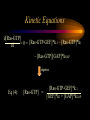Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
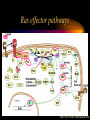
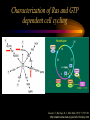
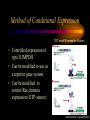
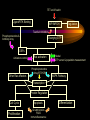
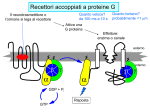
Modeling Guanine NucleotideRas Binding and Cell Behavior Kate Brown Anna Stevens Katy Wack Project Goals: • Understanding the quantitative relationship between IMPDH, intracellular GTP concentration, Ras mediated signaling and cell behavior • How does this relationship define a cell’s intracellular state and its decision making processes – Stem cell self renewal or maturation – Cancer cell proliferative capacity Implications of GTP in cell decisions • Stem Cell Self Renewal/asymmetric kinetics – Inhibition of IMPDH induces differentiation – Addition of guanine nucleotide precursors reverses this and restores exponential growth • Cancer cells-high proliferative/undifferentiated state (lose the ability to mature) – Some Cancer drugs (Tiazofurin), inhibit IMPDH, result in decrease of GTP and change in proliferative capacity, not just proliferative rate De Novo Nucleotide Synthesis De Novo Sythesis (R5P) Xanthosine or Xanthine Guanosine or Guanine Salvage Pathways IMP IMPDH XMP GMP GDP GTP IMPDH is the rate limiting step of De Novo Synthessis How does Ras Signaling Work? GEF Ras GDP GDP Ras GDP Pi GAP GEF Ras Ras GTP GEF Ras GTP GTP Ras effector pathways http://193.175.244.148/maps/ras.html Influencing the kinetics of Ras-GTP binding • Change intracellular GTP concentration – IMPDH inhibition/stimulation • Change GAP/GEF – GTPase dephosphorylation – Nucleotide binding • Change Ras behavior – oncogenic Ras has different nucleotide binding affinity Inhibition of IMPDH reduces GTP and Ras-GTP Percent Ras in GTP-bound state in K562 IMPDH activity and [GTP] in HL-60 cell 35 16 30 14 12 Control 15 Tiazofurin 10 nmol/10 20 10 Tiazofurin 7 cells 25 8 Control 6 4 5 2 0 0 Ras-GTP IMPDH activity [GTP] Tiazofurin inhibits IMPDH lowering cellular GTP concentration GMP and GDP concentrations do not change appreciably due to An excess of enzymes converting them to GTP Knight et al. Blood, 69 634-639 (1987) Hata et al. Oncol Res., 5 (4-5) 161-164 (1993) Equilibrium Model [Ras-GDP] + [GEF] Keq1 [Ras-GDP-GEF] Keq2 kGAP [GDP] [Ras-GEF] [GAP] Keq3 [GTP] Keq1 [Ras-GTP] + [GEF] [Ras-GTP-GEF] Assumptions • The system is at equilibrium • Pseudo steady state - d[Ras-GTP]/dt = 0 • GEF binds Ras-GTP and Ras-GDP with no bias • The Ras-GEF complex does not bind equally to GTP and GDP Haney et al., J. Bio. Chem 269 (24) 16541-16548 (1994) Lenzen et al., Biochem 37 7420-7430 (1998) Equilibrium Equations Eq (1): [Ras-GDP-GEF] Keq1= [Ras-GDP][GEF] Eq (2): [Ras-GEF][GDP] Keq2= [Ras-GDP-GEF] Eq (3): [Ras-GEF][GTP] Keq3= [Ras-GTP-GEF] = [Ras-GTP-GEF] [Ras-GTP][GEF] Kinetic Equations d[Ras-GTP] [Ras-GTP-GEF]*k-1 – [Ras-GTP]*k1 = 0 = dT – [Ras-GTP][GAP]*kGAP algebra Eq (4): [Ras-GTP-GEF]*k-1 [Ras-GTP] = [GEF]*k1 + [GAP]*kGAP Working Model Equation [Ras-GTP] [Ras-GDP] [GTP] Keq2 [GDP] * Keq3 = Keq2 = .625 uM-1 * Keq3 = 3.33 uM-1 35 [Ras-GTP]/[Ras-GDP] 30 [Ras-GTP/[Ras-GDP] = 0.1875[GTP]/[GDP] slope=Keq2/keq3 25 20 15 10 5 0 0 20 40 60 80 100 120 140 160 180 200 [GTP]/[GDP] *As determined by Lenzen et al., Biochem 37 7420-7430 (1998) Model Limitations/ Future Work Experimental %[GTP] Change 37 +/- 13 * Experimental % Ras-GTP change 35% +/- 13 § Model prediction for % Ras-GTP change 37% • Need to generate more data for better determination of kinetic parameters in order to test model. • Evidence that there is biphasic activation of Ras, so we may want to explore the full time course of Ras activation, and therefore generate a kinetic model using our system. • Would like to incorporate our model into current MAPK signaling models to quantitatively predict the effect of changing GTP pools on the cellular response to extracellular ligands. * Knight et al. Blood, 69 634-639 (1987) § Hata et al. Oncol Res., 5 (4-5) 161-164 (1993) Experimental Goals • Explore the relationship between IMPDH and GTP – Measure total vs. signaling [GTP] • Explore GTP “sensing” by Ras – Consider both phases of Ras activation – Kinetics of Ras activation • Explore the role of specific Ras effecter pathways in cell cycle and maintaining “stemness” • Characterize changes in cell state with GTP variation • Quantify signaling system – Consider changes in GTP Experimental Cell Lines • Stem Cell – Putative adult rat liver stem cell line-lig 8 • Cancer Cell – Hepatoma 3924A cell line • Primary Epithelial – Hepatocytes, freshly isolated Characterization of Ras and GTP dependent cell cycling Joneson, T., Bar-Sagi, D., J. Mol. Med (1997) 75; 587-593 http://www2.hama-med.ac.jp/w1a/bio1/index-j.html GTP “Sensing” Fluorescence Resonace Energy Transfer • Use FRET to measure signaling GTP • Understand the spatial aspect of Ras activation • Use GTP-sensor to monitor biphasic behavior of Ras activation Cullen, P.J., Lockyer P.J., Nature Reviews Molecular Cell Biology 3; 339-348 (2002) Method of Conditional Expression TET on/off Expression System • Controlled expression of type II IMPDH • Can be modified to use as a reporter gene system • Can be modified to control Ras chimera expression (GTP-sensor) www.clontech.co.jp/qa/tet.html Tools for defining intracellular state at the Protein level Proteomics and Phosphoproteomics Antibody array for Protein expression www2.mrc-lmb.cam.ac.uk/groups/arrays swehsc.pharmacy.arizona.edu/analysis/images/proteomics.gif Monitoring Cellular State www.acl.ac.uk/biology/new/admin/pix/astrossm.jpg www.icnet.uk/axp/facs/davies/brdu1.gif TET on/off switch Ligand/RTK Binding IMP/IMPDH Regulation Tiazofurin Inhibition Phosphoproteomics & Antibody array Changing GTP GEFs Activation control Other Ras effectors Ras activation Model GTP sensor & population measurement Phosphoproteomics Array/RT-PCR MAPK Pathway Transcription Protein Regulation Cell Cycle Apoptosis Proliferation Growth kinetics FACS Immunofluoresence Differentiation Acknowledgements • • • • Dr. James Sherley Ali Khademhosseini BE computer room population Doug and Paul References • • • • • • • • • • • Sherley, J.L., An Emerging Cell Kinetics Network:Integrated Control of Nucleotide Metabolism and Cancer Gene Function, submitted Sherley, J.L., Asymmetric Cell Kinetics Genes: The Key to Expansion of adult Stem Cells in Culture, Stem Cells, 2002 Wright,D.G., A Role for Guanine Ribonucleotides in the Regulation of Myeloid Cell Maturation. Blood, Vol. 69 (1987) 334-337 Knight,R.D., Mangum,J., Lucas,D.L., Cooney,D.A., Khan,E.C., Wright,D.G., Insoine Monophosphate Dehydrogenase and Myeloid Cell Maturation. Blood, vol. 69 (1987) 634-639 Collart,F.R., Huberman,E., Expression of IMP Dehydrogenase in Differentiong HL-60 Cells, Blood, vol.75 (3) (1990) 570-576 Colombo,R.S., Coccetti,P., Martegani,E., Role of guanine nucleotides in the regulation of the Ras/cAMP pathway in Saccharomyces cerevisiae. Biochima Biophys Acta, (2001) 181-189 Haney,S.A., Broach, J.R., Cdc25p, the guanine Nucleotide Exchange Factor for the Ras Proteins of Saccharomyces cervisiae, Promotes Exchange by stabilizing Ras in a Nucleotide-free State, J. Bio. Chem, vol. 269 (1994) 1654116548. Hata, Y., Natsumeda,Y., Weber,G., Tiazofurin decreases Ras-GTP complex in K4562 cells., Oncol Res (1993) 161164. Taylor S., Shalloway D., Cell cycle-dependent activation of Ras., Current Biology vol.6 (1996) 1621-1627 Nature Review Molecular Cell Biology 3; 339-348 (2002) Gille H., Downward J., Multiple Ras Effector Pathways Contribute to G1 Cell Cycle Progression, J. Biol CChem vol 274 (1999) 22033-22040