Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
BIOL 370 – Developmental Biology
Topic #7
A Continuation of
A First Look at Early Development:
More On Rapid Specification
in Snails and Nematodes
Lange
To understand cleavage, we need additional vocabulary:
• Vegetal pole – the yolk rich region
• Animal pole – the yolk devoid region
• Isolecithal – roughly equal distribution of yolk (such as in sea
urchins)
• Holoblastic cleavage – complete cleavage
• Meroblastic cleavage – partial cleavage where only some of the
cytoplasm is cleaved (insects, fish, reptiles, birds)
• Centrolecithal – centrally placed yolk (insects)
• Telolecithal – only one area is free of yolk (birds and fish)
• Discoidal cleavage – cleavage in the telolecithal eggs that occurs
only in the small disk of cytoplasm
• Holoblastic cleavage subtypes:
• Radial
• Spiral
• Bilateral
• Rotational
• Vegetal pole – the yolk rich region
• Animal pole – the yolk devoid region
• Isolecithal – roughly equal distribution of yolk (such as in
sea urchins)
• Centrolecithal – centrally placed yolk (insects)
• Telolecithal – only one area is free of yolk (birds and fish)
• Holoblastic cleavage – complete cleavage
• Meroblastic cleavage – partial cleavage where only some of the
cytoplasm is cleaved (insects, fish, reptiles, birds)
• Discoidal cleavage – cleavage in the telolecithal eggs that occurs
only in the small disk of cytoplasm
• Holoblastic cleavage subtypes:
• Radial
• Spiral
• Bilateral
• Rotational
Summary of the main patterns of cleavage
We will focus on each
type of cleavage in
greater detail AGAIN
in the next few slides.
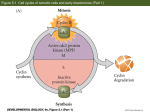
Summary of the main patterns of cleavage (Part 1)
Summary of the main patterns of cleavage (Part 2)
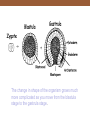
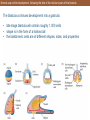
The change in shape of the organism grows much
more complicated as you move from the blastula
stage to the gastrula stage.
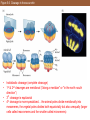
Types of cell movements during gastrulation
In this slide, we are seeing how during gastrulation, there is a noticeable
shift in terms of cell movement. Notice the five types shown:
• Invagination
• Involution
• Ingression
• Delamination
• Epiboly
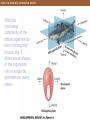
Axes of a bilaterally symmetrical animal
With this
increasing
complexity of the
whole organism do
the in-folding that
occurs, the 3
dimensional shape
of the organisms
will no longer be
symmetrical every
plane.
Figure 5.6 Cleavage in the sea urchin
•
•
•
•
holoblastic cleavage (complete cleavage)
1st & 2nd cleaveges are meridional ("along a meridian" or "in the north–south
direction“)
3rd cleavage is equitaorial
4th cleavage is more specialized… the animal poles divide meridionally into
mesomeres, the vegetal poles divides both equatorially but also unequally (larger
cells called macromeres and the smaller called micromeres)
Cleavage in the sea urchin (Part 1)
5th (16 cell stage) cleavage:
• mesomeres (animal) equitorially to
produce an1 and an2
• macromeres divide meridionally
• micromeres divide unequally
producing four small mircomeres
and four large micromeres
6th division animal hemisphere cells
divide meridionally and vegetal cells
divide equatorially.
7th division the vegetal cells divide
meriodionally and the animal cells
divide equatorially.
The embryo is now at 120 cells and is
called a blastula.
Micrographs of cleavage in live embryos of the sea urchin Lytechinus variegatus, seen from the
side
Fate map and cell lineage of the sea urchin Strongylocentrotus purpuratus (Part 1)
Embryo at 60 cell stage.
Oral – towards the
mouth
Aboral – away from
the mouth
Fate map and cell lineage of the sea urchin Strongylocentrotus purpuratus (Part 2)
Cell lineage and fate map.
Ability of micromeres to induce presumptive ectodermal cells to acquire other fates
In this series we can see how even in the dissected embryo missing
its vegetal portion, the micromeres can guide and direct
differentiation into a larva.
What does this
suggest as the role
for micomeres?
Normal sea urchin development, following the fate of the cellular layers of the blastula
The blastula continues development into a gastrula:
• late stage blastula will contain roughly 1,000 cells
• shape is in the form of a hollow ball
• the blastomeric cells are of different shapes, sizes, and properties
Gastrulation is a phase early in the embryonic
development of most animals, during which the singlelayered blastula is reorganized into a trilaminar ("threelayered") structure known as the gastrula. These three
germ layers are known as the ectoderm, mesoderm, and
endoderm.
Gastrulation is followed by organogenesis, when
individual organs develop within the newly formed germ
layers. Each layer gives rise to specific tissues and
organs in the developing embryo.
Simplified view of the change from blastula to gastrula.
The ectoderm gives rise to epidermis and also tissues that
will later form the nervous system.
The mesoderm is found between (meso = middle) the
ectoderm and the endoderm and gives rise to the muscular
system, cartilages, the dermis, the notochord, blood and
blood vessels, bone, and connective tissue.
The endoderm gives rise to the epithelium of the digestive
system and respiratory system, and organs associated with
the digestive system, such as the liver and pancreas.
Entire sequence of gastrulation in Lytechinus variegatus
Invagination of the vegetal plate
The archenteron is the primitive gut that forms during
gastrulation. It develops into the digestive tract of an
animal.
Cell rearrangement during extension of the archenteron in sea urchin embryos
The imaginal rudiment growing in the left side of the pluteus larva of a sea urchin
The imaginal rudiment is the precursor tissue for
the developing skeletal structure of the urchin.
A related line to the skeletal development seen in the
previous slides about the urchin, is the shell development
seen in a variety of snails. These molluscs that develop
conchae (spiral shaped shells), can develop often develop
either a right handed spiral or a left handed spiral.
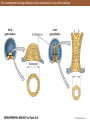
Spiral cleavage of the mollusc Trochus viewed from the animal pole (A) and from one side (B)
Looking down on the animal pole of left-coiling (A) and right-coiling (B) snails
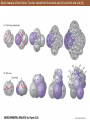
Fate map of Ilyanassa obsoleta
The glochidium is a microscopic larval stage seen in some
freshwater mussels, aquatic bivalve mollusks, and European
freshwater pearl mussels.
This larval form has hooks, which enable it to attach itself to
fish (for example to the gills of a fish host species) for a
period of time prior to when it detaches and falls to the
substrate and takes on the typical form of a juvenile mussel.
Ecologically, since fish are active and free-swimming, this
process helps distribute the mussel species to potential
areas of habitat that it could not reach via its own
locomotion.
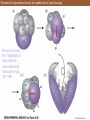
Formation of a glochidium larva by the modification of spiral cleavage
Be sure to read
the “Sidelights &
Speculations
associated with
this work on pp.
167-168.
Phony fish atop the unionid clam Lampsilis altilis
This fake “fish” is actually a brood pouch holding glochidium in this
species of clam. It attracts predatory fish and allows the glochidium easy
access to these fish.
The two-step process for specifying the marginal cells of the tunicate embryo
Macho-1 = the gene/RNA material
FGF = fibroblast growth factor
A fibroblast is a type of cell that synthesizes the extracellular matrix and
collagen, the structural framework for animal tissues, and plays a critical
role in wound healing. Fibroblasts are the most common cells of
connective tissue in animals.
Gastrulation in the tunicate
Convergent extension of the tunicate notochord
Caenorhabditis elegans is a free-living, transparent nematode
(roundworm), about 1 mm in length, which lives in temperate soil
environments. Research into the molecular and developmental biology of
C. elegans was begun in 1974 by Sydney Brenner (Nobel Prize in
Medicine in 2002) and it has since been used extensively as a model
organism.
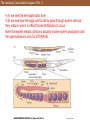
The nematode Caenorhabditis elegans (Part 1)
In A, we see the hermaphroditic form
In B, we see how the eggs are forced to pass through sperm cells as
they mature, which in effect forces fertilization to occur
Note that earlier meiotic divisions actually involve sperm production and
the spermatheca is only for STORAGE.
The nematode Caenorhabditis elegans (Part 2)
In the embryos formed, the “P”
cell represents the stem cells.
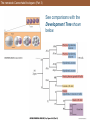
The nematode Caenorhabditis elegans (Part 3)
See comparisons with the
Development Tree shown
below.
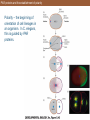
PAR proteins and the establishment of polarity
Polarity – the beginning of
orientation of cell lineages in
an organism. In C. elegans,
this is guided by PAR
proteins.
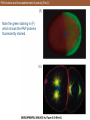
PAR proteins and the establishment of polarity (Part 2)
Note the green staining in (F)
which shows the PAR proteins
fluorescently stained.
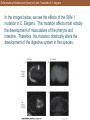
Deficiencies of intestine and pharynx in skn-1 mutants of C. elegans
In the images below, we see the effects of the SKN-1
mutation in C. Elegans. This mutation affects most notably
the development of musculature of the pharynx and
intestine. Therefore, this mutation drastically alters the
development of the digestive system in this species.
End.