Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
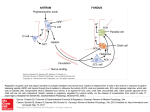
LECTURE 2 GASTROINTESTINAL HORMONES Gastrin The functions of gastrin are coordinated to promote hydrogen ion (H+) secretion by the gastric parietal cells. Gastrin, a 17-amino acid straight chain peptide, is secreted by G (gastrin) cells in the antrum of the stomach. The 17-amino acid form of gastrin, which is called G17 or "little" gastrin, is the form of gastrin secreted in response to a meal. A 34-amino acid form of gastrin, which is called G34 or "big" gastrin, is secreted during the interdigestive period (between meals). It is secreted at low basal levels. G34 is not a dimer of G17, nor is G17 formed from G34. Rather, each form of gastrin has its own biosynthetic pathway, beginning with its own precursor, a progastrin molecule. The minimum fragment necessary for biologic activity of gastrin is the C-terminal tetrapeptide. Although the C-terminal tetrapeptide is the minimum fragment necessary for activity, it still is only one-sixth as active as the entire gastrin molecule. Secretion of gastrin In response to eating a meal, gastrin (G17) is secreted from G cells located in the antrum of the stomach. The physiologic stimuli that initiate gastrin secretion all are related to ingestion of food. These stimuli include: the products of protein digestion (Among the products of protein digestion, the amino acids phenylalanine and tryptophan are the most potent stimuli for gastrin secretion). distention of the stomach by food vagal stimulation (Local vagal reflexes also stimulate gastrin secretion. In these local reflexes, the neurocrine released from vagal nerve endings onto the G cells is gastrin-releasing peptide (GRP, bombesin)). In addition to these positive stimuli, gastrin secretion is inhibited by a low pH of the gastric contents and by somatostatin. Actions of gastrin. Gastrin has two major actions: 1. It stimulates H+ secretion by gastric parietal cells, and 2. It stimulates growth of the gastric mucosa, a trophic effect. The physiologic actions of gastrin are nicely illustrated in conditions of gastrin excess or deficiency. For example, in persons with gastrin-secreting tumors (Zollinger-Ellison syndrome), H+ secretion is increased, and the trophic effect of gastrin causes the gastric mucosa to hypertrophy. Conversely, in persons whose gastric antrum is resected (which removes the source of gastrin, the G cells), H+ secretion is decreased, and the gastric mucosa atrophies. Zollinger-Ellison syndrome is caused by a gastrinsecreting tumor or gastrinoma, usually in the non-β-cell pancreas. The signs and symptoms of Zollinger-Ellison syndrome all are attributable to high circulating levels of gastrin: increased H+ secretion by parietal cells, hypertrophy of the gastric mucosa, and duodenal ulcers caused by the unrelenting secretion of H+. Steatorrhea, the increased H+ secretion also results in acidification of the intestinal lumen, which inactivates pancreatic lipase, an enzyme necessary for fat digestion. As a result, dietary fats are not adequately digested or absorbed, and fat is excreted in the stool (steatorrhea). Treatment of Zollinger-Ellison syndrome includes: administration of H2 receptor-blocking drugs (e.g., Cimetidine & Ranitidine); protone pump inhibitors (e.g., Omeprazole); removal of the tumor; the last resort is gastric resection, which removes gastrin's target tissue. Cholecystokinin The functions of cholecystokinin (CCK) are coordinated to promote fat digestion and absorption. CCK is a 33-amino acid peptide, which is structurally related to gastrin. The C-terminal five amino acids (CCK5) are identical to those of gastrin and include the tetrapeptide that is minimally necessary for gastrin activity. Thus, CCK has some gastrin activity. The minimum fragment of CCK necessary for its biologic activity is the C-terminal heptapeptide [CCK-7]. CCK is secreted by the I cells of the duodenal and jejunal mucosa in response to two types of physiologic stimuli: (1) monoglycerides and fatty acids (but not triglycerides), and (2) small peptides and amino acids. These stimuli alert the I cells to the presence of a meal containing fat and protein, which must be digested and absorbed. CCK will then ensure that appropriate pancreatic enzymes and bile salts are secreted to aid in this digestion and absorption. There are five major actions of CCK, and each contributes to the overall process of fat, protein, and carbohydrate digestion and absorption. 1. Contraction of the gallbladder with simultaneous relaxation of the sphincter of Oddi ejects bile from the gallbladder into the lumen of the small intestine. Bile is needed for emulsification and solubilization of dietary lipids. 2. Secretion of pancreatic enzymes. Pancreatic lipases digest ingested lipids to fatty acids, monoglycerides, and cholesterol, all of which can be absorbed. Pancreatic amylase digests carbohydrates, pancreatic proteases digest protein 3. Secretion of bicarbonate (HCO3-) from the pancreas. This is not a major effect of CCK, but it potentiates the effects of secretin on HCO3- secretion. 4. Growth of the exocrine pancreas and gallbladder. 5. Inhibition of gastric emptying. CCK inhibits or slows gastric emptying and increases gastric emptying time. This action is critical for the processes of fat digestion and absorption, which require a considerable amount of time. Secretin Secretin, a 27-amino acid peptide, is structurally homologous to glucagon. Fourteen (14) of the 27 amino acids of secretin are identical and in the same position as those of glucagon. In contrast to gastrin and CCK, which have active fragments, all 27 amino acids of secretin are required for its biologic activity. Secretin is secreted by the S cells (secretin cells) of the duodenum in response to H+ and fatty acids in the lumen of the small intestine. Thus, secretion of secretin is initiated when the acidic gastric contents (pH < 4.5) arrive in the small intestine. The function of secretin is to promote the secretion of pancreatic and biliary HCO3-, which then neutralizes H+ in the lumen of the small intestine. Neutralization of H+ is essential for fat digestion; pancreatic lipases have pH optimums between 6 and 8, and they are inactivated or denatured when the pH is less than 3. Secretin also inhibits the effects of gastrin on the parietal cells (H+ secretion and growth of gastric mucosa). Glucose-Dependent Insulinotropic Peptide. (Gastric inhibitory peptide, GIP) Glucose-dependent insulinotropic peptide (GIP), a 42amino-acid peptide. GIP has 9 amino acids in common with secretin and 16 amino acids in common with glucagon. Because of this homology, pharmacologic levels of GIP produce most of the actions of secretin. GIP is secreted by K cells of the duodenal and jejunal mucosa. It is the only gastrointestinal hormone that is secreted in response to all three types of nutrients: glucose, amino acids, and fatty acids. The major physiologic action of GIP is stimulation of insulin secretion by the pancreatic β cells. This action explains the observation that an oral glucose load is utilized by cells more rapidly than an equivalent intravenous glucose load. Oral glucose stimulates GIP secretion, which stimulates insulin secretion (in addition to the direct stimulatory action of absorbed glucose on the β cells). Glucose given intravenously stimulates insulin secretion only by the direct action on the β cells. The other action of GIP is inhibition of gastric H+ secretion (secretin-like action). Motilin, a 22-amino acid peptide. It is secreted from the upper duodenum during fasting states. Motilin is believed to increase gastrointestinal motility and, specifically, to initiate the interdigestive myo-electric complexes that occur at 90-minute intervals. Pancreatic polypeptide is a 36-amino acid peptide secreted by the pancreas in response to ingestion of carbohydrates, proteins, or lipids. Pancreatic polypeptide inhibits pancreatic secretion of HCO3- and enzymes, although its physiologic role is uncertain. Enteroglucagon is released from intestinal cells in response to a decrease in blood glucose concentration. It then directs the liver to increase glycogenolysis and gluconeogenesis. Somatostatin is secreted by D cells of the gastrointestinal mucosa in response to decreased luminal pH. In turn, somatostatin inhibits secretion of the other gastrointestinal hormones and inhibits gastric H+ secretion. In addition to these paracrine functions in the gastrointestinal tract, somatostatin is secreted by the hypothalamus and by the delta (δ) cells of the endocrine pancreas. Histamine is secreted by endocrine-type cells of the gastrointestinal mucosa, particularly in the H+-secreting region of the stomach. Histamine, along with gastrin and ACh, stimulates H+ secretion by the gastric parietal cells. NEUROCRINES Neurocrines are synthesized in cell bodies of gastrointestinal neurons. An action potential in the neuron causes release of the neurocrine, which diffuses across the synapse and interacts with receptors on the postsynaptic cell. Substance Source Acetylcholine (ACh) Cholinergic neurons Norepinephrine (NE) Adrenergic neurons Actions Contraction of smooth muscle in wall but Relaxation of sphincters ↑ Salivary secretion ↑ Gastric secretion ↑ Pancreatic secretion Relaxation of smooth muscle in wall but Contraction of sphincters ↑ Salivary secretion Vasoactive Intestinal Peptide (VIP) Neurons of mucosa and Gastrin-Releasing Peptide (GRP) Neurons of gastric mucosa Enkephalins (opiates) Neurons of mucosa and smooth muscle smooth muscle Neuropeptide Y Neurons of mucosa and smooth muscle Substance P Co-secreted with ACh Relaxation of smooth muscle ↑ Intestinal secretion ↑ Pancreatic secretion ↑ Gastrin secretion Contraction of smooth muscle ↓ Intestinal secretion Relaxation of smooth muscle ↓ Intestinal secretion Contraction of smooth muscle ↑ Salivary secretion The neurones are divided into: Cholinergic neurons: parasympathetic neurones and release Acetylcholine (Ach). Adrenergic neurons: sympathetic neurones and release norepinephrine. Peptidergic neurons: parasympathetic neurons but noncholinergic. They release the other neurocrines. Effect of Gut Activity and Metabolic Factors on Gastro-intestinal Blood Flow Under normal conditions, the blood flow in each area of the gastrointestinal tract, as well as in each layer of the gut wall, is directly related to the level of local activity. For instance, during active absorption of nutrients, blood flow in the villi and adjacent regions of the submucosa is increased as much as eightfold. Likewise, blood flow in the muscle layers of the intestinal wall increases with increased motor activity in the gut. For instance, after a meal, the motor activity, secretory activity, and absorptive activity all increase; likewise, the blood flow increases greatly but then decreases back to the resting level over another 2 to 4 hours. Possible Causes of the Increased Blood Flow During Gastrointestinal Activity. Although the precise cause or causes of the increased blood flow during increased gastrointestinal activity are still unclear, some facts are known. First, several vasodilator substances are released from the mucosa of the intestinal tract during the digestive process. Most of these are peptide hormones, including cholecystokinin, vasoactive intestinal peptide, gastrin, and secretin. Second, some of the gastrointestinal glands also release into the gut wall two kinins, kallidin and bradykinin, at the same time that they secrete their secretions into the lumen. These kinins are powerful vasodilators that are believed to cause much of the increased mucosal vasodilation that occurs along with secretion. Third, decreased oxygen concentration in the gut wall can increase intestinal blood flow at least 50 to 100 per cent; The decrease in oxygen can also lead to as much as a fourfold increase of adenosine, a well known vasodilator that could be responsible for much of the increased flow. “Countercurrent” Blood Flow in the Villi. The arterial flow into the villus and the venous flow out of the villus are in directions opposite to each other, and that the vessels lie in close apposition to each other. Because of this vascular arrangement, much of the blood oxygen diffuses out of the arterioles directly into the adjacent venules without ever being carried in the blood to the tips of the villi. As much as 80 per cent of the oxygen may take this short-circuit route and therefore not be available for local metabolic functions of the villi. Under normal conditions, this shunting of oxygen from the arterioles to the venules is not harmful to the villi, but in disease conditions in which blood flow to the gut becomes greatly curtailed, such as in circulatory shock, the oxygen deficit in the tips of the villi can become so great that the villus tip or even the whole villus suffers ischemic death and can disintegrate. Therefore, for this reason and others, in many gastrointestinal diseases the villi become seriously blunted, leading to greatly diminished intestinal absorptive capacity. Nervous Control of Gastrointestinal Blood Flow Parasympathetic stimulation of the nerves going to the stomach and lower colon increases local blood flow at the same time that it increases glandular secretion. This increased flow probably results secondarily from the increased glandular activity and not as a direct effect of the nervous stimulation. Sympathetic stimulation, by contrast, has a direct effect on essentially all the gastrointestinal tract to cause intense vasoconstriction of the arterioles with greatly decreased blood flow. After a few minutes of this vasoconstriction, the flow often returns almost to normal by means of a mechanism called “autoregulatory escape.” That is, the local metabolic vasodilator mechanisms that are elicited by ischemia become prepotent over the sympathetic vasoconstriction and, therefore, redilate the arterioles, thus causing return of necessary nutrient blood flow to the gastrointestinal glands and muscle. Importance of Nervous Depression of Gastro-intestinal Blood Flow When Other Parts of the Body Need Extra Blood Flow. (1) It allows shut-off of gastrointestinal and other splanchnic blood flow for short periods of time during heavy exercise, when increased flow is needed by the skeletal muscle and heart. (2) Also, in circulatory shock, when all the body’s vital tissues are in danger of cellular death for lack of blood flow—especially the brain and the heart—sympathetic stimulation can decrease splanchnic blood flow to very little for many hours. Sympathetic stimulation also causes strong vasoconstriction of the large-volume intestinal and mesenteric veins. This decreases the volume of these veins, thereby displacing large amounts of blood into other parts of the circulation. In hemorrhagic shock or other states of low blood volume, this mechanism can provide as much as 200 to 400 milliliters of extra blood to sustain the general circulation. GIT MOVEMENT Propulsion and Mixing Contractions of gastrointestinal smooth muscle can be either phasic or tonic. Two types of phasic movements occur in the gastrointestinal tract: (1) propulsive movements, which cause food to move forward along the tract at an appropriate rate to accommodate digestion and absorption, and (2) mixing movements, which keep the intestinal contents thoroughly mixed at all times. Propulsive Movements—Peristalsis A contractile ring appears around the gut and then moves forward, any material in front of the contractile ring is moved forward. Peristalsis is an inherent property of many syncytial smooth muscle tubes; stimulation at any point in the gut can cause a contractile ring to appear in the circular muscle, and this ring then spreads along the gut tube. (Peristalsis also occurs in the bile ducts, glandular ducts, ureters and many other smooth muscle tubes of the body.) Stimuli for Peristalsis: (1) The usual stimulus for intestinal peristalsis is distention of the gut. That is, if a large amount of food collects at any point in the gut, the stretching of the gut wall stimulates the enteric nervous system to contract the gut wall 2 to 3 centimeters behind this point, and a contractile ring appears that initiates a peristaltic movement. (2) Other stimuli that can initiate peristalsis include chemical or physical irritation of the epithelial lining in the gut. (3) Parasympathetic nervous signals to the gut will elicit strong peristalsis. Role of the Myenteric Plexus in Peristalsis. Effectual peristalsis requires an active myenteric plexus. Peristalsis occurs only weakly or not at all in any portion of the gastrointestinal tract that has congenital absence of the myenteric plexus. Also, it is greatly depressed or completely blocked in the entire gut when a person is treated with atropine to paralyze the cholinergic nerve endings of the myenteric plexus. Directional Movement of Peristaltic Waves Toward the Anus. Peristalsis, theoretically, can occur in either direction from a stimulated point, but it normally dies out rapidly in the orad direction while continuing for a considerable distance toward the anus. The exact cause of this directional transmission of peristalsis has never been ascertained, although it probably results mainly from the fact that the myenteric plexus itself is “polarized” in the anal direction, which can be explained as follows. Peristaltic Reflex and the “Law of the Gut.” When a segment of the intestinal tract is excited by distention and thereby initiates peristalsis, the contractile ring causing the peristalsis normally begins on the orad side of the distended segment and moves toward the distended segment, pushing the intestinal contents in the anal direction for 5 to 10 centimeters before dying out. At the same time, the gut sometimes relaxes several centimeters downstream toward the anus, which is called “receptive relaxation,” thus allowing the food to be propelled more easily anally than orad. This complex pattern does not occur in the absence of the myenteric plexus. Therefore, the complex is called the myenteric reflex or the peristaltic reflex. The peristaltic reflex plus the anal direction of movement of the peristalsis is called the “law of the gut.” Mixing Movements Mixing movements differ in different parts of the alimentary tract. (1) In some areas, the peristaltic contractions themselves cause most of the mixing. This is especially true when forward progression of the intestinal contents is blocked by a sphincter, so that a peristaltic wave can then only shake the intestinal contents, rather than propelling them forward. (2) At other times, local intermittent constrictive contractions occur every few centimeters in the gut wall. These constrictions usually last only 5 to 30 seconds; then new constrictions occur at other points in the gut, thus “chopping” and “shearing” the contents first here and then there. SHEWING Chewing has three functions: 1. It mixes food with saliva, lubricating it to facilitate swallowing; 2. it reduces the size of food particles, which facilitates swallowing (although the size of the swallowed particles has no effect on the digestive process); and 3. it mixes ingested carbohydrates with salivary amylase to begin carbohydrate digestion. Chewing has both voluntary and involuntary components. The involuntary component involves reflexes initiated by food in the mouth. Sensory information is relayed from mechanoreceptors in the mouth to the brain stem, which orchestrates a reflex oscillatory pattern of activity to the muscles involved in chewing. Voluntary chewing can override involuntary or reflex chewing at any time. Swallowing There are three phases involved in swallowing: oral, pharyngeal, and esophageal. The oral phase is voluntary, and the pharyngeal and esophageal phases are controlled by reflexes. Oral phase. The oral phase is initiated when the tongue forces a bolus of food back toward the pharynx, which contains a high density of somatosensory receptors. Activation of these receptors initiates the involuntary swallowing reflex in the medulla. Pharyngeal phase. The purpose of the pharyngeal phase is to propel the food bolus from the mouth through the pharynx to the esophagus in the following steps: 1. The soft palate is pulled upward, creating a narrow passage for food to move into the pharynx so food cannot reflux into the nasopharynx. 2. The epiglottis moves to cover the opening to the larynx, and the larynx moves upward against the epiglottis to prevent food from entering the trachea. 3. The upper esophageal sphincter relaxes, allowing food to pass from the pharynx to the esophagus. 4. A peristaltic wave of contraction is initiated in the pharynx and propels food through the open sphincter. Breathing is inhibited during the pharyngeal phase of swallowing. Afferent nerves. the sensory portions of the trigeminal and glossopharyngeal nerves. Reflex centre: The deglutition or swallowing center in the medulla and lower pons. Efferent nerves. To pharynx and upper esophagus by the 5th, 9th, 10th, and 12th cranial nerves and even a few of the superior cervical nerves. Effect of the Pharyngeal Stage of Swallowing on Respiration. The entire pharyngeal stage of swallowing usually occurs in less than 6 seconds, thereby interrupting respiration for only a fraction of a usual respiratory cycle. The swallowing center specifically inhibits the respiratory center of the medulla during this time, halting respiration at any point in its cycle to allow swallowing to proceed. Yet, even while a person is talking, swallowing interrupts respiration for such a short time that it is hardly noticeable.