 391, 357-362). Cerberus induces head structures Frzb expression and function Frzb binds to Xwnt8 Dorsal Pharyngeal endoderm Prechordal plate mesoderm Notochordal mesoderm Cerberus Dickkopf Frzb Chordin Noggin Anterior Follistatin Posterior Xwnt8 Nodal-related Ventral BMP4 Posterior transforming proteins: Wnt signals and retinoic acid In Xenopus embryos a gradient of Wnt signalling (Xwnt8) and -catenin is highest in the posterior and absent in the anterior. There is also a high concentration of retinoic acid (RA) at the posterior end of the neural tube. RA is especially important in patterning the hindbrain but not the forebrain (Dupé and Lumsden 2001 Development 128, 2199-2208) The anterior transforming proteins: Insulin-like growth factors In addition to proteins that block BMP4 and Wnt signalling in the head there is a positive signal that promotes anterior head development. Insulin-like growth factors (IGFs) are required to form the anterior-neural tube with its brain and sensory placodes. (Pera et al Dev Cell 1, 655-665) Summary Anterior O R G A Dorsal N I S E R IGFs Ventral BMP4 RA Wnt Posterior