Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Phospholipid-derived fatty acids wikipedia , lookup
Human microbiota wikipedia , lookup
Microorganism wikipedia , lookup
Triclocarban wikipedia , lookup
Plant virus wikipedia , lookup
Community fingerprinting wikipedia , lookup
Horizontal gene transfer wikipedia , lookup
Metagenomics wikipedia , lookup
Introduction to viruses wikipedia , lookup
Magnetotactic bacteria wikipedia , lookup
Disinfectant wikipedia , lookup
History of virology wikipedia , lookup
Bacterial morphological plasticity wikipedia , lookup
Bacterial cell structure wikipedia , lookup
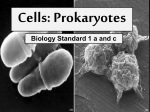
19 Bacteria, Archaea, and Viruses Chapter 19 Bacteria, Archaea, and Viruses Key Concepts • 19.1 Life Consists of Three Domains That Share a Common Ancestor • 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life • 19.3 Ecological Communities Depend on Prokaryotes • 19.4 Viruses Have Evolved Many Times Chapter 19 Opening Question How do Vibrio populations detect when they are dense enough to produce bioluminescence? Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor All organisms have: • Plasma membranes and ribosomes • Metabolic pathways (e.g., glycolysis) • Conservative DNA replication • DNA that encodes proteins Shared features indicate that all life is related, but major differences have also evolved. Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Three domains of life: • Bacteria—prokaryotes • Archaea—prokaryotes • Eukarya—eukaryotes Figure 19.1 The Three Domains of the Living World Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Prokaryotes differ from eukaryotes. • All are unicellular • Divide by binary fission, not mitosis • DNA is often circular, not in a nucleus • No membrane-enclosed organelles Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Genetic studies show that the three domains had a single common ancestor. Some eukaryote genes are most closely related to those of archaea, while others are most closely related to those of bacteria. Mitochondria and chloroplasts of eukaryotes originated through endosymbiosis with a bacterium. Table 19.1 The Three Domains of Life on Earth Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Study of prokaryotes was not possible until microscopes were developed. Before DNA sequencing, classification was based on phenotypic characters such as shape, color, motility, nutrition, and cell wall structure. Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Most bacteria cell walls contain peptidoglycan, which is unique to bacteria. Antibiotics target peptidoglycan because eukaryote cells don’t have it, thus there is no harm to human cells. Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Bacteria can be grouped by the Gram stain response, which is based on differences in cell wall structure: Gram-positive bacteria appear blue to purple. Gram-negative bacteria appear pink to red. Figure 19.2 The Gram Stain and the Bacterial Cell Wall (Part 1) Figure 19.2 The Gram Stain and the Bacterial Cell Wall (Part 2) Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Common bacteria cell shapes: • Sphere—coccus (plural cocci), occur singly or in plates, blocks, or clusters • Rod—bacillus (plural bacilli) • Spiral or helical—helix (plural helices) Rods and helical shapes may form chains or clusters. Other bacterial shapes form filaments and branched filaments. Figure 19.3 Bacterial Cell Shapes Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Sequencing of ribosomal RNA (rRNA) genes is useful for phylogenetic studies because: • rRNA was present in the common ancestor of all life. • All free-living organisms have rRNA. • Lateral transfer of rRNA genes among distantly related species is unlikely. • rRNA has evolved slowly. Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Whole genome sequencing has revealed that even distantly related prokaryotes sometimes exchange genetic material. Transformation, conjugation, and transduction allow exchange of genetic information between prokaryotes without reproduction. In lateral gene transfer, genes move “sideways” from one species to another. When sequenced, gene trees will not match the organismal tree. Figure 19.4 Lateral Gene Transfer Complicates Phylogenetic Relationships Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Genes that result in new adaptations that confer higher fitness are most likely to be transferred. Genes for antibiotic resistance are often transferred among bacterial species. Concept 19.1 Life Consists of Three Domains That Share a Common Ancestor Many prokaryote species, and perhaps whole clades, have not been described by biologists. Many have resisted efforts to grow them in pure culture. Biologists now examine gene sequences collected from random samples of the environment. Many new sequences imply there are thousands more prokaryotic species. Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Prokaryotes are the most successful organisms on Earth in terms of number of individuals. The number of prokaryotes in the ocean is perhaps 100 million times as great as the number of stars in the visible universe. They are found in every type of habitat on Earth. We will describe eight bacterial groups. Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Low-GC Gram-positives (Firmicutes) Low ratio of G-C to A-T base pairs in DNA. Some are gram-negative, and some have no cell wall. Some produce heat-resistant endospores that can survive unfavorable conditions. Some can survive for 1,000 years. Includes Clostridium and Bacillus. Figure 19.5 A Structure for Waiting Out Bad Times Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Bacillus anthracis produces an exotoxin that causes anthrax. The endospores have been used as a bioterrorism agent. Staphylococcus (staphylococci) are abundant on skin and cause boils and other skin problems. S. aureus can also cause respiratory, intestinal, and wound infections. Figure 19.6 Staphylococci Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Mycoplasmas have no cell wall, are extremely small, and have a very small genome. They have less than half as much DNA as other prokaryotes, which may represent the minimum amount of DNA needed for a living cell. Figure 19.7 Tiny Cells Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life High-GC Gram-positives (Actinobacteria) Higher ratio of G-C to A-T base pairs. Branched filaments; some form reproductive spores at filament tips. Most antibiotics are from this group. Mycobacterium tuberculosis causes tuberculosis; oldest know human pathogen. Figure 19.8 Actinomycetes Are High-GC Gram-Positives Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Hyperthermophilic bacteria Live at extreme high temperatures (extremophiles)—hot springs, volcanic vents, underground oil reservoirs. High temperatures may have been the ancestral condition on Earth when prokaryotes evolved. Monophyly of this group is not well established. Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Hadobacteria Also extreme thermophiles. Deinococcus survive cold as well as hot temperatures and are resistant to radiation. They can consume nuclear waste. Thermus aquaticus was isolated from a hot spring; source of the thermally stable DNA polymerase used in PCR. Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Cyanobacteria Photosynthetic; have blue-green pigments. Many species fix nitrogen. Chloroplasts of eukaryotes are derived from an endosymbiotic cyanobacterium. Some colonies differentiate into vegetative cells, spores, and heterocysts specialized for N-fixation. Figure 19.9 Cyanobacteria (Part 1) Figure 19.9 Cyanobacteria (Part 2) Figure 19.9 Cyanobacteria (Part 3) Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Spirochetes Gram-negative; motile Unique axial filaments (modified flagella) that rotate Many are human parasites, some are pathogens (syphilis, Lyme disease), others are free living. Figure 19.10 Spirochetes Get Their Shape from Axial Filaments (Part 1) Figure 19.10 Spirochetes Get Their Shape from Axial Filaments (Part 2) Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Chlamydias Can live only as parasites in cells of other organisms. Gram-negative; extremely small Can take up ATP from host cell with translocase Complex life cycle with two forms—elementary bodies and reticulate bodies Figure 19.11 Chlamydias Change Form during Their Life Cycle Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Proteobacteria: largest group of bacteria Mitochondria of eukaryotes were derived from a proteobacterium by endosymbiosis. Some are photoautotrophs that use light energy to metabolize sulfur; some are N-fixers (Rhizobium). Escherichia coli is one of the most studied organisms on Earth. Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Agrobacterium tumefaciens causes crown gall disease of plants and has a plasmid used in recombinant DNA studies. The proteobacteria include many pathogens— cholera, bubonic plague, salmonella. Figure 19.12 Proteobacteria Include Human Pathogens Figure 19.13 Crown Gall Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Separation of the Archaea domain from bacteria and eukaryotes is based on genome sequencing. Many archaea live in extreme habitats—high temperatures, low oxygen, high salinity, extreme pH. Many others are common in soil and in the oceans. Figure 19.14 What Is the Highest Temperature Compatible with Life? (Part 1) Figure 19.14 What Is the Highest Temperature Compatible with Life? (Part 2) Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Archaea are divided into two main groups, Euryarcheota and Crenarcheota Two recently discovered groups: • Korarchaeota (known only from DNA in hot springs) • Nanoarchaeota, a parasite on cells of a crenarchaeote in deep sea hydrothermal vents All lack peptidoglycan in the cell walls and have unique lipids in the cell membranes. Figure 19.18 A Nanoarchaeote Growing in Mixed Culture with a Crenarchaeote Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Bacterial and eukaryotic membranes have lipids with fatty acids connected to glycerol by ester linkages. Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Archaeal membranes have lipids with fatty acids linked to glycerol by ether linkages. This is a synapomorphy of archaea. Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Some archaeal lipids have glycerol at both ends and form lipid monolayers. Others have lipid bilayers. Figure 19.15 Membrane Architecture in Archaea Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Most Crenarcheota are thermophilic and/or acidophilic (acid loving). Sulfolobus lives in hot sulfur springs (70–75°C, pH 2 to 3). They can still maintain an internal pH of 5.5 to 7. Figure 19.16 Crenarchaeotes Like It Hot Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Some Euryarcheota are methanogens. CH4 is produced by reducing CO2; they are obligate anaerobes. Methanogens release 2 billion tons of methane per year. Many live in the guts of grazing mammals, termites, and cockroaches. Increased cattle farming and rice growing contributes methane to the atmosphere. Concept 19.2 Prokaryote Diversity Reflects the Ancient Origins of Life Extreme halophiles (salt lovers) have pink carotenoid pigments. Live in the most salty, most alkaline environments on Earth. Some have a light-absorbing molecule, microbial rhodopsin, to trap light energy and form ATP. Figure 19.17 Extreme Halophiles Concept 19.3 Ecological Communities Depend on Prokaryotes Many prokaryotes form complex communities. Biofilms—cells bind to a solid surface and secrete a sticky polysaccharide matrix that traps other cells. Cells in biofilms are hard to kill. Can form on any surface, including contact lenses, artificial joint replacements, metal pipes. Dental plaque and stromatolites are biofilms. Figure 19.19 Forming a Biofilm (Part 1) Figure 19.19 Forming a Biofilm (Part 2) Concept 19.3 Ecological Communities Depend on Prokaryotes The long evolutionary history of prokaryotes has led to a great diversity of metabolic pathways. They have evolved huge variation in use or nonuse of oxygen, energy and carbon sources, and waste products produced. Concept 19.3 Ecological Communities Depend on Prokaryotes Anaerobes do not use oxygen as an electron acceptor in respiration. Obligate anaerobes—oxygen is poisonous. Aerotolerant anaerobes—not damaged by oxygen. Facultative anaerobe—use both aerobic and anaerobic metabolic pathways. Obligate aerobes—require oxygen. Concept 19.3 Ecological Communities Depend on Prokaryotes Nutritional categories: • Photoautotrophs perform photosynthesis; use CO2 as carbon source. Cyanobacteria use chlorophyll a and produce O2. Others use bacteriochlorophyll and produce sulfur; H2S is the electron donor. Concept 19.3 Ecological Communities Depend on Prokaryotes Bacteriochlorophyll absorbs longer wavelengths than chlorophyll. Bacteria using this pigment can grow in deeper water under dense layers of algae, using light that is not absorbed by the algae. Figure 19.20 Bacteriochlorophyll Absorbs Long-Wavelength Light Concept 19.3 Ecological Communities Depend on Prokaryotes • Photoheterotrophs use light as an energy source, but get carbon from organic compounds made by other organisms. Sunlight provides the ATP through photophosphorylation. Concept 19.3 Ecological Communities Depend on Prokaryotes • Chemolithotrophs (chemoautotrophs) get energy by oxidizing inorganic substances and use it to fix carbon. Inorganic compounds oxidized include ammonia, nitrite, hydrogen gas, hydrogen sulfide, sulfur, and other materials. Concept 19.3 Ecological Communities Depend on Prokaryotes • Chemoheterotrophs get both energy and carbon from organic compounds that have been synthesized by other organisms. Most known bacteria and archaea are chemoheterotrophs—as are all animals, fungi, and many protists. Table 19.2 How Organisms Obtain Their Energy and Carbon Concept 19.3 Ecological Communities Depend on Prokaryotes Prokaryotes play a major role in the cycling of elements. Many are decomposers: they metabolize organic compounds in dead organic material. The inorganic products, such as CO2, are returned to the environment. Other prokaryotes oxidize inorganic compounds and also play key roles in element cycling. Concept 19.3 Ecological Communities Depend on Prokaryotes Denitrifiers: Bacteria that use nitrate (NO3–) as an electron acceptor in place of O2 in anaerobic conditions. They release N2 to the atmosphere. They play a key role in nitrogen cycling. Concept 19.3 Ecological Communities Depend on Prokaryotes Nitrogen fixers: Convert N2 to ammonia. Ammonia is a form of nitrogen that is useable by organisms. Nitrogen fixation is vital to life and is done only by certain prokaryote species. Concept 19.3 Ecological Communities Depend on Prokaryotes Nitrifiers: Chemolithotrophic bacteria that oxidize ammonia to nitrate. Nitrate is the form of nitrogen most easily used by many plants. Concept 19.3 Ecological Communities Depend on Prokaryotes Many prokaryotes live on or in eukaryotic organisms (e.g., nitrogen fixers that live in plant root nodules). Animals have many prokaryotes in their digestive tracts. Bacteria in the rumen of cattle produce the enzyme needed to digest cellulose. Concept 19.3 Ecological Communities Depend on Prokaryotes Humans have thousands of bacterial species on their skin and in their guts. Gut bacteria help digest and absorb nutrients and produce vitamins. Only a small percentage of bacteria are pathogens. Concept 19.3 Ecological Communities Depend on Prokaryotes Late 19th century—studies began to show the microbial basis of some diseases. Koch’s postulates were rules for establishing that a particular microorganism causes a particular disease. These rules were used to show that ulcers are caused by a bacterium Helicobacter pylori. Figure 19.21 Satisfying Koch’s Postulates (Part 1) Figure 19.21 Satisfying Koch’s Postulates (Part 2) Concept 19.3 Ecological Communities Depend on Prokaryotes In spite of the many defense mechanisms of potential hosts, some bacteria are very successful pathogens. Because they form biofilms, pathogens can be hard to combat. Consequences of bacterial infections depend on invasiveness of the pathogen (ability to multiply in host’s body) and its toxigenicity (ability to produce toxins). Concept 19.3 Ecological Communities Depend on Prokaryotes Endotoxins are released when certain Gramnegative bacteria lyse (burst); rarely fatal; they cause fever, vomiting, and diarrhea. Examples: Salmonella and Escherichia Exotoxins are released by living bacteria; highly toxic, often fatal. Examples: tetanus (Clostridium tetani), cholera (Vibrio cholerae), bubonic plague (Yersinia pestis), anthrax (Bacillus anthracis), botulism (Clostridium botulinum). Concept 19.4 Viruses Have Evolved Many Times Although viruses are not cellular, they have many characteristics of living organisms. Virus phylogeny is difficult to resolve: small genomes restrict phylogenetic analyses; rapid mutation and evolution rates cloud evolutionary relationships; there are no fossils. Instead, viruses are grouped based on genome structure. Concept 19.4 Viruses Have Evolved Many Times Viruses are obligate cellular parasites, but many may have once been cellular components. They may be “escaped” components cells that now evolve independently of their hosts. Concept 19.4 Viruses Have Evolved Many Times Negative-sense single-stranded RNA viruses: Negative-sense RNA—the complement of mRNA. They can make mRNA from their negative-sense RNA genome. These viruses probably arose by cellular escape many times independently across the tree of life. Includes viruses that cause measles, mumps, rabies, and influenza. Figure 19.22 Viruses Are Diverse (Part 1) Concept 19.4 Viruses Have Evolved Many Times Positive-sense single-stranded RNA viruses: The most abundant and diverse group; includes mosaic viruses of crop plants, polio, hepatitis C, and the common cold. They also appear to have evolved multiple times across the tree of life from different groups of cellular ancestors. Figure 19.22 Viruses Are Diverse (Part 2) Figure 19.23 Mosaic Viruses Are a Problem for Agriculture Concept 19.4 Viruses Have Evolved Many Times RNA retroviruses: Single-stranded RNA; probably evolved as escaped cellular components. Regenerate themselves by reverse transcription. DNA is produced and integrated into the host genome, where it is replicated along with host’s DNA. Only infect vertebrates; includes HIV, and some are associated with various cancers. Figure 19.22 Viruses Are Diverse (Part 3) Concept 19.4 Viruses Have Evolved Many Times Double-stranded RNA viruses: May have evolved repeatedly from singlestranded RNA ancestors. Cause many plant diseases and infant diarrhea in humans. Concept 19.4 Viruses Have Evolved Many Times Double-stranded DNA viruses: May represent highly reduced parasitic organisms that have lost their cellular structure and ability to survive as free-living species. Some have genomes as large as parasitic bacteria. Includes bacteriophage, smallpox, and herpes viruses. Figure 19.22 Viruses Are Diverse (Part 4) Figure 19.22 Viruses Are Diverse (Part 5) Figure 19.22 Viruses Are Diverse (Part 6) Answer to Opening Question Bacteria release chemical substances that are sensed by others of the same species. As population increases, concentration of chemical signal builds up. Bacteria then start activities such as forming a biofilm (quorum sensing). When populations are dense enough, Vibrio produce bioluminescence, which attracts fish that eat the phytoplankton on which the bacteria are growing, which gets the bacteria into a new host.