Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
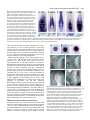
473 Development 125, 473-484 (1998) Printed in Great Britain © The Company of Biologists Limited 1998 DEV1253 Early posterior neural tissue is induced by FGF in the chick embryo Kate G. Storey1,†, Anne Goriely1, Catherine M. Sargent1, Jennifer M. Brown1, Helen D. Burns2, Helen M. Abud2,* and John K. Heath2 1Department 2Department of Human Anatomy, University of Oxford, South Parks Rd, Oxford, OX1 3QX, UK of Biochemistry, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK *Present address: Peter MacCallum Cancer Institute, Smorgon Family Building, Andrews Place, East Melbourne, Australia †Author for correspondence (e-mail: [email protected]) Accepted 21 November 1997: published on WWW 13 January 1998 SUMMARY Signals that induce neural cell fate in amniote embryos emanate from a unique cell population found at the anterior end of the primitive streak. Cells in this region express a number of fibroblast growth factors (FGFs), a group of secreted proteins implicated in the induction and patterning of neural tissue in the amphibian embryo. Here we exploit the large size and accessibility of the early chick embryo to analyse the function of FGF signalling specifically during neural induction. Our results demonstrate that extraembryonic epiblast cells previously shown to be responsive to endogenous neural-inducing signals express early posterior neural genes in response to local, physiological levels of FGF signal. This neural tissue does not express anterior neural markers or undergo neuronal differentiation and forms in the absence of axial mesoderm. Prospective mesodermal tissue is, however, induced and we present evidence for both the direct and indirect action of FGFs on prospective posterior neural tissue. These findings suggest that FGF signalling underlies a specific aspect of neural induction, the initiation of the programme that leads to the generation of the posterior central nervous system. INTRODUCTION hindbrain reticular neurons and spinal interneurons, the precursors of which arise adjacent to the anterior primitive streak during the early phase of neural induction (Sechrist and BronnerFraser, 1991; also see Lawson et al., 1991). The precursors of the posterior CNS (posterior hindbrain and spinal cord), therefore, appear to be among the first to respond to neuralinducing signals from Hensen’s node. We have recently shown that signals from the node induce expression of the basic helixloop-helix (bHLH) transcription factor cash4 and the homeodomain-containing gene Sax-1 (Henrique et al., 1997). These genes are expressed in the precursors of the posterior CNS at HH5-6 (Hamburger and Hamilton, 1951 stages) and therefore after L5, Sox 2 and FLIK. cash4 is a homologue of the Drosophila achaete-scute genes which are required for the formation of neural precursors in the fly (for review, see Campuzano and Modolell, 1992; Campos-Ortega, 1993). Overexpression of cash4 in the Xenopus embryo results in the expansion of the neural plate and an increase in the number of neurons produced. Further, expression of this chick gene in Drosophila reveals that it is a functional as well as structural homologue of the fly achaete-scute genes; it is able to rescue sensory bristle formation in flies lacking both achaete and scute (Henrique et al., 1997). Our functional studies therefore implicate cash4 in a similar early neural specification process in the posterior regions of the chick embryo (Henrique et al., 1997). We have also demonstrated that cash4 expression can be induced in anterior neural tissue following grafting of fibroblast growth factor (FGF)-soaked beads at primitive streak stages In higher vertebrates, the formation of the central nervous system (CNS) is initiated by signals that emanate from the anterior end of the primitive streak (Waddington, 1932; Gallera, 1971; Storey et al., 1992, 1995; Beddington, 1994). Grafts of this region (known as Hensen’s node in the chick embryo) induce extraembryonic epiblast cells to form an organised neural axis that expresses a range of anteroposterior marker genes and which contains differentiating neurons (Dias and Schoenwolf, 1990; Storey et al., 1992). However, the neural-inducing signals produced by Hensen’s node have yet to be identified. In the chick, cells competent to respond to such inducing signals express L5220 (a cell-surface glycoprotein, Roberts et al., 1991; Streit et al., 1997) whose domain of expression at early primitive streak stages includes the prospective neural plate and the region of extraembryonic epiblast cells responsive to Hensen’s node (Roberts et al., 1991). Expression of the L5 antigen can be maintained by Hepatocyte growth factor/scatter factor, a secreted factor that is present in Hensen’s node, but which does not induce neural tissue (Streit et al., 1995, 1997). A number of genes expressed in the early neural plate have also been identified and shown to be induced by Hensen’s node, these include the transcription factor Sox2 (Streit et al., 1997) and a follistatin-like (FLIK) molecule (Patel et al., 1996). However, the functions of these genes and the identity of the signals that regulate them have yet to be established. Among the first nerve cells to differentiate in the chick are Key words: Fibroblast growth factor, Neural induction, Hensen’s node, Chick 474 K. G. Storey and others (Henrique et al., 1997). This raises the possibility that FGF signalling plays a role in the anteroposterior patterning of the chick CNS. This result also implies that FGF signalling acts upstream of cash4 and therefore suggests that an early step in the neural programme could be the induction of cash4 by FGF signals. A number of FGFs are expressed in the anterior primitive streak when neural induction is taking place (FGF2, FGF3, FGF4, FGF8; Riese et al., 1995; Mahmood et al., 1995a,b; own observations) and might therefore play a role in the initiation of the neural programme. FGFs are characterised by shared features of sequence conservation and gene structure as well as the ability to bind to a conserved family of transmembrane signalling receptors of the tyrosine kinase class and heparan sulphate proteoglycans present in the extracellular matrix (reviewed Ornitz et al., 1996; Wilkie et al., 1995). FGF signals are therefore unlikely to diffuse far from their source and probably act locally on cells in their immediate vicinity. It is thus striking that FGFs are expressed during neural induction in and adjacent to the cells that will give rise to the posterior CNS. In the early amphibian embryo, FGFs have been shown to have general posteriorising effects (Slack and Tannahill, 1992) which act on both mesodermal and neural tissues (e.g. Isaacs et al., 1994; Cox and Hemmati-Brivanlou, 1995; Pownall et al., 1996). However, experiments assessing the function of FGFs in the induction of the amphibian nervous system are at present inconclusive. While FGFR1-mediated signalling appears not to be required for the induction and anteroposterior patterning of the nervous system in the intact embryo (Kroll and Amaya, 1996) and some explants of prospective epidermis (the animal cap) do not form neural tissue in response to FGF (Cox and Hemmati-Brivanlou, 1995), other reports suggest that this tissue does express neural genes in response to FGF and that FGF signalling in the ectoderm is required for its later response to neural-inducing signals (Kengaku and Okamoto 1993, 1995; Lamb and Harland, 1995; reviewed by Doniach, 1996; Launay et al., 1996; Sasai et al., 1996). Further, mice mutant for FGFR1 do not survive long enough for the role of FGF signalling in anteroposterior patterning to be assessed (Deng et al., 1994; Yamaguchi et al., 1994). Indeed, while some neural tissue appears to be induced in these embryos, they fail to undergo regression movements and may therefore fail to form more posterior regions of the CNS; a phenotype similar to that observed in chick embryos treated globally with molecules that bind FGF, heparin or suramin (Riese et al., 1995). Finally, while mouse embryos chimeric for FGF4 exhibit apparent overgrowth of the CNS (Abud, 1995), mice homozygous for the null allele of Fgf4 fail to develop far enough to assess the role of FGF signalling in early neural development (Feldman et al., 1995). In all these studies, cells are subjected to activation or inhibition of FGF signalling over prolonged periods of time in a spatially unrestricted manner. It is therefore difficult, using these experimental approaches, to define the action of FGF signalling at specific times during neural development. While the role of Fgf8 has recently been specifically assessed during midbrain development (Crossley et al., 1996a; Lee et al., 1997), there has been no study of FGF function during neural induction in higher vertebrates. We have accordingly addressed the role of FGF in the induction of neural tissue in the chick embryo by making use of local application of FGFs bound to beads, which permits tightly defined temporal and spatial activation of FGF-signalling pathways. Using this approach, we have assessed the neural-inducing activity of FGFs presented to extraembryonic epiblast cells that we have shown to be responsive to neural-inducing signals from Hensen’s node, but which normally give rise to extraembryonic membranes (Storey et al., 1992; 1995). We show that FGFsoaked heparin beads can maintain and/or induce the L5 epitope in extraembryonic epiblast cells and induce neural tissue that specifically expresses markers of the early posterior nervous system, including Sax-1 and cash4. Strikingly, this FGFinduced tissue does not express anterior neural markers and does not undergo neuronal differentiation. Induction of early posterior neural genes takes place in the absence of axial mesoderm. However, FGF signalling does induce prospective mesoderm (indicated by the presence of cells co-expressing Brachyury (Bra) and Delta-1). This raises the possibility that posterior neural genes are induced indirectly, by signals provided by prospective mesoderm. Significantly, this ‘indirect’ effect may also be FGF mediated as Fgf8 is induced shortly after Bra in response to FGF beads and is expressed in prospective mesoderm cells and the adjacent posterior neuroepithelium itself. The co-localisation of Bra and cash4 in epiblast cells around FGF beads also supports the possibility that FGF acts directly within the prospective neuroepithelium, downstream of bra expression in this tissue. This study is the first to assess the role of FGF signalling in the induction of neural tissue in an amniote embryo. It shows that FGF signals can mimic a very specific aspect of the neuralinducing capacity of the organiser region; the initiation of the programme that leads to the generation the posterior nervous system. MATERIALS AND METHODS Preparation of FGF proteins Human FGF-2 protein was expressed under the control of the trp promoter of plasmid pFC80 in E. coli strain FICE127 (both a gift of Dr Antonella Isacchi, Pharmacia Biopharmaceuticals) and purified by heparin affinity chromatography. Murine FGF-8A, FGF-8B and FGF9 were expressed in E. coli as glutathione-s transferase fusion proteins, purified by glutathione affinity chromatography, proteolytic cleavage and heparin affinity chromatography. Recombinant human FGF-4 protein was a gift of Dr David Rogers (Genetics Institute). Recombinant FGF-7 was purchased from R and D Systems (Europe) Ltd. The identity of all recombinant proteins was verified by amino acid sequencing. Preparation and detection of DIG-FGF-2 Digoxigenin-labelled FGF-2 (DIG-FGF-2) was prepared as described by Gleizes et al. (1994). Digoxigenin malemide was purchased from Boehringer Mannheim. DIG-FGF-2 exhibited comparable affinity and biological activity to wild-type FGF-2 as determined by binding to immobilised FGF receptors and the mitogenic response of fibroblasts. DIG-labelled FGF2 was detected in whole-mount embryos with an anti-DIG antibody conjugated with alkaline phosphatase (Boehringer Mannheim) using standard immunocytochemical techniques. Implanting FGF-soaked beads Heparin-coated acrylic beads (selected to be approx. 75-100 µm in diameter; Sigma H5263) were soaked in a FGF of choice for 30 minutes at room temperature and washed three times in phosphate-buffered saline (PBS) prior to implantation. Heparin-coated beads simply rinsed in PBS served as controls. Embryos at stages HH3-4+ were set up in New culture (New, 1955) ventral side up and individual beads were positioned Early posterior neural genes induced by FGF in contact with extraembryonic epiblast cells. Embryos with implanted beads were incubated at 37oC for desired periods of time up to 36 hours. DiI-labelling Epiblast cells were labelled with the lipophilic dye DiI (1,1′-dioctadecyl-3,3,3′,3′ tetramethyl indocarbocyanineperchlorate, Molecular Probes) using standard procedures (e.g. Stern, 1990; Izpisúa-Belmonte et al., 1993). The spatial relationship between the bead and labelled cells was confirmed using a cooled CCD camera to detect low level fluorescence through a fluorescein filter set (to reduce photobleaching and the production of toxic free-radicals) prior to incubation of embryos for the desired period. Embryos were then fixed in formal saline/EGTA and the pattern of fluorescently labelled cells observed using a rhodamine filter set to reveal cell movement. Fluorescent and bright-field images were superimposed using Adobe Photoshop. In situ hybridisation and immunocytochemistry Following New culture, embryos were fixed in 4% formaldehyde/PBS for 1 hour to overnight. Whole-mount in situ hybridisation protocol was adapted after Izpisúa-Belmonte et al. (1993). In some cases, embryos were further processed for immunocytochemistry with the Bra antibody (TN-1, kindly provided by Prof. Bernhard Herrman). Briefly, embryos were fixed for 30 minutes in formol saline at 4°C, washed in PBS and placed in a blocking solution (PBS containing 3% bovine serum albumin (BSA), 1% Triton X-100, 0.01% thimerosal and 5% heat-inactivated normal goat serum for 4 hours at room temperature). Polyclonal antibody TN-1 was added 1:500 and embryos incubated for 6 hours at room temperature. After extensive washing in PBS, embryos were incubated in blocking solution with peroxidase-conjugated goat anti-rabbit IgG antibody (Jackson ImmunoResearch) (1:50) overnight at 4°C. Embryos were then washed, underwent the usual diaminobenzidine tetrahydrochloride reaction and were postfixed in 4% formol saline. Whole-mount immunocytochemistry with Not-1, Inv4D9 and 3A10 antibodies were as described in Storey et al. (1992, 1995) and the L5 antibody was used as described by Streit at al. (1995). Marker gene expression patterns Mesoderm markers All the following genes or proteins are expressed in mesodermal tissues, however, all but the first are also expressed in the other tissues including the nervous system: •The Not-1 antibody recognises a notochord-specific antigen (Yamada et al., 1991) and is expressed shortly after this axial mesoderm emerges from Hensen’s node at HH7. •goosecoid is expressed in the node and later in the emerging prechordal plate (mesoderm and endoderm) and in the anterior neural folds from HH8 (Izpisúa-Belmonte et al., 1993). •Brachyury (Bra) is expressed in posterior regions of the chick embryo from primitive streak stages; it is found in prospective mesodermal cells in the primitive streak and in surrounding epiblast cells (Kispert et al., 1995). These domains of expression retain their spatial relationship with the regressing node as the posterior CNS is laid down. Bra is later also expressed in the notochord from HH5, in prospective posterior endoderm, and in the neural tube anterior to the first formed somite (Kispert et al., 1995; Knezevic et al., 1996). Bra is therefore a marker of all three germ layers in posterior regions of the embryo. •Fgf8 is also expressed in multiple germ layers; it is found in the anterior primitive streak (excluding the node) and flanking epiblast and in a number of discrete sites in the developing CNS, including the forebrain, the isthmus of the midbrain and the newly formed posterior neural plate (Heikinheimo et al., 1994; Crossley and Martin 1995; Mahmood et al., 1995; own observations). •cDelta-1 is expressed in presomitic mesoderm and nascent neurons (Henrique et al., 1995). 475 Neural markers •tailless is a steroid-hormone receptor expressed in the diencephalon from HH10 (Yu et al., 1994). •Engrailed-2 (En-2) is expressed in the posterior midbrain and rhombomere 1 of the hindbrain (Patel et al., 1989). •Krox-20 is expressed in the hindbrain in rhombomeres 3 and 5 from HH9 (Wilkinson et al., 1989). •cash4 is expressed at HH5 in the epiblast flanking Hensen’s node which will give rise to the posterior CNS and maintains its spatial relationship with the regressing node during the laying down of this region of the nervous system. (Henrique et al., 1997). cash4 expression extends laterally into prospective epidermal tissue and also has an extraembryonic domain in the forming blood islands. Thus while it is the first gene to distinguish prospective posterior neural tissue from the anterior neural plate, it is not a neural-specific gene. •Sax1 is expressed after cash4 in the prospective posterior CNS either side of the regressing Hensen’s node at HH6-7 (Spann et al., 1994; own observations). In contrast with cash4, Sax-1 expression is confined to the nervous system and is therefore the earliest neuralspecific marker expressed in the posterior regions of the embryo. •Hox b9 is expressed (at HH8-9) in prospective spinal cord and posterior mesoderm, its anterior boundary of expression is later established below the level of the somite 9 in the central nervous system, while its mesodermal domain is located more posteriorly at the 18/19 somite level. •The L5 antibody recognises an antigen expressed in the epiblast cells throughout the anterior two thirds of primitive streak stage embryos but which becomes restricted to the neural plate by HH8 (Roberts et al., 1991; Streit et al., 1995, 1996). •The 3A10 antibody (a gift of Dr Jane Dodd) recognises a neurofilament-associated antigen expressed in the chick by in early differentiating neurons in the posterior hindbrain, anterior spinal cord and diencephalon at HH11-12 (Yamada et al., 1991; Storey et al., 1992). RESULTS Grafts of Hensen’s node can induce extraembryonic epiblast cells to form an ectopic neural axis consisting of both anterior and posterior CNS that contains differentiating neurons (Storey et al., 1992). We have used this in vivo approach to assess the extent to which FGF signalling can mimic these organiser properties by presenting FGF4 on heparin-coated beads to the same extraembryonic epiblast cells. FGF4 was chosen because it contains a signal sequence and because it binds to a number of FGF receptors and is therefore likely to emulate the activity of other FGFs. In all cases, beads were soaked in 50 µg/ml FGF4 (FGF beads) (see below) grafted into HH3-3+ embryos (Fig. 1) and then incubated for 20-22 hours in New culture (New, 1955) until they reached HH8-9 (unless stated otherwise). FGF induces early posterior neural tissue in extraembryonic epiblast FGF beads, but not control PBS washed beads (PBS beads), induce ectopic structures expressing the early posterior neural markers Sax1 and cash4 (Table 1; Fig. 2A-D). Hoxb9, which is also expressed posteriorly in neural tissue and mesodermal tissue at HH8-9, is also induced in response to FGF beads (Table 1; Fig. 2E). FGF beads implanted in embryos incubated until they reached HH5 (n=4/4) as well as to HH8-9 (Table 1) maintain and/or induce expression in extraembryonic tissue of the L5 epitope (Fig.2H), an early pan-neural marker and possible indicator of neural competence (Roberts et al., 1991; Streit et al., 476 K. G. Storey and others accumulation of epiblast cells. In contrast with grafts of the young Hensen’s node, therefore, FGF signalling does not induce anterior neural tissue, but specifically elicits expression of genes characteristic of the inchoate posterior CNS. Fig. 1. FGF or control PBS beads were positioned against the extraembryonic epiblast in HH3-3+ host embryos and maintained in New culture for the desired period. 1997). These findings show that FGF signalling can induce neural tissue, which expresses a combination of transcription factors characteristic of early posterior neural tissue. FGF does not induce anterior neural markers To ascertain that FGF4 specifically initiates the generation of the posterior neural tissue, we assessed its ability to induce ectopic expression of a panel of neural genes characteristic of anterior regions: a forebrain marker, tailless, a midbrain/hindbrain boundary marker, Engrailed 2 and a marker of rhombomeres 3 and 5 of the hindbrain, Krox20. FGF beads do not induce ectopic expression of any of these genes when placed in the extraembryonic epiblast of HH3-3+ embryos (Table 2; Fig. 2F,G and 2I-L). Hensen’s nodes grafted into later stage hosts (HH4) induce truncated axes, which appear to constitute largely anterior CNS (Storey et al., 1992). It has also been observed in the amphibian embryo that competence to respond to FGF changes during development (Lamb and Harland, 1995) such that ageing ectoderm loses competence to respond to FGF by expressing posterior neural markers, but gains the ability to express progressively more anterior neural genes. We therefore assessed the ability of the extraembryonic epiblast to respond to FGF beads in later stage embryos. While the ability to induce ectopic Sax1 and cash4 is lost by HH4, this activity is not replaced by the induction of the more anteriorly expressed markers (tailless, Engrailed and Krox20) (Table 2). An alternative hypothesis is that, while high FGF concentrations induce expression of posterior neural genes, low levels induce genes characteristic of the anterior CNS (Kengaku and Okamoto, 1995; Lamb and Harland, 1995). We therefore also assessed the ability of FGF to induce expression of tailless and Krox-20 at lower concentrations (Fig. 4). Neither Krox-20, nor tailless are induced by FGF concentrations 10-fold lower (beads soaked in 5 µg/ml FGF4) than those able to induce cash4 and Sax-1 (Fig. 4). Indeed, FGF beads placed in HH4 hosts and beads soaked in this lower FGF concentration elicit only a small ectopic Table 1. Induction of neural genes by FGF in HH3-3+ extraembryonic epiblast FGF beads PBS beads cash4 sax1 L5 Hoxb9 12/18 (66%) 0/18 (0%) 9/16 (56 %) 0/8 (0%) 10/10 (100%) 0/6 (0%) 5/7 (71%) 0/4 (0%) FGF signalling does not elicit neuronal differentiation To determine whether FGF expression in the extraembryonic epiblast is sufficient to elicit a neural programme that proceeds to the onset of neuronal differentiation, embryos were grafted with FGF beads or PBS beads and allowed to develop to a later stage, HH12-13. The resulting ectopic structures were then assessed for the expression of a neurofilament-associated protein recognised by the antibody 3A10. Hensen’s nodes derived from HH3-4 embryos and implanted in extraembryonic epiblast of HH3-3+ embryos induce ectopic neural axes expressing this neurofilament-associated antigen within this period of time (Storey et al., 1992). FGF beads however, do not induce 3A10positive cells within the same period (n=12; Fig. 3A) nor is this antigen detected around FGF beads implanted within the extraembryonic epiblast in ovo and cultured for a much longer period to HH20-24 (n=6; Fig. 3B,C). These data suggest that, while FGF can initiate the neural programme in extraembryonic epiblast cells, it is not sufficient to elicit neuronal differentiation, which therefore requires the action of additional signals. FGF DOES NOT INDUCE AXIAL MESODERM IN EXTRAEMBRYONIC EPIBLAST It is important to establish in this assay whether FGF beads induce posterior neural tissue directly or as a consequence of the prior induction of axial mesoderm. To test this, we therefore assayed for the induction of the Not-1 antigen, which specifically identifies the notochord from HH7 and for the expression of goosecoid, which at the time assessed is expressed in prechordal mesendoderm anterior to the emerging notochord as well as in the ventral diencephalon. Neither the Not-1 antigen (0/6 FGF beads, 0/4 PBS beads) nor goosecoid mRNA (0/7 FGF beads, 0/6 PBS) were induced in response to FGF or PBS beads placed in contact with the extraembryonic epiblast at HH3+ (Fig. 5A-C). Together these findings suggest that axial mesoderm is not formed in response to FGF. However, brachyury (bra) (Herrman et al., 1990; see Beddington et al., 1992 for review) is induced in response to FGF beads at HH3-3+ and in older HH4-4+ hosts (Table 2; Figs 4, 5C). Expression of this T-box-containing transcription factor is an immediate early response to FGF signalling (Smith et al., Table 2. FGF induces posterior, but not anterior neural genes in HH3-3+ hosts and does not elicit neural genes in older HH4-4+ host embryos tlx En2 Krox20 sax1 cash4 Bra HH3-3+FGF beads 0/2 (0%) 0/11 (0%) 0/8 (0%) 9/16 (56%) 7/10 (70%) 12/12 (100%) PBS beads 0/4 (0%) 0/4 (0%) 0/4 (0%) 0/8 (0%) 0/18 (0%) 0/6 (0%) HH4-4+FGF beads 0/5 (0%) 0/6 (0%) 0/6 (0%) 0/4 (0%) 0/4 (0%) 4/4 (100%) PBS beads 0/4 (0%) 0/4 (0%) 0/4 (0%) 0/4 (0%) 0/4 (0%) 0/4 (0%) Early posterior neural genes induced by FGF 477 Fig. 3. The neurofilament-associated antigen recognised by the antibody 3A10 is not present in ectopic structures induced by FGF beads implanted in extraembryonic epiblast at HH3-3+ and maintained in New culture to HH12-13 (A) or cultured in ovo to HH20-24 (B,C). (A) Whole-mount showing 3A10-positive neurons in the diencephalon and hindbrain of the host embryo and the absence of this antigen in tissue around the FGF bead (arrowhead); (B) 3A10-negative ectopic structure associated with a FGF bead (arrowhead) after 4 days culture in ovo, only brown background staining is visible; (C) Section of ectopic growth in B confirming the absence of the 3A10 antigen. Fig. 2. FGF beads, but not control PBS beads, induce expression of a combination of transcription factors characteristic of posterior neural tissue and maintain and/or induce expression of the L5 epitope in the extraembryonic epiblast (A-E,H). However, neither FGF beads nor PBS beads induce anterior neural markers (F,G,JL). In all cases, beads were implanted at HH3-3+ and gene expression assessed at HH8-9, except for (F,G,J-L) which were allowed to develop to HH10-12, when the host embryos express the marker genes assessed). Whole-mount in situ hybridisation techniques using DIG-labelled RNA probes were used to detect relevant genes, unless stated otherwise. (A) FGF beads induce local ectopic expression of Sax-1, which is expressed in prospective posterior neural tissue in the host embryo; (B) closeup view showing ectopic Sax-1 expression; (C) PBS beads do not induce expression of cash4 (shown here) or any other gene assessed; (D) cash4 is induced in epiblast cells close FGF beads; (E) Hoxb9 is induced in response to FGF beads; (F) The steroid hormone receptor tailless is expressed in the diencephalon and is not induced by FGF beads; (G) Close-up view of FGF bead in F showing absence of tailless expression; (H) The L5 antigen is maintained and/or induced by FGF signalling in the extraembryonic epiblast, as revealed by immunocytochemistry using a peroxidase conjugated secondary antibody; (I) Krox-20, a marker of rhombomeres 3 and 5 of the hindbrain is not induced by FGF beads; (J) Close up view of FGF bead in I showing absence of Krox20 expression; (K) Close up view of FGF bead in L showing absence of En2 expression, detection by immunocytochemistry with a peroxidase conjugated secondary antibody; (L) The posterior midbrain/anterior hindbrain marker Engrailed 2 is not induced by FGF beads (arrowhead). 1991; Isaacs et al., 1994) and in the chick Bra is expressed in presumptive mesodermal tissues of the primitive streak and subsequently in notochord and endodermal tissues as well as in the posterior CNS (Kispert et al., 1995; Knezevic et al., 1996). As Bra is only co-expressed with Delta-1 in the primitive streak, we combined in situ hybridization for Delta-1 with immunocytochemistry for the Bra antibody in order to identify cells that have characteristics of the prospective mesoderm. In most cases (16/17), Bra and Delta-1 are co-expressed in response to FGF. In many cases (8/16), they are expressed together in a block of cells at a distance from the bead which resembles the regressed primitive streak (Fig. 5D-F). However, not all Braexpressing cells co-express Delta-1 (Fig. 5F) and some Brapositive cells are also found in the immediate vicinity of the bead (Fig. 5E). Together, these findings indicate that, while ectopic FGF does not induce axial mesoderm, it can induce expression of genes characteristic of prospective mesoderm. Induction of early posterior neural genes is preceded by expression of brachyury and Delta-1 The presence of Bra/Delta-expressing cells in this assay indicates that prospective mesodermal cells might be induced by FGF and it is possible that, in turn, these cells produce signals that elicit expression of early posterior neural genes. We investigated this possibility further by assessing the temporal sequence in which bra, Delta-1 and cash4 are expressed. FGF induces bra and Delta-1 in extraembryonic epiblast cells within 6 hours, while cash4 is only expressed 10-12 hours after FGF 478 K. G. Storey and others is presented to extraembryonic epiblast cells (Table 3; Fig. 6AD). The expression of bra and Delta-1 is therefore a prelude to cash4 induction. It is thus possible that FGF signalling acts ‘indirectly’ via non-FGF signals provided by prospective mesodermal tissue, to induce early posterior neural genes. FGF4 induces Fgf8 In view of the different kinetics of bra/Delta-1 and cash4 induction by FGF signalling, we investigated the possibility that FGF4 might also induce activation of other FGF genes (as reported by Crossley et al., 1996b). We find that Fgf8 is induced by FGF4 beads in the extraembryonic epiblast within 8 hours and therefore prior to cash4 (Fig. 6E; Table 4). Fgf8 is normally expressed in both prospective mesoderm (the primitive streak) and also in the posterior neural plate where its domain of expression overlaps with Bra and cash4 as well as Sax1 (see below Fig. 7). This finding therefore identifies Fgf8 as a signal downstream of FGF4 signalling which may act ‘directly’ on or within prospective neural tissue to induce early posterior neural genes (see below). Bra and cash4 are co-expressed in a subset of epiblast cells After 6 hours incubation, Bra is expressed in cells surrounding FGF beads, however, by 12 hours cells closest to the bead now express cash4. In embryos in which cash4 in situ hybridisation is combined with the Bra antibody, we find that some cells express both cash4 and Bra (Fig. 6F). This suggests that exposure to FGF elicits a succession of gene expression from Bra to cash4, in the same cell population. Strikingly, this sequence of events mirrors the temporal pattern in which these two genes are normally expressed in prospective posterior neural tissue. Bra is expressed prior to the onset of cash4 at HH5-6 in Table 3. Response times of Bra, Fgf8 and cash4 to FGF signalling bra Delta-1 Fgf8 cash4 4 hours 6 hours 8 hours 10 hours 0/4 (0%) ND 0/4 (0%) 0/4 (0%) 8/8 (100%) 6/12 (50%) 0/4 (0%) ND 10/10 (100%) 10/10 (100%) 6/10 (60%) 0/8 (0%) ND ND ND 5/12 (71%) the epiblast adjacent to the primitive streak, (Fig. 3 in Kispert et al., 1995; Henrique et al., 1997) and, by HH8-9, the overlap between these two genes (and Sax-1 and Fgf8) is confined to the most recently formed posterior neural plate (Fig. 7A-J; and see Fig. 4 in Kispert et al., 1995). These findings further support the direct action of FGF signals on the prospective neuroepithelium, which could act via brachyury expression in this tissue. A specific subset of FGFs induce cash4 and Bra We assessed the ability of a variety of FGF family members with differing receptor specificities to induce expression of cash4 and Bra in order to establish the ligand-specificity of this effect and to deduce the possible receptor-mediated signalling pathways involved (Table 5). While FGF7 does not induce Bra or cash4, FGF2, FGF4 FGF9 and FGF8b all induce expression of both these genes. Given the binding specificities of these FGFs for specific receptors (reviewed by Orntiz et al., 1996), we can rule out transduction through FGFR2 (IIIb) which specifically mediates the activity of FGF7. Chick FGFR1 and FGFR2 are both expressed in the extraembryonic epiblast at HH3-3+, while FGFR3 is expressed most highly in the primitive streak and only weakly in surrounding epiblast cells (data not shown). The most likely receptors transducing FGF presented to extraembryonic epiblast are therefore the IIIC splice variants of FGFR1 and/or FGFR2. It is significant that FGF8b is able to induce cash4 as well as Bra, as this supports an endogenous role for this gene in the induction of early posterior neural genes. Concentration effects and cell movements in response to FGF Previous studies of the activity of FGF bound to heparin-coated beads (e.g., Cohn et al., 1995; Crossley et al., 1996) have not defined the concentration of FGF required for signalling. It is, however, important to establish that FGF-induced neural induction occurs at physiologically relevant concentrations of ligand (Slack, 1994). We soaked heparin-coated beads in a range of FGF4 concentrations (0.5-50 µg/ml) and assessed their ability to induce expression of Bra (Fig. 4). We find that beads soaked in a concentration between 0.5 and 5 µg/ml elicit Bra expression in half the cases examined, while a maximal response is achieved when beads are soaked in 50 µg/ml FGF4. By comparison, the data of Ornitz et al. (1996) indicate that Fig. 4. Heparin-coated beads were soaked in a range of FGF4 concentrations (µg/ml) and juxtaposed with extraembryonic epiblast cells in HH3-3+embryos, which were then cultured until they reached HH8-9. The activity of different concentrations of FGF presented by the beads was assessed in terms of the ability to induce expression of Bra, cash4, Sax-1, Krox20 and Tlx. While Bra is induced by FGF presented by beads soaked in 0.5 µg/ml. cash4 and Sax-1 are only expressed in response to higher FGF concentrations. Krox20 and tailless expression are not elicited by FGF signalling. (‘n’ values on columns tops). Table 4. A subset of FGFs induce expression of Bra and cash4 FGF2 FGF4 FGF7 FGF8a FGF8b FGF9 conc. Bra cash4 50 µg/ml 50 300 60 6 50 4/4 6/6 0/12 0/10 2/2 4/4 4/4 4/6 0/12 0/10 2/7 4/4 Early posterior neural genes induced by FGF the Ed50 for FGF4 activating FGF-R2c and R1c in transfected cell lines is about 0.5 nM. Combined with a clear doseresponse curve (Fig. 4), this suggests that FGF beads present physiological levels of ligand, although we cannot calculate the exact amount of FGF available to surrounding cells. FGFs bind locally to cell surface receptors and heparan sulphate proteoglycans present in the extracellular matrix (Wilkie et al., 1995). FGFs should therefore only be presented to cells in the immediate vicinity of a bead. In order to define the cell population directly exposed to FGFs in this assay, the distance of diffusion from heparin beads was assessed by immunocytochemical detection of digoxigenin (DIG)-labelled FGF2. While this growth factor is detected only on the surface of beads soaked in 0.5 µg/ml DIG-FGF (Fig. 8A), DIG-FGF is restricted to cells immediately adjacent to beads soaked in concentrations of 5 µg/ml (Fig. 8B). At 50 µg/ml DIG-labelled FGF2 is detected 4-6 cell diameters from the beads (Fig. 8C; also assessed in sections, not shown). The lower level of our immunocytochemical detection of DIG-FGF therefore correlates with the lowest concentrations at which we can detect Bra expression around a bead. Together, these data indicate that beads soaked in 50 µg/ml FGF present local physiological levels of growth factor. FGFs are known to stimulate both cell movement and proliferation (Wilkie et al., 1995). In order to define further the cell population exposed to FGF, we compared the movement of DiI-labelled extraembryonic epiblast cells in response to beads washed in PBS or soaked FGF. We observe no dramatic difference between the movement of cells in contact with PBS (n=14) or FGF beads (n=23). In both cases, DiI-labelling is found in the vicinity of the bead and in trails of cells that have moved away from the bead. Thus, although cells move away from FGF beads placed in the extraembryonic epiblast, this behaviour appears to reflect normal morphogenetic movements in this region, rather than a response evoked by FGF signalling (Fig. 8D,E). We also assessed the possibility that FGF-stimulated extraembryonic epiblast cells recruit cells from the embryonic epiblast. Epiblast cells at the lateral edge of the prospective neural plate were labelled with DiI at HH3-3+ and an FGF bead placed in the adjacent extraembryonic epiblast (Fig. 8F). In all cases (14/14), no DiI-labelled cells were found in the vicinity of the bead, following overnight culture (to HH8-9) (Fig. 8G). Thus, while cell proliferation may expand the population of cells responding to this local source of growth factor, we show that embryonic epiblast cells are not recruited in response to local application of FGF to the extraembryonic epiblast. DISCUSSION This is the first study to examine the role of FGF signalling in the induction of neural tissue in an amniote embryo. FGFs are expressed in the organiser region Hensen’s node/anterior primitive streak; a source of signals that induces both anterior and posterior neural tissue which subsequently undergoes neuronal differentiation. We demonstrate that, in comparison with Hensen’s node, FGF signalling at primitive streak stages leads to the induction of a very specific subset of genes associated with the laying down of the posterior nervous system. We show that ectopic FGF maintains and/or induces expression of the early neural marker L5 in extraembryonic epiblast cells and elicits the expression of the transcription factors cash4 and 479 Sax-1, genes expressed transiently during the formation of the posterior nervous system. Further, this early posterior neural tissue forms without the induction of anterior neural markers and does not undergo the complete neural programme culminating in neuronal differentiation. We present evidence for both direct and indirect actions of FGF signalling on the prospective neuroepithelium. While axial mesoderm does not form in response to FGF signalling, the co-expression of Bra and Delta1 in some cells suggests that presumptive mesodermal tissue is formed. Neural-inducing signals could therefore be provided indirectly by presumptive mesoderm. However, these ‘indirect’ signals may also be FGF-mediated as FGF4 beads induce Fgf8, which is expressed in both prospective mesoderm and the forming posterior neural plate. The co-localisation of Bra and cash4 in cells around FGF beads further suggests that the induction of posterior neural genes could be mediated directly via Bra expression in the prospective neuroepithelium. These findings therefore show that FGF signalling in an early amniote embryo can mimic a specific subset of node activities, which include the maintenance of neural competence as well as the induction of early posterior neural genes. The assay We have used the large and accessible chick embryo to analyse the effects of FGF activity in a spatiotemporally defined manner. By placing FGF-soaked heparin beads in contact with extraembryonic epiblast cells, previously shown to be competent to respond to endogenous neural-inducing signals, we have tested the neural-inducing activity of FGF. While the extraembryonic epiblast may not be a completely naïve tissue, it contrasts favourably with Xenopus animal cap ectoderm which contributes to embryonic structures and appears to be prepatterned (Sokol and Melton, 1991). In comparison, chick extraembryonic epiblast is fated to give rise to extraembryonic membranes and is physically remote from the embryonic axis. Our DiI-labelling studies also demonstrate that FGF beads placed in the extraembryonic epiblast do not recruit embryonic epiblast cells that might have been exposed to neural-inducing signals from Hensen’s node. Further, although only the medial half of the extraembryonic epiblast is competent to respond to neural-inducing signals (Storey et al., 1992), it is striking that ectopic gene expression is often found on the side of FGF beads furthest from the host embryo (see for example Figs 2A, 6B). This pattern is inconsistent with the influence of other neuralinducing signals emanating from the embryonic axis and may reflect the radial expansion of the embryo at these stages. By assessing the effects of FGF signalling with respect to the FGF concentration in which beads were soaked, we have also identified a ‘soaking’ concentration that delivers physiologically relevant levels of FGF. Further, using DIG-labelled FGF2, we show that at these concentrations FGF diffuses from the heparin bead and becomes associated with cells in its immediate vicinity. The induction of Fgf8 in response to FGF4, however, may relay FGF signals beyond this region. Indeed, Fgf8 expression, along with the morphogenetic movements of extraembryonic epiblast cells may account for patterns of gene expression that extend outside the domain defined by the spread of DIG-labelled FGFs. Comparison of the effects of six different FGF members has also allowed us to establish the ligand-specificity of observed effects and to deduce the possible receptor-mediated signalling pathways, which include IIIC splice variants of FGFR1 and/or 480 K. G. Storey and others Fig. 5. Axial mesoderm is not induced in response to FGF beads (A-C), but expression of Bra and Delta-1 is detected (D-F). (A) goosecoid expression marking the cells of the prechordal region (as well as overlying diencephalic tissue) is not induced by FGF beads; (B) the notochord-specific antigen Not-1 is shown in the host embryo; (C) Not-1 is not induced by FGF beads; (D) Delta-1 is expressed, frequently in a block of cells at a distance from the FGF bead; (E) Delta-1-expressing cells (purple) also express Bra (brown) as revealed by immunocytochemistry with the TN antibody following in situ hybridisation for Delta-1 (shown in D). Not all Bra-expressing cells co-express Delta-1, some Bra only positive cells are also found in the vicinity of the bead (arrowhead); (F) Section (level indicated by bar in E) through region of Delta-1-expressing cells in E, showing that a central group of Delta-1 cells co-express bra (arrowhead). FGFR2. Our study therefore further defines the extraembryonic epiblast assay and characterises this in vivo method of delivering secreted factors. FGF can initiate the neural programme in the chick embryo In common with grafts of Hensen’s node (Streit et al., 1997), FGF signalling is able to maintain and/or induce expression of the L5 antigen in the extraembryonic epiblast. The L5 antibody recognises the LeX oligosaccharide epitope present in two different glycoproteins one of which (L5220) is present early in extraembryonic tissue and is associated with neural competence, while the second (L5450) is a later pan-neural marker (Streit et al., 1996, 1997). As FGF beads implanted for only a short period (6-8 hours) are surrounded by an ectopic region of L5 expression, it is possible that FGF signalling maintains L5220. However, FGF may also induce neural tissue that subsequently expresses L5450. Thus, while these findings and the induction of neural-specific Fig. 6. Temporal sequence of gene expression (detected by in situ hybridisation) in response to FGF beads; bra and Delta-1 are induced prior to Fgf8 and all three are induced prior to cash4 (A-E, see Table 3). (A) bra is induced within 6 hours in response to FGF signalling (arrowhead); (B) Delta-1 is also present after 6 hours (although the signal shown here is stronger than that shown for bra in A, this is not typical and Delta-1 is detected in only half the cases assessed at 6h while is bra is always present, see Table 3); (C) cash4 is first detected around FGF beads (arrowhead) after 10-12 hours; (D) High magnification of ectopic cash4 expression around the FGF bead in C; (E) Fgf8 is first detected after 8 hours (arrowhead); (F) Following overnight culture (16-18 hours) cash4 and Bra are co-expressed in cells close to FGF beads (arrowhead); shown here in section following in situ hybridisation for cash4 (blue labelling; same bead is shown in Fig. 2C) in combination with immunocytochemistry with an anti-brachyury antibody (brown labelling). genes (see below) implicate FGF in neural competence, the specific maintenance of L5220 by FGF signalling has yet to be determined. Like Hensen’s node, FGF also induces ectopic expression of the transcription factors cash4 and Sax1, which are markers of early posterior neural tissue (Henrique et al., 1997; Spann et al., 1994). However, unlike organiser activity (Storey et al., 1992), FGF signalling in extraembryonic epiblast does not elicit the expression of the early neuronal marker recognised by the 3A10 antibody even in embryos cultured to HH24, suggesting that FGF does not trigger the complete neural programme leading to neuronal differentiation. The failure to induce neurons is consistent with the induction of only early posterior neural genes (cash4 and Sax-1). cash4 is expressed transiently in precursors of the posterior CNS in a rostrocaudal wave of expression that moves in concert with the regressing node/primitive streak and is induced by signals emanating from the node (Henrique et al., 1997). It can substitute for the neural specification activity of homologous genes (achaete and scute) in Drosophila and expands the neural plate Early posterior neural genes induced by FGF 481 Fig. 7. During normal development Bra, cash4, Sax-1 and Fgf8 are expressed in overlapping domains in the posterior neural plate at HH9-10 (A-J). (A) The posterior neural plate (white arrowhead) expresses bra as well as the cells in the regressing primitive streak (PS) and the notochord (black arrowhead); (B) section through A (white bar) showing expression of bra in both open neural plate and underlying mesoderm. (C) cash4 is expressed in the neural plate at this time, the caudal-most region of this domain overlaps with bra expression; (D) Section through B (white bar) showing cash4 expression in the open neural plate but not in underlying mesodermal tissues; (E) Sax-1 is expressed in a similar domain to cash4 in the posterior neural plate; (F) Section through E (white bar) showing Sax-1 expression confined to the neural plate; (G) Fgf8 is expressed in the regressing primitive streak and in the caudal end of the forming posterior neural plate in a domain that overlaps with cash4, Sax-1 and bra at HH9; (H) Section through (white bar) G showing Fgf8 expression in the mesodermal and prospective neural tissue; (I) By HH10 Fgf8 is clearly expressed in the elevating posterior neural folds; (J) Section through I (white bar) showing localisation of Fgf8 throughout neural folds as well as underlying mesodermal tissues. when overexpressed in the frog embryo (Henrique et al., 1997). The expression of cash4 in neural precursors in the chick, together with these functional studies therefore suggests that cash4 acts early to assign neural cell fate in posterior regions of the chick embryo. Our current findings suggest that expression of cash4 is however, insufficient to trigger the cascade of gene expression that leads to neuronal differentiation. During neural development cash4 is expressed prior to Delta1, which in the CNS identifies the first post-mitotic neurons (Henrique et al., 1995). In this assay, Delta-1 is always coexpressed with Bra (a combination indicative of prospective mesodermal tissue in the primitive streak; Fig. 5F) suggesting that post-mitotic neurons (which do not express Bra) do not form in response to FGF. Consistent with this interpretation, overexpression of Fgf8 in the developing midbrain leads to a dramatic expansion of the neural precursor population in the ventricular zone (Lee et al., 1997). It is therefore tempting to speculate that, in line with in vitro data (Gensburger et al., 1987; Murphy et al., 1994; Temple and Qian, 1995) a common function of FGF signalling in the developing nervous system is the induction and/or maintenance of precursor cells. Indeed, the generation of post-mitotic cells and their subsequent differentiation as neurons or glia may require the down regulation of cash4 and Sax-1 and the activity of other signals, such as retinoic acid, bone morphogenetic proteins, sonic hedgehog and neurotrophins (Papalopulu and Kintner, 1996; Sharpe and Cross, 1997; Liem et al., 1995; Shah et al., 1996; Averbuch-Heller et al., 1994; Roelink et al.,1995; Jungbluth et al., 1997). FGF signalling in this assay, therefore, appears to elicit only the initial steps in the neural programme, which includes maintenance of neural competence and the induction of a combination of transcription factors characteristic of posterior neural precursors. FGF does not induce anterior neural tissue FGF does not induce any of the anterior neural markers tested in this assay; these include genes expressed in the diencephalon (tailless), at the midbrain/hindbrain boundary (Engrailed 2) and in the hindbrain (Krox20). This suggests that neural tissue induced in response to FGF has a more posterior regional character, which may include posterior hindbrain (at least Fig. 8. Digoxigenin-labelled FGF2 was used to detect the spread of FGF from beads implanted in the extraembryonic epiblast (A-C), and DiI was used to follow the movement of cells exposed to FGF or PBS beads (D,E) as well as to test the possibility that FGF beads recruit embryonic epiblast cells from the host embryo (F,G). The spread of DIG-FGF2 from beads soaked in concentrations of (A) 0.5 µg/ml, (B) 5 µg/ml and (C) 50 µg/ml, was detected with an antiDIG-alkaline-phosphatase conjugated antibody following implantation in HH3-3+ embryos and overnight incubation in New culture. The dorsal surface of the extraembryonic epiblast cells was labelled with DiI and (D) PBS beads or (E) FGF beads were placed over the labelled cells and in HH3-3+ embryos, which were cultured overnight. Cells exposed to FGF did not appear to move more than those exposed to PBS beads; (F) DiI labelling of embryonic epiblast cells at the lateral edge of the prospective neural plate at HH3-3+ adjacent to a FGF bead (arrowhead); (G) Following overnight culture no DiI-labelled cells are recruited by FGF beads (arrowhead). 482 K. G. Storey and others below the level of Krox20 expression in rhombomere 5) and spinal cord, as judged by the induction of Hoxb9. The absence of more anterior neural markers might also reflect the undifferentiated state of this FGF-induced neural tissue, which is characterised by the expression of only early markers of the posterior neural precursor population. In this assay, anterior neural markers are also not induced by low FGF concentrations. This contrasts with reports in amphibian embryos in which high FGF concentrations elicit expression of posterior neural genes, while low levels of FGF induce anterior neural markers (Kengaku and Okamoto, 1995). However, these experiments were carried out with ectoderm cells which had been previously dissociated, a treatment that can in itself lead to the formation of anterior neural tissue (Grünz and Täcke, 1989; Godsave and Slack 1991; Wilson and Hemmati-Brivanlou, 1995). Indeed, Lamb and Harland (1995) report only weak concentration effects, but they do find anterior neural markers, such as Engrailed, induced when old (late gastrula) animal caps are exposed to FGF. Ectoderm in these experiments, however, also underwent treatment that may have led to the partial dissociation of cells. A further problem with these Xenopus experiments is that it is not possible to distinguish between multiple actions of FGF signalling during development. Using more precise methods afforded by the use of FGFcoated beads, our results show that in the chick there is a defined window of competence during which epiblast cells respond to FGF by expressing early posterior neural genes and that anterior neural genes are not induced in their stead in older epiblast cells. Further, using the same technique Crossley et al. (1996a) have demonstrated that much later during the regionalisation of the nervous system and formation of the midbrain, FGF signals do now elicit Engrailed expression. This emphasises, along with other studies (e.g. Shimamura and Rubenstein, 1997) that FGF signalling acts at specific times and in distinct regions of the developing nervous system. By showing that FGF signalling underlies the initial steps in the generation of posterior neural tissue, our study identifies a new role for such signals at primitive streak stages. Do FGF signals act directly or indirectly to induce early posterior neural genes? It is possible that non-FGF signals provided by mesodermal tissues induce the expression of early neural genes in this assay. Axial mesodermal tissue has been shown to provide neuralinducing signals in many vertebrates, including the chick embryo (Storey et al., 1995; A. Rowan, C. D. Stern and K. G. S., unpublished data). We demonstrate that FGF signalling in this assay does not induce markers of axial mesoderm, the prechordal plate marker goosecoid and the notochord-specific antigen Not-1, confirming observations made in the Xenopus embryo (Cho et al., 1991; Pownall et al., 1996). However, we do find that FGF beads induce the co-expression of genes characteristic of prospective para-axial mesoderm in the primitive streak (Delta-1 and Bra). Evidence for the indirect action of FGF Several lines of evidence suggest that early posterior neural genes could be induced indirectly by signals emanating from these prospective mesoderm cells: (i) bra and Delta-1 expression precede cash4 induction by at least 4 hours; and (ii) the competence to respond to FGF by expressing bra persists beyond that for cash4 and Sax-1 (Table 2). As neural competence is lost shortly after HH4 (Gallera, 1971; Storey et al., 1992; Streit et al., 1995), it may be that signals produced by bra/Delta-1-expressing tissue are not produced soon enough to elicit cash4 and Sax-1 expression. However, the proposition that these prospective mesoderm cells are a source of neural-inducing signals is not consistent with the results of grafting experiments that compare the neuralinducing abilities of prospective axial and non-axial mesoderm cells in the anterior primitive streak (Storey et al., 1995). These experiments show on the one hand that, while prospective axial mesoderm (medial mesendoderm) is a potent neural inducer, prospective non-axial mesoderm (posterolateral mesendoderm) does not induce neural tissue in extraembryonic epiblast cells (Storey et al., 1995). On the other hand, this failure to induce neural tissue was assessed only on morphological grounds and it may be, therefore, that prospective non-axial mesoderm can induce expression of early posterior neural genes. Evidence for direct effects In this context, it is significant that an early response to the FGF beads is the induction of Fgf8. During development Fgf8 is expressed in both this prospective non-axial mesoderm in the primitive streak, as well as later in adjacent developing posterior neural tissue in a domain that overlaps with those of cash4 and Sax-1 (Fig. 7G-I). It is therefore possible that FGF signalling mediates the action of non-axial mesoderm and also that it may act directly within the posterior neural plate. We show that Fgf8 is induced prior to cash4 in response to FGF beads and also that FGF8 protein can induce cash4 (as well as bra) identifying it as a potential endogenous regulator of this early neural gene. The presence of cells that co-express Bra and cash4 close to FGF beads (Fig. 6F) further suggests that direct action of FGFs on prospective neural tissue could be mediated via Bra expression in this tissue. This Bra-expressing cell population most likely corresponds to those Bra-positive cells close to FGF beads that do not co-express Delta-1 (Fig. 5E). In fact, the spatiotemporal pattern in which Bra and cash4 are expressed in the embryo during the laying down of the posterior nervous system suggests that bra is a prelude to the expression of cash4 in the same cell population (Fig. 7A-D) (Kispert et al., 1995; Henrique et al., 1997). Strikingly, a requirement for bra for the maintenance of Sax-1 expression has been demonstrated in the T-mutant mouse (Schubert et al., 1995), although this may reflect the failure to form mesodermal tissue as well the absence of bra in the posterior neural plate. A role for bra in the induction of early posterior neural genes might also account for the formation of an apparently normal posterior nervous system in transgenic frogs in which FGFR1-mediated-signalling is blocked at early gastrula stages, but after the induction of bra (Kroll and Amaya, 1996). The concentration effects that we observe (Fig. 4) may also support the direct action of FGFs on prospective neuroepithelium. These data show that bra is induced by both low and high FGF doses while posterior neural genes are only induced at high FGF concentrations. This is consistent with the early expression of bra in the vicinity of the bead and its later expression at a distance, while cash4 is expressed close to FGF beads. One interpretation of these results is that cells close to the FGF bead initially express bra, but later acquire a neural fate. Alternatively, cells may initially turn on mesodermal genes, move away and induce neural tissue in their wake. Early posterior neural genes induced by FGF However, as DiI-labelling of the extraembryonic epiblast exposed to FGF (or PBS) beads shows that some cells do remain in the vicinity of these beads while others move away we cannot distinguish between these two possibilities. In short, we find evidence for both direct and indirect effects of FGF signalling on prospective neural tissue. These two routes are not mutually exclusive. Indeed, given the induction of Fgf8 in this assay and its expression in both prospective mesodermal and neural tissue, direct and indirect effects could be mediated by the same signal in this region of the embryo. The role of FGF in neural induction Our results implicate FGF signalling in neural competence and show that FGF signalling can elicit expression of early posterior neural genes in the absence of anterior neural tissue. This suggests that the initial steps in the generation of posterior neural tissue take place downstream of FGF signalling independently of other node-derived signals. This contrasts with many studies in the Xenopus embryo which show that initial neural-inducing signals (such as Noggin and Chordin) elicit formation of anterior neural tissue and that this step is a prerequisite for the later induction of posterior neural genes (Nieuwkoop et al., 1952; reviewed by Hemmati-Brivanlou and Melton, 1997; see Taira et al., 1997). It is also striking that in our assay FGF signalling elicits only early and transient markers of prospective posterior neural tissue and it is possible that expression of these genes may not be indicative of caudal neural character, but rather reflects the induction of a particular cell state associated with the formation of neural precursors in this region. In conclusion, while our data strongly suggest that FGF signalling functions during neural induction and that it specifically plays a role in the generation of precursors of the posterior nervous system, spatiotemporally defined loss-offunction experiments are required to confirm these activities. The early posterior neural genes induced in this assay are present only transiently; their expression ceases when the cells of the posterior neural plate are left behind by the regressing Hensen’s node/FGF source and no longer express Fgf8. It will be intriguing to assess the regulation of this transition in the context of signals that instigate later phases of the neural programme leading to neuronal differentiation. We thank Helen Skaer, Jonathan Slack, Claudio Stern and Andrea Streit for critical reading of the manuscript. We are grateful to Yosef Gruenbaum for the gift of Sax-1 probe; Bernard Herrman for the gift of the Bra antibody; Domingos Henrique for gifts of cash4 and Delta1 plasmids; Robb Krumlauf for the gift of Hoxb9 plasmid; Gail Martin and Clive Dickson for the gift of mouse Fgf8 plasmids; Jim Smith for the gift of the bra plasmid and to Andrea Streit for the L5 antibody. This work was supported by Joint MRC small project grant for the MRC to K. G. S. and J. K. H. and by grants from the Wellcome Trust to J. K. H. and K. G. S. REFERENCES Abud, H. E. (1995). In Examination of methods for the study of FGFs during mouse development. D. Phil. Thesis, University of Oxford. Averbuch-Heller, L., Pruginin, M., Kahane, N., Tsoulfas, P., Parada, L., Rosenthal, A. and Kalcheim, C. (1994). Neurotrophin-3 stimulates the differentiation of motoneurons from avian neural tube progenitor cells. Proc. Natl. Acad. Sci. USA 91, 3247-3251. Beddington, R. S. P. (1994). Induction of a second neural axis by the mouse node. Development 120, 613-620. 483 Beddington, R. S. P., Rashbass, P. and Wilson, V. (1992). Brachyury – a gene affecting mouse gastrulation and early organogenesis. Development 1992 supplement, 157-165. Campos-Ortego, J. (1993). Early neurogenesis in Drosophila melanogaster. In The Development of Drosophila (ed. M. Bat and A. Martinez Arias) Vol. 2. pp 1132-1172. New York: Cold Spring Harbour Laboratory Press. Campuzano, S. and Modolell, J. (1992). Patterning of the Drosophila nervous system: the achaete-scute gene complex. Trends. Genet. 8, 202-208. Cho, K. W. Y., Blumberg, B., Steinbreisser, H. and De Robertis, E. M. (1991). Molecular nature of Spemann’s organiser: the role of the Xenopus homeobox gene goosecoid. Cell 7, 1111-1120. Cohn, M. J., Izpisua-Belmonte, J-C, Abud, H., Heath, J. K. and Tickle, C. (1995). Fibroblast growth factors induce additional limb development from the flank of chick embryos. Cell 80, 739-746. Cox, W. G. and Hemmati-Brivanlou, A. (1995). Caudalisation of neural fate by tissue recombination and bFGF. Development. 121, 4349-4358. Crossley, P. H. and Martin, G. R. (1995). The mouse Fgf8 gene encodes a family of polypeptides and is expressed in regions that direct outgrowth and patterning in the developing embryo. Development. 121, 439-451. Crossley, P. H., Martinez, S. and Martin, G. R. (1996a). Midbrain development induced by FGF8 in the chick embryo. Nature. 380, 66-68. Crossley, P. H., Minowada, G., MacArthur, C. A. and Martin, G. R. (1996b). Roles for FGF8 in the induction, initiation, and maintenance of chick limb development. Cell 84, 127-136. Dias, M. and Schoenwolf, G. C. (1990). Formation of ectopic neuroepithelium in chick blastoderms : age related capacities of induction and self-differentiation following transplantation of quail Hensens’s node. Anat. Rec. 229, 437-448. Deng, C-X., Wynshaw-Boris, A., Shen, M., Daugherty, C., Orntiz, D. and Leder, P. (1994). Murine FGFR-1 is required for early postimplantation growth and axial organisation. Genes Dev. 8, 3045-3057. Doniach, T. (1995). Basic FGF as an Inducer of Anteroposterior Neural pattern. Cell 83, 1067-1070. Feldman, B., Poueymirou, W., Papaioannou, V. E., DeChiara,T. M. and Goldfarb, M. (1995). Requirement of FGF4 for post-implantation mouse development. Science 267, 246-249. Gallera, J. (1971). Primary induction in birds. Adv. Morph. 9, 149-180. Gensburger, C., Labourdette, G. and Sensenbrenner, M. (1987). Brain basic fibroblast growth factor stimulates the proliferation of rat neuronal precursor cells in vitro. FEBS Lett. 217, 1-5. Gleizes, P. E., Noaillac-Depeyre, J. and Gas, N. (1994). Labelling of basic fibroblast growth factor with digoxigenin: a non-radioactive probe for biochemical and cytological applications. Anal. Biochem. 219, 360-7. Godsave, S. and Slack, J. M. W. (1991). Single cell analysis of mesoderm formation in the Xenopus embryo. Development 111, 523-530. Grunz, H. and Tacke, L. (1989). Neural differentiation of Xenopus laevis ectoderm takes place after disaggregation and delayed reaggregation without inducer. Cell Diff. Dev. 28, 211-218. Hamburger, V. and Hamilton, H. L. (1951). A series of normal stages in the development of the chick embryo. J. Morph. 88, 49-92. Heikinheimo, M., Lawshe, A., Shakleford, G. M., Wilson, D. B. and MacArthur, C. A. (1994). FGF8 expression in the post-gastrulation mouse suggests roles in the development of the face limbs and central nervous system. Mech. Dev. 48, 129-138. Hemmati-Brivanlou, A. and Melton, D. (1997). Vertebrate Embryonic Cells Will Become Nerve cells Unless Told Otherwise. Cell 88, 13-17. Henrique, D. Tyler, D. Kintner, C. Heath, J. K., Lewis, J., Ish-Horowicz, D. and Storey, K. G. (1997). cash4 a novel achaete-scute homologue induced by Hensen’s node during the generation of the posterior nervous system. Genes Dev. 11, 603-611. Henrique, D., Adam, J., Myatt, A., Chitnis, A., Lewis, J. and IshHorowicz, D. H. (1995). Expression of a Delta homologue in prospective neurons in the chick. Nature 375, 787-790. Herrmann, B. G., Labeit, S. Poustka, A., King, T. R. and Lehrach, H. (1990). Cloning of the T gene required in mesoderm formation in the mouse embryo. Development 113, 891-911. Isaacs, H. V., Pownall, M. E. and Slack, J. M. (1994). eFGF regulates Xbra expression during Xenopus gastrulation. EMBO J. 13, 4469-4481. Izpisua-Belmonte, J. C., De-Robertis, E. M., Storey, K. G. and Stern, C. D. (1993). The homeobox gene goosecoid and the origin of organiser cells in the early chick blastoderm. Cell 74, 645-659. Jungbluth, S. Koentges, G. and Lumsden, A. (1997). Coordination of early neural tube development by BDNF/trkB. Development 124, 1877-1885. Kengaku, M. and Okamoto, H. (1993). Basic fibroblast growth factor induces 484 K. G. Storey and others differentiation of neural tube and neural crest lineages of cultured ectoderm cells from Xenopus gastrula. Development 119, 1067-1078. Kengaku, M. and Okamoto, H. (1995). bFGF as a possible morphogen for the anteroposterior axis of the central nervous system in Xenopus. Development 121, 3121-3130. Kispert, A., Ortner, H., Cooke, J. and Herrmann, B. G. (1995). The chick Bra gene: Developmental expression pattern and response to axial induction by localised activin. Dev. Biol. 171, 458-470. Knezevic, V., De santo, R. and Mackem, S. (1996). T-box genes in chick gastrulation. Development 123, 411-419. Kroll, K. L. and Amaya, E. (1996). Transgenic Xenopus embryos from sperm nuclear transplantations reveal FGF signalling requirements during gastrulation. Development 122, 3173-3183. Lamb, T. M. and Harland, R. M. (1995). Fibroblast growth factor is a direct neural inducer, which combined with noggin generates anterior-posterior neural pattern. Development 121, 3627-3636. Launay, C., Fromentoux, V., Shi, D-L. and Boucaut, J-C. (1996). A truncated FGF-receptor blocks neural induction by endogenous Xenopus inducers. Development 122, 869-880. Lawson, K. A., Meneses, J. J. and Perdersen, R. A. (1991). Clonal analysis of the epiblast during germ layer formation in the mouse embryo. Development 113, 891-911. Lee, S. M. K., Danielian, P. S., Fritzsch, B. and McMahon, A. P. (1997). Evidence that FGF8 signalling from the midbrain-hindbrain junction regulates growth and polarity in the developing midbrain. Development 124, 959-969. Liem, K. F., Tremml G., Roelink, H. and Jessell, T. M. (1995). Dorsal differentiation of neural plate cells induced by BMP-mediated signals from epidermal ectoderm. Cell 82, 969-979. Mahmood, R. Bresnick, J., Hornbruch, A., Mahony, C., Morton, N., Colquhoun, K., Martin, P., Lumsden, A., Dickson, C. and Mason, I. (1995a). A role for FGF-8 in the initiation and maintenance of vertebrate limb bud outgrowth. Curr. Biol. 5, 797-806. Mahmood, R., Kiefer, P., Guthrie, S., Dickson, C. and Mason, I. (1995b). Multiple roles for FGF-3 during cranial neural development in the chicken. Development 121, 1399-1410. Murphy, M., Reid, K., Ford, M., Furness, J. B. and Bartlett, P. F. (1994). FGF2 regulates proliferation of neural crest cells, with subsequent neuronal differentiation regulated by LIF or related factors. Development 120, 35193528. New, D. A. T. (1955). A technique for the cultivation of the chick embryo in vitro. J. Embryol. Exp. Morph. 3, 326-331. Nieuwkoop, P. D., Boterenbrood, E. C., Kremer, A., Bloesma, F. F. S. N., Hoessels, E. L. M. J., Meyer, G. and Verheyen, F. J. (1952). Activation and organisation of the central nervous system in amphibians. J. Exp. Zool. 120, 1-108. Ornitz, D. M., Xu, J., Colvin, J. S., McEwen, D. G., MacArthur, C., Coulier, F., Gao, G. and Goldfarb, M., (1966). Receptor specificity of the fibroblast growth factor family. J. Biol. Chem. 271, 1592-15297. Patel, K., Connolly, D. J., Amthor, H., Nose, K. and Cooke, J. (1996). Cloning and early dorsal axial expression of Flik, a chick follistatin-related gene: evidence for involvement in dorsalization/neural induction. Dev. Biol. 178, 327-342. Patel, N. H., Martin-Blanco, E., Coleman, K. G., Poole, S. J., Ellis, M. C., Kornberg, T. B. and Goodman, C. S. (1989). Expression of engrailed proteins in arthropods, annelids and chordates. Cell 58, 955-968. Papalopulu, N. and Kintner, C. (1996). A posteriorising factor, retinoic acid, reveals that anteroposterior patterning controls the timing of neuronal differentiation in Xenopus neuroectoderm. Development 122, 3409-3418 Pownall, M. E., Tucker, A. S., Slack, J. M. W. and Isaacs, H. V. (1996). eFGF, Xcad3, and Hox genes form a molecular pathway that establishes the anteroposterior axis in Xenopus. Development 122, 3881-3892. Riese, J., Zeller, R. and Dono, R. (1995). Nucleo-cytoplasmic translocation and secretion of fibroblast growth factor-2 during avian gastrulation. Mech. Dev. 49, 13-22. Roberts, C., Platt, N., Streit, A., Schachner, M. and Stern, C. D. (1991). The L5 epitope: an early marker for neural induction in the chick embryo and its involvement in inductive interactions. Development 112, 959-970. Roelink,H., Porter, J. A., Chiang, C., Tanabe, Y., Chang, D. T., Beachy, P. A. and Jessell, T. M. (1995). Floor plate and motor neuron induction by different concentrations of the amino-terminal cleavage product of sonic hedgehog autoproteolysis. Cell 81, 445-455. Sasai, Y. Lu, B. Piccolo, S. and De Robertis, E. M. (1996). Endoderm induction by the organiser secreted factors chordin and noggin in the Xenopus animal caps. EMBO J. 15, 4547-4555. Schubert, F. R., Fainsod, A., Gruenbaum, Y. and Gruss, P. (1995). Expression of a novel murine homeobox gene Sax-1 in the developing nervous system. Mech. Dev. 51, 99-114. Sechrist, J. and Bronner-Fraser, M. (1991). Birth and differentiation of reticular neurons in the chick hindbrain: Ontogeny of the first neuronal population. Neuron 7, 947-963. Shah, N., Groves, A. and Anderson, D. J. (1996). Alternative neural crest fates are instructively promoted by TGFb superfamily members. Cell 85, 331-343. Sharpe, C. R. and Goldstone, K. (1997). Retinoid receptors promote primary neurogenesis in Xenopus. Development 124, 515-523. Shimamura, K. and Rubenstein, J. L. (1997). Inductive interactions direct early regionalisation of the mouse forebrain. Development 124, 2709-2718. Slack, J. M. W. (1994). Inducing factors in Xenopus early embryos. Current Biology 4, 116-125. Slack, J. M. W. and Tannahill, D. (1992). Mechanisms of anteroposterior axis specification in vertebrates: lessons from amphibians. Development 114, 285-302. Smith, J. C., Price, B. M. J., Green, J. B. A., Weigel, D. and Herrmann, B. G. (1991). Expression of the Xenopus homologue of Bra (T) is an immediate early response to mesoderm induction. Cell 67, 79-87. Sokol, S. and Melton, D. A. (1991). Pre-existent pattern in Xenopus animal cap pole cells revealed by induction with activin. Nature 351, 409-11. Spann, P., Ginsburg, M., Rangini, Z., Fainsod, A., Eyal-Giladi, H. and Gruenbaum, Y. (1994). The spatial and temporal dynamics of Sax1(CHox3) homeobox gene expression in the chick’s spinal cord. Development 120, 1817-1828. Stern, C. D. (1990). The marginal zone and its contribution to the hypoblast and primitive streak of the chick embryo. Development 109, 667-682. Storey, K. G., Crossley, J. M., De Robertis, E. M., Norris, W. E. and Stern, C. D. (1992). Neural induction and regionalisation in the chick embryo. Development 114, 729-741. Storey, K. G., Selleck, M. A. and Stern, C. D. (1995). Neural induction and regionalisation by different subpopulations of cells in Hensen’s node. Development 121, 417-428. Streit, A., Stern, C. D., Thery, C., Ireland, G. W., Aparicio, S., Sharpe, M. J. and Gherardi, E. (1995). A role for HGF/SF in neural induction and its expression in Hensen’s node during gastrulation. Development 121, 813824. Streit, A.,Yuen, C. T., Loveless, R. W., Lawson, A. M., Finne, J., Schmitz, B., Feizi, T. and Stern, C. D. (1996). The Lex carbohydrate sequence is recognised by the antibody L5, a functional antigen in early neural development. J. Neurochem. 66, 834-844 Streit, A., Socknathan, S., Pérez, L. Rex, M., Scotting, P. J., Sharpe, P. T., Lovell-Badge, R. and Stern, C. D. (1997). Preventing loss of competence for neural induction: HGF/SF, L5 and Sox2. Development 124, 1191-1202. Taira, M., Saint-Jeannet, J.-P. and Dawid, I. (1997). Role of the Xlim-1 and XBra genes in anteroposterior patterning of neural tissue by the head and trunk organsier. Proc. Natl. Acad. Sci. USA 94, 895-900. Temple, S. and Qian, X. (1995). bFGF, neurotrophins and the control of cortical neurogenesis. Neuron 15, 249-252. Thomas, P. and Beddington, R. (1996). Anterior primitive endoderm may be responsible for patterning the anterior neural plate in the mouse embryo. Current Biol. 6, 1487-1496. Waddington, C. H. (1932). Experiments on the development of chick and duck embryos cultivated in vitro. Phil. Trans. Roy. Soc. Lon. B 221, 179230. Wilkie, A. O. M., Morriss-Kay, G. M., Jones, E. Y. and Heath, J. K. (1995). Functions of fibroblast growth factors and their receptors. Curr. Biol. 5, 500507. Wilkinson, D. G., Bhatt, S., Chavrier, P., Bravo, R. and Charnay, P. (1989). Segment-specific expression of a zinc-finger gene in the developing nervous system of the mouse. Nature 112, 959-970. Wilson, P. A. and Hemmati-Brivanlou, A. (1995). Induction of epidermis and inhibition of neural cell fate by BMP-4. Nature 376, 331-333. Yamada, T., Placzek, M., Tanaka, H., Dodd, J. and Jessell, T. M. (1991). Control of cell pattern in the developing nervous system: polarising activity of the floorplate and notochord. Cell 64, 635-647. Yamaguchi, T., Harpal, K., Henkemeyer, M. and Rossant, J. (1994). Fgfr1is required for embryonic growth and mesoedrm patterning during mouse gastrulation. Genes Dev. 8, 3032-3044. Yu, R. T., McKeown, M., Evans, R. M. and Umesono, K. (1994). Relationship between Drosophila gap gene tailless and a vertebrate nuclear receptor Tlx. Nature 370, 375-379..