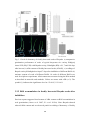
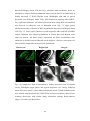
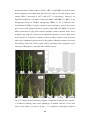
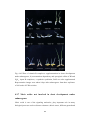
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Hong Kong Baptist University HKBU Institutional Repository Open Access Theses and Dissertations Electronic Theses and Dissertations 2013 Molecular studies of some developmental and reproductive traits of Rayada rice Rubaiyath Bin Rahman A N M Hong Kong Baptist University Follow this and additional works at: http://repository.hkbu.edu.hk/etd_oa Recommended Citation Bin Rahman A N M, Rubaiyath, "Molecular studies of some developmental and reproductive traits of Rayada rice" (2013). Open Access Theses and Dissertations. Paper 51. This Thesis is brought to you for free and open access by the Electronic Theses and Dissertations at HKBU Institutional Repository. It has been accepted for inclusion in Open Access Theses and Dissertations by an authorized administrator of HKBU Institutional Repository. For more information, please contact [email protected]. Molecular Studies of Some Developmental and Reproductive Traits of Rayada Rice BIN RAHMAN A N M Rubaiyath A thesis submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy Principal supervisor: Prof. ZHANG Jianhua Hong Kong Baptist University November 2013 Declaration I hereby declare that this thesis represents my own work which has been done after registration for the degree of PhD at Hong Kong Baptist University, and has not been previously included in a thesis or dissertation submitted to this or any other institution for a degree, diploma or other qualifications. Signature Date: November 2013 i ii Abstract Crop domestication and subsequent breeding or directional selection have narrowed the genetic diversity of elite varieties whereas land races, ecotypes, wild relatives growing on native preferences still keep genetic diversities of stress tolerances. Rayada is such an exceptional ecotype, variant of typical deepwater rice, completely endemic to certain areas of Madhumati river tracts of Bangladesh and still shares some features of wild rices. Multiple physiological features of Rayadas are distinctly different from typical deepwater rice. In this PhD project, we have studied the specialty of Rayada rice and identified that Rayada has special tolerances to prolonged flood, submergence and cold along with longer root system and prompt recovery capacity after water stress. All these features make it as an elite resource of stress tolerance and might become a new focus of rice germplasm research. Among all deepwater rices, Rayada is the only exception, having virtually no seed dormancy, but both physiological and molecular bases of this trait are completely unknown. We examined the non-dormant nature of Rayadas as a natural variant of deepwater rice. After comparing features of freshly harvested seeds of Rayada with those of typical deepwater rice variety, we identified several concerted features; for instance, less ABA content in freshly harvested seeds; faster ABA catabolism and enhanced ROS accumulation after imbibition. Moreover, after analyzing stepwise gene expressions of 32 bZIPs in seed germination, mild and severe water stresses among three extreme ecotypes including Rayadas together with homology search with reported genes, we identified OsbZIP84 as a candidate gene for the regulation of ABA catabolism in Rayada rice. ABA content and expression analysis of OsbZIP84 and ABA8oxs in four growth and developmental stages along with phenotyping of mutant revealed the function of OSbZIP84 in the dormancy regulation of Rayada rice. Submergence tolerance during seed germination is one of the rare traits of rice, even among cereals. Except few physiological indications of tolerance, most other molecular signaling network is not known. We identified several positive and negative regulators of shoot development under submergence inducting the capacity of shoot development of Rayada rice under oxidative stress. We iii successfully developed a condition supplemented with riboflavin and H2O2 where intolerant genotypes successfully developed shoot under submergence. However, induced shoot development was completely inhibited by glucose, ABA and mitochondrial complex IV inhibitor signifying ABA and glucose as negative regulators, whereas ROS, riboflavin and mitochondrial complex IV as positive regulators. Gene supplementation expression mimicked analysis aerobic of gene α-amylases expression revealed pattern. H2O2 Plausible mechanisms of riboflavin and H2O2 function in submergence tolerance were also discussed. Finally, we isolated a novel mutant of Rayada variety with Kaladigha background and having four interesting phenotypes of practical implications. Mutant plant shows purple pigmentation throughout the plants organs along with dense and elongated trichomes on the adaxial leaf surface. In addition, the same mutant also shows high frequency of stigma exsertion. But ultimately, we observed that the mutant plant is completely sterile. The possible reason of the sterility was found being related to the stigma receptivity. Severe reduction of ROS accumulation in stigmas of mutant plant was observed after fluorescent H2DCF-DA staining. However, pollen grains are completely viable with normal shape and size. Interestingly, the fertility was partially restored after humidifying the panicles. Mutant progeny showed dense black coloration in seeds with significant reduction of grain weight. Moreover, it showed segregating ratio of 3:1 for purple pigmentation, suggesting single gene mutation nature. Other phenotypic features confirmed the mutant as a Rayada variety with Kaladigha background, not a seed contamination. After extensive data mining of these four phenotypes, we identified maize Lc gene with three similar phenotypes reported earlier excluding stigma exsertion, hence considered as candidate gene of this mutant. The gene expression of maize Lc homolog of rice, OsbHLH13, was exceptionally upregulated in the purple mutant. Further studies of genetic characterization may open up the practical implications of this mesmerizing mutant. In summary, Rayada is a primitive deepwater rice ecotype that can offer many traits and genetic resources that are badly needed in rice breeding for stress tolerance and the time is mature to do the more detailed research with rapid advances in genome research weaponry . iv Acknowledgements I would like to express my deepest gratitude to my principal supervisor, Professor Jianhua Zhang for his supervision, encouragement, supports and inspiration over the past three years. Without his support, guidance and care, I could never run my projects as well as my life smoothly in abroad. If any credit earns this thesis, naturally it belongs to him too. Similarly, I want to thank my co-supervisor, Professor Yiji Xia for his advice, guidance as well as research assistantship he offered me. Lucky me, have such two notable scientists as my supervisors. Special thanks to our Dean of Science, Professor Tang Tao, for his generosity as well as arrangement of prayer room after my request. Certainly, thanks to all of my teachers of Biology Department, HKBU, especially to Chris Wong, Head of the department. All of you are so kind and cooperative. I would remember Professor Ricky Wong ever, for his cordial behavior, I met him in my very first day of HKBU, apart my supervisor. Nothing would satisfy me unless I thank Hong Kong Baptist University as well as the Research Grant Council for studentship of my PhD tenure. Sincere gratitude to all of the scientific officers of Biology department who helped me in different ways in my studies, especially to Ms. Olivia Chau Ching Wah and Louise Lai Ha Ng. Similarly, thanks to Ho Yee, Fion, Pui Ling, Lo Kam Fai for their cooperation and support. My thesis completion would be impossible without the help my lab mates. I must remember you all. You were too kind to me, made my life easy abroad, I am grateful to you all, especially to Dr. Ye Nenghui, Jia Liguo, Shi Lu, Liu Rui, Zhi Hui. All others including Professor Xia’s group are also acknowledged for their favor and attention. Hey, Larry! I wished to thank you some three years ago; Google was my dependence since the beginning of the study. While I was alone as an alien in the Sino Island, you played music for me. I was never been to library last three years, only because of your awesome services of Google scholar, Note, YouTube and v certainly Google itself. I would be miser if I do not acknowledge you with my gratefulness. It’s my privilege to acknowledge the best scientists of the world, the farmers, especially rice farmers who kept Rayada rice still growing in the field. I just revealed scientifically whatever you know since long. Finally, I do dedicate my thesis to my beloved parents, my lovely wife and to my two little angels: Rudaba & Namira. Without your love, affection, encouragement, spirit I would never finish it. vi Table of Contents Declaration i Abstract iii Acknowledgements v Table of Contents vii List of Figures and Tables xii List of Abbreviations xv Chapter 1 Introduction 1 1.1 General Introduction 1 1.2 Rice ecosystems 1 1.2.1 Irrigated lowland ecosystem 2 1.2.2 Rainfed lowland ecosystem 2 1.2.3 Upland rice ecosystem 3 1.2.4 Flood-prone ecosystem 3 1.2.4.1 Rayada ecotype 1.3 4 Molecular mechanisms of adaptation to flood-prone area 5 1.3.1 Prolonged flood with partial submergence 5 1.3.2 Flash flood of complete submergence 6 1.4 Seed dormancy and germination: two interrelated complex processes 7 1.5 ABA 8’-hydroxylases: key player after imbibition 11 1.6 ABA signaling pathway 12 1.7 ABA responsive transcription factors 13 1.7.1 bZIP transcription factors 14 1.7.2 bZIPs in rice 15 1.8 Crosstalk between ROS and ABA signaling 16 1.9 Seed germination under submergence 17 1.10 Anthocyanin biosynthesis and trichome development: linked 19 processes 1.11 Objectives of this study 22 vii Chapter 2 Rayada specialty: the forgotten resource of elite features of 23 rice 2.1 Introduction 23 2.2 Deepwater rice in Bangladesh 24 2.3 Types of deepwater rice in Bangladesh 25 2.3.1 Rayada rice 25 2.4 Distribution of Rayada 27 2.5 Yearlong life cycle 28 2.6 Elongation ability of Rayada 29 2.7 Submergence tolerance 31 2.8 Seed dormancy 32 2.9 Cold tolerance 34 2.10 Longer root systems of Rayada 35 2.11 Photoperiod sensitivity 36 2.12 Quick recovery after water stress 38 2.13 Origin and evolution of Rayada 39 2.14 Rayada: the forgotten resource of elite features of rice 41 Chapter 3 OsbZIP84 is involved in seed dormancy regulation of 43 Rayada rice through ABA catabolism 3.1 Introduction 43 3.2 Materials and methods 46 3.3 3.2.1 Plant materials 46 3.2.2 Growth conditions and stress treatments 46 3.2.3 Microarray data analysis 53 3.2.4 RNA extraction and quantitative real-time (qRT)-PCR 53 3.2.5 Extraction and measurement of ABA 53 3.2.6 ROS staining and confocal laser scanning microscopy 54 3.2.7 Statistical analysis 54 Results 55 3.3.1 Lack of dormancy of freshly harvested Rayada seeds 55 3.3.2 ROS accumulation in freshly harvested Rayada seeds after 56 imbibition 3.3.3 Differential expressions of viii selected OsbZIPs in seed 58 germination 3.3.4 Expression pattern of OsbZIP84 in mild and severe water 60 stresses 3.4 3.3.5 Homology search and possible functional homolog 62 3.3.6 Putative functions of VIP1or RSG homolog of rice 63 3.3.6.1 Inundation 64 3.3.6.2 Seed maturation 65 3.3.6.3 Water stress recovery 67 3.3.6.4 Seed germination 68 3.3.7 Dormancy phenotype of osbzip84 69 3.3.8 Elevated ABA in osbzip84 seeds but reversed in water stress 71 Discussion 73 3.4.1 Natural variation of seed dormancy 73 3.4.2 ABA, ROS and seed dormancy: linked altogether 74 3.4.3 Regulation of ABA catabolism: function of OsbZIP84 in 75 Rayada rice 3.4.4 Elevated ABA in osbzip84 seed results in dormancy phenotype Chapter 4 Shoot development under submergence is regulated by ROS 76 79 and riboflavin in Rayada rice 4.1 Introduction 79 4.1.1 Submergence tolerance during seed germination: a rare trait 80 4.1.2 Physiological features of tolerant genotypes 80 4.1.3 Alternative strategy to reveal factors regulating shoot 81 development 4.2 4.3 Materials and methods 82 4.2.1 Plant materials and growth conditions 82 4.2.2 Different treatments 82 4.2.3 Uncoating of Rayada seeds 83 4.2.4 Electric conductivity measurement 83 4.2.5 RNA extraction and quantitative real-time (qRT)-PCR 84 4.2.6 Extraction and measurement of ABA 84 Results 86 4.3.1 High ABA restricts shoot development, not root ix 86 4.3.2 Rayada seedlings: more tolerant to composite stress 87 4.3.3 Augmentation of seed germination by hydrogen peroxide 89 4.3.4 Shoot development of Rayadas under composite submergence 90 stress 4.3.5 Coatless Rayada seeds unable to develop shoot under 92 submergence 4.3.6 Riboflavin induces shoot development under submergence 94 4.3.7 Nitric oxide: not involved in in shoot development under 95 submergence 4.3.8 Both exogenous ABA and glucose restrict shoot development 96 under submergence 4.3.9 Mitochondrial complex IV inhibition: restrict seed germination 98 under submergence 4.3.10 H2O2 affects carbohydrate metabolism under submergence 4.4 99 Discussion 100 4.4.1 Rayada rice: is elixir of stress tolerances? 100 4.4.2 Riboflavin richness of red rice 101 4.4.3 Riboflavin: a signaling compound 101 4.4.4 How might riboflavin induce shoot development in 102 submergence? 4.4.5 Hidden player masked riboflavin 103 4.4.6 Involvement of ABA and glucose signaling 104 Chapter 5 A novel natural mutant of Rayada rice, plausibly in 107 OsbHLH13, shows multiple phenotypes with potential use in hybrid seed technology 5.1 5.2 Introduction 107 5.1.1 Stigma exsertion 107 5.1.2 Morphological markers 108 5.1.3 Trichome development 108 5.1.4 Anthocyanin and trichome development: linked processes 109 Materials and methods 110 5.2.1 Plant materials and growth condition 110 5.2.2 Determination of total anthocyanin content 110 x 5.3 5.2.3 RNA extraction and real-time PCR analysis 111 5.2.4 Estimation of trichome density and distribution 111 5.2.5 Determination of stigma exsertion 111 5.2.6 Pollen viability assay 112 5.2.7 Determination of stigma receptivity 112 5.2.8 Humidification of panicles 112 5.2.9 ROS staining and confocal laser scanning microscopy 113 Results 114 5.3.1 Phenotypic characterization of natural purple mutant 5.3.1.1 Purple pigmentation 114 5.3.1.1.1 Anthocyanin content 5.4 114 116 5.3.1.2 Profuse and elongated trichomes 117 5.3.1.3 Stigma exsertion 119 5.3.1.4 Sterility of purple mutant 120 5.3.2 Stigma receptivity: plausible cause of sterility 120 5.3.3 Reduced ROS accumulation in mutant stigma 121 5.3.4 Partial restoration of fertility 124 5.3.5 Phenotype of purple progeny 125 5.3.6 Candidate gene of theses phenotypes 125 5.3.7 Expression analysis of OsbHLH13 126 Discussion 128 5.4.1 Natural mutation, not seed contamination 128 5.4.2 Sterility and reduced ROS accumulation of stigmas: linked 129 together 5.4.3 Humidity and sterility 130 5.4.4 Black colored but reduced grain weight 130 5.4.4 Selection of OsbHLH13 as candidate gene 131 Chapter 6 General Discussion and Conclusion 133 References 139 xi List of Figures and Tables Fig. 1.1 Distribution of rice cultivation among different ecosystems in 2 Asia Fig. 1.2 Total cultivation area of rice in different ecosystems in top five 4 rice producing countries in mid 1990s Fig. 1.3 Two different strategies of survival of rice after flooding 6 Fig. 1.4 Vivipary or preharvest sprouting of different crops 9 Fig. 1.5 Generalized model of mechanism of seed dormancy induction 10 and release Fig. 1.6 Simplified ABA biosynthetic and catabolic pathways in plants 12 Fig. 1.7 Model on the major ABA signaling pathway 13 Fig. 1.8 Submergence tolerant cultivars during seed germination and 18 phenotype of comparative tolerance Fig. 1.9 Simplified anthocyanin biosynthesis and trichome development 21 regulation Fig. 2.1 Flood-affected areas of Bangladesh 23 Fig. 2.2 Status of deepwater rice cultivation in Bangladesh 24 Table 1 Comparative features of different types of deepwater and wild 26 rice (O. rufipogon) of Bangladesh Fig. 2.3 Distribution of Rayada rice in Bangladesh 28 Fig. 2.4 Internode elongation pattern of Rayada after inundation 30 Fig. 2.5 Comparative submergence tolerance of Sub1A introgressed 32 variety, BRRI 52 with Rayada variety, Kaladigha Fig. 2.6 Climatic features of Rayada growing areas 33 Fig. 2.7 Comparative phenotype of cold tolerance of Rayada variety, 34 Kaladigha with upland rice, Manikmodhu Fig. 2.8 Comparative root growth of Rayadas with others varieties in 36 different conditions Fig. 2.9 Comparative time course leaf derolling after rewatering of water 38 stressed Rayada rice variety, Kaladigha and aus genotype, Dudsor Fig. 2.10 Phylogenetic analysis of snorkel protein sequences among xii 40 deepwater and wild rices Table 2 List of primer sequences used in this study 47 Fig. 3.1 Lack of dormancy of freshly harvested seeds of Rayada 56 Fig. 3.2 Comparative ROS accumulation of freshly harvested seeds 57 Fig. 3.3 Expression analyses 59 of selected OsbZIPs among three ecotypes/varieties during seed germination Fig. 3.4 Expression profile of OsbZIP84 after mild, severe water stresses 61 and recovery in three extreme ecotypes/varieties Fig. 3.5 Phylogenetic tree of OsbZIP84 homolog among wild and 63 cultivated species Fig. 3.6 Time course ABA content and selected genes expression after 65 inundation in Rayada variety, Kaladigha Fig. 3.7 Time course ABA content and selected genes expression during 66 seed maturation in Rayada variety, Kaladigha Fig. 3.8 Time course ABA content and ABA8ox1 and OsbZIP84 67 expression during water stress recovery in Rayada variety, Kaladigha Fig. 3.9 Time course ABA content and gene expression of ABA8ox1, 68 OsbZIP84 and water absorption pattern during imbibition in Rayada variety, Kaladigha and dose dependent effect of ABA on seed germination of Kaladigha Fig. 3.10 Time course relative gene expression of NCEDs after imbibition 69 Fig. 3.11 Germination and seedling growth performance of osbzip84 70 Fig. 3.12 Change of ABA content, grain size and weight of osbzip84 72 Fig. 4.1 87 Effect of high ABA and mild salinity on seed germination of Rayadas Fig. 4.2 Composite stress tolerance capacity of Rayadas to modern rice 88 varieties Fig. 4.3 Effect of H2O2 on seed germination, coleoptile growth and ABA 90 content Fig. 4.4 Exceptional submergence tolerance capacity of Rayada varieties 91 under composite submerged stress Fig. 4.5 Effect of seed coat on germination and electric conductivity xiii 93 Fig. 4.6 Effect of vitamin-B complexes supplementation in shoot 95 development under submergence Fig. 4.7 Effect of SNP either singly or in combination with H2O2 under 96 submergence Fig. 4.8 Effect of glucose, ABA and osbzip84 on shoot development 97 under submergence Fig. 4.9 Effect of mitochondrial complex IV inhibition on shoot 98 development under submergence Fig. 4.10 Gene expression patterns of selected amylases in either 99 coleoptile or caryopsis after submergence Fig. 5.1 Comparative pigmentation of purple mutant with control plants Fig. 5.2 Comparative total anthocyanin content in different parts of both 116 115 control and purple mutants Fig. 5.3 Trichome density and distribution of control and purple mutant 118 Fig. 5.4 Stigma exsertion of purple mutant 119 Fig. 5.5 Percentage of seed setting in control, purple and humidified 120 purple mutant plants Fig. 5.6 Comparative reproductive features of mutant plants Fig. 5.7 ROS accumulation of whole stigma and stigmatic papillae level 123 121 in both control and purple mutant plants Fig. 5.8 Seed setting in purple mutant panicles after humidification 124 Fig. 5.9 Phenotype of purple mutant progeny 125 Fig. 5.10 Relative expression OsbHLH13 in leaf, leaf sheath and 126 internode of both control and mutant plants xiv List of Abbreviations ABA abscisic acid ABA8ox ABA 8'-hydroxylase ABA-GE ABA-glucose ester ABF ABRE binding factor abi ABA insensitive ABRE ABA-responsive element ano anoxia ATP adenosine 5'-triphosphate B1 thiamine B2 riboflavin B3 niacin B5 pantothenic acid B6 pyridoxine BAP 6-benzylaminopurine bHLH basic helix-loop-helix BRRI Bangladesh Rice Research Institute BVP basic vegetative phase bZIP basic leucine zipper CaMV cauliflower mosaic virus Col Columbia Cvi Cape Verde Islands cpm counts per minute xv CYP cytochrome DAH days after harvest Din diniconazole DNA deoxyribonucleic acid DOG delay of germination DPA dihydrophaseic acid DPI diphenylene iodonium DWR deepwater rice EC electric conductivity EGL enhancer of glabrous era enhanced response to exogenous ABA ERF ethylene-responsive factor EUI elongated uppermost internode FAD flavin adenine dinucleotide FMN flavin mononucleotide FR flood resistant FW fresh weight GA gibberellic acid GA20ox gibberellin 20 oxidase GDP gross domestic product GL glabrous GOI gene of interest GWAS genome-wide association study H2DCF-DA 2',7'-dichlorodihydrofluorescein diacetate xvi H2O2 hydrogen peroxide HBSS Hanks’ balanced salt solution IRRI International Rice Research Institute KD Kaladigha Lc leaf color min minute NADH nicotinamide adenine dinucleotide NaN3 sodium azide NCED 9-cis-epoxycarotenoid dioxygenase NO nitric oxide O2- superoxide radical OH. hydroxyl radical PA phaseic acid PAC1 pale aleurone color 1 PCD programmed cell death PCR polymerase chain reaction PEG polyethylene glycol PHS preharvest sprouting POSTECH Pohang University of Science and Technology PP2C protein phosphatase 2C PSP photoperiod sensitive phase PYL PYR1-like PYR1 pyrabactin resistance1 qRT-PCR quantitative reverse transcription PCR xvii QTL quantitative trait loci RAP-DB rice annotation project -database rboh respiratory burst oxidase RCAR regulatory components of ABA receptors RIA radioimmunoassay rin ripening inhibitor RLD root length density RNA ribonucleic acid ROS reactive oxygen species rpm revolution per minute RSG repression of shoot growth RWC relative water content SAPK stress activated protein kinase SD standard deviation sdr seed dormancy rice sec second SK snorkel SLR1 slender rice-1 SLRL1 slr1 like-1 SNF1 sucrose nonfermenting 1 SNP single nucleotide polymorphism SNP sodium nitroprusside SnRK2 SNF1-related protein kinase 2 SS Sarsari xviii Sub1 submergence 1 TF transcription factor TMV tobacco mosaic virus TT transparent testa TTG transparent testa glabra VIP VirE2 interacting protein YD6 Yang Dao 6 xix xx Chapter 1 Introduction 1.1 General Introduction Rice (Oryza sativa L.) belongs to the family Poaceae. It is the single most important crop of primary food source for half of the world’s population. Interestingly, nearly all, i.e. 90% rice is produced in Asia (Maclean et al. 2002). Rice accounts for 35% to 75% of the calories consumed by more than 3 billion Asians. Moreover, rice is also the single most important food source of the poor. It accounts over 70% of the daily calorie intake in some developing countries like Bangladesh, Cambodia, Laos, Myanmar (Khush 1997). The majority of the population in rice-producing areas, particularly in many Asian and African countries, is still suffering from hunger, malnutrition and extreme poverty (Hirano et al. 2008). It is projected that global rice production need to be doubled by 2030 to cope with the impending population increase (Li and Zhang 2013). This will not be possible without new discoveries in breeding and growing methods as well as development of new plant types with high yield potential (Varshney and Koebner 2006). Scientifically, rice is also very important since being a model crop plant offering various advantages for functional genomics research (Varshney and Koebner 2006). Moreover, information obtained from rice is also helpful for studying other cereal crops as these seem to have a monophyletic origin (Clark et al. 1995). 1.2 Rice ecosystems Rice grows in different ecosystems. It can be classified in different ways. Some countries like Bangladesh , classify rice ecosystems based on growing seasons and it includes 1) boro (winter rice), 2) aus (summer rice), 3) transplanted aman (autumn rice), 4) broadcast aman (typical deepwater rice-DWR), 5) Rayada (long duration DWR), 6) ashwina (photoinsensitive DWR) and 7) hill rice (Khush 1997). Out of 7, three of them are flood-prone rice ecosystems i.e. deepwater rice ecosystems. However, International Rice Research Institute (IRRI) classifies rice ecosystems into four broad groups: i) irrigated, ii) rainfed lowland, iii) upland and 1 iv) flood-prone ecosystems (Fig.1.1) (Maclean et al. 2002). The distribution of rice cultivation among these four ecosystems in Asia is shown in Figure 1.1. Fig. 1.1 Distribution of rice cultivation among different ecosystems in Asia. (a) irrigated lowland, (b) rainfed lowland, (c) upland, and (d) flood‐prone (deepwater) rice. Adopted from Bouman et al. (2007). 1.2.1 Irrigated lowland ecosystem Half of the global rice cultivated area covers irrigated ecosystem, accounts 75% of global rice production (Fig. 1.1a). Generally semi-dwarf improved modern varieties are cultivated in irrigated rice ecosystems. Genetic diversity of irrigated rice ecosystem has been significantly narrowed due to high yielding varietal dominance (Maclean et al. 2002). 1.2.2 Rainfed lowland ecosystem Rainfed lowland ecosystem comprising about 34% of the global rice cultivated area (Fig. 1.2b), is non-irrigated but lack of water control hence suffer from both 2 flood and drought, occasionally. The rainfed lowland rice ecosystem again can be divided into four sub-ecosystems: (1) favorable rainfed lowland, (2) droughtprone, (3) submergence-prone, and (4) both drought and submergence-prone. Both traditional and modern varieties are cultivated in these ecosystems depending on risk prone nature of the respective areas (Maclean et al. 2002). 1.2.3 Upland rice ecosystem Globally 13% of total world rice area with only 4% total world rice production is from upland rice ecosystem (Fig. 1.1c). Since traditional cultivars are cultivated in upland ecosystem hence overall productivity is very low (1 ton/hectare) (Maclean et al. 2002). Several common features such as fine and large root systems, quick recovery after stress, prompt stomata closure are associated with drought tolerance, frequently found in in upland rice; hence considered as source of drought tolerant novel genes. 1.2.4 Flood-prone ecosystem Finally, the flood-prone ecosystem is the complex environment for rice production for being variation of nature, depth and timing of flooding. This ecosystem consists of several subtypes: 1) deepwater - flooding exceeding 100 cm for 10 days to several months, 2) flash flood (more than 10 days), 3) costal tidal areas, 4) problem soils due to excess water (Maclean et al. 2002). The main production areas of deepwater rices are the deltas of the Ganges-Bramaputra river of India and Bangladesh, the Mekong river of Vietnam and Cambodia, the Irrawady River in Myanmar and the Chao Phraya river of Thailand (Fig. 1.1d) (Maclean et al. 2002). Over 100 million people in South and Southeast Asia largely depends on this ecosystem for their livelihood (Bhuiyan et al 2004). Deepwater rices are exclusively local varieties or landraces. Generally, before the monsoon, deepwater rice fields are broadcasted with traditional varieties. This rice ecosystem represents about 8% of the total rice cultivation area. However, in Bangladesh, almost 12 % of rice cultivation area under flood-prone ecosystem, highest among rice producing countries (Fig. 1.1d and 1.2) (Maclean et al. 2002). Bangladesh is rich in rice biodiversity due to its wide variation of land, topography, and seasons. Almost 700 deepwater rice genotypes have been 3 collected from Bangladesh and stored in the BRRI and IRRI gene banks (Bashar et al. 2004). Among them, Rayada rice is a specialized ecotype of deepwater rice, completely endemic to certain area of Bangladesh (Perez and Nasiruddin 1974; Catling 1992; Bashar et al. 2004). Fig. 1.2 Total cultivation area of rice in different ecosystems in top five rice producing countries in mid 1990s (Data adopted from Huke and Huke (1997). 1.2.4.1 Rayada ecotype Genetic variation is the raw materials of evolution; define the genetic basis of phenotypes. Ecotype is a population of a species developed with distinct morphological or physiological characteristics due to specific environment and persists even after changed to different environments (Lowry 2012). Ecotypes of extreme environment or habitat are good source of novel genes because of natural selection of adaptive mechanisms for survival in such extreme conditions whereas artificial breeding activity has directed selection towards increasing economic yield of cultivated species (Alonso-Blanco et al. 2009). So crop domestication and subsequent selection or breeding narrowed the genetic diversity of most cultivated varieties, hence ecotypes with endemic nature could still be good source of genetic variation. Rayada is such an exceptional ecotype of deepwater rice, completely endemic to only specific areas of Bangladesh (Perez and Nasiruddin 4 1974; Catling 1992; Bashar et al. 2004). Multiple distinct physiological features of Rayadas are different than typical deepwater rice varieties. In addition, still it shares some features of wild rices (Khush 1997). Tolerances to prolonged flood, submergence and cold are special features along with strong photosensitivity and lack of dormancy (Perez and Nasiruddin 1974; Catling 1992; Bashar et al. 2004). Moreover, longer root system (Gowda et al. 2012) and prompt recovery make it as an elite resource of stress tolerances. However, it has long been neglected because of its yearlong life cycle and poor yield. Rayada type varieties might be eliminated due to these two undesired traits (Vergara and Chang 1985). In this thesis, we focus on Rayada specialty discussed in Chapter 2 and following three chapters for specific traits with detailed studies. 1.3 Molecular mechanisms of adaptation to flood-prone area Based on water depth, duration and geological pattern, flooding of rice fields can be classified into two types: 1) flash flood, shallow depth with weeks duration and 2) deepwater or prolong flood of several months. Interestingly, some rice ecotypes or land races posess efficient mechanisms of complete withstand both partial and complete submergence. 1.3.1 Prolonged flood with partial submergence All deepwater rices possess the special capacity of internode elongation after flooding and efficiently adapted to prolong flooding of several months. Wild rice, O. rufipogon is also able to withstand prolong flood by faster internode elongation. Three hormones, ethylene, ABA and GA play key role to internode elongation or adaptation to partial submergence. Hattori et al. (2009) identified the Snorkel1 (sk1) and Snorkel2 (sk2) QTLs from deepwater rice with strong effect of internode elongation. Over-expression of SK1 and SK2, driven by the rice actin promoter, in non-deepwater rice result internode elongation, even in air. Snorkel (SK) genes encoding ERF transcription factors, allow rice to adapt in deepwater condition by enhancing GA responsiveness by an unknown manner (Fig. 1.3a) 5 (Hattori et al. 2009). However, the GA content increase in the deepwater rice after flooding is slight (Hoffmann-Benning and Kende 1992; Hattori et al. 2009). Moreover, still, how the snorkel genes enhance GA responsiveness is unknown. Fig. 1.3 Two different strategies of survival of rice after flooding. a) deepwater rices escape flooding stress by elongation of internodes mediated by ethylene and snorkel genes that improves GA responsiveness , b) flash flood tolerant rice overcome stress by quiescence strategy using SUB1A that restricts GA responsiveness through GA repressors SLR1 and SLRL1 (adopted from Voesenek and Bailey-Serres 2009) 1.3.2 Flash flood of complete submergence Unfortunately, most modern varieties are highly susceptible to flash flood; die within 7 days after complete submergence. However, some land races can tolerate complete submergence up to 2 weeks using complete opposite mechanism of deepwater rice; restrain elongation after submergence. FR13A is most tolerant genotypes; 100% survival after 7 days submergence of 10 days seedlings. Several physiological features are associated with complete submergence tolerance. These are keeping high carbohydrate concentration with least elongation and optimized alcoholic fermentation, up-regulation of antioxidant systems after desubmergence and low synthesis or sensitivity to ethylene. 6 The QTL studies after cross between submergence tolerant line (IR 40931-26) derived from FR13A with sensitive japonica line (PI543851) successfully identified Sub1 QTL located on chromosome 9 that confer the submergence tolerance (Bailey-Serres et al. 2010). This QTL defined 70% of the phenotypic variance of submergence tolerance where FR13A was the donor of the phenotype. In this locus, there are three similar genes, Sub1A, Sub1B and Sub1C. Recent studies identified that submergence tolerant is governed by submergenceinducible Sub1A gene encodes an ethylene response factor (ERF) -type transcription factor. However, other 2 genes (Sub1B and Sub1C) are also present in some submergence intolerant varieties, not involved in submergence tolerance. Sub1A decreases ABA content and significantly restricts ethylene stimulated GA responsiveness by increased the accumulation of the GA signaling repressors, Slender Rice-1 (SLR1) and SLR1 Like-1 (SLRL1) (Fig. 1.3b) (Xu et al. 2006; Bailey-Serres et al. 2010). Interestingly, submergence tolerance capacity was observed in 1950s, systematically screened in 1970s, finally Sub1A introgressed into high yielding varieties (Swarna, Samba Mahsuri, IR64, Thadokkam 1, CR1009, and BR11) and released recently in several countries including Bangladesh (Bailey-Serres et al. 2010). So it takes 60 years to successful varietal development journey of submergence tolerance from tolerant landraces. So the tolerance to complete and partial submergence use complete opposite strategy to overcome such stresses. Remarkably, similar type ERF transcription factors (group VII of ERF subfamily) regulate these features where SKs increase GA responsiveness, oppositely Sub1A restrict GA responsiveness. Interestingly, in both cases reduction of ABA is a general phenomenon (Fig. 1.3a,b). 1.4 Seed dormancy and germination: two interrelated complex processes Seed dormancy and germination are two complex interrelated processes of plant development. Seed germination is defined as physiological process culminating in the emergence of the embryo after imbibition of dry quiescent seed and subsequently become active metabolically and initiates the embryonic radicle and 7 or coleoptile growth (Bewley et al. 2013). Oppositely, seed dormancy is defined as inherent inability of germination under favorable condition of germination. Most plants seed at maturity are usually dormant. Seed dormancy could be several consequences such as physiological, morphological, morpho-physiological, physical and combinational (Finch-Savage and Leubner-Metzger 2006). In wild plants, seeds dormancy is a common phenomenon. It ensures wild plants to survive in unfavorable conditions. However, most cultivated species either loss or alleviate the seed dormancy period to some extent for synchronous germination along with rapid seedling establishment (Finch-Savage and Leubner-Metzger 2006). Lack of seed dormancy is a trait of crop domestication. Since in earlier days rice was grown once a year (single crop) in most of tropical and subtropical Asia hence, strong dormancy was not a problem. However, after availability of short-duration, photoperiod insensitive varieties and the development of irrigation facilities, double cropping of rice became a widespread practice in Asia. So strong and prolong seed dormancy are not suitable with double cropping system (Khush and Virk 2005). The acquisition of germination capacity during seed development or complete lack of dormancy can cause preharvest sprouting (PHS) or vivipary. It means the germination occurs in humid conditions while seed is still attached to mother plants, before the seed dispersal. Preharvest sprouting causes a great problem in some cereals (e.g. rice, wheat, barley, sorghum) (Fig. 1.4a-d). However, vivipary also can be equally an adaptive feature, for instance mangrove plant Rhizophora mangle (Bewley et al. 2013). ABA biosynthesis or signaling mutants usually result vivipary phenotypes. Key ABA biosynthesis gene, NCED mutant of maize (vp14) shows vivipary phenotype (Fig. 1.4a). Similarly, ABA signaling mutant vp1 or abi3 also shows vivipary. Recently, Sdr4, a QTL locus has been identified in rice determining seed dormancy through the global regulator of seed maturation player VP1. Wild rice O. rufipogon possesses Sdr4-k allele whereas japonica type (Sdr4-n) shows at least two mutations (Fig. 1.4d-f) (Sugimoto et al. 2010). Most seed dormancy is acquired during seed development and maturation when the seed is still attached with mother plants. Maternally supplied ABA contributes to seed maturation and desiccation. Dormancy is started early maturation stage 8 and continues to increase till fully mature seed (Fig. 1.5) (Raz et al. 2001). At maturity both desiccation tolerance and primary dormancy establish. Several hormones, most importantly, ABA plays important roles in seed development and maturation. It includes a) promotion of accumulation of seed storage proteins, b) acquisition of desiccation tolerance, c) induction and maintenance of dormancy (Bewley et al. 2013). Fig. 1.4 Vivipary or preharvest sprouting of different crops. (a) vp14 of maize, (b) wheat, (c) rin tomato, (d) vivipary-susceptible japonca cv. Nipponbare, (e) vivipary-resistant indica cv. Kasalath and (f) the near isogenic Nipponbare line carrying dormancy QTL (Sdr4-k) from Kasalath. Figures adopted from Bewley et al. (2013). Generally, mutants of ABA biosynthesis or deficiency of ABA during seed development lead to lack of primary seed dormancy. Oppositely, over-expressing ABA biosynthesis genes result higher ABA accumulation in seeds, thus enhance seed dormancy as well as delay seed germination. Moreover, regulation of ABA catabolism also plays equal importance of acquisition and maintenance of seed dormancy through regulating ABA content (Bewley et al. 2013). However, strong and prolong dormancy is also regulated by the native capacity of ABA 9 biosynthesis of seed itself, not the maternal ABA source only, during development and maturation (Holdsworth et al. 2008). In most studied plants, ABA content increases during initial seed development and decrease at maturity. However, seeds of arabidopsis, rapeseed, cotton, and barley show two ABA peaks, one around mid-maturity another one at late maturation (Bewley et al. 2013). ABA synthesis of both maternal and zygotic tissue contributes first peak whereas last peak is due to ABA synthesized by from zygotic tissues that regulates the induction and maintenance of dormancy. Apart ABA content, responsiveness to ABA also play important role in dormancy. ABA-insensitive mutants (abi1, abi2, and abi3) also show varying degree of reduced dormancy. Oppositely, hypersensitivity of ABA (era1) enhances seed dormancy. Recently, a major QTL for dormancy in arabidopsis reveals the accumulation of DOG1 protein during seed maturation and remain stable throughout seed storage and imbibition (Fig. 1.5) (Nakabayashi et al. 2012). Low temperature enhances the DOG1 expression (Kendall et al. 2011). Fig. 1.5 Generalized model of mechanism of seed dormancy induction and release. Seed dormancy trends to increase during seed maturation with active involvement of ABA whereas decrease during seed storage or after-ripening period. GA and ABA act antagonistically, GA enhances germination whereas 10 ABA restricts. ROS accumulation decreases the dormant period (adopted from Graeber et al. 2012). 1.5 ABA 8’-hydroxylases: key player after imbibition ABA content depends on the balance among ABA biosynthesis, catabolism and conjugation (Fig. 1.6). ABA catabolism is abundantly reported on regulation of ABA homeostasis in many plants such as barley (Chono et al. 2006; Millar et al. 2006), Arabidopsis (Millar et al. 2006; Okamoto et al. 2006; Umezawa et al. 2006; Okamoto et al. 2010), bean (Yang and Zeevaart 2006), maize (Ren et al. 2007) and rice (Yang and Choi 2006; Saika et al. 2007; Zhu et al. 2009; Ye et al. 2012a). However, after imbibition no significant up-expression of ABA biosynthesis occurs, rather ABA catabolic genes up-regulate before radicle emergence, even after exogenous ABA application during imbibition (Kushiro et al. 2004; Liu et al. 2010; Ye et al. 2012a). CYP707A2 is responsible for rapid decrease of ABA during imbibition in Arabidopsis, and expectedly, cyp707a2 mutant shows hyperdormancy phenotype, accumulates 6 fold higher ABA than normal (Kushiro et al. 2004). Moreover, triple mutant of ABA catabolic genes cyp707a1a2a3 accumulates 70 fold higher ABA thus shows deep seed dormancy (Okamoto et al. 2010). ABA catabolic pathway is rather simple than biosynthesis (Ye et al. 2012b). Among the hydroxylated products of C-7’, C-8’, and C-9’, only the 8’-hydroxyABA can be spontaneously isomerized to phaseic acid (PA) (Fig. 1.6) then further reduced to dihydrophaseic acid (DPA) (Nambara and MarionPoll, 2005). DPA does not have any biological activity of ABA. In rice, there are three ABA 8’-hydroxylases (Saika et al. 2007; Yang and Choi 2006). Among them, OsABA8ox3 is suppressed by glucose and delay glucose induced delay in seed germination (Zhu et al. 2009). OsABA8ox1 higher up-regulates after reentering of water stressed plants (Ye et al. 2011). Moreover, up-regulation of ABA8ox2 and 3 is also reported earlier in rice seed germination. Apart ABA oxidation, ABA can be conjugated with sugar to form ABA-glucose ester (ABAGE) catalyzed by ABA glucosyltransferase (Fig. 1.6). Role of ABA catabolic genes in dormancy regulation is already established, however, no clear indication of ABA conjugation in dormancy regulation yet been established through functional analysis of mutants or overexpressing lines. Neither atbg1 (Lee et al. 11 2006) nor bg2 (Xu et al. 2012) shows dormancy phenotype hence the role of ABA conjugation in seed dormancy is more unlikely. Fig. 1.6 Simplified ABA biosynthetic and catabolic pathways in plants. The collective effect of biosynthesis, inactivation through 8’ hydroxylation and conjugation defines the ABA content and physiological effect (adopted from Seki et al. 2007). 1.6 ABA signaling pathway Significant progress has been achieved in the last couple of years in understanding ABA perception and signaling pathway, especially after identification of its receptors in Arabidopsis. Generalized ABA signaling pathway is illustrated in Fig. 1.7. Two key regulators of ABA signaling are protein phosphatase 2C (group A PP2C) and SNF1-related protein kinase 2 (subclass III SnRK2). SnRK2 is positive regulator of the signaling pathway whereas PP2C acts negatively (Umezawa et al. 2010). Recent reports suggest that ABA signaling starts with binding of ABA to the ABA receptors RCARs/PYR1/PYLs leads to inactivation of type 2C protein phosphatases. The inactivation of protein phosphatases results the activation of 12 SnRK2 that targets ABA-dependent gene expression and ion channels (Raghavendra et al. 2010). So under normal conditions, PP2C constantly suppress kinase activity of SnRK2 through dephosphorylation, thus repress ABA signaling (Fig. 1.7). But when the soluble ABA receptor recognizes the ABA molecule, it binds PP2C and inhibits its phosphatase activity. Then SnRK2 is activated and phosphorylates its downstream substrates, including bZIP transcription factors (AREB/ABFs) etc., to activate ABA-responsive gene expression or other responses (Fig. 1.7) (Umezawa et al. 2010). Kobayashi et al. (2004) have identified 10 SnRK2 protein kinases in the rice genome and all family members are activated by hyperosmotic stress but only three (SAPK 8, SAPK 9 and SAPK 10) are activated by ABA. Fig. 1.7 Model on the major ABA signaling pathway. Detail description in the text (Adopted from Umezawa et al. 2010) 13 1.7 ABA responsive transcription factors Transcription factors (TFs) are any protein that takes part in DNA transcription due to its ability to bind to DNA sequences but not the part of DNA polymerases. Transcriptional control of gene expression is one of the key regulatory processes of genes expression involved in many biological processes of plant growth and development. Transcription regulators are thought to be the evolutionary switches or dominant class of genes changed during the course of domestication (Doebley and Lukens 1998; Doebley et al. 2006). In rice, a total of 1611 transcription factor encoding genes comprises around 2.6% in rice genome (Xiong et al. 2005). Plenty of experiments have identified several transcription factors family that are upregulated in ABA upregulated situations such water and salinity stress. It includes bZIP, zinc finger proteins, bHLH, NAC, MYB, MYC, WRKY etc. (Rabbani et al. 2003; Shinozaki and YamaguchiShinozaki 2007 ; Chen et al. 2002; Seki et al. 2003; Oh et al. 2009). Among these different transcription factor families, most responsive to ABA signaling as well as water stress are bZIP, NAC, and MYB (Ashraf 2010). Moreover, some ciselements, bound by these transcription factors, have been identified. ABAresponsive element (ABRE, PyACGTGGC), an important cis-element, is associated with the transcriptional regulation of ABA- and/or stress-responsive genes. In plants, ABRE binding factors (ABFs)/ABRE binding proteins (AREBs) are basic region leucine zipper (bZIP) class TFs (Jakoby et al. 2002) . In our study, we selected bZIP as candidate transcription factor family. 1.7.1 bZIP transcription factors The bZIP proteins compose a large family of transcription factors having bZIP domain composed of a basic region and a leucine zipper (Jakoby et al. 2002). The more conserved basic region is responsible for sequence specific DNA binding, whereas the less conserved leucine zipper region confers dimerization specificity. In plants, bZIP transcription factors regulate different cellular processes including seed maturation, flower development, pathogen defense, light and stress signaling (Jakoby et al. 2002, Nijhawan et al. 2008). bZIPs preferentially bind to DNA 14 sequences with an ACGT core. Binding specificity is also regulated by flanking nucleotides. Plant bZIPs preferentially bind to the A-box (TACGTA), C-box (GACGTC) and G-box (CACGTG) although there are also examples of nonpalindromic binding sites (Jakoby et al. 2002). By analyzing the promoters of ABA-inducible genes, it was found that ABA-responsive gene expression requires multiple cis-elements, designated as ABA-responsive elements (ABREs) or the combination of an ABRE with a coupling element (Jakoby et al. 2002). 1.7.2 bZIPs in rice There are 89 bZIP transcription factor-encoding genes in rice genome (Nijhawan et al. 2008). However, Plant Transcription Factor Database version 2 (Zhang et al. 2011) indicates 99 in indica spp. and 140 in japonica whereas Lu et al. (2009) reports 84. To date, several bZIP transcription factors have been identified or functionally characterized from rice. Among these, TRAB1 (bZIP66) (Hobo et al. 1999), OsABF2 (bZIP46) (Hossain et al. 2010a), OsbZIP23 (Xiang et al. 2008), OsABF1 (bZIP12) (Hossain et al. 2010b), OSBZ8 (bZIP05) (Nakagawa et al. 1996; Lee et al. 2003), OsbZIP72 (Lu et al. 2009), OsbZIP16 (Chen et al. 2012), OsbZIP71 (Liu et al. 2013) are positive regulators of ABA signaling, enhances abiotic stress tolerance. OsbZIP50 (named OsbZIP74) is associated with heat stress and salicylic acid mediated systemic acquired resistance (Lu et al. 2012). Moreover, a bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance (Zou et al. 2008). Two bZIPs, LIP19 (bZIP38) (Shimizu et al. 2005) and OsOBF1 (bZIP87) (Aguan et al. 1993) are involved in cold signaling. OsZIP-1a has a putative role in induction of the Em gene promoter by ABA (Nantel and Quatrano, 1996). OsbZIP58 regulates starch biosynthesis (Wang et al. 2013). RISBZ1 (Onodera et al. 2001) and RITA-1 (Izawa et al. 1994), involves in the regulation of rice genes expressed during seed development (Yamamoto et al. 2006) and REB is a transcriptional activator in rice grains (Yang et al. 2001). RF2a and RF2b function in vascular development and also play a role in symptom development of rice tungro disease. OsbZIP52/RISBZ5 could function as a negative regulator in cold and drought stress environments (Liu et al. 2012). OsbZIP39 is involved in the regulation of many ER stress-responsive genes 15 (Takahashi et al. 2012). Other rice bZIP proteins, RISBZ4, RISBZ5, OsZIP-2a and OsZIP-2b functions are not clear (Nantel and Quatrano 1996; Dai et al. 2003, 2004; Nijhawan et al. 2008). Rest others still to functionally characterize. 1.8 Crosstalk between ROS and ABA signaling The atomic structure and spin restriction of impaired electrons of O2 molecules give rise to reactive oxygen species if partially reduced. Reactive oxygen species (ROS), including superoxide (.O2-), hydrogen peroxide (H2O2) and hydroxyl radicals (•OH) are reactive molecules of oxygen resulting from the incomplete reduction of molecular oxygen. Being potential damaging capacity, high ROS leads the ‘oxidative stress’ concept, however, recent studies clearly identified ROS as more ‘oxidative signaling’ components, rather ‘oxidative stress’ molecules (Foyer and Noctor 2005). Now it is widely accepted that ROS are key regulators of plant metabolism, growth, development and certainly pathogen defense (Rio and Puppo 2009). Chloroplasts, mitochondria and peroxisomes are the major source of ROS production in plant cells. Among different ROS, being the most stable and ability to cross plant membranes and hydrogen peroxide is the well-studied molecule for cell to-cell signaling. ROS accumulation and seed germination appear to be linked together (Fig. 1.5). Progressive ROS accumulation after imbibition of after ripened seed has already been evident (Leymarie et al. 2012). Incubation of dormant sunflower seeds in the presence of ROS inducing compounds such as hydrogen cyanide or methyl viologen also alleviate seed dormancy (Oracz et al. 2007). Moreover more ROS accumulation is also evident in arabidopsis non-dormant seeds than that of dormant seeds whereas NADPH oxidase mutant (rbohD) shows deep dormancy (Leymarie et al. 2012). After ABA treatment, suppression of hydrogen peroxide (H2O2) occurs in barley aleurone cells whereas induced by GA (Fig. 1.5). Exogenous (H2O2) both enhances seed germination as well as alleviates seed dormancy in many plants along with enhanced seedling establishment. Oppositely, exogenous ABA application restricts ROS accumulation (Ye et al. 2012a) 16 Reactive oxygen species are also essential for leaf extension in the elongation zone of maize leaves (Rodriguez et al. 2002), root hairs and pollen tube growth (Foreman et al. 2003, Potocky et al. 2007), auxin signaling and gravitropism (Joo et al. 2001). Root hair defective 2 (rhd2) mutant of arabidopsis cannot accumulate ROS at the hair tip and fail to develop afterwards the initiation stage. The role of ROS in programmed cell death (PCD) has become an important topic in recent years (Breusegum et al. 2008). In deepwater rice, both aerenchyma formation and adventitious root initiation require PCD mediated by H2O2 (Steffens and Sauter 2009; Steffens et al. 2011). Moreover, in several systems, it has been shown that ROS can modulate the activity of specific transcription factors, thereby directly affecting gene expression. Moreover, DELLA proteins can regulate plant growth and defense processes by modulating the levels of ROS (Achard et al. 2008). ABA can interact with both the production and signaling functions of ROS. ABA induced H2O2 production is involved in stomatal closure in in V. faba guard cells (Zhang et al. 2001). ABA is the key inducer of H2O2 production in leaves of maize plants under water stress (Hu et al. 2006). 1.9 Seed germination under submergence Seed germination under submergence is much more complex than that of aerobic condition as complete submergence always leads to oxygen deficiency where oxygen is one of the prerequisites of seed germination. Most of the cereals are highly susceptible to submerged seed germination. However, among them, only rice is exceptional, possesses efficient capacity to germinate under submergence or complete anoxic condition (Ismail et al. 2012). However, the germination pattern is typically different than that of aerobic condition as completely lack of development of root under submergence, only the coleoptile develops. However, inability of subsequent shoot development results failure of seedling establishment thus causes significant loss of rice in direct seeded fields if sallow flooding or waterlogging occurs after direct seeding. However, direct seeding is popular method in irrigated and rainfed along with flood-prone ecosystems (Ismail et al. 2012). 17 Submergence tolerance during seed germination is a rare phenomenon in cereals, even among angiosperms. Only a few species can successfully germinate under water. It includes Trapa natans, Erithrina caffra, Nuphar luteum, Scripus mucronatus, Echinochloa spp. Except Trapa natans, all others germinate only with shoot, and no root develops. But, germination with radicle emergence happens only to Trapa natans (Jackson et al. 1996). In rice, after large scale screening of over 8000 germplasms, only a few varieties (19) are found capable of efficient seedling establishment after complete submergence (Angaji et al. 2010; Ismail et al. 2012). Subsequent studies identify only a few (5 to 6) tolerant genotypes, (Khao Hlan On, Khaiyan, Kalonchi, Nanhi, Ma-Zhan Red and Cody (Fig.1.8a) (Angaji et al. 2010; Ismail et al. 2009; Miro and Ismail 2013). Phenotypic comparison of tolerant and intolerant genotypes is shown in Figure 1.8b. Interestingly, initial screening also identified a Bangladeshi deepwater rice variety, Dholamon 64-3 as a tolerant variety with higher survival percentage (80%) (Fig. 1.8a) (Angaji et al. 2010). Moreover, Yamauchi et al. (1993) also identified germplasms of Bangladesh that are adapted to deepwater condition showed superior performance among tested genotypes. Since Rayadas are more primitive than typical deepwater rice, completely endemic to Bangladesh, well adapted to flooding condition along with practice of direct seeding, so the capacity of submergence tolerance during seed germination in Rayada varieties demands exploration. 18 Fig.1.8 Submergence tolerant cultivars during seed germination (a) and phenotype of comparative tolerance (b). list of submergence tolerance cultivars was adopted from Angaji et al. (2010). Image of phenotypic tolerance was downloaded from IRRI Flickr account: IRRI images’ photo stream (publicly available at http://www.flickr.com/photos/ricephotos/2199544108/in/photostream/, and labeled according). IR64 and IR42 are submergence sensitive varieties; oppositely Khao Hlan On and Khaiyan are tolerant genotypes. Only few physiological differences have been identified comparing tolerant genotypes with intolerant one. It includes keeping high soluble sugar, as well as high expression of Ramy3D whereas the progressive decline of starch content intolerant genotypes (Ismail et al. 2009; Ismail et al. 2012; Miro and Ismail 2013). Two QTL studies using crossing tolerant genotypes (Khao Hlan On, Ma-Zhan Red) with intolerant (IR64, IR42), respectively , identified several QTLs but they only share limited regions (Angaji et al. 2010, Septiningsih et al. 2013) . The specific genetic basis of submergence tolerance is still in an enigma. 1.10 Anthocyanin biosynthesis and trichome development: linked processes One of the most intensively studied metabolic systems in plants is flavonoid biosynthesis pathways. One of the specific branches of flavonoid biosynthesis pathways develops anthocyanin. Anthocyanin is one of the major pigments of plants defining red to blue colors in vegetative, reproductive as well as storage organs. The exact role of anthocyanins in plant growth and development is a matter of debate although classically explains the functions with photoprotection of chloroplasts; attenuation of UV-B radiation and free radical scavenging or antioxidant activity. However, recent studies suggest modulating role of anthocyanins in reactive oxygen signaling cascades involved in plant growth and development, as well as responses to stress (Hatier et al. 2009). 19 Similarly, leaf trichomes also play important role in biotic and even abiotic stress tolerances. Moreover, trichome development is an excellent model system for studying different aspect of cell differentiation apart biological significances (An et al. 2011). Recent studies confirm the intimately connected process involved in both anthocyanin biosynthesis and trichome development. A ternary activator complex (MYB-bHLH-WDR) comprising of R2R3 family MYB transcription factor (TF), a basic helix-loop-helix (bHLH) TF and a WD40 protein (Fig. 1.9a), regulate the spatiotemporal expression of structural genes involved in anthocyanin biosynthesis and trichome development (Patra et al. 2013). In Arabidopsis, GLABROUS 1(GL1- an R2R3-MYB TF), GLABROUS 3 (GL3) and ENHANCER OF GLABROUS 3 (EGL3) (both are bHLH transcription factors) together with TRANSPARENT TESTA GLABRA 1 (TTG1, a WD40 protein) regulate the structural genes of anthocyanin biosynthesis and trichome development (Fig. 1.9a) (Patra et al. 2013). Based on bHLHs structural features, it seems bHLHs act as docking protein between MYB and WD 40 proteins (Fig. 1.9b). In Arabidopsis, anthocyanin and trichome regulating bHLHs belonged to subgroup IIIF (Heim et al. 2003). The basic region of the bHLH domain consisting of 15-17 amino acids interacts with “CANNTG” consensus sequences of DNA. N-terminal acidic regions (MIR-MYB interacting region, 200 amino acids) interact with MYB transcription factors (Fig. 1.9b). The next negatively charged region (about 200 amino acids) might be necessary for interaction with WD40 proteins and/or the RNA polymerase III complexes (Fig. 1.9b) (Hichri et al. 2011). Finally, the C-terminal HLH region is responsible for either homo (such as GL3) or heterodimerization (such as GL3/EGL3) (Hichri et al. 2011) (Fig. 1.9b). A recent study on maize Lc (bHLH) by Tominaga-Wada et al. (2012) identifies regulation trichome development by N-terminal region whereas full length is necessary for anthocyanin synthesis (Fig. 1.9c). Ectopic expression of Lc in different plants like petunia (Fig. 1.9d) (Bradley et al. 1998), alfalfa (Ray et al.2003), tomato (Bovy et al.2002), and Arabidopsis (Lloyd et al. 1992) develops pigmented organs. 20 Fig. 1.9 Simplified anthocyanin biosynthesis and trichome development regulation. a) ternary activator complex comprising R2R3 family MYB transcription factor (TF), a basic helix-loop-helix (bHLH) TF and a WD40 protein, b) general structure of the bHLH transcription factors regulating flavonoid biosynthesis and trichome development, c) diagrammatic representation of R type bHLH parts required for both trichome and anthocyanin synthesis, d) ectopic expression of maize Lc (bHLH) in petunia under high-light conditions. Diagrams and figures adopted from (a) Koes et al. 2005, (b) Hichri et al. 2011, (c) Tominaga-Wada et al. 2012, and (d) Albert et al. 2009). 21 1.11 Objectives of this study The generalized aim of the study was to reveal the special traits of Rayada rice. Hence for the first time, we comprehensively reviewed and reexamined the special features of Rayadas (described in chapter 2). In addition, we studied in details, one of the notable physiological traits of Rayada rice, the lack of dormancy that is distinctly different from typical deepwater rice. We identified both physiological and molecular bases of the lack of seed dormancy (described in chapter 3). Furthermore, we characterized submergence tolerance capacity during seed germination of Rayada rice varieties that are rare in rice, even among cereals (described in chapter 4). Finally, we identified a novel mutant in Rayada rice background having multiple interesting phenotypes with practical implications, especially in hybrid rice technology (described in chapter 5). So specifically our studies will answer several key questions of Rayada rice and these are: 1. What are the special features of Rayada rice? And how they are different from typical deepwater rice? 2. What is the molecular and physiological basis of lack of dormancy in Rayada rice? 3. Are H2O2 and riboflavin the key signaling molecules of shoot development under submergence? 4. What are the factors that negatively regulate submergence tolerance? 5. How red or purple pigmentation of stigma can lead to sterility? 22 Chapter 2 Rayada specialty: the forgotten resource of elite features of rice 2.1 Introduction Flood and rice are two indispensable features of Bangladesh landscape, one of the most flood-prone countries in the world. Most areas of this country are less than 5 meters above sea level and hence almost every year, more than 20% areas become inundated with an additional 42 % with risk of inundation of varied intensity (Fig. 2.1) (Ahmed and Mirza 2000; Dasgupta et al. 2011). Rice is the lifeline of Bangladesh as it accounts for 76% of the daily calorie intake and 66% of protein needs for more than 94% of the population. Moreover, rice production contributes to 18% of national GDP and 55% of labor employment (Nasiruddin and Hassan 2009). Rice grows all the seasons round the year in Bangladesh. Beyond as staple food, rice is the culture and heritage of Bangladesh. Huge genetic diversity of rice exists in Bangladesh; among over 113,000 IRRI genebank accessions, 14,931 accessions are from Bangladesh (Wang et al. 2013). Still now more than 1,000 local varieties/landraces are cultivated in Bangladesh (Hossain and Jaim 2012). Fig. 2.1 Flood-affected areas of Bangladesh. 23 2.2 Deepwater rice in Bangladesh Deepwater rice is one of the special rices cultivated in Bangladesh since time immemorial. Deepwater rice, locally regarded as Broadcast aman, is defined as rice cultivated in flood plain deeper than 50 cm for one month or longer during the growing season (Catling et al. 1988; Catling 1992). In 2005-06, deepwater rice (Broadcast aman) was cultivated in 505.46 thousand hectare area, i.e. 4.08% of total rice cultivation area (Fig. 2.2a). In comparison, deepwater rice cultivation area was 18.14 % in 1971-72 (Fig. 2.2a). The gradual decreasing trends of deepwater rice cultivation over the past 34 years was due to the increasing cultivation of high yielding modern varieties with irrigation facilities in boro season since the yield of deepwater rice is generally low. The yield of deepwater rice over this period has always been around 1 ton/hectare (Fig. 2.2b), implying the absence of varietal development, whereas yields of other rice types have increased gradually due to both improved modern varieties and agricultural practices. Fig. 2.2 Status of deepwater rice cultivation in Bangladesh. Comparative scenario of cultivation area (a) and average yield (b) of deepwater (DWR) and total rice cultivation in Bangladesh from 1971-72 to 2005-06. Data were collected from Bangladesh Agricultural Research http://www.barc.gov.bd) 24 Council website, available at 2.3 Types of deepwater rice in Bangladesh There are three major types of deepwater rice cultivated in Bangladesh. They are: 1) Typical deepwater, 2) Bhadoia/Ashwina and 3) Rayada (Table 1) (Catling 1992; Bashar et al. 2004). All of them are collectively considered as deepwater rice having similarity in elongation capacity after rising flood water level, kernel color and yield, although there are some distinct differences among them, especially to Rayada rice. Ashwina/Bhadoia are almost similar to typical deepwater rice except less or no sensitivity to photoperiods (Bashar et al. 2004) and they usually flower in Bengali calendar months of Bhadra/Ashwin (August/September), hence coined the term Bhadoia / Ashwina (Haque 1974). Among these three types, Rayadas are most primitive, still sharing some features of wild rices (Khush 1997). 2.3.1 Rayada rice Rayadas are unique rice ecotypes, distinctive group of deepwater rice, totally endemic to Bangladesh, grown only in certain areas of Madhumati river tracts and have multiple physiological features different from typical deepwater rice (Table 1) (Perez and Nasiruddin 1974; Catling 1992; Bashar et al. 2004). They are photoperiod insensitive in early vegetative stage but strongly sensitive for flowering in late vegetative stage (Vergara and Chang 1985). Moreover, they completely lack seed dormancy, typically different to other deepwater rices along with longer life cycle. However, there are two types of Rayada rice based on duration of their life cycle i.e. (1) 12 months Rayada and (2) 6-8 months Rayada (Table 1) (Perez and Nasiruddin 1974; Bashar et al. 2004). Rayada rice was described as distinct group in very early literature (Watt 1891). Some biochemical studies also identified and supported its separate status to deepwater rice (Glaszmann 1987). Rayada rice was ranked separate group (Group-IV) from typical deepwater rice varieties (Group-III) based on fifteen polymorphic loci coding for 8 enzymes on 1688 traditional Asian varieties. In our study, we focuses on specialty of Rayada rice; elaborated their physiological and other features that are distinctly different from typical deepwater rice. 25 Table 1 Comparative features of different types of deepwater and wild rice (O. rufipogon) of Bangladesh. Wild rice Features Typical deepwater rice Ashwina / Bhadoia Rayada rice (O. rufipogon) Elongation ability after flooding yes yes yes (faster) yes (faster) Life cycle (months) 6-8 6-8 12 and 6-8 perennial Cold tolerance no no yes varying degree of tolerance varying degree of sensitivity less or no strongly sensitive sensitive Seed dormancy high high no high Responsiveness of short day in early vegetative stage yes yes no (not known) South and Southeast Asia Bangladesh and India Bangladesh South and Southeast Asia, Latin America, Australia, West- Africa Habigonj Aman I -VIII Ashwina Kaladigha, Beto Jhoradhan Photoperiod sensitivity Distribution Examples 26 2.4 Distribution of Rayada The Rayada rice is only cultivated in certain areas of natural lowland (locally called beel) beside the Madhumati river. The Rayada growing area is estimated at 5,180 hectares bordered by Sreepur, Magura (North) to Mollahat , Bagerhat (South), Magura and Narial (West) to Gopalgonj (East) (Fig. 2.3, R marked area) (Perez and Nasiruddin 1974). However, the 12-month Rayada is only cultivated at Kalia in Narail, Mollahat in Bagerhat, and Gopalganj (Perez and Nasiruddin 1974; Bashar et al. 2004) whereas, typical deepwater rice is cultivated throughout the country except three hill tract districts. Deepwater rice cultivation areas can be divided into mainly two zones. Zone-1 includes greater Faridpur, Jessore, Kushtia, Rajshahi and Pabna districts whereas zone-2 encompasses greater Sylhet, Comilla, Tangail, Dhaka, Noakhali and Mymensingh districts (Ahmed 1974). Soil properties of these two zones are generally different i.e. zone-1 characteristically natural to alkaline, rich in calcium and phosphorus but deficient of nitrogen and potassium. Oppositely, soil of another group is distinctly acidic, poor in calcium and phosphorus and comparatively higher in nitrogen and potassium (Ahmed 1974). Interestingly, Rayadas are only available in zone-1, shown in Fig. 2.3. Long term (1948-2008) climatic features (month-wise average rainfall, humidity, temperature and bright sunshine) of Rayada growing area are illustrated in Fig. 2.6. 27 Fig. 2.3 Distribution of Rayada rice in Bangladesh. Only the R (Rayada) marked areas (approximate) have Rayada rice cultivation. 2.5 Yearlong life cycle Numerous morphological and physiological differences exist among rice varieties (Chang and Bardenas 1965). For instance, some varieties mature within 80 days whereas Rayada needs a year to complete its life cycle (Khush 1997). Rayadas are cultivated rice of longest life cycle in Bangladesh. It is traditionally cultivated as mixed crop with boro in November/December. Boro is harvested in April/May before onset of flood while Rayada keeps growing and is harvested in the next 28 November/December (Perez and Nasiruddin 1974; Bashar et al. 2004). So from seedling to harvest is around 12 months or a year. Water level increases in around June, peaks in early August and recedes by the end of September or early October. Rayada usually flowers after flood water recedes in late September (Hasanuzzaman 1974), at that time day length already becomes shortened to induce flowering. Rayada type varieties might be eliminated because of very long life cycle (Vergara and Chang 1985). 2.6 Elongation ability of Rayada It is evident that deepwater rice elongates its internode after flooding as a survival strategy (Keith et al. 1986). However, the pattern of Internode elongation of Rayada is not clearly evident. In our experiments, interestingly, we observed all internodes under water were significantly elongated after prolonged inundation (Fig. 2.4a) and plants became more than 3 meters tall (Fig. 2.4d). Artificial inundation induced internode elongation of 2-4 times longer than control (Fig. 2.4a, c). Notably, basal internode above soil was found equally elongated to middle internodes and significantly different from control plants (Fig. 2.4a, c). We also compared internode elongation pattern of Rayada variety, Kaladigha with typical deepwater rices (Habigonj aman I-VIII) reported earlier (Hasanuzzaman 1974). Both air grown plants showed an almost similar pattern except much elongated distal nodes in typical deepwater rice (Fig. 2.4a). However, the basal Internode elongation pattern in Rayada was significantly different from that of deepwater rice. Some middle internodes of typical deepwater rice became similar to air grown plants (Fig. 2.4a). In addition, numbers of nodes were found significantly increased after inundation in both types, deepwater varieties showed more internodes than Rayada because of natural flood grown condition whereas Rayada variety was grown in greenhouse in 2.5 meter water tube (Fig. 2.4a,b). Internodes above the water level became same as control but developed both primary and secondary nodal tillers. However, highest elongated internode was the elongated uppermost internode (EUI) both in control and inundated plants (Fig. 2.4a). The eui mutant was reported to accumulate exceptionally large 29 amounts of biologically active gibberellins (GAs) in the uppermost internode (Zhu et al. 2006). Fig. 2.4 Internode elongation pattern of Rayada after inundation. (a) internode elongation pattern of Rayada and typical deepwater rice (Habigonj amam I-VIII after Hasanuzzaman (1974), (b) number of nodes per plants, (c) phenotypic comparison of basal internodes (above soil) of Rayada rice between control and inundated condition (bar = 5cm), (d) comparative phenotypes BRRI52, Kaladigha, and inundated Kaladigha (left to right, respectively) at plants maturity. Rayada variety, Kaladigha plants (4.5 months old) were grown in water tube with increasing water level keeping top leaves above water for 45 days reaching 2.5 meter. Then the plants were kept under around 2.5 meter inundation (water renewal fortnightly) for rest of months (around 7 months) till seed mature. Basal 30 internodes/nodes (3-4) beneath the soil were excluded in estimation and comparison. 2.7 Submergence tolerance Tolerance or resistance to complete submergence and partial submergence are regulated by similar type ERF (ethylene response factor) transcription factors by opposite mechanism. Under complete submergence, SUB1 restricts GA responsiveness by enhanced SLR1 expression (Xu et al. 2006; Bailey-Serres et al. 2010) whereas in partial submergence, SK genes enhance GA responsiveness by some unknown mechanisms (Hattori et al. 2009). Some aus ecotypes such as FR13A has SUB1A gene whereas deepwater even wild rice species possess SK genes for internode elongation (Hattori et al. 2009). Rayada rice is efficiently able to elongate its internodes after partial submergence. Similarly, in our experiments we imposed 2.5 meter submergence of equal sized plants and found equal capacity to withstand complete submergence of 2 weeks with similar performance to Sub1A introgressed rice (BRRI 52) (Fig. 2.5abc). But chlorosis or senescence of older leaves was more prominent in Rayada rice (Fig. 2.5 b, e). However, significant reduced senescence was observed in Sub1A introgressed variety, almost one third to Rayada variety (Fig. 2.5c,e). However, senescence of older leaves is common in deepwater rice varieties even under partial submergence. Despite older leaves senescence, survival performance showed no significant difference (Fig. 2.5 d). Similar observation of complete submergence tolerance was also previously reported in Rayada rice (Perez and Nasiruddin 1974). Complete submergence tolerance of Rayada rice might be regulated by SUB1A or some other means. However, two Sub1A like alleles (OrSub1A-1, OrSub1A-2) have already been reported in wild rice O. rufipogon (Li et al. 2011). So it is more likely that Sub1A type submergence tolerance regulates complete submergence in Rayada rice. 31 Fig. 2.5 Comparative submergence tolerance of Sub1A introgressed variety, BRRI 52 with Rayada variety, Kaladigha. a) before submergence, b) 14 days after submergence, c) 2 days after de-submergence, d) survival performance , e) leaf senescence . Plants of equal height (Kaladigha and BRRI 52 of 3 , 4.5 months , respectively) were completely submerged under 2.5 meter water for 14 days in water tube in greenhouse condition of 14/10 hrs. day-night cycle. Submergence tolerance performance was calculated 2 days after de-submergence. Representative image and figure were from two independent biological replicates. 2.8 Seed dormancy Prolonged seed dormancy is one of the distinctive features of the deepwater rices (Beachell 1975). Deepwater rice cultivars have a dormancy of 25-56 days (Catling 1992). Moreover, most wild rices also have strong seed dormancy (Cai and Morishima 2000). Dormancy is essential for deepwater rice to prevent inadvertent germination prior to harvesting of mature seeds while still on plant under high 32 humid or wet even inundated conditions. Among deepwater rices, Rayada is the only exception, having no seed dormancy. Since yearlong life cycle, Rayada seeds are sown within weeks or even days after harvest (Catling 1992). We also reexamined this feature and found efficient seed germination after freshly harvested (data shown in chapter 3, Fig 3.1). Lack of seed dormancy seems an adaptive feature of Rayada rice because of year long life cycle, highly photosensitive nature and flowering after flood water recedes. Moreover, during seed maturation period (late October-November), the rainfall, humidity and temperature, drop to lower extent to avoid unexpected germination (Fig. 2.6abc). Fig. 2.6 Climatic features of Rayada rice growing areas. a) average rainfall, b) humidity and c) lowest and highest temperature and d) bright sunshine hour. Long term (1948-2008) (a-c) average climatic status of Rayada rice growing areas were calculated based on data of two nearby weather stations (Faridpur and Jessore). Database of bright sunshine hours is only limited to 1967-2008 (Jessore) and 1985-2008 (Faridpur). Metrological data were collected from Bangladesh Agricultural Research Council website (http://www.barc.gov.bd/data_stat.php). 33 2.9 Cold tolerance Generally japonica rice is considered more cold tolerant than the indica (Andaya and Mackill 2003). Among indicas, cold tolerance is another important feature of Rayada rice and distinctive feature from typical deepwater rice (Catling 1992). Rayadas are tolerant to chilling temperature of 10°C. In the winter months the lowest temperature drops to around 10°C in the Rayada growing areas (Fig. 2.6c) and the plants across whole winter months during its initial vegetative growth stages. Typical deepwater rice is usually planted in April-May after the temperature rises (Fig. 2.6c) whereas Rayadas in November-December (Perez and Nasiruddin 1974; Bashar et al. 2004). We also re-examined the cold tolerance of Rayada by incubating plants in chilling temperature of 10°C for 4 days which resulted aus cultivars to cold injury whereas Rayada varieties showed no effect at all (Fig. 2.7). Fig. 2.7 Comparative phenotype of cold tolerance of (a) Rayada variety, Kaladigha with (b) upland rice, Manikmodhu. Three months old plants were incubated chilling temperature of 10°C for 4 days. Similar phenotypes were observed in 2 biological replicates. 34 2.10 Longer root system of Rayada Longer root system is unanimously considered as the feature of drought adaptation or tolerance (Blum 2011). Upland rice usually has longer root system and is hence adapted to drought prone area. Single nucleotide polymorphisms (SNPs) among 20 diverse rice varieties and landraces (OryzaSNP panel) including Rayada have been identified for deep exploration of rice diversity and gene-trait relationships and eventually for future rice improvement (McNally et al. 2009). Henry et al. (2011) evaluated variation in root system architecture and drought response among these 20 diverse rice genotypes of OryzaSNP panel and identified the aus isozyme group, particularly Dular genotype having highest drought resistance by deeper root growth with highest drought response index. Similarly, Gowda et al. (2012) examined root growth, water uptake and shoot growth among these 20 OryzaSNP panel under both well-watered and water-stressed conditions. We compared root growth pattern of Rayada to Dular genotype from their (Gowda et al. 2012) results. After analyzing their data, we identified longer root system of Rayada, even better than Dular both in well-watered and water-stressed condition (Fig. 2.8a). In well-watered condition, Rayada showed root length density (RLD) of 12.87 cmcm-3 in surface to 30 cm soil profile, more than 5 times higher than Dular (2.50 cmcm-3) (Fig. 2.8a). Interestingly , in water-stressed condition from surface to 100 cm depth , RLD of Rayada was always higher than Dular, especially at 30-45 cm soil profile, more than doubled in Rayada (RLD 3.77 cmcm-3) than Dular (1.60 cmcm-3) (Fig. 2.8a). However, inappropriately, Gowda et al. (2012) generalized Rayada as aus group although they are distinctly different in all aspects of growth and development. We have also re-examined root growth capacity at seedling establishment stage among selected Rayada, typical deepwater rice, flood tolerant and aus varieties. Although variation existed among ecotypes/ varieties but we observed that Rayadas always displayed longer root systems (Fig. 2.8b,c). Some upland varieties (eg. Dudsor and Boupagol) also showed elevated root growth initially but Rayada varieties superseded that due to higher growth rate (data not shown). Flash flood tolerant Sub1A varieties (BRRI51and 52) showed least growth for 35 root and shoot (Fig. 2.8bc). Month old seedlings showed distinct differences in phenotypes, especially in root length in Rayada variety, Kaladigha (Fig. 2.8b). Fig. 2.8 Comparative root growth of Rayadas with other varieties in different conditions. a) comparative root length density between Rayada and Dular both well-watered and water-stressed conditions, figure generated from data of Gowda et al. (2012), b) phenotypes of months old seedlings of selected varieties of Rayada, flood tolerant and upland rice , c) comparative root length at seedling establishment stage. Seeds were germinated on wet filter paper at 28°C in dark. 2.11 Photoperiod sensitivity Life cycle of rice consists of three phases: 1) vegetative growth stage, from germination to panicle initiation , 2) reproductive stage, from panicle initiation to flowering (generally 35 days) , and 3) the ripening stage, from flowering to ripen grain (usually 30-35 days , could be even 2 months depend on some factors such as water , nutrient richness) (Vergara and Chang 1985) . So the duration of the life 36 cycle of rice varieties mainly depends on the vegetative growth phase. Based on photosensitivity, the vegetative growth phase can be again divided into two phases: a) basic vegetative phase (BVP) -juvenile growth stage, independent to photoperiod and b) photoperiod sensitive phase (PSP). Rice varieties are classified into four types based on photoperiodic sensitivity: 1) Type A- both BVP and PSP are short, such as most modern varieties, 2) Type B - long BVP but short PSP, such as Habigonj Boro 5, BVP and PSP are 57 and 5 days, respectively , 3) Type C- short BVP and long PSP, such as typical deepwater rice varieties and 4) Type D- both long BVP and PSP. Rayadas are the only group of rice categorized type D (Vergara and Chang 1985). Most of the previous literatures reported BVP range from 10 to 63 (Gomosta and Vergara 1983). In typical deepwater rice varieties, BVP range from 3 to 43 days whereas that of Rayadas is 70 to 74 days, longest among all rice varieties reported earlier (Vergara and Chang 1985). Longest BVP is the characteristic feature of Rayada making it non-responsive in early growing stage to short photoperiod in February and early March (Vergara and Chang 1985). Possibly it is because of low temperature in winter months (Nov to Jan) that extended BVP (Gomosta and Vergara 1983). The range of PSP in photoperiod insensitive varieties is 0 to 30 days whereas PSP of photoperiod sensitive cultivars are above 31 days to years (Vergara and Chang 1985). Continuous long photoperiods resulted some cultivars to keep in vegetative stage more than 12 years (Kondo et al. 1942). So experimental duration for PSP determination was usually concluded after 200 days. All of the Rayada varieties tested earlier showed the highest limit of PSP, same as typical deepwater rice varieties (Vergara and Chang 1985). Critical photoperiod of Rayadas is around 12 hrs. Moreover, photoinductive cycle is also prolonged in Rayadas, more than 2 weeks (Vergara and Chang 1985). Interestingly, most of the wild rices also showed strong photosensitivity (Oka and Chang 1960). 37 2.12 Quick recovery after water stress Water stress recovery is crucial for Rayadas as it may be totally lost or damaged if it fails to recover from drought stress before onset of flood. Previous screening of IRRI in the 1981 identified 4 Bangladeshi varieties (Sarsari, Bhabani, Hijaldigha and Tilbazal) as promising deepwater varieties for drought tolerance (Datta et al. 1982). Interestingly, Sarsari showed similar recovery score as that of two drought tolerant check, Salumpikit and IR442-2-58 (Datta et al. 1982). In our study, we used Rayada variety, Kaladigha with aus variety Dudsor. Kaladigha showed leaf rolling in just minutes after transferring high humidity (90%) growing plants to low humidity (60%) condition (data not shown). High humidity induces ABA 8'hydroxylase to regulate local and systemic ABA responses in Arabidopsis (Okamoto et al. 2009). So we compared how quickly Rayada and Dudsor respond to recovery after watering. Leaf rolling is a useful indicator of leaf water potential in rice (O'Toole and Cruz 1980). Hence we compared water stress recovery rate by measuring the leaf drilling rate after watering. After analyzing the time course photographs, it revealed that Rayada variety sensed watering in just minutes and 50% derolled leaves (mostly young leaf) in 4-5 minutes (Fig. 2.9) whereas Dudsor sensed and recovered significantly slower than Rayada variety. Excellent performance of recovery of Rayada rice here suggests as it could be used as promising materials for future studies of water stress recovery and stress tolerance. 38 Fig. 2.9 Comparative time course leaf derolling after rewatering of water stressed Rayada rice variety, Kaladigha and aus genotype, Dudsor. Three months old plants were water stressed by water withdrawal for 5 days then rewatered. Time course image was derived by Sony Vegas Pro software from HD video taken during rewatering and image analysis was carried out in Adobe Photoshop 7. From enlarged photographs, fastest five derolled leaves breadth were measured and averaged on three position of each derolled leaf. Data representation was based on two independent biological replicates. Bar represents the standard deviation. 2.13 Origin and evolution of Rayada The origin and evolution of rice is little bit of a puzzle with considerable debate mainly on single or multiple ancestries with domestication place (Londo et al. 2006; Molina et al. 2011; Huang et al. 2012; Sang and Ge 2013). Most of the scientists believe O. rufipogon as progenitor of deepwater rice because it has still deepwater types, adapted to deepwater habitat and having photoperiod sensitivity too (Glaszmann 1987; Catling 1992; Bashar et al. 2004). As elongation ability after flooding is the main feature of both Rayada and typical deepwater rice, hence evolutionary study of key genes responsible for internode elongation after flooding is also relevant to Rayada. Snorkel genes (SK1 and SK2) allow rice to adapt in deepwater condition by enhancing GA responsiveness (Hattori et al. 2009). Phylogenetic analysis of snorkel protein sequences among deepwater and wild rices were shown in Fig. 2.10. Both of the wild species (O. rufipogon and O. nivara) possess complete SK1, but SK2 of O. nivara is truncated due to insertion of transposon. However, O. glumaepatula, in another wild rice species, have SK2 and SK2-like genes, but deficient of SK1 (Fig. 2.10). Phenotypic effect of SK2 is more prominent and wild rice species (O rufipogon and O. glumaepatula) possess the SK2 genes, suggesting these genes may have been evolved before or during wild rice speciation (Hattori et al. 2009). 39 Fig. 2.10 Phylogenetic analysis of snorkel protein sequences among deepwater and wild rices. Phylogenetic tree was derived using the Neighbor-Joining method by MEGA5 (Tamura et al. 2011). DDBJ accession numbers of SNORKEL1 and SNORKEL2 are as follows (name, accession numbers): O. sativa DWR Bhadua, (AB510480 and AB510481), O. rufipogon (AB510482 and AB510483); and O. nivara (AB510484 and AB510485). AB510486 and AB510487 are SNORKEL2 and SNORKEL2-like genes in O. glumaepatula. Interestingly, Bangladeshi deepwater rice varieties (all three types) were found to possess special isozyme alleles, Est10-4 and Amp5-4 which are frequently found in Asian common wild rice O. rufipogon but rare in other cultivars (Cai and Morishima 2000). This finding suggests Bangladesh deepwater rice has inherited these alleles from its wild progenitor. Generally, it has been considered that deepwater rice varieties belong to indica spp. only , although a very few reports on isozyme analysis showed some of floating rice of Bangladesh including Rayada displayed japonica relevance (Cai and Morishima 2000, Wang et al. 2013). However, none of the 188 deepwater rice varieties tested from Thailand, Vietnam and Cambodia showed japonica pattern (Catling, 1992). Analyzing the polymorphism of nuclear, mitochondria and chloroplast DNA of 193 genotypes, Sun et al. (2002) identified japonica type nuclear and chloroplast DNA but indica 40 type mitochondria DNA in some Bangladeshi deepwater landraces including Rayadas. Since some of Bangladeshi deepwater rice varieties including Rayada showed japonica relevance with presence of Est10-4 allele and japonica type nuclear and chloroplast DNA, so considering other features of Rayadas altogether (such as yearlong life cycle, strong photosensitivity, multiple stress tolerances and elongation ability after flooding that are resemble to wild rice), collectively might deduce Rayadas as a possible missing link between indica-japonica differentiation. However, most of the large scale molecular (GWAS etc.) or evolutionary studies of rice omitted Bangladeshi unique varieties such as Rayadas as experiment material hence the enigma lingers. 2.14 Rayada: the forgotten resource of elite features of rice Being sessile organisms, plants have evolved with special capacity to sense, respond, and adapt to unfavorable environmental conditions. Water stresses, either excess (flooding) or shortage (drought) of water, significantly limit plant growth and development and obviously crop production. The natural capacity of these stress tolerance mainly depends on plants genome specialty and efficiency of activation of adaptive mechanisms (Chinnusamy et al. 2004). However, crop domestication and thereafter gradual selection or directional breeding have narrowed the genetic diversity of elite varieties, even promoted gathering of deleterious mutations in their stress response mechanisms (Tang et al. 2010), whereas local ecotypes, landraces and wild relatives growing on native environment and preferences still keep genetic diversities for features like stress tolerance, disease resistance etc. (Suslow et al. 2002) Rayadas are resources of desirable features like flood, cold and drought tolerance and still share some features of wild rice (Perez and Nasiruddin 1974; Glaszmann 1987; Khush 1997; Bashar et al. 2004). In addition, we have found features like longer root system (Fig. 2.8), quicker recovery (Fig. 2.9), and faster stomatal responses (data not shown) etc. in Rayada rice that are associated with drought tolerance. However, mainly because of two undesired features like yearlong life cycle and poor yield diminished the focus on it although development of 41 deepwater rice had initiated since 1917 (Zaman 1977). Despite the limitations, deepwater rice was considered as a model plant to study stem elongation and basic aspects of plant growth (Kende et al. 1998). Functional role of snorkel genes in deepwater rice has been already revealed although how SK genes regulate GA responsiveness is still unknown (Hattori et al. 2009). Moreover, several other minor QTLs (qTIL2 and qTIL4) identified in deepwater rice (Nagai et al. 2012) need to be characterized. Moreover, focused study on Rayada or deepwater rice would enrich our understanding on GA/ABA antagonism which is important to know how plants cope with changing environments. In addition, Rayada rice seems to be an excellent material for natural stress memory phenomenon study (Chinnusamy and Zhu 2009) as it always passes through multiple stresses (flooding, cold, sometimes drought) before completion of life cycle. In summary, Rayada is such a primitive deepwater rice ecotype that can offer many traits that are badly needed in rice breeding , especially for stress tolerance. We expect Rayada rice will become a new focus of rice germplasm research with the rapid advances in genome research weaponry. 42 Chapter 3 OsbZIP84 is involved in seed dormancy regulation of Rayada rice through ABA catabolism 3.1 Introduction Seed dormancy and germination are two of the complex interrelated processes of plant development. Myriad of regulators of both endogenous hormonal signaling and exogenous factors are involved in these processes. However, abscisic acid (ABA) is the key regulator of both of the processes (Nambara et al. 2010). Despite extensive research, still seed dormancy is being considered as a least understood phenomenon in seed biology (Finkelstein et al. 2008). Seed dormancy, a temporary quiescent state of seed, is one of the natural ways of preventing seed germination. Complete lack of dormancy can cause pre-harvest sprouting or vivipary in wet or high humid environment, thus might cause significant economic loss of crops due to reduction of yield as well as seed quality (Gubler et al. 2005). Moreover, it can also increase the risk of seedling mortality in unfavorable conditions. Oppositely, strong and prolong seed dormancy significantly hamper synchronous germination, hence not desired in agriculture. Therefore, well-balanced seed dormancy is necessary for crop plants (Graeber et al. 2012). Most of the wild rice accessions possess strong seed dormancy (Vaughan 1994; Veasey et al. 2004). Generally seed dormancy is considered as an adaptive trait of wild species. However, reduction or loss of seed dormancy is one of the traits of the domestication syndrome (McCouch et al. 2012). All deepwater rice varieties except Rayadas possess extended period of seed dormancy (Catling 1992). However, among deepwater rices, Rayadas is considered most primitive based on yearlong life cycle, multiple stress tolerance capacity, strong photosensitivity and endemic nature (Catling 1992). Moreover, it still shares some features of wild rices (Khush 1997). Interestingly, Rayada varieties completely lack seed dormancy; Rayada seeds are sown within weeks or even days after harvest (Catling 1992). Therefore, Rayadas are good example of natural variation of seed dormancy among deepwater rices. Physiological features along with endemic nature suggest lack of dormancy may be also an adaptive feature of Rayadas. The 43 genetic basis of such adaptive feature is the key question of both fundamental and evolutionary biology. But both physiological and molecular bases of this trait of Rayadas are completely unknown. In this study we, for the first time, examined Rayadas as a natural variant of deepwater rice to reveal both physiological and molecular bases of their lack of dormancy. Numerous studies with both ABA and GA biosynthesis and signaling mutants in different plants enriched our understanding of antagonistic role of the two phytohormones in seed dormancy and germination (Nambara et al. 2010; Graeber et al. 2012). ABA biosynthesis mutants show reduced dormancy in freshly harvested seeds whereas ABA catabolic mutants act oppositely, enhances seed dormancy. After imbibition of seeds, catabolic activities of ABA8’hydroxylases rapidly decrease ABA content in non-dormant seeds whereas remain high in dormant seeds (Graeber et al. 2012). Key ABA catabolic gene of barley (HvABA8’OH-1) is highly expressed in non-dormant seeds and surprisingly, localized onto coleorhiza (Millar et al. 2006). Apart these, some individual genes such as DOG1 (Bentsink et al. 2006), SDR4 (Sugimoto et al. 2010) also regulate seed dormancy by some unknown means. Moreover, some other signaling molecules such as NO, H2O2 also modulate the sensitivity of ABA and consequently regulate seed dormancy and germination. H2O2 interact with ABA signaling, up-regulate ABA catabolic genes hence decrease ABA content during seed germination (Liu et al 2010; Ye et al. 2012a). Among ROS, H2O2 received special attention due to more stability and long lived feature (half-life 1 ms). It is well evident that plants use ROS as signaling molecules or second messenger in different growth and development as well as biotic and abiotic stresses (Rodriguez et al. 2004; Zhang et al 2001) including dormancy alleviation, after-ripening and germination (Oracz et al. 2007; ElMaarouf-Bouteau and Bailly 2008; Ye et al. 2012a). Inhibition of ROS production by diphenylene iodonium (DPI) delays endosperm weakening and seed germination. Similarly, NADPH oxidase mutant (atrbohB) fails to after ripen due to reduced ROS accumulation (Műller et al. 2009). Therefore, we paid our special attention to observe ROS accumulation in freshly harvested seeds of Rayada whether they have any involvement in the feature of lack of dormancy. 44 During the course of domestication, transcription regulators are assumed to be the evolutionary switches or dominant class of genes that have changed (Doebley and Lukens 1998; Doebley et al. 2006). bZIPs are very ancient and evolutionally conserved family of eukaryotic transcription factors and some of their DNAbinding specificities evolved before the divergence of the metazoa and fungi (Amoutzias et al. 2007). In plants, some bZIP transcription factors are directly linked with ABA signaling pathways and regulate different cellular processes including seed maturation, flower development, pathogen defense, light and stress signaling (Jakoby et al. 2002, Nijhawan et al. 2008). A bZIP TF, ABI5 is one of the key transcription factors that confer ABA responsiveness (Nambara et al. 2010). Stored mRNA profiling in dry seed of Arabipsis revealed enrichment of genes with ABREs in promoter (Nakabayashi et al. 2005). Recently, Tsugama et al. (2012) claimed a bZIP transcription factor, VIP1 enhance the promoter activities of ABA 8'-hydroxylase thus regulate ABA catabolism , although VIP1 is classically explained as VirE2 interacting protein and required for nuclear import and Agrobacterium tumorigenicity (Tzfira et al. 2001). We first examined the physiological basis of lack of dormancy in Rayada rice by determining ABA content as well as ROS accumulation in freshly harvested seeds after imbibition. Then after analyzing stepwise gene expression of 32 bZIPs in three ecotypes, we selected OsbZIP84 as our gene of interest (GOI), possibly involved in ABA catabolism regulation. We hypothesized the role of OsbZIP84 in seed dormancy, after analyzing gene expression of GOI and ABA8oxs in 4 different growth and developmental situations in Rayada rice together was ABA content and sequence homology of reported genes. Finally we ascertain our hypothesis with phenotyping of osbzip84 in japonica background representing dormancy phenotype along with similar phenotypes of pharmacological studies of ABA catabolic inhibitor. 45 3.2 Materials and methods 3.2.1 Plant materials Eight different varieties of rice variety (O. sativa L.) were used in this study. It included Rayada (Kaladigha, Sarsari), typical deepwater rice (Habigonj aman VIII), flash flood tolerant (BRRI51, BRRI52), Aus (Dudsor, Boupagol) and Japonica type (Donjing) rice. Except Donjing, all other varietal seeds were collected from Bangladesh Rice Research Institute (BRRI), Bangladesh. Mutant of OsbZIP84 was collected from POSTECH, Korea (Jeon et al. 2000). The homozygous mutant was identified by PCR using T-DNA and gene specific primers listed in Table 2. 3.2.2 Growth conditions and stress treatments Intact seeds (with hull) were used for germination performance of freshly harvested seeds of Rayadas and deepwater rice. However, dehulled seeds were used in other experiments with downstream ABA or RNA extraction. Seeds were sterilized with diluted chlorex and germinated on sterile moist filter paper at 28°C in dark. Germinated seedlings were grown on Kimura B nutrient solution (renewal cycle 3 day) in greenhouse at 25-30°C with 14 /10h photoperiod shift. Different treatments were performed mentioned in respective figure legends. Samples of leaves, seeds, nodes, internodes under different treatments were collected in liquid nitrogen and stored at -80°C (if necessary) for ABA determination and RNA isolation. For each treatment, 10-30 seeds were used and triplicate experiment sets were used unless otherwise mentioned. 46 Table 2 List of primer sequences used in this study. SL no Name MSU ID RAP ID Forward primer (5’-3’) 1 bZIP04 LOC_Os01g36220 Os01g0542700 GGCGTTGTGTGTACTACTGT TAGTAGTAGTAGTTGCAGTGCA 2 bZIP05 LOC_Os01g46970 Os01g0658900 GGAAATTGATGGTGCGGCTATGT GTTGGCACTTCAGTTACTGGGG 3 bZIP13 LOC_Os02g03580 Os02g0128200 ATGGCAAAGCATCGTCTGGTTC GTGCCATTGGTGTCGTTCTGTT 4 bZIP14 LOC_Os02g03960 Os02g0132500 AAGAGGATGCTGTCGAACAGGGA GGGGAGGATCTGAAGAGGTCGTC 5 bZIP16 LOC_Os02g09830 Os02g0191600 AGCATCACCATGATTCCAGTGACT GGCTACTTCAGCATCTTCCATTTCC 6 bZIP18 LOC_Os02g10860 Os02g0203000 ACAGAATGAGAACCAGATGCTCAGA GTGCTGTTGGGCATCCTGTGG 7 bZIP24 LOC_Os02g58670 Os02g0833600 AGCTCCTTCCTTTCCATCTCCAG ATGAGCTTCTTCGGTGGGAACT 8 bZIP26 LOC_Os03g13614 Os03g0239400 CATCGGCGACACAGAATGAACA GTGATTGCCCGGGATAGGGATT 9 bZIP29 LOC_Os03g20650 Os03g0322700 ATGACGCTCGAGGACTTCCTGTC AGCTGATTCTTTGGCTCCTGTTG 10 bZIP34 LOC_Os03g59460 Os03g0809200 ATGCTGACGAGAAGGGTTTGT GTGCTTATTTAGCTGGCTACTGC 47 Reverse Primer (5’-3’) SL no Name MSU ID RAP ID Forward primer (5’-3’) 11 bZIP37 LOC_Os04g54474 Os04g0637000 TGTGGAGTCACGAGCAGTTCAA ATTTGCATACACAGCCTGCTGC 12 bZIP39 LOC_Os05g34050 Os05g0411300 TCGCGGACCGCTGTGGATT TGGGAAATGCATGCCGGGAAT 13 bZIP40 LOC_Os05g36160 Os05g0437700 CTAATGTAACGGAGGGTGTGGGT CCTTATGACAACTGCAACGGCTT 14 bZIP42 LOC_Os05g41070 Os05g0489700 CTGGTACGTCTGATTTGAACGCTG TTGACCTGGCAGCTGATTCCCT 15 bZIP44 LOC_Os05g41540 Os05g0495200 ACATCAAGGAGGACCTGACGAA GCCATTGAAGCACATACCTGGA 16 bZIP45 LOC_Os05g49420 Os05g0569300 TGGCAGGAAACGGAGATTGGAT CAGATCCTCTGTTTCAGCCTGC 17 bZIP48 LOC_Os06g39960 Os06g0601500 GATAGCGACGACGAGGAGATAGT TGTTCTTCAGCACCTGCCTAAGC 18 bZIP51 LOC_Os06g42690 Os06g0633400 ATGGCGTACCAATCGTTC TTAATCCATTGGCGCCA 19 bZIP53 LOC_Os06g50310 Os06g0716800 CTGTGCATGACCTCTCACACAC CACCACCAAGATAACTTCCCTGG 20 bZIP57 LOC_Os07g03220 Os07g0124300 CGGATACGATGGTGTTGCCTC TAGGAGGCGGGACAGCCCTA 21 bZIP59 LOC_Os07g10890 Os07g0209800 GACCACAAGGATTCGTCTGCAC ATCCAGGTGAGCAACACCATCA 48 Reverse Primer (5’-3’) SL no Name MSU ID RAP ID Forward primer (5’-3’) 22 bZIP63 LOC_Os07g48820 Os07g0687700 TGTTGATGGCATAATGGCGCAT CAGAAGACCCTGATGAAGCGAG 23 bZIP67 LOC_Os08g38020 Os08g0487100 AGCTGGAGCGCATCCTGGAAGAA AGCAGCGGTAGTCGTAGTGCTGT 24 bZIP68 LOC_Os08g43090 Os08g0543900 TGGGGAGTTCACTGAAGCTGAG TGAGGACTAATGCTGATGAGGGT 25 bZIP69 LOC_Os08g43600 Os08g0549600 GTCGTTTCAGGACGGGCTTCCGTC GCCTTCAGCTGGTGGCACTGG 26 bZIP70 LOC_Os09g10840 Os09g0280500 GCAACTACGACATGGCCGATTT CTTCCTTCCTTCTGCAAGCCAC 27 bZIP71 LOC_Os09g13570 Os09g0306400 TCCAGGCCCAGATGGAGAAGAA AGTGGTGGCTGAAATTGTAGGAGT 28 bZIP77 LOC_Os09g36910 Os09g0540800 GTGCTCTCCGCGCCATTCTGA GCACTGCCACAGCTAAGCTTCCT 29 bZIP78 LOC_Os10g38820 Os10g0531900 ACCCTCTCCGCTCAGCTGACTC CGTGCCATTCTGCTGCTCTGGCTG 30 bZIP81 LOC_Os11g06170 Os11g0160500 CTGCTCCAGGCGTACTGGGATG TGGCCGCTCATCGATTGGCCT 31 bZIP84 LOC_Os12g06520 Os12g0162500 AAGAGAATTCTGGCGAACAGGC GTGGCCGCCTAAATAAGGCA 32 bZIP85 LOC_Os12g09270 Os12g06334500 TGGGATTCGAGACTGATGCTCC CGAGACCTCTGAGCATACTGCA 49 Reverse Primer (5’-3’) SL no Name MSU ID RAP ID Forward primer (5’-3’) 33 bZIP89 LOC_Os12g43790 Os12g06334500 CTTCCTCGCCGACATTGGCTTC CTACGCGATCAACGAGGCGAG 34 LEA3 LOC_Os05g46480 Os05g0542500 TAGGATCAATGGCTTCCCACCAG GATGGCAGAGTCCTTGGTGTACT 35 SAPK8 LOC_Os03g55600 Os03g0764800 ACCGTGATTTGAAGTTGGAG CCGTATTTCTGGGATTGTTA 36 SAPK9 LOC_Os12g39630 Os12g0586100 GATTGCCGATGTTTGGTC TGTTGCTTCTGCCAGTATCT 37 SAPK10 LOC_Os03g41460 Os03g0610900 ATGCCGATAATGCACGACG TGGTTGCGAATGAAGAACAGAC 38 PP2C4 LOC_Os01g36080 Os01g0541900 GTCACCATTGCCACTGATCTAG TTGCTGCTGTAAGTTGTTGTTCG 39 PP2C9 LOC_Os01g62760 Os01g0846300 TCGCACTATCCGTTGACCACA GCCAACTTCGTCAGCAGCAT 40 qbZIP13 LOC_Os02g03580 Os02g0128200 GGCCTACTCGGCAATTCCACCG CGTAAGGTGGCACCATGGGCTG 41 qbZIP40 LOC_Os05g36160 Os05g0437700 CCTCGGGAGGGCTATCTGCTCC AGCCAGGAAATCCTCCAGCGTCA 42 qbZIP42 LOC_Os05g41070 Os05g0489700 CCCAGGTTAGGTCAGCCTCGCT ATCAGCCCCCGAGGATTCCACC 43 qbZIP44 LOC_Os05g41540 Os05g0495200 TTCTCCCATCCGGAGGTGCCTG GGATTGCAGGTGTGGGTGTGGG 50 Reverse Primer (5’-3’) SL no Name MSU ID RAP ID Forward primer (5’-3’) Reverse Primer (5’-3’) 44 qbZIP53 LOC_Os06g50310 Os06g0716800 CACCTGCGTCCATGTCCACACC GGCCGCTTCTTGGAGGCATTGT 45 qbZIP59 LOC_Os07g10890 Os07g0209800 GGGCGGCACTTCCTAATGCCAC ACCCAGGACTTGCTTCCCCCTG 46 qbZIP63 LOC_Os07g48820 Os07g0687700 GTGAGTCTGTGCCGTTTCCCCG GTGGCGGTTGGGGGAAATCTCG 47 qbZIP68 LOC_Os08g43090 Os08g0543900 CGAGTCACCGAAGCTGCCACTG GCACCAAACAGAGATGCAGCGC 48 qbZIP71 LOC_Os09g13570 Os09g0306400 CACCTCGCAGGTGAACCAGCTG TGTGCCTGCACTGCCACAAGG 49 qbZIP84 LOC_Os12g06520 Os12g0162500 GGTGGGCTACCGGACTACGC TCTCTTTGCGCGCTTGGGGTC 50 qOsABA8ox1 LOC_Os02g47470 Os02g0703600 CCTGCCCGGAAAAGTTCGACCC CGCACCGGGAAAGGAAAGGGAG 51 qOsABA8ox2 LOC_Os08g36860 Os08g0472800 ACAGCAGCAGGTGCTTGACACG CATTGCATCCACACACACGGCG 52 qOsABA8ox3 LOC_Os09g28390 Os09g0457100 GCTACGGCCATGAGAAAGCCGTC AGTAGAGCTGAAGGGTCTCGCCG 53 qGA20ox1 LOC_Os03g63970 Os03g0856700 TTCCTCTGCCCGGAGATGGACAC ACGTGAAGTCCGGGTACACCCTC 54 qNCED1 LOC_Os02g47510 Os02g0704000 ATCATGCACTCGGTGAACGCGTG 51 CGTGCTCGATGGAGAGGACGTTG SL no Name MSU ID RAP ID Forward primer (5’-3’) Reverse Primer (5’-3’) 55 qNCED2 LOC_Os12g24800 Os12g0435200 GTTGATGAGCTGCCCCCGACAAG GGTGCTGTGGGTTGGGACCATTG 56 qNCED3 LOC_Os03g44380 Os03g0645900 AGCTCACCACCCAGGCCTGATC CAGGCTCCCTCTGGTCACTTCC 57 qNCED4 LOC_Os07g05940 Os07g0154100 CCGTCACCGAGAACTATGCCGTC CGACGCCTTCTCCCTGTCGTATAC 58 qNCED5 LOC_Os12g42280 Os12g0617400 CGCCAACTCCAGCTCTCACCTC GGCATGGCTGTCCTACAAGAGC 59 qActin1 LOC_Os03g50890 Os03g0718100 AGAGGAGCTGTTATCGCCGTCCTC GCCGGTTGAAAACTTTGTCCACGC 60 Tubulin α LOC_Os03g51600 Os03g0726100 TGTGCATGATCTCCAACTCCAC AATCAGGCCTTGACACACAGAA LOC_Os12g06520 Os12g0162500 ACATTGTGAAACGGAGGGAG GAAGGTGTAATCTCCCCGTG PFG_ 2A61 50025.R (Mutant) 62 Middle primer 1 (T-DNA) CATCACTTCCTGATTATTGACC 63 Middle primer 2 (T-DNA) AACGCTGATCAATTCCACAG 52 3.2.3 Microarray data analysis To identify the OsbZIP family members, first all the OsbZIPs mentioned in Plant Transcription Factor Database (http://plntfdb.bio.uni-potsdam.de) was manually checked using BLAST. To avoided confusion, Nijhawan et al. (2008) nomenclature was followed. The gene expression was manually checked from RiceGE: Gene Expression Atlas and NCBI GEO Profiles. For RAP and MSU rice gene id conversion, RAP-DB ID converter was used (http://rapdb.dna.affrc.go.jp/tools/converter/run). 3.2.4 RNA extraction and quantitative real-time (qRT)-PCR RNA was extracted from respective samples using the Agilent Plant RNA isolation Mini Kit and then digested with RNase-free DNase I to eliminate genomic DNA contamination. First-strand cDNA was synthesized with oligo (dT) primers using a Promega cDNA kit accordingly. Transcript levels of each gene were measured by qRT-PCR using Mx3000p QPCR System with iQ SYBR Green Supermix. The data were normalized to the amplification of a rice α-tubulin or actin 1 gene. Primer sequences of genes used for RT and qRT-PCR expression analysis and mutant screening were listed Table 2. 3.2.5 Extraction and measurement of ABA Measurement of endogenous ABA content was carried out using the radioimmunoassay (RIA) method as described by Jia et al. (2012). Respective samples of 0.2 g (or 10 seeds) were ground in liquid nitrogen and homogenized in 1 ml of distilled water and shaken overnight at 4°C. The homogenates were centrifuged at 12000g for 20 min at 4°C and the supernatant were directly used for ABA assay. The 450 µl reaction mixture comprised of 200 µl of phosphate buffer (pH 6.0), 100 µl of diluted antibody (Mac 252) solution, 100 µl of [3H] ABA (@ 8000 cpm) solution, and 50 µl of crude extract. The mixture was then incubated for 45 min at 4°C and the bound radioactivity was measured in pellets precipitated with 50% saturated (NH4)2SO4 with a liquid scintillation counter. 53 3.2.6 ROS staining and confocal laser scanning microscopy Total ROS staining was dichlorodihydrofluorescein determined diacetate using (H2DCF-DA) fluorescent dye (Invitrogen). 2',7'Freshly harvested seeds were washed gently with Hanks’ balanced salt solution (HBSS) buffer and stained with H2DCF-DA at 37 °C in the dark for 30 min, then washed briefly with same buffer and observed under Olympus Fluoview FV1000 confocal laser scanning microscope with the following settings: excitation=488 nm, emission=535 nm, frame 1024×1024. To compare ROS staining between seeds of Rayadas and typical deepwater rice, identical exposure conditions were used for all samples. Representative images of 10 seeds were used for comparison. 3.2.7 Statistical analyses ANOVA and Duncan’s multiple range test were done using Excel Add-ins develop by Onofri 2007. The values are the means and SD of at least three biological replicates from for each experiment unless otherwise mentioned. The lowercase a, b or c etc. in figures represents ranking test and different letters indicate significant differences at the P<0.05 level. For other simple experiments, the student t-test was used and significance level was mentioned in respective figure. 54 3.3 Results 3.3.1 Lack of dormancy of freshly harvested Rayada seeds We examined the feature of lack of dormancy of freshly harvested seeds of Rayada variety, Kaladigha. Seeds of different time course after harvest (0, 7 and 180 days after harvest (DAH), were used to compare seed germination performance of freshly harvested Kaladigha seeds with that of typical deepwater rice variety, Habigonj Aman VIII seeds (Fig. 3.1 a). Germination performance of freshly harvested seeds of Rayada variety clearly demonstrated non-dormant nature (Fig. 3.1a,c). However, absolute freshly harvested seeds of Kaladigha, i.e. 0 DAH showed delayed and reduced percentage of germination whereas seeds of 7 DAH showed high germination percentage comparing that of typical deepwater rice (Fig. 3.1a). As ABA is the master regulator of seed dormancy, hence we compared the ABA content of seeds of freshly harvested Rayada variety, Kaladigha with that of typical deepwater rice variety, Habigonj Aman VIII. Remarkably, freshly harvested seeds of Habigonj Aman VIII possessed significantly higher ABA than that of Rayada variety (Fig. 3.1b). Although we examined ABA content of only one variety of each type, but we assume other varieties of Rayada might show similar trends. Phenotypes of seedlings of 7 days after imbibition of Kaladigha variety were shown in Fig. 3.1c. Except reduced root growth, seeds of 7 DAH showed virtually no difference with that of 180 DAH (Fig. 3.1c). Interestingly, moisture content was found significantly different between complete freshly harvested seeds (0 DAH) with that of 180 DAH. Freshly harvested seeds usually contained around 25% moisture whereas it was only 13.38% in dry seeds of 180 DAH. Seeds of 7 DAH showed intermediary moisture content (Fig. 3.1d). 55 Fig. 3.1 Lack of dormancy of freshly harvested seeds of Rayada. a) comparative germination performance of seeds of typical deepwater rice variety Habigonj Aman VIII (Hbj VIII) with Rayada variety, Kaladigha (KD) of 0, 7 and 180 days after harvest, b) ABA content of freshly harvested seeds (0 DAH) , c) seedlings of Rayada variety Kaladigha developed 7 days after imbibition of different DAHs. d) moisture content of seeds of different DAHs. 10 seeds of different DAH were used for triplicate experiments. ABA content was measured using the RIA method as described in materials and methods. Values are means with ±SD (n=3). The symbol (*) indicates the significance level at * P<0.05, ** P<0.01 3.3.2 ROS accumulation in freshly harvested Rayada seeds after imbibition Previous reports suggested involvement of ABA content on ROS accumulation in seed germination (Oracz et al. 2007; Ye et al. 2012a). Since Rayada showed reduced ABA content and we observed partial overriding of dormancy of freshly 56 harvested Habigonj Aman VIII after H2O2 treatment (data not shown), hence we attempted to compare ROS accumulation status of seeds after 24 hrs imbibition in freshly harvested (7 DAH) Rayada seeds (Kaladigha) with that of typical deepwater rice (Habigonj Aman VIII). After fluorescent staining with H2DCFDA, significant difference was observed between them. Huge ROS accumulation was observed in embryonic axis of Kaladigha seeds (Fig. 3.2, upper panel) whereas noteworthy reduction of ROS accumulation appeared in Habigonj Aman VIII (Fig. 3.2, lower panel). Moreover, both exogenous ABA and DPI (NADPH oxidase inhibitor) also reduced germination in freshly harvested Rayada seeds (data not shown). All these clearly represented the ROS accumulation after imbibition in freshly harvested Rayada seeds might be a decisive consequence of reduced ABA content thus promoted seed germination. Fluorescent Bright field Merged Fig. 3.2 Comparative ROS accumulation of freshly harvested seeds of Rayada variety, Kaladigha (upper panel) and typical deepwater rice variety, Habigonj aman VIII (lower panel). 24 hrs imbibed freshly harvested (7DAH) dehulled seeds were stained with fluorescent dye H2DCF-DA and observed under confocal laser scanning microscope with similar image acquisition settings. Representative image of 10 seeds was shown here. 57 3.3.3 Differential expressions of selected OsbZIPs in seed germination Although previous studies already functionally characterized several OsbZIPs involved in ABA signaling as well as abiotic stress tolerance in rice (Hobo et al. 1999; Xiang et al. 2008; Nakagawa et al. 1996; Lee et al. 2003; Lu et al. 2009; Hossain et al. 2010ab; Chen et al. 2012; Liu et al. 2013) but majority of them are still to characterize. After searching publicly available microarray databases in details, 32 putative bZIPs were selected (Table 2) based on (a) either having high expression profile in ABA mediated stress tolerance situations (drought and/or salinity), (b) high sequences difference between indica and japonica spp. and even (c) the genes that do not have expression data . Both upstream and downstream marker genes of ABA signaling pathways were also monitored along with bZIPs. Out of 10 SnRK2s in rice genome, only three of them (SAPK8, SAPK9 and SAPK10) were activated by ABA (Kobayashi et al. 2004), therefore we used them as SnRK2 marker genes. Moreover, expression of two phosphatase (2C) genes (PP2C04, PP2C09) and widely used downstream marker gene (LEA3) expression was also monitored. Moreover, we also checked expression of bZIP05 as bZIP control since rapidly induced by ABA in rice, reported earlier (Nakagawa et al. 1996). As previous report, we observed significant up-expression of bZIP05 after ABA treatment in both Rayada varieties as well as salinity in Sarsari (Fig. 3.3). Among other marker genes, SAPK8 was expressed in all treatment in Rayada rice including anoxia (Fig. 3.3) However, ABA and saline treatments showed highest expression in Rayada variety, Sarsari. Interestingly, it was neither expressed in any treatment of Sub1A rice nor in other upland rices. SAPK9 was mainly expressed in ABA treatment of both Rayada varieties including saline treatment in Sarsari (Fig. 3.3). Slight expression of SAPK10 was observed in Rayada rice in ABA (Kaladigha) and saline treatment (Sarsari). Similarly, expression of PP2C04 was also limited to Rayada rice ABA treatment whereas PP2C09 was different i.e. Kaladigha showed slight expression in both ABA and saline treatments whereas Sarsari showed high expression in saline but not in ABA treatment (Fig. 3.3). LEA3 showed very high ABA, saline treated seed germination of Rayada rice. 58 Fig.3.3 Expression analyses of selected OsbZIPs among three ecotypes/varieties during seed germination. Treatments were germination in presence of i) 10µm ABA, ii) 50mM Nacl, iii) anoxia (ano). For anoxic germination, seed were placed in 50ml tube and filled with degassed then nitrogen humidified (for 15 min) water and capped carefully underwater as no air bubble was trapped inside. RNA was isolated 48 hrs after imbibition and DNase treated RNA from respective samples were used for cDNA synthesis. Equal amount of cDNA were used for PCR amplification of fixed 30 cycles. Names of bZIPs are shown on the right panel. Tubulin was used as internal control whereas bZIP05 as bZIP control as rapidly induced by ABA in rice (Nakagawa et al. 1996) 59 Among 32 bZIP family members, 10 bZIPs (14, 18, 48, 51, 57, 67, 69, 70, 81, and 85) showed no expression among these ecotypes hence omitted in the figure. Twelve bZIPs (13, 40, 42, 44, 45, 53, 59, 63, 68, 71, 77 and 84) were expressed in Rayada varieties either ABA or Nacl or both treatments (Fig. 3.3). Rice seed germinated on Nacl or ABA showed highest number (18) bZIP expression (bZIP05, 13, 24, 26, 37, 39, 40, 42, 44, 45, 53, 59, 63, 68, 71, 77, 78, and 84) mainly in Rayadas. In anoxic germination, slight expression was observed in seven bZIP family members (bZIP05, 13, 45, 63, 71, 77, and 84) also limited to Rayadas. Generally, bZIP expression was either weak or not expressed in Sub1A rice (BRRI51) and other two upland varieties (Fig. 3.3). However, in both Rayada varieties, expression of bZIP84 was huge in both ABA and Nacl treatments. In addition, slight expression was also observed in anoxic germination, highest among all 33 bZIPs including bZIP05 (Fig. 3.3). However, gene expression determination using semi-quantitative RT PCR at seed germination might not reflect the exact expression profile, hence 10 bZIPs (bZIP13,40,42,44,53,59,63,68,71,and 84) were selected for another round of screening using real-time quantitative RT-PCR . Here we used in 3 months old mature plants of three ecotypes/varieties in two experiment sets for both mild and severe water stresses along with exogenous ABA application and recovery status after watering assuming highly expressed genes of either ABA or Nacl treatment in seed germination (bZIP13,40,42,44,53,59,63,68,71,and 84) might reflect clear ABA induce ability. 3.3.4 Expression pattern of OsbZIP84 in mild and severe water stresses After analyzing selected 10 bishops genes quantitative expression patterns (QRTPCR), expression of OsbZIP84 was found pretty interesting (others expression data not shown). In mild water stress in BR52-BP-KD set, OsbZIP84 showed opposite trends among ecotypes, i.e. up-regulated in both upland and Rayada rice varieties in all treatment conditions but down-regulated in flash flood tolerant variety, BRRI52 (Fig. 3.4a). Highest expression (12 fold) was observed in exogenous ABA in upland variety, Boupagol. However, in Rayada rice maximum 60 expression was observed in the recovery stage although both drought and exogenous ABA showed up-regulation too. After severe water stress in another set (BR51-DS-SS), consistent up-regulation of OsbZIP84 was observed with significantly elevated fold in both upland and Rayada rice (Fig. 3.4b). However, in Sub1A rice, (BRRI51) expression was also consistently down-regulated. Fig. 3.4 Expression profile of OsbZIP84 after mild, severe water stresses and recovery in three extreme ecotypes/varieties. Three months old plants were imposed gradual water stress (mild and severe for 8 and 10 days respectively) along with recovery status 30min after watering. Each experiment set consists of one variety of flood tolerant Sub1A (BR51, 52), one upland (BP, DS) and one Rayada rice (KD, SS) and grown in the same pot to reduce environmental interactions. 10µM ABA was sprayed at midday (12.00PM) each day from withholding the water only on designed experiment set. Control plants were 61 sprayed equal amount of ddH2O. DNase treated RNA from respective samples were used for cDNA synthesis. Transcript levels were quantified by qRT-PCR, as described in Materials and Methods. Values are means with ±SD (n=3). 3.3.5 Homology search and possible functional homolog The opposite expression pattern of OsbZIP84 among ecotypes and huge upregulation at recovery stage in Rayada rice and contrasting phenotype of shoot growth after inundation (Sub1a vs. Rayada) made us interested to search homologous genes. We found two homolog members of OsbZIP84 already been functionally characterized, i.e. one in tobacco as repression of shoot growth (RSG) , having the function of GA regulation (Fukazawa et al. 2000) and another in Arabidopsis as VirE2 Interacting Protein 1 (VIP1) (Tzfira et al. 2001). We generated phylogenetic tree based on homologs protein (around 50% and over similarity of different plants) sequences of cultivated and wild species (Fig. 3.5). Among wild species, predicted proteins of Triticum uratu and Aegilops tauschii showed highest similarity (85% and 75%, respectively) whereas other monocot cultivated members (corn, sorghum, barley) showed more than 70% homology. Unfortunately, none of them (either wild or cultivated) are functionally characterized. A. tauschii is considered as predominant source of variation for drought tolerance for cultivated wheat because of great adaptability in extreme habitat (Sohail et al. 2011). However, both of the functionally characterized proteins showed nearly 50% homology only. However, in the recent report of Tsugama et al. (2012) showed that VIP1 might involve in osmosensory regulation through regulating ABA catabolism and that made us more curious to follow gene expression in other situations of Rayada rice. 62 Fig. 3.5 Phylogenetic tree of OsbZIP84 homolog among wild and cultivated species. Phylogenetic tree was derived using the Neighbor-Joining method by MEGA5 (Tamura et al. 2011). Genbank or NCBI accession numbers for the sequences are as follows: Aegilops tauschii (EMT09464.1), Triticum urartu (EMS64321.1) , Hordeum vulgare (BAK03813.1), Brachypodium distachyon (XP_003578876.1), Oryza sativa spp. japonica (NP_001066224.1), Oryza sativa spp.indica (EAY82358.1), Sorghum bicolor (XP_002441871.1), Zea mays (NP_001151647.1), Arabidopsis thaliana (AAM63070.1), Nicotiana tabacum (BAA97100.1) 3.3.6 Putative functions of VIP1 or RSG homolog of rice As the reported two homolog members showed different functions so what might be the function of VIP1 or RSG homolog of rice. There are four possibilities: (a) GA regulation (as RSG) (b) ABA catabolism regulation as VIP1 (Tsugama et al. 2012) (c) both (regulating GA/ABA balance with both VIP1 and RSG functions altogether) or (d) some new functions. Since based on previous reports both drought stress and rehydration conditions, all ABA catabolic genes (CYP707A in 63 arabidopsis) were upregulated whereas after rehydration was significantly high (Kushiro et al. 2004) and similar expression pattern of bZIP84 were observed in Rayadas (Fig. 3.4), hence we conceived OsbZIP84 may have function in regulation of ABA catabolism. To check whether bZIP84 play any such role conceiving the idea of “overexpressed situations might lead to decreased ABA content and subsequently GA/ABA balance will be in favor of GA”. Since GAABA balance still being considered as decisive theory for either loss or maintenance of dormancy (Finkelstein et al. 2008), so to check our hypothesis, we analyzed the expression of OsbZIP84 and ABA catabolic genes in Rayada rice, Kaladigha together with determining ABA content in four different growth and developmental situations where ABA trends to decrease as reported earlier. 3.3.6.1 Inundation Although complete submergence tolerant genotypes such as FR13A and deepwater rices overcome flooding stress by different strategies, but ABA decrease is common to both after flooding (Fukao and Bailey-Serres; 2008 Yang and Choi, 2006). Similarly, a rapid decrease of ABA content after inundation was observed in Kaladigha (Fig 3.6a). Moreover, prolong inundation (45 days) also kept ABA at lowest level (Fig. 3.6a). We also compared ABA between node and internode and observed significant higher ABA in node than internode in both control and inundated conditions (Fig. 3.6b). Internodes become mainly elongated after flooding. Interestingly, GA20ox1, key gene of GA biosynthesis, was strongly up-regulated, especially in 6 hrs. (Fig. 3.6c) when ABA already being lower extents (Fig. 3.6a). However, key ABA catabolic gene, ABA8ox1 was upregulated for couple of hours and after prolong time (24hrs.) down-regulated (Fig. 3.6d). Interestingly, similar expression pattern of ABA8ox1 was observed in OsbZIP84 (Fig. 3.6e) with higher fold level. 64 Fig. 3.6 Time course ABA content and selected genes expression after inundation in Rayada variety, Kaladigha. (a) Time course ABA content in internode ,b) comparative ABA content between node and internode, gene expression in internodes of (c) GA20ox1, (d) ABA8ox1 and (e) OsbZIP84 after inundation. Rayada variety, Kaladigha plants (3 months old) were inundated in water tube with increasing water level keeping top leaves above water for 45 days. ABA and gene transcript levels measured by RIA and qRT-PCR, respectively, as mentioned in Materials and methods. Values are means with ±SD (n=3). 3.3.6.2 Seed maturation On the progression of seed maturity, embryo growth ceases, accumulates storage products and most importantly, develops the capacity of dormancy and desiccation tolerance (Gutierrez et al. 2007). Moreover, in most studied plants, ABA content increases during initial seed development and decreases at maturity (Bewley et al. 2013). Similarly, in this study, we also observed higher ABA at milk and dough stages and progressively declined to maturity (Fig. 3.7a). Similarly, moisture content declined progressively whereas fresh weight increased 65 instead (Fig. 3.7f). All three ABA catabolic genes were down-regulated in milk and dough stages (Fig. 3.7b,c,d), however, ABA8ox1 was significantly downregulated (more than 20 folds) suggesting its role on ABA content determination or ABA homeostasis. But, in developing barley grains main regulator of ABA rise was reported ABA biosynthesis (Sreenivasulu et al. 2010). In mature seeds, ABA8ox1 expression was same as dry seed whereas other two ABA catabolic genes (ABA8ox2, 3) were partially up-regulated instead. Interestingly, OsbZIP84 was also found significantly downregulated during all three stages (Fig. 3.7e) comparing to dry seed but huge down-regulation showed in mature seeds. So gene expression of ABA8ox1 and OsbZIP84 in mature seed suggested the decisive role on keeping high ABA in mature seed. Fig. 3.7 Time course ABA content and selected genes expression during seed maturation in Rayada variety, Kaladigha. (a) time course ABA content and gene expression of (b) ABA8ox1, (c) ABA8ox2 (d) ABA8ox3, (e) OsbZIP84 during seed developmental stages , (f) changes of fresh weight and moisture content. Milk, dough and mature stages were of 7, 14, 28 days after anthesis, respectively. Dry seeds were used from the previous harvest (180 DAH) stored in desiccator. ABA and gene transcript levels measured by RIA and qRT-PCR, respectively, as mentioned in Materials and Methods. Values are means with ±SD (n=3). 66 3.3.6.3 Water stress recovery Numerous experiments in Arabidopsis, barley and beans (Kushiro et al. 2004; Saito et al. 2004; Millar et al. 2006; Yang and Zeevaart 2006) revealed that under stress and recovery, ABA catabolism was mainly regulated at the transcriptional level. Water stress increases the expression of ABA catabolic genes (Kushiro et al. 2004). Alike to previous reports, we also observed very high ABA accumulation after water stress (0 min of Fig.3.8a). But high ABA level was sharply decreased after watering with time progression (Fig.3.8a). Interestingly, 50% of ABA decreased in just 2 hrs. ABA8ox1 was significantly up-regulated for hour and pick expression was observed at 30 min of watering (Fig. 3.8b). Apparently it viewed down-expression after two hours, but actually it was then still up-regulated because we compared with water stressed condition. Similarly, we observed up-expression of OsbZIP84 in both mild and severe water stresses (in Fig. 3.4 a,b) in both upland and Rayada varieties. So apparently gene expression of OsbZIP84 showed partial up-regulated initially in recovery stage, down-regulated later on (Fig 3.8c); actually highly up-regulated if we compare with unstressed normal plants. Fig. 3.8 Time course (a) ABA content , (b) ABA8ox1 and (c) OsbZIP84 expression during water stress recovery in Rayada variety, Kaladigha . Rayada variety, Kaladigha plants (3 months old) were water stressed by withholding watering for 5 days, decreasing relative water content around 70%. Then plants were rewatered and leaf samples (usually 2nd leaf) were collected. ABA and gene transcript levels were measured by RIA and qRT-PCR, respectively, as mentioned in Materials and Methods. Values are means with ±SD (n=3). 67 3.3.6.4 Seed Germination Several hormone interact during seed dormancy release and germination, among them GA promotes whereas ABA suppresses germination (Kucera et al. 2005). Decrease of ABA level is prerequisite of germination (Weitbrecht et al. 2011). We measured ABA content in imbibed seeds of Rayada variety, Kaladigha in time course and observed sharp decrease in the initial hours of imbibition and later on keeping low and steady (Fig. 3.9a). Oppositely, the water absorption pattern was observed quick and sharp increase during the first few hours of imbibition, gradual then steady afterwards (Fig. 3.9d). Exogenous ABA showed a dose dependent effect, 0.5 µM ABA delayed the germination and growth whereas 10µM ABA completely restricted shoot development and growth but root growth was suppressed but still developed (Fig. 3.9e). ABA8ox1 was increasingly upregulated till 4 hrs (Fig. 3.9b), later on up-regulated but lesser extent whereas ABA8ox 2 and 3 was decreased instead (data not shown). Remarkably, OsbZIP84 was significantly up-regulated during first 8 hrs of imbibition (Fig. 3.9c). Fig. 3.9 Time course (a) ABA content and gene expression of (b) ABA8ox1, (c) OsbZIP84 and (d) water absorption pattern during imbibition and (e) dose dependent inhibitory effect of ABA on seed germination in Rayada variety, Kaladigha. We also examined time course gene expression level of all 5 NCED genes after imbibition. Interestingly, none of them were found to up-regulated after 68 immibition rather decreased instead (Fig. 3.10). As none of the NCED is upregulated, then certainly main regulator of ABA decrease after imbibition is ABA 8’-hydroxylases. Fig. 3.10 Time course relative gene expression of NCEDs after imbibition. RNA was isolated from imbibed seeds of different time point and compared with that of dry seed (0hrs) . Values represent average of triplicate samples. 3.3.7 Dormancy phenotype of osbzip84 Among tested four different stages of growth and development stages, OsbZIP84 and ABA8ox1 showed almost similar expression pattern except mature seeds during seed maturation (Fig. 3.7b, e). All these results clearly suggested involvement of OsbZIP84 in regulation of ABA catabolism. To confirm the function of OsbZIP84, we collected, screened and identified T-DNA insertional homozygous mutant of OsbZIP84. Mutant osbzip84 clearly showed dormancy phenotype in seed germination (Fig. 3.11a). Seed germination was significantly delayed than wild type (Fig. 3.11b). Almost 90 percent of control seeds germinated within 36 hrs of imbibition, however a few mutant seeds germinated, mostly rupture appeared (Fig. 3.11b). To reach 100% mutant seed germination, it 69 required 3-4 days. Seedling establishment and growth was significantly retarded in mutant. We measured root (Fig. 3.11 c, d) and shoot length (Fig. 3.11e, f) in 6 and 10 days after imbibition and always found significant reduction than control. To reconfirm the phenotypic explanation, pharmacological approach was used. Wild type seeds were germinated in presence of ABA catabolic inhibitor, diniconazole and found almost similar phenotype (Fig. 3.11a inset). Diniconazole is a potent inhibitor of ABA 8'-hydroxylase regarded as ABA catabolic inhibitor (Kitahata et al. 2005). These similar phenotypes of mutant and ABA catabolic inhibitors strongly supported our hypothesis of involvement of OsbZIP84 in ABA catabolic regulation. Fig 3.11 Germination and seedling growth performance of osbzip84. (a) comparative phenotype of osbzip84 with wild type and in presence of ABA catabolic inhibitor, diniconazole in control seeds (inset), (b) germination percentage and comparative root (c, d) and shoot length(e, f) after 6 and 10 days, respectively. Seeds of 30 DAH were used for germination test in both wild and 70 mutants. Each batch contained 10 seeds with three replicates. The symbol (*) indicates the significance level, * for P<0.05, *** for P<0.001. 3.3.8 Elevated ABA in osbzip84 seeds but reversed in water stress Apart dormancy phenotype, osbzip84 also showed another feature; significantly reduced grain weight (Fig. 3.12b) and grain size (Fig. 3.12c) in two successive generations of homozygous mutant. Although dormancy phenotype of osbzip84 confirmed our hypothesis of ABA catabolism regulation, but reduced grain weight and size lead us to check the other possibilities. Several factors can regulate grain filling, thus grain weight and size such as carbon assimilation rate , remobilize of pre-stored in vegetative tissue (Yang and Zhang 2006) , shorter grain filling period (Kobata and Uemuki 2004), high temperature (Yamakawa et al. 2007) as well as drought. However, to ascertain our hypothesis, we measured ABA content in both wild type and mutant seed. Interestingly, mutant seeds showed significantly elevated ABA content (nearly double) (Fig. 3.12a) despite its reduced grain weight (Fig. 3.12b). So certainly delayed seed germination was due to elevated ABA content despite reduced size and weight. Although osbzip84 showed elevated ABA in seed but significant reduced ABA content was observed in leaves in normal (Fig. 3.12d), dehydrated (Fig. 3.12e) conditions (detached leaf). So it seems OsbZIP84 might have played other important role in ABA homeostasis, not only seed dormancy; need further investigation. 71 Fig. 3.12 Change of ABA content, grain size and weight of osbzip84 (a) elevated ABA content in mutant seeds, (b) reduced grain weight (c) grain size and comparative change of ABA content in (d) normal (e) dehydrate leaves of osbzip84. ABA was determined using RIA methods described in materials and methods. Random seed samples were used to compare grain weight and size. ABA content of detached leaf was determined after air drying for 30 min. All ABA samples were replicated thrice. The symbol (**) represented significance level at P<0.01 72 3.4 Discussion In this study, we first examined and compared the physiological basis of dormancy in deep water and Rayada rice then searched for the plausible gene of interest that regulate ABA catabolism through expression analysis of 32 OsbZIPs in three extreme ecotypes/varieties during seed germination . After analyzing the expression and functional homologs, we selected OsbZIP84 and examined ABA content and expression analysis of ABA catabolic genes in four growth and developmental stages. Our results clearly demonstrate that OsbZIP84 play an important role in ABA homeostasis. 3.4.1 Natural variation of seed dormancy Natural variations among accessions are useful to study the molecular mechanism of seed dormancy. Variation of seed dormancy among Arabidopsis accessions exists, some accession such as Cape Verde Islands (Cvi) possess strong seed dormancy whereas the laboratory strain Landsberg erecta (Ler) and Columbia (Col) possess low dormancy. Similarly, after ripening requirement also significantly different between them , dry storage of 74-185 days are required to achieve 50% germination for Cvi while Ler only requires 12-17 days (AlonsoBlanco et al. 2003). Through QTL analysis of recombinant inbred line derived from Cvi and Ler identified major QTL of seed dormancy, DOG1 (delay of germination), common in dormant accessions. Recent studies identified DOG1 as one of the key regulator as dog1 shows completely lack of seed dormancy without any other pleiotropic phenotypes except reduced seed dormancy (Nakabayashi et al. 2012). DOG1 regulate seed dormancy through enhanced ABA and sugar responsiveness through enhanced ABI4 (Teng et al. 2008). Wide variation of dormancy was observed in both cultivated rice species O. sativa and O. glaberrima. But generally both of them possess less intense seed dormancy comparing to wild species. Mean dormancy period among 137 varieties of O. sativa found ranged from 0 to 110 days (Roberts 1961) whereas that of 35 cultivars of O. glaberrima showed 85-155 days (Misra and Misro, 1970). However, among wild species O. glumaepatula and O. rufipogon, showed the 73 strongest dormancy (180 days) (Veasey et al. 2004). However, the degree of dormancy significantly varies on harvesting stage ripeness as well as storing temperature. Cooler temperature prolongs dormancy whereas higher temperature alleviate dormant period (Veasey et al. 2004). Numerous studies were performed to identify QTLs of rice seed dormancy but successful identification of relevant gene is scarce. To date , only successful dormancy regulatory gene has been characterized in rice from QTLs , sdr4 through backcross inbred line derived from the japonica cultivar Nipponbare and the indica cultivar Kasalath (Sugimoto et al. 2010). Since Rayadas possess yearlong life cycle, hence lack of seed dormancy seems an adaptive feature of Rayada rice. Interestingly, as mentioned in the preceding chapter, the climatic features such as the rainfall, humidity and temperature drop to the lower extent during the seed maturation period to reduce inadvertent germination. 3.4.2 ABA, ROS and seed dormancy: linked altogether Recently, plenty of research has identified reactive oxygen species (ROS) as signal molecules regulating growth and development as well as biotic and abiotic stresses (Rodriguez et al. 2004; Jones et al. 2007; Zhang et al. 2010; Zhang et al 2001), programmed cell death (Bethke and Jones 2001) including dormancy alleviation, after-ripening, and germination (Oracz et al. 2007; El-MaaroufBouteau and Bailly 2008; Ye et al. 2012a). Several cellular organelle such as chloroplasts, mitochondria, peroxisomes (Neill et al. 2002, Shigeoka et al, 2002) as well as some enzyme complexes such as class III peroxidases, oxalate oxidases, amine oxidases, lipoxygenases, quinone reductases and plant NADPH oxidases (NOXs or rboh), can generate ROS in cells. . Production of H2O2 after imbibition was observed in several plants, i.e. barley (Ishibashi et al. 2012), soybean (Puntarulo et al. 1988), maize (Hite et al. 1999), wheat (Caliskan and Cuming 1998), and Zinnia elegans (Ogawa and Iwabuchi, 2010), rice (Ye et al 2012), Arabidopsis (Liu et al. 2010). Oppositely, exogenous application of ROS scavenger significantly suppressed seed germination 74 (Ishibashi et al 2012, Ye et al. 2012a). Here we observed freshly harvested Rayada seeds accumulated excessive ROS in embryonic axis whereas freshly harvested seeds of typical deepwater rice accumulated sparingly low amount (Fig. 3.2). Moreover, exogenous ABA suppressed ROS accumulation as well as seed germination of freshly harvested Rayada seeds (data not shown). Moreover, ABA content was significantly reduced than typical deepwater rice (Fig. 3.1b). So it was clear huge ROS accumulation was due to reduced ABA content. Similarly, ROS accumulation of non-dormant seeds in Arabidopsis was evident than that of dormant seeds (Leymarie et al. 2012). Moreover, NADPH oxidase mutant (rbohD) showed deep dormancy due to less H2O2 production capacity. So ROS accumulation after imbibition presumably played key role in defining seed germination of freshly harvested seeds of Rayada variety. 3.4.3 Regulation of ABA catabolism: function of OsbZIP84 in Rayada rice The balance between ABA biosynthesis and catabolism are determinants of ABA content. NCEDs are the main regulatory genes of biosynthesis whereas ABA8oxs are that of catabolism (Qin and Zeevaart 1999; Nambara and Marion-Poll 2005). Five NCED and three ABA8ox genes have been reported in rice (Saika et al. 2007; Zhu et al. 2009). Numerous experiments reported the importance of ABA catabolism regulating ABA levels and homeostasis in barley (Chono et al. 2006; Millar et al. 2006), Arabidopsis (Millar et al. 2006; Okamoto et al. 2006; Umezawa et al. 2006; Okamoto et al. 2010), bean (Yang and Zeevaart 2006), maize (Ren et al. 2007) and rice (Yang and Choi 2006; Saika et al. 2007; Zhu et al. 2009; Ye et al. 2011). The significance of ABA catabolism after submergence or flooding induced petiole or internode elongation was reported earlier in flooding-tolerant Rumex palustris (Benschop and Jackson 2005) and rice (Yang and Choi 2006), respectively. Decreased ABA content is a prerequisite for increased GA responsiveness and that leads to the enhanced elongation as exogenous ABA can completely demolish internode elongation capacity (Hoffmann-Benning and Kende 1992; Kende et al. 1998). Decreasing ABA and increasing phaseic acid (PA) level were reported in rice after flooding despite the 75 rice types whether Sub1A (Fukao and Bailey-Serres 2008) or deepwater (Yang and Choi 2006) even in japonica rice (Saika et al. 2007). Among three OsABA8oxs, only the expression of OsABA8ox1 was significantly increased and the induction was controlled partially by ethylene (Saika et al. 2007). We also observed similar pattern of ABA decrease (Fig. 3.6a) and ABA8ox1 expression increase (Fig. 3.6d) after inundation in Rayada rice. In addition, OsbZIP84 was also showed similar pattern of expression both after inundation (Fig. 3.68e) and imbibition of seeds (Fig. 3.9c) suggesting the possible connection between the genes in ABA homeostasis. In water stress recovery stage, we observed significant up-expression of OsABA8ox1 in 30 min after rehydration (Fig. 3.8b). Similarly, up-expression of OsABA8ox1 was also observed earlier in rice (Ye et al. 2012a). Both upland and Rayada rice showed huge up-expression of OsbZIP84 in water stress and recovery stages (Fig. 3.4a,b). However, expression of OsbZIP84 was consistently downregulated in two Sub1A rice both in mild and severe stresses (Fig. 3.4) made the explanation difficult. Perhaps either Sub1A or semi-dwarf nature might have interactions or some other factors involved herein; it certainly demands further study. However, Sub1A showed cross talk with drought tolerance (Fukao et al. 2011) and ubiquitous expression of Sub1A (Ubi:Sub1A) delayed both seed maturation and germination (Fukao and Bailey-Serres 2008) suggesting Sub1A might interfere the process. 3.4.4 Elevated ABA in osbzip84 seed results in dormancy phenotype ABA plays central roles in the induction and maintenance of seed dormancy. Although exogenous ABA restricts the germination potential and delay endosperm rupture in rice, Arabidopsis and barley (Muller et al. 2006) but high endogenous ABA accumulating mutant (cyp707a1a2a3) in Arabidopsis showed deep seed dormancy (Okamoto et al. 2010) suggesting ABA catabolism is one of the key factors that regulate seed dormancy and germination. ABA catabolic triple mutant (cyp707a1a2a3) accumulated 70-fold higher ABA than wild type in 76 Arabidopsis and remained at high levels after seed imbibition (Okamoto et al. 2010) whereas cyp707a2 mutants accumulated 6 fold higher ABA (Kushiro et al. 2004). Similarly, high accumulation of ABA was also observed in osbzip84 (Fig. 3.12a) connecting the dormancy phenotype (Fig. 3.11a). Lack of seed dormancy is one of the main features of Rayadas (Table 2). Moreover, Rayadas showed high expression of OsbZIP84 after imbibition (Fig. 3.9c) suggested the faster catabolic activity reduced the ABA thus lack seed dormancy. Tsugama et al. (2012) clearly demonstrated using both in gel shift and chromatin immunoprecipitation analyses that VIP1 (OsbZIP84 homolog in Arabidopsis) directly bound to DNA fragments of the CYP707A1/3 promoters. All these results clearly suggest the importance ABA 8-hydroxylases for regulating seed dormancy by the regulation of OsbZIP84. However, OsbZIP84 might play dual regulatory role in ABA homeostasis. Because after water stress, OsbZIP84 showed up-expression in both Rayada and upland rice (Fig 3.4ab and 3.8c), consequently osbzip84 accumulated significantly less ABA after water stress imposed by air drying of detached leaves (Fig.3.12e), even possessed less ABA in normal leaves (Fig.3.12d) . These decreased ABA content signified the increased expression of OsbZIP84 in water stress conditions. Moreover, osbzip84 displayed increased water loss and slower recovery after water stress and recovery (data not shown). Certainly, further experiments are essential to confirm the dual regulatory role of OsbZIP84 in ABA homeostasis. In summary, lack of dormancy of Rayada might be an adaptive trait and several concerted features; for instance, less ABA content in freshly harvested seeds; faster ABA catabolism and enhanced ROS accumulation after imbibition, collectively might involved in the regulation of dormancy. Gene expression analysis of OsbZIP84 and OsABA8ox1 together with ABA content in four growth and developmental stages suggest the function of OsbZIP84 in ABA catabolism. Dormancy phenotype of osbzip84 along with similar phenotype after ABA catabolic inhibitor , diniconazole conclude it’s function in ABA catabolism. 77 78 Chapter 4 Shoot development under submergence is regulated by ROS and riboflavin in Rayada rice 4.1 Introduction Rainfed and irrigated lowland including flood-prone ecosystems consist of over 90% of total rice cultivation area; account around 95% of global rice production (Maclean et al. 2002). Direct seeding of rice is the popular method of seeding in these ecosystems because of its operational simplicity, low labor cost along with week’s earlier maturation (Balasubramanian and Hill 2002). However, sudden sallow flooding due to heavy rain fall or waterlogging limit the seedling establishment in direct seeded fields since rice is sensitive to submergence during seed germination (Ismail et al. 2012). All other cereals (wheat, maize, barley, oat and sorghum) are much more sensitive to submergence during seed germination, even cannot germinate (Ismail et al. 2012). In comparison with them, rice is more tolerant to submergence; efficiently able to germinate under submergence. However, the tolerance is only limited to coleoptile development but unable to develop subsequent shoot and root or fails to establish seedling. Complete submergence always leads to oxygen deficiency where lack or limited oxygen (anoxia or hypoxia) limit the energy budget as O2 is the ultimate electron acceptor. Moreover, anoxia or hypoxia also restricts the function of some enzymes that restrict the mobilization of stored carbohydrate (Ismail et al. 2012). Molecular mechanism of flash and prolong flood tolerance at vegetative stage of rice has been already revealed. Flash flood or complete submergence tolerance, governed by a single gene SUB1A restricting GA responsiveness, identified from landrace, FR13A (Xu et al. 2006; Bailey-Serres et al. 2010). However, SUB1A regulating submergence tolerance is limited to vegetative stage, not involved in seed germination stage (Magneschi and Perata 2009). Oppositely, prolonged either stagnant or rising flood tolerance of deepwater or floating rice is regulated by snorkel genes (SK1, SK2), enhancing GA responsiveness by unknown manner (Hattori et al. 2009). However, flooding tolerance during seed germination and early seedling growth and establishment is still in obscure. 79 4.1.1 Submergence tolerance during seed germination: a rare trait Submergence tolerance in seed germination of rice or the feature of shoot and root development under submergence is a rare trait. Attempts were made in past to identify submergence tolerance capacity and integrate in breeding program but resulted limited success, mainly lack of tolerant genotypes and inadequacy of comprehensive knowledge of physiological basis of tolerance (Ismail et al. 2009). Recently, after large scale screening of over 8000 germplasms, only few varieties (19 only) were identified that can successfully establish seedlings under submergence (Angaji et al. 2010). However, subsequent replicated trails limited the tolerant capacity to only 5-6 genotypes i.e. Khao Hlan On, Khaiyan, Kalongchi, Nanhi, Ma-Zhan Red and Cody (Angaji et al. 2010; Ismail et al. 2009; Miro and Ismail 2013). Interestingly, out of these six, two of them, Khaiyan, Kalongchi are traditional varieties of Bangladesh. In addition, Khao Hlan On and Nanhi are traditional varieties of Myanmar and India, respectively (Angaji et al. 2010). Interestingly, only these two countries share border with Bangladesh along with cultural and agricultural practices. So it might not be unlikely that tolerant varieties might have developed or originated in similar region, even in same territory. Remarkably, preliminary screening of IRRI identified another Bangladeshi deepwater rice variety Dholamon 64-3 as a tolerant genotype with high survival percentage (80%) (Fig. 1.8a) (Angaji et al. 2010). Interestingly, some germplasms adapted to deepwater conditions of Bangladesh were also reported superior performance of seedling establishment after submergence (Yamauchi et al. 1993). Since Rayadas are more primitive among deepwater rices having multiple stress tolerance capacity and also endemic to certain deepwater area of Bangladesh, so the capacity of submergence tolerance during seed germination in Rayada rice certainly demands exploration. 4.1.2 Physiological features of tolerant genotypes Some physiological differences between tolerant and intolerant genotypes have already been revealed (Ismail et al. 2009; Ismail et al. 2012; Miro and Ismail 2013). Sugar metabolism through the glycolytic pathway plays the key role in 80 energy production; hence regulation of this process under submergence is very critical for seedling establishment. After submergence starch concentration is progressively declined in tolerant lines but remained high in intolerant genotypes suggesting the importance of carbohydrate mobilization after submergence (Ismail et al. 2009). Moreover, amylases play key role in breakdown of starch of rice during seed germination; so total amylase activity expectedly is observed keeping significantly high in tolerant genotypes than intolerant one. Notably, high amylase activity is positively correlated with shoot and root growth along with seeding survival (Ismail et al. 2009; Ismail et al. 2012; Miro and Ismail 2013). Similarly, submergence tolerant genotypes also keep higher soluble sugars during seed germination and seedling establishment stage. Among α-amylases, only RAmy3D is reported having differential expression between tolerant to intolerant genotypes, others showed almost same with no remarkable difference (Ismail et al. 2009; Ismail et al. 2012). 4.1.3 Alternative strategy to reveal regulatory factors of shoot development under submergence As submergence tolerance is a rare phenomenon in rice and no specific indication of tolerance mechanism, especially in context of molecular signaling, hence we have attempted with an unconventional strategy to develop shoot in intolerant varieties first in artificial induced condition under submergence. Then we have tried to hinder or augment shoot development by interfering plausible interacting pathways with known interacting chemicals to reveal the factors that interact with the submergence tolerance. We, for the first time have successful developed a condition inducting Rayada rice phenomenon and identified several positive and negative regulators that either induce or hinder the shoot development process under submergence. 81 4.2 Materials and methods 4.2.1 Plant materials and growth conditions Rice seeds of diverse origin and type were used in these studies. It included Rayada (Kaladigha, Beto), typical deepwater rice (Habigonj aman VIII), flash flood tolerant SUB1A rice (BRRI51, BRRI52) with BRRI52 background variety BR 11, indica variety (YD6) and japonica type (Nipponbare, Donjing). YD6 is popular Chinese varieties whereas Nipponbare, Donjing are varieties of Japanese and Korean origin, respectively; rest others are Bangladeshi origin. Intact seeds (with hull) were used to ascertain the natural condition and ability whereas dehulled seeds were used in some experiments for downstream RNA and ABA extraction, mentioned in respective figures. Normally 50-100 seeds of YD6 were used in all treatments with three biological replicates whereas 10-20 seeds for other varieties including Rayada. Control (air) seeds were germinated on moist filter papers in dark at 28ºC. Submerged germination was setup with degased deionized distilled water (ddH2O) in DURAN laboratory bottle of 500ml with height of 17 cm submergence. Special precautions were taken to avoid air bubble trapped inside bottle. 4.2.2 Different treatments Different treatments were used with several known chemicals of specific pathways .Most of the chemicals used in these studies were of reagent grade, mostly from Sigma-Aldrich. a) H2O2: Freshly prepared 10 mM H2O2 was used for supplementation. b) Vitamin B complexes: Different vitamin B complexes (B1, B2, B3, B6, B9) were used in the experiments of fixed 200 µM concentration. However, different concentration of riboflavin (B2) were used in later experiments and dissolved only in ddH2O (within in the limit of solubility in water) although riboflavin is slightly soluble in water. Despite high solubility in alkalies, we avoided due to instability of riboflavin in alkaline solution. Since riboflavin is highly photolabile in solution, hence we warped the germination setup with 82 aluminum foils to protect light for first four days (product information sheet Sigma-Aldrich). Then the setup was unwrapped and placed under light in green house conditions with 14/10 hrs day-night cycle. c) ABA: A concentration 1µM ABA was used in submergence; the concentration is not completely inhibitory for seed germination in aerobic condition. High concentration of ABA (10 µM) was used to follow the inhibition pattern on root and shoot growth in Rayada rice varieties, Kaladigha and Sarsari. d) NaN3: Sodium azide is a potent inhibitor of mitochondrial complex IV. A concentration of 1 mM of NaN3 was supplemented in submergence either singly or with riboflavin and H2O2. e) Nacl: Mild salinity stress was imposed with 50 mM Nacl (equivalent to 3.1 dSm-1 EC and 0.25 MPa of osmotic pressure). f) PEG mediated water stress: 15% PEG 6000 based on Kimura B nutrient solution were used to induce artificial water stress for months old seedlings of different varieties. Water stress experiments were carried out in phytotron at 28°C, 60% relative humidity with 16/8hrs day night cycle. 4.2.3 Uncoating of Rayada seeds Seeds of Rayada variety Kaladigha were uncoated manually using single edge razor blade. Special attention was paid to embryo region to avoid injury. 4.2.4 Electric conductivity (EC) measurement Orion 105 basic handheld conductivity meter was used for EC measurement. Intact and uncoated 100 seeds of YD6 variety were imbibed in ddH2O or 10mM H2O2 and the change of EC was observed in time course among control, coatless and H2O2 treatment. All experiments were done at least of three biological replicates. 83 4.2.5 RNA extraction and quantitative real-time (qRT)-PCR RNA was extracted from respective samples (only coleoptile or caryopsis) using the Agilent Plant RNA isolation Mini Kit (Qiagen, Hilden, Germany) and then digested with RNase-free DNase I (Amersham, USA) to eliminate genomic DNA contamination. First-strand cDNA was synthesized with oligo (dT) primers using a Promega cDNA kit as according to the manufacturer’s instructions (Promega, USA). Transcript levels of each gene were measured by qRT-PCR using a Mx3000p QPCR System (Agilent, USA) with iQ SYBR Green Supermix (BioRad, USA). The data were normalized to the amplification of a rice actin-1 gene. Primer sequences of genes used for qRT-PCR expression analysis are as follows: RAmy1AF-5’CTACGCATACATCCTCACCCAC3’, RAmy1AR-5’CTGCCGGTTTCTGATTGACAC3’, RAmy3DF-5’TAGCAGCAACTCACTATCGAACA3’, RAmy3DR-5’CAGTTGAAACCCTGGAAGAGGAC3’, RAmy3EF-5’TGGAGAAGAAGCAGAGTTGAGAC3’, RAmy3ER-5’TAAACCCCTGGAAGAGAACTTGG3’, Actin1F-5’AGAGGAGCTGTTATCGCCGTCCTC3’ and Actin1R-5’GCCGGTTGAAAACTTTGTCCACGC 3’. 4.2.6 Extraction and measurement of ABA Measurement of endogenous ABA content was carried out using the radioimmunoassay (RIA) method as described by Jia et al. (2012). 200 mg seedlings of 5 days after imbibition was ground in liquid nitrogen and homogenized in 1 ml of distilled water and shaken overnight at 4°C. The homogenates were centrifuged at 12000g for 20 min at 4°C and the supernatant were directly used for ABA assay. The 450 μl reaction mixture comprised of 200 μl of phosphate buffer (pH 6.0), 100 μl of diluted antibody (Mac 252) solution, 100 μl of [3H] ABA (@ 8000 cpm) solution, and 50 μl of crude extract. The 84 mixture was then incubated for 45 min at 4°C and the bound radioactivity was measured in pellets precipitated with 50% saturated (NH4)2SO4 with a liquid scintillation counter. 85 4.3 Results 4.3.1 High ABA restricts shoot development, not root Multiple stress tolerances are one of the unique features of Rayada rice varieties (discussed in Chapter 2). A 5 µM ABA concentration was reported as completely inhibitory to rice seed germination (Ye et al. 2012a; Kim et. al. 2012). However, we germinated Rayada seed on as much as twice of the inhibitory concentration. Exogenous 10 µM ABA is very high concentration during seed germination; rice seeds never reach such a high level of ABA concentration, even in deep dormant seed. Most others varieties we tested showed complete inhibition of seed germination on such high concentration of ABA. However, suppression of germination in Rayada varieties was also observed, but not at inhibitory level. Inhibitory effect of high concentration ABA was observed mainly on shoot development (Fig. 4.1) in two Rayada varieties. Although reduced but remarkable root growth was still observed in both of the Rayada varieties we tested (Fig, 4.1, inset zoom in figures). These data clearly indicated that high ABA tolerance of Rayada seeds during seed germination. Moreover, root development was less sensitive to ABA but shoot development was opposite to root. Similar results of preferential shoot suppression of ABA were reported earlier in water stresses condition (Sharp and Davies 1989). Mild salinity stress imposed by 50 mM Nacl during seed germination, however, did not restrict either root or shoot development but growth in two Rayada rice varieties (Fig. 4.1). Lin and Kao (1995) reported that GA3 counteracted Naclinhibited shoot growth, but not root growth suggesting differential behavior of root and shoot growth and development after stress during seed germination as we observed after high exogenous ABA. Since Rayada rice possess high elongation ability after flooding (Fig. 2.4c,d) or high GA responsiveness (due to SK genes) and GA counteracts salinity induced shoot growth inhibition(Lin and Kao 1995), and moreover, even freshly harvested Rayada seeds possess less ABA along with both partial and complete submergence tolerant, so we hypothesized Rayada rice might be more tolerant to submergence tolerance during seed germination, especially to shoot growth. 86 Fig. 4.1 Effect of high ABA and mild salinity on seed germination of Rayadas. Upper panel (Kaladigha), Lower panel (Sarsari) 5 days after imbibition. Salinity and ABA stress was imposed by 50 mM Nacl and 10 µM ABA, respectively. Representative image taken 5 days after imbibition from three replicates. Zoom in image of a seed showing growth inhibition pattern on shoot and root (top right (Kaladigha), bottom right (Sarsari). 4.3.2 Rayada seedlings: more tolerant to composite stress Some of the natural stress situations are with pattern of composite stress i.e. multiple stresses together. For instance, in drought the main stress imposed due to soil water deficit, however, some other stress also occasionally developed along with such as high daytime temperature, reduced nutrient availability as well as sometimes increased salinity (Oliver et al. 2011). Similarly, salinity also imposes two types of stresses i.e. ionic along with oxidative stress. Hence we attempted to examine the composite stress tolerance capacity of Rayada rice to modern rice varieties imposing successive stress. Month’s old seedlings were pretreated with 10mM H2O2 for 24 hrs then imposed water stress using 15% PEG 6000 on Kimura B nutrient solution. Within 12 hrs after imposing water stress, H2O2 87 pretreated plants of both modern varieties (BR 11 and BRRI 52) showed water stress symptoms (leaf rolling and flaccid leaves) (Fig. 4.2 a) and relative water content (RWC) decreased to 68.23 and 63.57 % respectively. However, no significant difference was observed between BR11 and BRRI 52. BR11 is the background variety of SUB1A introgressed BRRI 52. So it reflected noninvolvement of SUB1A in these composite stress tolerances. After 4 days water stress, most of the plants of modern varieties (either BR 11or BRRI 52) died in H2O2 pretreated set (Fig.4.2b) whereas Rayada varieties showed water stress symptom only (Fig.4.2c), still possess significant capacity to recover from water stress (Fig.4.2c). Around 15% plants of these modern varieties were recovered after stress withdrawal whereas most of the plants of Rayada varieties recovered successfully (Fig.4.2d). These results clearly represented the composite stress tolerance of Rayada rice, especially after oxidative stress. Fig. 4.2 Composite stress tolerance capacity of Rayadas to modern rice varieties. a) oxidative following water stress phenotype of modern varieties 12 hrs after water stress impose, b) that of 4 days , c) comparative phenotypes between 88 Rayadas and modern varieties after 4 days , d) recovery performance 2 days after withdrawing water stress. Month’s old seedlings were pretreated with 10mM H2O2 for 24 hrs then imposed water stress using 15% PEG 6000 based on Kimura B nutrient solution. Composite stress experiment was carried out in phytotron at 28°C , 60% relative humidity with 16/8hrs day night cycle. Representative images of five replicates were shown here. 4.3.3 Augmentation of seed germination by hydrogen peroxide We examined effect of hydrogen peroxide on seed germination both in aerobic and submerged conditions of diverse varieties and it clearly represented that H2O2 speeded up the germination and the phenomenon of speedup by H2O2 was global i.e. irrespective of varieties (Fig.4.3a). Generally, seed germination under submergence was delayed than aerobic control, almost a day. Interestingly, all the varieties showed elevated germination performance after H2O2 treatment in both aerobic (after 30 hrs. of imbibition) and submerged conditions (48 hrs.) and it was almost doubled although variation observed among varieties (Fig.4.3a). Differential effects of gradient H2O2 on root and shoot growth were observed in both aerobic and submerged conditions. In aerobic condition, increasing H2O2 concentration trended to decrease root development and growth whereas shoot growth and development was much more tolerant to oxidative stress (up to 50mM), keeping no change , even promoted shoot growth. (Fig. 4.3b upper panel). Likewise, in submerged condition lower concentration of H2O2 increased the coleoptile length, however, 5mM or above concentration trended to decrease coleoptile length (Fig. 4.3b, lower panel). No root development was observed in submerged condition, even after H2O2 supplementation. We also examined change of ABA content after 10mM H2O2 during seed germination and seedling establishment stage in Rayada rice variety, Kaladigha. It clearly showed H2O2 treatment significantly reduced ABA of seeding establishment stage (Fig. 4.3c). 89 Fig. 4.3 Effect of H2O2 on seed germination, coleoptile growth and ABA content. (a) effect of H2O2 in seed germination in aerobic and submerged conditions (b) effect of gradient H2O2 concentration on root and shoot growth in aerobic (upper panel) and submerged conditions (lower panel) , (c) change of ABA content after H2O2 treatment in Rayada rice variety, Kaladigha seedlings, 4 days after imbibition in aerobic condition. Germination rate was observed after 30 hrs imbibition in aerobic condition whereas submerged after 48 hrs. 4.3.4 Shoot development of Rayadas under composite submergence stress Since we observed excellent performance of composite stress tolerance (oxidative followed water stress) of Rayada rice varieties (Fig. 4.2c, d) and tolerance of shoot or coleoptile development after oxidative stress in aerobic and submerged condition, respectively (Fig. 4.3b) hence we imposed Rayada seeds with another 90 type composite stress during seed germination (submergence together with oxidative stress). Surprisingly, we observed the unique feature of shoot development under such stress condition in Rayada variety, Kaladigha (Fig. 4.4). Then we tested diverse varieties from indica to japonica, Rayada to deepwater but observed merely Rayada varieties to develop shoot under such composite submerged stress. The interesting phenotypes of shoot development of Kaladigha in such composite submerged stress made us interested to check in other Rayada variety. Similar result was observed in another Rayada variety, Beto (Fig. 4.4). In addition to shoot development, Beto developed distinct and longer mesocotyl and coleoptile. Two other indica varieties including a Chinese variety YD6, showed partial shoot initiation capacity but cannot precede further develop further (Fig.4.7). Fig. 4.4 Exceptional submergence tolerance capacity of Rayada varieties under composite submerged stress. Representative image of several replicates taken 7 days after submergence. Composite stress (submergence plus oxidative stress) was imposed with 17 cm 10 mM H2O2 submergence. Details are in Materials and methods. 91 4.3.5 Coatless Rayada seeds unable to develop shoot under submergence As only Rayada varieties developed shoot after oxidative submerged stress, then we tried to identify how Rayada seeds were different than others. The only visible difference was seed coat color. Generally Rayada varieties have red seed coat or kernels whereas most others type varieties were white; however, some traditional varieties have also red kernel color. So we attempted to check whether the seed coat or pericarp color had any involvement on shoot development under such submergence as pigmentation is confined to the pericarp (Juliano and Bechtel 1985). We removed the seed coat manually and germinated under same condition where Rayada seeds developed shoot. Interestingly, coatless Rayada seeds were completely unable to develop shoot (Fig. 4.5a). To inquest the inability of shoot development of coatless seeds of Rayadas, we compared electric conductivity (EC) of intact to coatless seeds together with H2O2 treatment and observed a sharp and highly significant increase of electric conductivity in coatless seeds than control or H2O2 treatment (Fig. 4.5b) . H2O2 treatment also increased the electric conductivity but not significantly different from control seeds. We also compared the germination percentage of coatless seeds; coatless seed germinated faster than intact seed, even ABA inhibition was also observed significantly overruled (Fig. 4.5c), however, significant decreased of shoot growth was observed in coatless seed in aerobic condition. 92 Fig. 4.5 Effect of seed coat on germination and electric conductivity. a) suppression of shoot development under composite submerged stress of Kaladigha. b) electric conductivity of coatless seed with H2O2 treatment c) germination performance of coatless and intact seeds. Representative image of three replicates taken 5 days after submergence. 93 4.3.6 Riboflavin induces shoot development under submergence Vitamin B complexes are most abundant vitamin present in rice seed. Moreover, wild or red rice had been reported to have elevated vitamin B complexes in seed coat or bran. So we supplemented different vitamin B complexes along with H2O2 in submerged seed germination in YD6 (indica) variety that showed partial shoot initiation. Among different B complexes, surprisingly only vitamin B2 or riboflavin promoted shoot development under oxidative submergence (Fig. 4.6a). Apart vitamin B complexes, we also supplemented cytokinine, BAP that was widely used in shoot development in rice callus culture. However, BAP showed no augmenting effect on shoot development comparing to riboflavin effect (Fig. 4.6a). Supplementation of riboflavin and H2O2 successfully developed shoot in all varieties especially intolerant japonica type variety, Donjing (Fig. 4.8b) and Nipponbare. Both of the varieties were completely unable to develop shoot in submergence, even after 10 mM H2O2. We also checked whether lower combinational concentration or singly riboflavin can induce the shoot. We found neither high concentration of riboflavin (200 µM) singly nor in combination with lower concentration of H2O2 (100 µM) with high concentration of riboflavin (200 µM) able to induce shoot under submergence (Fig, 4.6b). So riboflavin and H2O2 acted synergistically with a concentration dependent manner in all varieties we tested whereas in Rayadas only H2O2 singly can efficiently develop shoot under submergence. 94 Fig. 4.6 Effect of vitamin-B complexes supplementation in shoot development under submergence, b) concentration dependency and synergistic effect of B2 and H2O2. Apart B complexes, a synthetic cytokinine, BAP was also supplemented Representative images were taken 9 days after submergence from three replicates of 100 seeds of YD6 varieties. 4.3.7 Nitric oxide: not involved in shoot development under submergence Nitric oxide is one of the signaling molecules, play important role in many biological processes such as disease resistance, abiotic stress, different growth and 95 development processes, including seed germination and dormancy release (Liu et al. 2010). NO has been reported as a germination enhancer in many plants. Moreover, under hypoxic condition significant amount NO produced in plants than normoxic condition, in addition nitrite can act as electron acceptor under O2 limiting environment. So we used NO donor, SNP either singly or in combination with H2O2 whether it has any effect on shoot development or can replace riboflavin instead. But we observed singly SNP did not show any promoting effect, rather reduced coleoptile growth (Fig. 4.7) but along with H2O2 showed weak synergistic effect, coleoptile rupture appeared, however no significant difference in YD6 variety as it can partially initiate shoot development under submergence under the same experimental conditions (Fig. 4.6a , only H2O2 ). Fig. 4.7 Effect of SNP either singly or in combination with H2O2 under submergence. Representative image of three replicates taken after 9 days after submergence. Each set was replicated thrice with 100 seeds of YD6. 4.3.8 Both exogenous ABA and glucose restrict shoot development under submergence Shoot growth is much more susceptible to ABA than that of root (Fig. 4.1) (Sharp and Davies 1989). Since glucose play important role in ABA signaling during seed germination, hence we examined the involvement of glucose signaling in 96 shoot development during seed germination by supplementing 3% glucose together with both riboflavin and H2O2 that efficiently induce shoot develop under submergence. Interestingly, complete inhibition of shoot development was observed after glucose supplementation (Fig. 4.8a). Similarly, complete inhibition of shoot development was also observed in ABA (1µM) (Fig. 4.8a). The concentration of 1µM ABA is not completely inhibitory of seed germination in aerobic condition during seed germination. Since osbzip84 accumulates high ABA than normal seeds, hence we examined whether it hiders shoot development after submergence in shoot inducing condition. Interestingly, we observed osbzip84 was unable to restrict shoot development in presence of both riboflavin and H2O2 (Fig 4.8b) under submergence, despite the elevated ABA content. However, developed seedlings were too weak to survive (data not shown), not as strong as normal seeds (Fig 4.8b). Fig. 4.8 Effect of a) glucose and ABA b) osbzip84 on shoot development under submergence. Control stands for shoot developing condition (10 mM H2O2+ 200 µM riboflavin) under submergence, Seeds of YD6 variety were used in glucose and ABA signaling effect, shown in (a) whereas osbzip84 background was Donjing seed, shown in (b). Representative image was 9 days after submergence. 97 4.3.9 Mitochondrial complex IV inhibition: restrict seed germination under submergence Sodium azide (NaN3) is a potent inhibitor of mitochondrial complex IV. We supplemented NaN3 (1mM) in shoot inducing condition (riboflavin and H2O2) under submergence whether the role of electron transport chain under submergence. Interestingly, we observed complete inhibition of seed germination in presence of 1mM NaN3 under submergence. However, the same concentration of sodium azide (NaN3) in aerobic condition neither inhibit seed germination nor shoot and root development (Fig. 4.9) although germinating time and seedling growth were significantly delayed (data not shown). Clearly, it means importance of mitochondrial complex IV or electron transport system even after submergence for germination as well as shoot development . Figure 4.9 Effect of mitochondrial complex IV inhibition on shoot development under submergence. A concentration of 1mM NaN3 was supplemented in aerobic germination whereas riboflavin and H2O2 was also supplemented under submergence. Representative image was taken 9 days after imbibition or submergence from 100 seeds of YD6 varieties of three biological replicates. 98 4.3.10 H2O2 affects carbohydrate metabolism under submergence As the sole source of energy of seed germination comes from stored material, mainly carbohydrate in rice seed, hence H2O2 and riboflavin might affect carbohydrate metabolism. So we analyzed the expression of several genes of αamylases in either coleoptile (or shoot) or caryopsis samples in Rayada variety, Kaladigha. After comparing gene expression, it clearly represented that H2O2 treatment in Rayada varieties mimicked the almost aerobic pattern of gene expression of Ramy1A and Ramy3D (Fig. 4.10). Ramy1A severely downregulated whereas Ramy3D up-regulated under submergence. But in H2O2 treatment, the only slight down expression was observed in Rmay1A whereas Ramy3D unchanged. However, when we compared same gene expression in samples excluding coleoptile or shoot, i.e. caryopsis only , then we observed significant up-regulation of Ramy3E after H2O2 although similar result of up expression was never been reported after submergence in rice. Fig. 4.10 Gene expression patterns of selected amylases in either coleoptile or caryopsis after submergence of Rayada. Dehulled seeds of Rayada (Kaladigha) were germinated under submerged condition (according to experimental design). RNA was isolated from either coleoptile or caryopsis samples 5 days after submergence. DNase treated RNA from respective samples were used for cDNA synthesis. Transcript levels were quantified by qRT-PCR, and compared with those of air germinated seeds as described in Materials and Methods. Values were means with ±SD (n=3). 99 4.4 Discussion Submergence tolerance during seed germination is one of the rare traits in rice, even among cereals. As regulator of submergence tolerance during seed germination is still unknown, hence we attempted with an alternative strategy to identify both positive and negative regulator(s). In this study, we for the first time, identified and developed a condition for efficient development of shoot, inducting the feature of Rayada rice. After successive experiments, we also identified several regulators of shoot development under submergence. 4.4.1 Rayada rice: is elixir of stress tolerances? Rayada is one of the special ecotypes with multiple stress tolerance capacity such as flood, submergence and cold. We observed Rayada variety, Kaladigha showed equal survival rate after 2 weeks of complete submergence comparing flash flood tolerant Sub1A rice variety , BRRI 52 (Fig. 2.5c,d) (data shown in chapter 2) . This unique feature of dual submergence (both partial and complete) tolerances made the variety special although the mechanism of complete submergence tolerance of Rayadas is still tentative. It is more likely that Sub1A type submergence tolerance mechanism also regulates complete submergence in Rayada rice too because of the presence of two Sub1A like alleles (OrSub1A-1, OrSub1A-2) in wild rice O. rufipogon (Li et al.2011). Rayadas are most primitive among deepwater rices and it still shares some feature of wild rice of O. rufipogon (Khush 1997). We also observed high oxidative stress tolerance in Rayada varieties (Fig. 4.2), mild salinity stress tolerance (Fig. 4.1, middle) and germination ability even after high concentration of ABA (Fig. 4.1, right). All these features lead us to be curious to check submergence tolerance during seed germination. But, our hypothesis were partially turned down after the screening. However, we observed successful shoot development in two Rayada varieties (Fig. 4.5) after oxidative stress imposed by 10 mM H2O2 in 17 cm submergence whereas similar condition did not induce shoot development in any other type rice varieties. Rayada varieties either might be more oxidative stress tolerant or use H2O2 as pronounced signaling molecules for growth and development 100 (pronounced oxidative signaling). Several features of deepwater rice or Rayadas such as lysigenous aerenchyma (Steffens et al. 2011), adventitious root formation (Steffens and Sauter 2009) are regulated by ROS signaling along with cell wall loosening for elongation after flooding, so it is more likely H2O2/ROS trigger shoot development under submergence. 4.4.2 Riboflavin richness of red rice Since coatless Rayada seeds are completely unable to develop shoot (Fig. 4.5a) after imposing oxidative stress during submergence but intact seed develop shoot in similar condition (Fig. 4.4). It clearly argues of presence of some components in seed coat that induce or essential for shoot development. After extensive data mining of seed coat materials, especially in red rice varieties, we found richness of vitamin-B complexes especially in red and wild rices; other vitamins are relatively poor. Among the B complexes , thiamine (B1), riboflavin (B2), niacin (B3), pantothenic acid (B5), and vitamin B6 are present in substantial amounts (Chen et al. 2012). After supplementing all of them separately (Fig. 4.6), we observed only riboflavin successfully induced shoot under submergence along with oxidative stress in all varieties that were completely unable to develop unless riboflavin supplement. Although we did not quantified the riboflavin content in Rayada and other modern varieties due to inadequacy of seeds of Rayada varieties, but, previous reports clearly mentioned significant richness of riboflavin in red rice than white grained modern varieties (Villareal and Juliano 1989) (http://ndb.nal.usda.gov/). We speculate all the tolerant varieties (Khao Hlan On, Khaiyan, Kalonchi, Nanhi and Cody) might have elevated riboflavin and ROS signaling along with red or black kernel color. At least two of them (Khao Hlan On and Khaiyan) probably possess red or black kernel or seed coat as seen in Fig 1.8b. However, we tried to collect such information from IRRI but failed to get their response. 101 4.4.3 Riboflavin: a signaling compound Riboflavin (vitamin B2) plays important role in plant normal growth and development as well as defense responses as flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD) are part of numerous flavoproteins. Riboflavin was reported as both biotic and abiotic signaling molecules activating stress tolerance (Dong and Beer 2000; Taheri and Tarighi 2010; Deng and Jin 2013). Systemic resistance against Peronospora parasitica and Pseudomonas syringae pv. tomato was observed in Arabidopsis after exogenous application of riboflavin (Dong and Beer 2000). Similarly, tomato plants showed systemic resistance to tobacco mosaic virus (TMV) and Alternaria alternate (Dong and Beer 2000) after riboflavin . Likewise, riboflavin acted as a defense activator in rice against economically important sheath blight diseased caused by Rhizoctonia solani (Taheri and Tarighi 2010). In a very recent report, Deng and Jin (2013) showed delayed leaf senescence and extended survival time conferring increased waterlogging tolerance after exogenous application of riboflavin in tobacco. Wild or purple/red rice is rich in riboflavin and most of the riboflavin resides in the seed coat (Villareal and Juliano 1989). So from all these previous reports along with our results clearly represent riboflavin as important signaling molecule. 4.4.4 How might riboflavin induce shoot development under submergence? We identified riboflavin as an inducer of shoot development in diverse type of varieties from indica to japonica under submergence in presence of 10mM H2O2. The major limitation under submergence is poor energy efficiency being limited O2 under submergence (hypoxia) or even absent (anoxia). Moreover, it is already evident that riboflavin promotes assembly and activity of mitochondrial complex IV (Grad and Lemire 2006). As oxygen binds to mitochondrial complex IV or cytochrome c oxidase, hence we checked whether respiratory inhibitor of mitochondrial complex IV (sodium azide) can inhibit the process of shoot development under submergence (Fig. 4.9). In aerobic condition, sodium azide delayed seed germination and seedling establishment but seeds successfully 102 developed shoot and root (Fig. 4.9) whereas completely restricted seed germination under submergence. However, apart mitochondrial complex IV, sodium azide can also inhibit mitochondrial ATPase, superoxide dismutase and DNA synthesis (Wood et al. 2012). Whatever, since H2O2 supplementation induced shoot development under submergence in Rayadas (Fig. 4.4), mimicked aerobic gene expression pattern of α-amylases (Fig. 4.10) whereas mitochondrial complex IV inhibition completely restricted seed germination (Fig. 4..9) , so it reasonably argue the importance of enhanced respiration for shoot development. In some way, H2O2 (in Rayadas) with riboflavin (in all other varieties) plausibly enhanced respiration. Lumichrome and lumiflavin are two derivative of the riboflavin formed under light. Moreover, Pseudomonas can also degrade riboflavin into lumichrome (Yanagita and Foster 1956). Lumichorome is already being reported as growth promoting compound produced by soil bacteria Sinorhizobium meliloti. Lumichrome enhanced 21% root respiration and increased net carbon assimilation in alfalfa (Volpin and Phillips 1998; Phillips et al. 1999). Similarly, lumichrome also enhanced root biomass in both Lotus japonicus and tomato. Apart legumes, lumichrome also stimulated seedling development in other cereals (Matiru and Dakora 2005; Gouws and Kossmann 2009). Surprisingly, exogenous application of lumichrome in both root (3 nM) and seed (5 nM) in alfalfa showed significant growth enhancement but it’s only in the shoot (Volpin and Phillips 1998; Phillips et al. 1999). So shoot development or enhancement feature of riboflavin or riboflavin derivative (lumichrome) is not only evident in rice but also in alfalfa. 4.4.5 Hidden player masked riboflavin Among cereals, rice has the least riboflavin content (weight basis) than others. The riboflavin content per 100g seeds of rice, wheat, maize, millet, sorghum, rye and oats was reported as 0.04, 0.10, 0.10, 0.33, 0.13, 0.25 and 0.14 mg, respectively (Juliano 1993). Then the question arises why not they develop shoot under submergence, if riboflavin induce shoot development. We also examined effect of riboflavin in other cereals (maize , wheat) but none of them developed 103 shoot under submergence (actually failed to germinate) even after riboflavin and H2O2 supplementation (data not shown) suggesting some hidden player(s) that might be triggered by riboflavin or riboflavin derivative lumichrome might be missing in these cereals whereas present in rice and alfalfa at least . Sub1A gene mainly responsible for submergence tolerance, similarly, mainly limited to some aus type varieties, none other type rice even other cereals lack such genes, hence submergence tolerance only limited to rice among cereals. 4.4.6 Involvement of ABA and glucose signaling ABA is considered as the stress hormone. Myriad of reports are available on ABA interacting functions in plants, mostly in water stress and seed dormancy (Nakashima and Yamaguchi-Shinozaki 2013). However, role of ABA in submerged seed germination is not evident. Generally ABA suppresses seed germination, subsequent seedling development. Shoot growth is more susceptible to ABA than root growth (Fig. 4.1) as previously reported (Sharp and Davies 1989). However, based on in silo analysis Mohanty et al. (2012) speculated ABA as a positive regulator of rice seed germination under submergence (Miro and Ismail 2013). Here we identified ABA as one of the regulators that inhibited shoot development both in aerobic (Fig. 4.1) and submerged condition (Fig. 4.8a), so undoubtedly ABA is the negative regulator of shoot development. Our results completely ruled out the speculation of Mohanty et al. (2012). We observed ABA content reduction after H2O2 treated seed in Rayada rice (Fig. 4.3c). Moreover, we also observed shoot development inhibition after exogenous glucose addition (Fig. 4.8a) suggesting involvement of glucose signaling in this process. Zhu et al. (2009) identified glucose induced delayed seed germination in rice due to suppression of ABA catabolism. However, one of the upstream regulatory mutant of ABA catabolism, osbzip84, (we identified using Rayada rice, described in Chapter 3) did not suppress shoot development at all. Glucose induced suppression of ABA8oxs was observed in OsABA8ox2 and OsABA8ox3, not in OsABA8ox1 (Zhu et al. 2009) and hence osbzip84 might be unable to restrict shoot development because osbzip84 acts as an upstream regulator of 104 OsABA8ox1. However, in osbzip84 developed shoot under submergence were too weak to establish seedling and eventually died. Crossing between tolerant line Khao Hlan On, with intolerant, IR64 and subsequent QTL analysis identified five QTLs defining 18-34% phenotypic variation in chromosomes 1, 3, 7, 9 (two QTL) (Angaji et al. 2010; Ismail et al. 2012). Similarly, another QTL study using population resulted from cross between Ma-Zhan Red (tolerant landrace) with intolerant IR42 identified six significant QTLs on chromosomes 2, 5, 6, and 7 (Septiningsih et al. 2013). These two QTL analysis share only limited region; so still a long way to run to identify the genetic basis of submergence tolerance. In summary, we successfully developed a condition supplemented with riboflavin and H2O2 where intolerant genotypes successfully developed shoot under submergence. However, induced shoot development was completely inhibited by glucose, ABA and mitochondrial complex IV inhibitor signifying ABA and glucose as negative regulators, whereas ROS, riboflavin and mitochondrial complex IV as positive regulators. Further research using our developed conditions, along with results may open up a new window to identify more regulators of submergence tolerance and enrich of our understanding of submergence tolerance mechanisms. 105 106 Chapter 5 A novel natural mutant of Rayada rice, plausibly in OsbHLH13, shows multiple phenotypes with potential use in hybrid seed technology 5.1 Introduction Extensive breeding efforts along with high-input agricultural practices doubled the rice productivity since first green revolution in 1960s. The second jump in rice productivity was observed after the development and successful commercialization of hybrid rice technology since it offers 15-20% yield advantage over best traditional varieties (Li and Zhang 2013). However, to meet up the demand of the increasing world population, global rice production needs to be again doubled by 2030 (Li and Zhang 2013). Development of super inbred and hybrid rice along with manipulation of inter-subspecific heterosis could offer the third leap (Li and Zhang 2013) although some constrains such as many unfilled grains, poor grain-filling, slow grain-filling rate etc. need to overcome first (Yang and Zhang 2010). 5.1.1 Stigma exsertion As rice is self-pollinated crop, hence the commercial production of hybrid seed depends on forced cross pollination through male sterile female plants to pollen donor male plants. However, asynchronous and differential timing of flowering hinder many spiklets to cross pollination (Yan et al. 2009). The feature of stigma exsertion in female lines, however, offers extended opportunity of cross pollination (Kato and Namai 1987; Yan et al. 2009). Most of the wild rice accessions of O. rufipogon possess high frequency of exserted stigma (Uga et al. 2003). Increasing frequency of stigma exsertion in female lines resulted increased hybrid seed setting and production (Zetian and Yanrong 2009). So the trait of stigma exsertion has received consistent attention from rice researchers as a desired feature, essential for hybrid seed production (Yan et al. 2009). However, the genetic basis of the trait is largely unknown. Most of the research identified 107 QTL markers, resides in all along 12 chromosomes of rice (Yamamoto et al. 2003; Uga et al. 2003, 2010; Miyata et al. 2007; Yan et al. 2009). 5.1.2 Morphological markers Morphological traits such as leaf, sheath, kernel color, awn length etc. are visible morphological markers are still being used by breeders for linkage map construction and convenient selection along with molecular markers (Yue et al. 2006). Since purity of hybrid seed possesses a great importance , so introgression of simple morphological markers into male sterile or restorer lines are very useful in identifying seed purity and have also shown great potential (Shen et al. 2004; Yue et al. 2006). Anthocyanins are pigments that are responsible for blue to red coloration of plants. Genetic basis of such coloration has been characterized in many plants including corn, petunia, and arabidopsis. 5.1.3 Trichome development Trichomes are hair like specialized structure developed from epidermal cells of leaf. Apart biological significances, trichomes are considered as an excellent model system for studying different aspect of cell differentiation (An et al. 2011). Trichomes have been reported with functions in both biotic and abiotic stress tolerances although protection against damage from herbivores most noticeable (Dalin et al. 2008). It can increase drought tolerance by reducing solar radiation and modulating moisture onto the leaf surface. Moreover, protection from UV radiation and cold temperature by profuse trichomes was also reported earlier (Dalin et al. 2008). 108 5.1.4 Anthocyanin and trichome development: interrelated processes Both anthocyanin pigmentation and trichome development have been extensively studied in Arabidopsis. Genetically, anthocyanin biosynthesis and trichome development are intimately connected. Recent studies have identified a ternary activator complex (MYB-bHLH-WDR) comprising of R2R3 family MYB transcription factor (TF), a basic helix-loop-helix (bHLH) TF and a WD40 protein , regulate the spatio-temporal expression of structural genes involved in anthocyanin biosynthesis and trichome development (Patra et al. 2013). In Arabidopsis, GLABROUS 1(GL1- an R2R3-MYB TF), GLABROUS 3 (GL3) and ENHANCER OF GLABROUS 3 (EGL3) (both are bHLH transcription factors) together with TRANSPARENT TESTA GLABRA 1 (TTG1, a WD40 protein) regulate the structural genes of anthocyanin biosynthesis and trichome development (Patra et al. 2013). We identified a novel natural mutant in Rayada variety, Kaladigha background having phenotypes of purple pigmentation throughout the plants and profuse trichomes in the leaves. Moreover, mutant plants showed high frequency of stigma exsertion. But the most important feature of the natural purple mutant was complete sterility. We first characterized the phenotypes and then attempted to identify the physiological basis of sterility and successfully recovered partially fertility. Based on phenotypic characterization, data mining, previous reports and expression analysis, we selected OsbHLH13 as the plausible candidate gene for these phenotypes. 109 5.2 Materials and methods 5.2.1 Plant materials and growth condition Seeds were germinated on moist filter papers in dark at 28°C. The natural purple mutant was identified after germination through distinct purple coleoptile and first leaf. Seedling of mutant along with control plants were raised on Kimura B nutrient solution (pH 5.6) in phytotron at 28°C, 60% relative humidity with 16/8hrs day-night cycle. Nutrient solution was renewed twice a week. After three weeks, seedlings of both mutant and control plants were transferred to the same soil pot and grown in greenhouse with 14/10 hrs day-night cycle. 5.2.2 Determination of total anthocyanin content Total anthocyanin content was determined using quick and simple spectrophotometric pH differential method (Rapisarda et al. 2000; Yuan et al. 2009). 100 mg of homogenized samples (young and mature leaf, leaf sheath and internodes) were extracted in 2 ml of pH 1.0 buffer containing 50 mM KCl and 150 mM HCl. Another set of similar samples were extracted in pH 4.5 buffer containing 400 mM sodium acetate and 240 mM HCl. Then the extracts were centrifuged at 12,000g for 15 min at 4oC. Absorbance of collected supernatants was measured at 510 nm. Total anthocyanin content was calculated using the following formula: Total anthocyanin content (mg g-1FW) = (Absorbance of pH 1.0 extractsabsorbance of pH 4.05 extracts) X 484.8/24 825 X dilution factor Where, 484.8 and 24,825 were the molecular mass of cyanidin-3-glucoside chloride and its molar absorptivity at 510 nm, respectively (Yuan et al. 2009). Triplicate samples were analyzed for each type of samples. 110 5.2.3 RNA extraction and real-time PCR analysis Total RNA was isolated from different sample (young leaves, leaf sheath and light exposed internode) using the Agilent Plant RNA isolation Mini Kit and then digested with RNase-free DNase I to eliminate genomic DNA contamination. First-strand cDNA was synthesized with oligo (dT) primers using a Promega cDNA kit accordingly. Transcript levels of each gene were measured by qRTPCR using Mx3000p QPCR System with iQ SYBR Green Supermix. PCR amplification was performed using three-step cycling conditions of 95°C for 10 s, followed by 40 cycles of 95°C for 10 s, 56°C for 30 s, and 72°C for 30 s. Actin1 was used as the internal standard. Primer sequences were listed below: OsbHLH13 F -5’ACTGTTGGCGTACAGCTGCAAA3’ OsbHLH13 R-5’CGCAACCAGAGTGTCGTGCGT3’ Actin1 F -5’ AGAGGAGCTGTTATCGCCGTCCTC3’ and Actin1 R -5’GCCGGTTGAAAACTTTGTCCACGC3’ 5.2.4 Estimation of trichome density and distribution Trichome density and distribution were examined under stereomicroscope with ocular grid and micrometer. Fully open mature leaves (usually 4rd leaf) were used for trichome distribution and density measurement. Trichomes were counted on both abaxial and adaxial surfaces and trichome density was measured on 10 leaves for both surfaces (three observations per leaf). 5.2.5 Determination of stigma exsertion Since stigma color of purple mutant plants was completely purple, hence it was easy to determine stigma exsertion percentage in purple mutant. A total 10 panicles of purple mutants were observed using magnifying glass to determine the percentage of stigma exsertion. However, exserted stigmas of control plants were observed under stereomicroscope as their stigmas were hyaline, difficult to 111 differentiate under light from lemma or palea color. The frequency of the double or single exserted stigma was calculated as the percentage (%) of exserted stigmas (either double or single) to the total spiklets of 10 panicles. 5.2.6 Pollen viability assay Pollen viability was measured using Alexander staining method (Alexander 1969). Stamen collected after anthesis and placed on slide. A few drops of Alexander stain buffer were used. Alexander stain buffer was made up with 95% ethanol- 10 ml; Malachite green (1% in 95% ethanol)-1 ml; Fuchsin acid (1% in water)-5 ml; Orange G (1% in water)-0.5 ml; phenol- 5 g; chloral hydrate-5 g; glacial acetic acid- 2 ml; glycerol- 25 ml and distilled water- 50 ml. Stained pollen grain was observed under stereomicroscope. Viable pollen stained red to redpurple whereas aborted pollen grain was green (Schoft et al. 2011). 5.2.7 Determination of stigma receptivity Stigma receptivity was determined by simple hydrogen peroxide (H2O2) method (Dafni and Maués 1998). A 6 % of hydrogen peroxide was placed on the stigma and observed production of bubbles from stigmas under stereomicroscope. Bubble production was compare under microscope where less bubble generation represented reduced stigma receptivity. 5.2.8 Humidification of panicles Since mutant plants were completely sterile, then we tried to recover fertility using different approaches, including humidification of panicles. Panicles including flag leaves were enclosed with transparent polybag. To ensure gas exchange, several tiny holes were made at the bottom of polybag. Humidity was measured using handheld hygrometer. 112 5.2.9 ROS staining and confocal laser scanning microscopy Total ROS staining was determined using fluorescent dye 2',7'- dichlorodihydrofluorescein diacetate (H2DCF-DA) (Invitrogen). Freshly dissected stigmas were washed gently with Hanks’ balanced salt solution (HBSS) buffer and stained with H2DCF-DA at 37 °C in the dark for 30 min, then washed briefly with the same buffer and observed under Olympus Fluoview FV1000 confocal laser scanning microscope with the following settings: excitation=488 nm, emission=535 nm, frame 1024×1024. To compare ROS staining between mutant and control stigmas, identical exposure conditions were used for all samples. Representative images of 10 stigmas were used for comparison. 113 5.3 Results 5.3.1 Phenotypic characterization of natural purple mutant Phenotypically a very distinct plant with multiple interesting phenotypes was isolated from Rayada variety, Kaladigha plants. Due to sharp and consistent difference with normal plants, we paid special attention to the plant considering as a natural mutant. The mutant plant was grown on same soil pot with control plants to nullify the environmental interactions on mutant phenotypes. The mutant plants were found completely sterile later on; hence tillers, nodal buds and branches were used to amplify the number of plants to characterize their special features. The four specific phenotypes were characterized in the mutant plants. 5.3.1.1 Purple pigmentation Rayada rice variety, Kaladigha is typically a green variety with occasional purple leaf sheath. Interestingly, main differential feature of the mutant plant was observed in the coloration of the plants. Initially, at seedling stage, mutant plant showed dense purple colored leaves (Fig. 5.1a), certainly distinct from normal plants. On maturation of leaves, deep purple color trended to fade. However, in abaxial side of mature leaves, it showed patches of purple pigmented spots, especially in regions of mid ribs and veins (Fig. 5.1b). To check the stem color, leaf sheath of old leaves were removed and observed pale purple color. However, in few days after exposed to light, stem color turned into dense purple (almost black) (Fig. 5.1c). Section of internodes also showed sharp difference in pigmentation throughout the tissues (Fig. 5.1d). The color of the spiklets was also changed. The color of the lemma and palea turned into light reddish, however, deep red at the tip (Fig. 5.1e). Inside the spiklets, interestingly, the stigmas of mutants plants were completely purple whereas in normal plants it was hyaline (Fig. 5.1f). Remarkably, roots at crown zone that were exposed to air and light also showed red pigmentation (Fig. 5.1g). In addition, lateral branches of these roots also showed changed pigmentation (Fig. 5.1h). Section of these roots clearly represented anthocyanin pigment accumulation only in the side of cortex that was exposed to light (Fig. 5.1i). Apart these, the leaf sheath (Fig. 5.1j) and ligule (Fig. 114 5.1k) also showed red to purple pigmentation. So the complete change in pigmentation throughout the plant organs (from root to stigma) was observed in the mutant plant organs. Fig. 5.1 Comparative pigmentation of purple mutant with control plants. a) young leaf, b) purple pigment patch in abaxial surface of leaf, c) internode pigmentation after light exposure, d) internode section , e) spiklets , f) stigmas , g) roots at crown zone , h) lateral braches of roots , i) section of roots , j) leaf sheath and k) ligule of purple mutant. In images with dual panels (a-f), first one represented control plants feature and the next one that of purple mutant. Representative images of at least 10 samples were shown here. 115 5.3.1.1.1 Anthocyanin content Total anthocyanin content was measured using the pH differential method (Rapisarda et al. 2000). Highly significant difference of anthocyanin content was observed between samples of control and purple mutant plants (Fig. 5.2). Light exposed internodes showed highest anthocyanin accumulation. Similarly, young leaves of seedlings also showed highly elevated anthocyanin content but significantly reduced in mature leaves, although still significantly higher than normal plants. Among different types of samples of control plants, only leaf sheath showed highest total anthocyanin content as Kaladigha variety occasionally developed purple leaf sheaths (Fig. 5.2). Other control samples such as young, mature leaves or internodes of normal or control plants showed low total anthocyanin content, range from 0.012 to 0.03 mg/g fresh weight. Fig. 5.2 Comparative total anthocyanin content in different parts of both control and purple mutants. Each type of samples was analyzed in triplicate and represented data were the mean of three replicates. Bars indicates the standard deviation whereas star (*) symbol indicates significance level at P< 0.01. 116 5.3.1.2 Profuse and elongated trichomes Development of profuse and elongated trichomes was observed as another altered feature in purple mutant plants. Normally Kaladigha variety possesses tiny leaf trichomes on its adaxial surface (Fig. 5.3a). However, in mutant plants profuse and elongated trichomes were observed on adaxial surface of leaves (Fig. 5.3b). Such elongated trichomes even could be seen in naked eyes against light (Fig. 5.3 c). Number of trichomes (per cm2) was significantly increased, mainly in adaxial surface (Fig. 5.3d). Moreover, in abaxial surface of leaves, elongated trichomes were also developed, but in lower extent. Length of the trichomes was around 700 µm in purple mutant plants, highly elongated than control plants (Fig. 5.3e, f). Although number of trichomes in abaxial surface was reduced but length of that was same as adaxial surface in purple mutant plants (Fig. 5.3e). 117 Fig. 5.3 Trichome density and distribution of control and purple mutant plants a) control, b) purple mutant leaf, c) naked eye view of control and purple mutant leaf, d) comparative trichome number, e) length of both adaxial and adaxial surfaces of both control and mutant plants , f) close up image of adaxial surface of leaf purple mutant plants . Representative images were from at least 10 different samples and the symbol (*) indicated the significance level at P<0.001. 118 5.3.1.3 Stigma exsertion Stigma exsertion was the third distinct phenotypes of the purple mutants (Fig. 5.4a). As the stigma color were changed to purple in mutant plants hence it was easy to notice even in naked eyes. Moreover, styles of purple pistil became bended and came out from lemma-palea (Fig. 5.4a-right panels). We observed almost 65 % of spiklets of mutant plants showed exserted stigmas. Among the exserted spiklets, double exserted stigmas were prominent in mutant, i.e. nearly 40%. However, the percentage of single exerted stigmas was also increased in mutant plants but not significantly different from normal plants (Fig. 5.4b). Fig. 5.4 Stigma exsertion of purple mutant. a) phenotype of exserted stigma, b) frequency of stigma exsertion between control and purple mutant spiklets. The symbol (*) indicated the significance level at P<0.001. 119 5.3.1.4 Sterility of purple mutant The last but most important feature of mutant plants was the complete sterility. A total of 24 individual plants was developed from a single mutant plant. All of them flowered after yearlong life cycle. However, none of them produced a single seed whereas almost 60% of the control plants spiklets produced seeds (Fig. 5.5). However, complete sterility made us more interested to the mutant plants. Fig. 5.5 Percentage of seed setting in a) control, b) purple and c) humidified purple mutant plants. Seed setting was calculated based on 10, 19 and 5 panicles of control, mutant, humidified purple mutant plants s. Bar represents the standard deviation. 5.3.2 Stigma receptivity: plausible cause of sterility As the mutant plants were found completely sterile, hence we attempted to reveal the possible physiological causes of sterility. To check the probable involvement of male or pollen based sterility, pollen viability was examined using Alexander staining (Alexander 1969). Pollen staining pattern clearly demonstrated that pollen were completely viable with normal shape and size, as control plants (Fig. 5.6b). Then we examined the ovary and observed no change in pigmentation. Color of ovary along with shape and size was similar to control plants (Fig. 5.6a). 120 Interestingly, after analyzing over 100 spiklets under stereomicroscope, we observed never a single pollen attached with stigmas of mutant plants (Fig. 5.6c), whereas plenty of pollens usually were attached with normal stigmas (Fig. 5.6d, arrows indicated the pollen grains). So we anticipated some problems might be associated with stigmas as its color were changed in mutants. Then we analyzed and compared receptivity of stigmas of mutant plants with control plants (Fig. 5.6 e, f). Surprisingly, we observed less receptivity of stigmas in mutant than control plants through analyzing bubble production after 6% H2O2. Fig. 5.6 Comparative reproductive features of mutant plants. a) ovary of mutant plant , b) pollen of purple mutants stained with Alexander stain, c) no pollen attachment in stigma of mutant plant, d) stigma with attached pollens in normal plants (arrows indicates the pollen) e) stigma receptivity test of purple mutant stigma and, f) of normal plants stigma . Representative image were of at least 10 replicates or samples. 5.3.3 Reduced ROS accumulation in mutant stigma The receptivity of stigma surface to pollen grain is only limited to short period and the timing for receptivity and pollination is critical for fertility or sterility. Receptivity of stigma is characterized by high level of peroxidase activity (McInnis et al. 2006) although the function of the enzymes in stigma is mostly unknown. Oppositely, antioxidant activity of anthocyanins was excessively reported. So we checked whether changed pigmentation in stigmas of mutant 121 plants had any involvement of ROS accumulation in stigmas. After ROS staining with H2DCF-DA, we compared the florescence signal of stigmas of mutants with that of control plants. It clearly demonstrated the significant reduction of ROS accumulation in stigmas of mutant plants than that of control plants (Fig. 5.7a). Similarly, in stigmatic papillae level, severe ROS reduction was observed in purple mutant (Fig. 5.7b). So both H2O2 bubbling assay (Fig.5.6 e, f) and H2DCFDA staining pattern of stigmas (Fig. 5.7a, b) clearly demonstrated reduced ROS accumulation in the mutant plants stigmas might be one of the reasons of sterility. 122 Fig. 5.7 ROS accumulation of a) whole stigma and b) stigmatic papillae level in both control and purple mutant plants. Freshly dissected stigmas were loaded with H2DCF-DA for 30 min in dark at 37 C and observed under confocal laser scanning microscope with similar image acquisition settings. Representative image of at least 10 stigmas were shown here. 123 5.3.4 Partial restoreation of fertility Since most of the mutant plants flowered in less humid season (late winter), stigmas of mutant plants were exserted (Fig. 5.4) and showed non-receptivity (Fig. 5.6e,f and 5.7a,b) , hence we artificially increased humidity of the panicles by wrapping the panicles with flag leaf by polybag as reduced humidity can reduce the receptivity of exserted stigmas. After enhanced humidity of 5 panicles we observed few seed setting occurred in each panicle but of very low percentage (Fig. 5.8). A total 30 seeds were obtained from these 5 panicle reflected partial restoration of fertility by increased humidity. Seed setting rate was increased to over 10% of spiklets after humidification whereas 60% of control spiklets developed into seed (Fig. 5.5). Although humidification partially restored fertility but other factors might be involved in the sterility phenomenon of the mutant plants as only limited percentage of spikelets overcome sterility after enhanced humidity. Moreover, bagging may also help pollen stick to the exserted stigma since wind or other physical interference on the pollen attachment is largely reduced. Fig. 5.8 Seed setting in purple mutant panicles after humidification. Untreated purple mutant panicle with complete sterility (upper panel), some seeds were developed (arrow marked) in humidified panicle bottom panel. Representative image of 5 humidified panicles with some seed settings were shown here. 124 5.3.5 Phenotype of purple mutant progeny We checked the progeny of the purple mutant seeds, developed from humidified panicles. Surprisingly, we observed seeds of the purple mutants were neither red (as normal) nor purple but it turned into completely black (Fig. 5.9a). Moreover, mutant plant seeds showed significantly reduced grain weight than control plant seeds (Fig. 5.9b). We germinated the seeds 7 days after harvest (DAH) and observed efficient germination but showed segregating nature of purple phenotype in a ratio of 3:1 although the number of seeds were very few to conclude. Fig. 5.9 Phenotype of purple mutant progeny. a) color of control and purple seed, b) seed weight , c) purple progeny. 5.3.6 Candidate gene of these phenotypes We extensively searched plausible candidate genes based on these four phenotypes and previous reported genes of similar phenotypes. No specific gene was reported with function of exserted stigma development. Most of the studies of exserted stigma are still in QTL level (Yamamoto et al. 2003; Uga et al. 2003, 2010; Miyata et al. 2007; Yan et al. 2009). Oppositely, plenty of genes dysfunction develops sterility phenotype. As the mutant plants possessed other two phenotypes (high anthocyanin and profuse and elongated trichome), hence we 125 focused on genes that control both of the feature. Interestingly, recent studies identified a ternary activator complex comprising members of R2R3-MYB, bHLH and WD40 proteins regulate both anthocyanin biosynthesis and trichome development (Patra et al. 2013). So we searched any of these protein members reported having involvement in sterility along with trichome and anthocyanin regulation. Fortunately, among numerous possibilities, we found only a single gene, Lc of maize that was reported earlier with all of these phenotypic features except exserted stigma, i.e, anthocyanin pigmentation, elongated trichome formation and sterility (Lloyd et al.1992; Tominaga-Wada et al. 2012; Li et al 2013). 5.3.7 Expression analysis of OsbHLH13 OsbHLH13 is the rice homolog of maize Lc gene. We analyzed the expression of OsbHLH13 using qRT-PCR between control and purple mutants using different types of samples. The expression analysis clearly showed exceptionally huge upexpression of OsbHLH13 in mutant plant samples (Fig. 5.10). In young leaves of developing tillers, the expression was up-regulated almost 150 fold than that of control plants. Similarly, significant up-expression of OsbHLH13 was observed in both leaf sheath and internodes samples but a lower extent than young leaves (Fig. 5.10). 126 Fig. 5.10 Relative expression OsbHLH13 in leaf, leaf sheath and internode of both control and mutant plants. DNase treated RNA from respective samples were used for cDNA synthesis. Transcript levels were quantified by qRT-PCR, as described in Materials and Methods. Values are means with ±SD (n=3). The symbol (*) indicates the significance level at P<0.0001 127 5.4 Discussion We identified a novel natural mutant of Rayada variety with Kaladigha background having distinct four phenotypes of the practical implications in hybrid seed technology. We also identified the receptivity of stigmas as probable reason of sterility in mutant plants. Based on phenotypic characterization, data mining of previous reports and expression analysis of putative gene, we selected OsbHLH13 as candidate gene for these mutant phenotypes. 5.4.1 Natural mutation, not seed contamination The first question raised after the mutant plant identification whether it was a natural mutant or seed contamination from purple variety since the probability of natural mutation resulting dominant phenotype is low (Brooks et al. 2008). As the seeds of the Kaladigha variety were collected from gene bank of Bangladesh Rice Research Institute (BRRI) and generally gene bank collections are good at purity and integrity, so the chance of contamination is very low or absent. However, to ascertain the same varietal origin, the mutant plant was grown with normal Kaladigha plants (as control) in same soil pot to compare the features of Rayada variety, Kaladigha. Except four noticed phenotypes, all other features such as shape and size of young (Fig. 5.1a) and mature leaves (Fig. 5.3a, b), stem (Fig. 5.1c, d), ligule, spiklets (Fig. 5.1e), and overall plant stature were same as control plants of Kaladigha. We also examined the seed dormancy of the mutant seeds, developed after enhanced humidity treatment and we observed the freshly harvested seeds (7 DAH) germinated efficiently same as control plants (Fig. 5.9c). Rayada varieties specially lack seed dormancy while all deepwater rice varieties even wild rice (O rufipogon) possessed significantly prolonged seed dormancy. The others features of Rayadas like yearlong life cycle, strong photosensitivity were also present in the mutant plants. Moreover, seeds of the mutant plants showed segregating phenotypes for purple pigmentation. All these features clearly ruled out the possibility of seed contamination and signified the mutant as Rayada variety, Kaladigha background. Brooks et al. (2008) identified a natural revert mutation in RC from 56 tons of foundation seeds of rice. 128 5.4.2 Sterility and reduced ROS accumulation of stigmas: linked together Stigma is the one of specific regions of plant reproductive organs that directs the success of the progeny development as pollen first attach to it, gets initial nutrient and guidance for pollen germination and tube growth (Li et al. 2007). Cyanidin-3glucoside is the most common anthocyanin in purple pigmentation with high antioxidant activity, especially superoxide and hydroxyl radical scavenging activity (Ichikawa et al. 2001). Stigmatic ROS / H2O2 were shown to be involved stigmatic receptivity, pollen germination and pollen tube growth (McInnis et al. 2006). Similarly, in kiwifruit ROS was found to involve in pollen tube initiation (Speranza et al. 2012). Moreover, stigmatic ROS/H2O2 is also involved in biotic resistance of stigma although how stigma differential pollen to pathogen, especially fungal spore is still unknown. Angiosperms possess large numbers of class III peroxidases; same in rice i.e. a total 138 class III peroxidase genes (Passardi et al. 2004). Mature and receptive stigmas of angiosperms also possess high level of peroxidase activity. McInnis et al. (2006) identified a stigma specific peroxidase from Senecio squalidus. Significantly, large amount of ROS, specifically H2O2 was detected in stigma where stigma papillae specific peroxidases expressed in S. squalidus (McInnis et al. 2006). However, exact function of these peroxidases is unknown. Although membrane bound NADPH oxidases are major sources of H2O2 production, however, some peroxides is also shown in the production of H2O2 (McInnis et al. 2006). As the purple mutants possessed deep purple stigmas and showed complete sterility, then we attempted to check whether the changed stigma pigmentation had any involved in this sterility. We compared the H2O2 production level in the stigmas both in control and purple mutant by ROS staining using fluorescent dye H2DCF-DA. The ROS staining pattern clearly represented the severe reduced ROS accumulation in stigma of purple mutant (Fig. 5.7a,b). Thus the reduced ROS level in purple mutant stigmas might be due to the high anthocyanin level or scavenging activity that might limit both receptivity of stigma and germination of pollen. 129 5.4.3 Humidity and sterility The impact of temperature and humidity on the sterility of rice was studied earlier (Weerakoon et al. 2008). Desiccation of stigma, loss of pollen viability and reduction of pollen germination on stigma might be the reason of increasing sterility after low relative humidity as observed in rice whereas high humidity significantly prolonged the stigma receptivity duration (Weerakoon et al. 2008). Simultaneously, high humidity also reduced the pollen dehiscence capacity from anther, might be another limiting factor of restoring complete fertility after humidification of purple mutant. In Arabidopsis, mutation of a family 8 glycosyltransferase gene resulted humidity sensitive sterility that can be partially rescued by high humidity (Lao et al. 2003) however, this gene was neither involved in anthocyanin pigmentation nor in trichome development, even in stigma exsertion 5.4.4 Black colored but reduced grain weight Humidified panicles developed some seeds with complete changed (dark purple or black) pigmentation (Fig.5.9a). Moreover, it showed significant reduction of grain weight (Fig.5.9b). So it came into consideration whether huge anthocyanin accumulation (since black grain) resulted reduced grain weight. Interestingly, in nature large fruits with dark or black color are severely scarce (Steyn 2009). Moreover, in some wild berries, huge anthocyanin accumulates but their fruit size is relatively small, along with thin fruit skin (Moyer et al. 2002; Steyn 2009). Moreover, treatments that induce anthocyanin biosynthesis also reduce cell growth rate in cell culture (Steyn 2009). So in culture system gain of anthocyanin production is offset by reduction of growth rate (Steyn 2009). Thus it seems the intense pigmentation in mutant seeds may be linked with reduced grain weight. 130 5.4.5 Selection of OsbHLH13 as candidate gene We selected OsbHLH13 as plausible candidate gene of the mutant, mostly based on phenotypic features and intense literature mining of relevant features. Recent studies identified a ternary activator complex (MYB-bHLH-WD40 complex) regulates both anthocyanin biosynthesis and trichome development (Patra et al. 2013). Since mutant plant showed both high anthocyanin along with prolonged & profuse trichomes, hence we assumed that the mutant plants might have mutation among the members of these regulatory ternary complexes. Structural features of bHLHs members involved in these regulatory processes suggest that it (bHLHs) might act as docking proteins among them. Because its Nterminal regions interact with MYB proteins, negatively charged successive 200 amino acid may interact with either pol III or WD40 complex and finally C terminal HLH region participate in either homo or hetero dimerization (Fig. 1.6) (Hichri et al. 2011) . Considering these structural features, we then emphasized on bHLHs. Among bHLHs of Arabidopsis, maize, petunia and Antirrhinum , only maize Lc was reported with features of trichomes , root hair and anthocyanin pigmentation in both leaf and seed coat (Ramsay and Glover 2005). Oppositely, no gene among MYBs was reported having all three functions all together whereas among WD40s, TTG1 of arabidopsis (Walker et al. 1999) and PAC1 of maize (Carey et al. 2004) were reported with all three functions. Interestingly, maize PAC1 can complement Arabidopsis ttg1 mutants (Carey et al. 2004). However, maize pac1 mutants only show a reduction in anthocyanin pigmentation in specific tissues (Carey et al.2004). In addition, over-expression of maize Lc in Arabidopsis ttg1 mutant background complements the phenotypes (Ramsay and Glover 2005). So it indicates mutation in WD40 genes (especially in TTG1 or PAC1 type) is more unlikely regulating all these phenotypes. Li et al. (2007) identified 548 genes using both 57 K Affymetrix rice wholegenome array and 10 K rice cDNA microarray in japonica rice variety, Nipponbare that were either stigma specific or predominantly expressed in stigmas of rice. Interestingly, they found OsbHLH13 (maize Lc homolog in rice) 131 as one of the highly expressed probes in stigma although stigma of Nipponbare was hyaline or pale green (Li et al. 2007). Since the mutant plants showed both purple and exserted stigmas hence OsbHLH13 might be the more likely candidate gene of these phenotypes. Moreover, as purple pigmentation was observed throughout the mutant plants organs, hence we searched genes that can change pigmentation in diverse organs, not only in leaf or seed coat. In transgenic petunia expressing maize Lc resulted deep purple foliage after up-regulation of at least 8 biosynthesis genes (Bradley et al. 1998), whereas when over-expressed in Arabidopsis resulted elevated number of trichomes along with high level anthocyanins (Lloyd et al. 1992). In addition, in an a very recent report of Li et al (2013) showed the sterility regulating feature of Lc in seventeen independent transgenic rice plants expressing maize Lc gene under control of the CaMV 35S promoter. Surprisingly, we also observed huge up-expression of OsbHLH13 in purple mutant (Fig. 5.10). Thus the candidate gene search narrowed to only maize Lc homolog in rice although mutation in other upstream regulatory genes could be still possible. In summary, we isolated a novel mutant of Rayada variety with Kaladigha background having multiple interesting phenotypes and identified the possible reason of the sterility of the mutant was found being related to the stigma receptivity. OsbHLH13 was identified as the candidate gene of these phenotypes. Further studies of genetic characterization either through genetic mapping or functional cloning of the candidate gene may open up the practical implications of this mesmerizing mutant. 132 Chapter 6 General Discussion and Conclusion Natural selection or evolution tends to increase plant adaptability through genetic diversity, whereas crop domestication and subsequent breeding or directional selection narrow the genetic diversities of crop plants (Alonso-Blanco et al. 2009); even promoted gathering of deleterious mutations in their stress response mechanisms (Tang et al. 2010). Thus natural variation is the assets of crop improvement program. Generally wild species are rich in genetic diversity. However, its direct use in breeding program is limited due to different incompatibilities as well as its inferior yield or quality. Landraces or ecotypes that are still growing in native preferences do share some feature of wild species. They could offer more opportunities for breeders as demand for more genetic resources increases. In some areas of Bangladesh where typical deepwater rice cannot be cultivated due to high depth of flooding, Rayadas are the only option to be cultivated (Perez and Nasiruddin 1974; Catling 1992). However, due to the invasion of high yielding modern rice varieties, traditional rice varieties tend to disappear rapidly in their native ecosystems. No improvement program for deepwater rice varieties exists although it was initiated as early as in 1917 (Zaman 1977). Moreover, even IRRI has left deepwater rice variety improvement program out of its priority list. Bangladesh Rice Research Institute first drew the attention of the scientific community through its first deepwater rice conference in 1974. Then in successive years, several countries like India, Thailand etc. also expanded and collaborated in the deepwater rice research including IRRI. However, in 1990s research of deepwater rice lost its pace although some research groups around the globe have identified several important molecular insights of deepwater rice such as snorkel genes (Hattori et al. 2009). Being poor yield performance and yearlong life cycle, Rayada has always been neglected. Moreover, longest life cycle might be a reason for such long duration varietal disappearance (Vergara and Chang 1985). Some of the features of Rayadas were reported almost 40-50 years ago. Still the same data are used by others such as Rayada growing area. No update information is available whether such types of varieties are still being grown in the field or already been eliminated 133 by high yielding varietal invasion. However, after reviewing and analyzing their physiological specialty, we are convinced with their special features that could be certainly used as excellent resources for future crop improvement program as well as basic scientific studies. For instance, we observed its significantly prompt recovery from water stress (Fig. 2.9). Despite being big stature of Rayada, after water stress, it possesses excellent and rapid recovery capacity. Moreover, longer root system of Rayada is being already reported (Fig.2.8a) (Gowda et al. 2012). Similar patterns of the longer root system are also observed in well-watered conditions in our study (Fig.2.8b). In addition, prompt stomata response through leaf rolling that we observed after transferring high humidity growing Rayada to low humidity condition suggests efficient water sensing mechanism present in Rayada rice. Despite complete and partial submergence tolerance mechanism are different (Xu et al. 2006; Hattori et al. 2009), we observed somehow Rayada variety withstand two weeks complete submergence with similar recovery performance to Sub1A rice (Fig. 2.5c, d) along with its natural ability to elongate after partial flooding (Fig. 2.4d). So the dual submergence tolerance capacity of Rayada rice makes it not only special but also unique. Similar feature of dual tolerance capacity was reported some 40 years ago through farmers but still unexplored scientifically (Perez and Nasiruddin 1974). Initially it was reported that wild rice does not possess Sub1A type allele but recent studies identified Sub1A like two alleles (OrSub1A-1, OrSub1A-2) in O. rufipogon (Li et al. 2011). Hence being most primitive among deepwater rices (Catling 1992), Rayada still shares some features with wild rice (Khush 1997), thus it is most likely that Rayada rice also possesses Sub1A type pathway for complete submergence tolerance. It would be fascinating to identify how do Rayadas sense and activate signaling pathways after such stress. Moreover, longer root system, faster recovery and quicker stomata response are considered traits of drought tolerance or adaptation; interestingly, all of these three features in some extent are present in some varieties of Rayadas such as Kaladigha. So considering all of these features altogether we expect Rayada rice will become a new focus of rice germplasm research with the rapid advances in genome research weaponry. 134 In this study, we for the first time examined Rayadas as natural variation to identify both physiological and genetical bases of one of their special features, the lack of dormancy that is different from other deepwater rice type (Table 1). Based on our results it shows several concerted events starting with less ABA in freshly harvested seeds (Fig.3.1b) enable to accumulate huge ROS after imbibition (Fig. 3.2) as well as faster ABA catabolism through ABA8ox1 (Fig. 3.9) might enable Rayada seeds to germinate even after freshly harvest. Similar results of high ROS accumulation in non-dormant seeds of Arabidopsis was reported earlier (Leymarie et al. 2012). Since in osbzip84 T-DNA insertion was in promoter region, hence ABA accumulation in seed was not that much higher. In Arabidopsis triple mutant, cyp707a1a2a3 showed strong dormancy phenotype with 70 fold higher ABA whereas osbzip84 showed nearly 2 fold only. Interestingly, it was reported earlier that the expression of ABA8oxs are up-regulated after water stress (Kushiro et al. 2004), moreover we also observed the same (Fig. 3.8b) but osbzip84 showed reduced ABA content both in water stress (Fig. 3.12e), normal condition (Fig. 3.12d). So reduced ABA content in both water stress and normal conditions suggest that OsbZIP84 might not only regulate ABA catabolism but also regulate ABA homeostasis. Certainly, further study is necessary to confirm the exact role in ABA homeostasis. ABA is one of the key phytohormones of plants; regulating many of growth and developmental processes including abiotic stress tolerance, seed maturation, dormancy acquisition and germination. Enormous research on ABA perception, signaling, transport and function have been done since the discovery of the compound in the 1960s although the naming of the compound as abscicis acid is a misnomer, initially thought as a regulator of abscission, however, not involved directly in that process, rather an ethylene regulated process (Schwartz and Zeevaart 2010). Usually acting in the downstream of ABA signaling, ROS is becoming a key signaling regulator of diverse types of plants growth and development pathways. It has become increasingly evident of ROS accumulation and dormancy release. H2O2 both enhances seed germination (Fig 4.3 a) both in aerobic and submerged conditions as well as alleviates seed dormancy in many plants. Moreover, hydrogen cyanide or methyl viologen also alleviate seed dormancy (Oracz et al. 135 2007). Apart dormancy, ROS also plays important function in root hair development. (Foreman et al. 2003), pollen tube growth (Potocky et al.2007), leaf expansion (Rodriguez et al. 2002), PCD (Breusegum et al. 2008). In our studies, we observed ROS is necessarily important to Rayada rice too for their growth and development. Apart ROS accumulation of freshly harvested seeds, we also observed ROS accumulation also plays a defining role in fertility (Fig. 5.7) as we observed in purple mutant with reduced ROS accumulation and sterility. Similarly, Rayada rice also developed shoot under submergence in the presence of H2O2 (Fig. 4.5). Previous reports establish the essentiality of PCD of adventitious root development (Steffens and Sauter 2009) as well as aerenchyma formation (Steffens et al. 2011). Suppression of shoot development by ABA and glucose might be mediated by restriction of ROS accumulation. Since coatless Rayada seeds were completely unable to develop shoot (Fig. 4.5a), whereas intact (coated) seed germinated with shoot in the presence of H2O2, inferring key regulatory factor reside in kernel or seed coat. After supplementing different vitamin B complexes we identified riboflavin as the key shoot development inducing molecule. A good number of reports are available on biotic resistant signaling by riboflavin (Dong and Beer 2000; Taheri and Tarighi 2010; Deng and Jin 2013). Similarly, riboflavin derivative, lumichrome is a natural growth promoting compound released by soil bacteria Sinorhizobium meliloti. Moreover, exogenous lumichrome application enhances root respiration as well as shoots growth in alfalfa. Since the H2O2 and riboflavin supplemented condition develop shoot in intolerant varieties, so it is the time to identify both positive and negative regulators of this process. Moreover, if lumichrome supplementation can develop the same under submergence then the involvement of riboflavin will be confirmed. Finally, we isolated an eye-catching mutant with interesting four phenotypes on Rayada variety, Kaladigha background. After phenotypic characterization, we identified the biggest hurdle of the mutant is its complete sterility. We tried to identify possible reason of such sterility. After stigma receptivity test (Fig. 5.6 e, f) and fluorescent H2DCF-DA staining (Fig. 5.7) we have largely identified the reason of sterility as non-receptivity of stigmas. Interestingly, humidifying the panicle partially overcomes sterility. Based on phenotypic characterization and 136 previous reports along with gene expression of OSbHLH13 we have identified the maize Lc gene as the candidate gene of this mutant. The gene expression of OsbHLH13 is extremely up-regulated in the purple mutant. Further studies of genetic characterization may open up the practical implications of this mesmerizing mutant. In summary, Rayada is such primitive deepwater rice that can offer many traits and genetic resources that are badly needed in rice breeding for stress tolerance. More detailed research is needed to characterize its special features and mine its useful genes. We think the time is mature to do so with the rapid progress in genome research. 137 138 References Achard P, Renou JP, Berthome R, Harberd NP, Genschik P (2008) Plant DELLAs restrain growth and promote survival of adversity by reducing the levels of reactive oxygen species. Current Biology 18: 656-660 Aguan K, Sugawara K, Suzuki N, Kusano T (1993) Low-temperature dependent expression of a rice gene encoding a protein with a leucine zipper motif. Molecular and General Genetics 240: 1-8 Ahmed AU , Mirza MMQ (2000) Review of causes and Dimensions of Floods with Particular Reference to Flood ’98: National Perspectives. In: Ahmed QK, Chowdhury AKA, Imam SH, Sarker M (eds) Perspectives on Flood 1998, University Press Limited, Dhaka Ahmed MS (1974) Ecology, distribution and production of deep-water rice. In: Proceedings of the International Seminar on Deepwater Rice, August 21-26, 1974. No. 15. Bangladesh Rice Research Institute, 1975, pp 1-9 Albert NW, Lewis DH, Zhang H, Irving LJ, Jameson PE, Davies KM (2009) Light-induced vegetative anthocyanin pigmentation in Petunia. Journal of experimental botany 60: 2191-2202 Alexander MP (1969) Differential staining of aborted and non-aborted pollen. Stain Technol 44:117-122 Alonso-Blanco C, Aarts MGM, Bentsink L, Keurentjes JJB, Reymond M, Vreugdenhil D, Koornneef M (2009) What has natural variation taught us about plant development, physiology, and adaptation? Plant cell 21:18771896 Amoutzias GD, Veron AS , Weiner J , Robinson-Rechavi M , Bornberg-Bauer E , Oliver SG , Robertson, DL(2007) One billion years of bZIP transcription factor evolution: conservation and change in dimerization and DNA-binding site specificity. Molecular biology and evolution 24:27-835. An L, Zhou Z, Yan A, Gan Y (2011) Progress on trichome development regulated by phytohormone signaling. Plant signaling & behavior 6: 1959-1962 139 Andaya V, Mackill DJ (2003) QTLs conferring cold tolerance at the booting stage of rice using recombinant inbred lines from a japonica × indica cross. Theoretical and applied genetics 106:1084-1090 Angaji SA, Septiningsih EM, Mackill DJ, Ismail AM (2010) QTLs associated with tolerance of flooding during germination in rice (Oryza sativa L.). Euphytica 172:159-168 Ashraf M (2010) Inducing drought tolerance in plants: Recent advances. Biotechnology advances 28: 169-183. Bailey-Serres J, Fukao T, Ronald P, Ismail AM, Heuer S, Mackill D (2010) Submergence tolerant rice: SUB1’s journey from landrace to modern cultivar. Rice 3: 138-147 BalasubramanianV, Hill JE (2002) Direct seeding of rice in Asia: emerging issues and strategic research needs for the 21st century. In: Pandey S, Mortimer M, Wade L, Tuong TP, Lopez K, Hardy B, eds. Direct seeding: research strategies and opportunities, Philippines: International Rice Research Institute, Los Baños Bashar KM, Haque MM, Zaman HMS (2004) Rice biodiversity and genetic wealth of flood-prone environment of Bangladesh. In: Bhuiyan SI, Abedin MZ, Singh VP, Hardy B, (eds) Rice research and development in the floodprone ecosystem. Proceedings of the international workshop on flood-prone rice systems held in Gazipur, Bangladesh,9-11 January 2001, International Rice Research, Los Baños Beachell (1974) Characteristics for consideration in deepwater breeding programme. In: Proceedings of the International Seminar on Deepwater Rice, August 21-26, 1974. No. 15. Bangladesh Rice Research Institute, 1975, pp 92-97 Benschop J, Jackson M (2005) Contrasting interactions between ethylene and abscisic acid in Rumex species differing in submergence tolerance. The plant journal 44: 756-68 140 Bentsink L, Jowett J, Hanhart CJ, Koornneef M (2006) Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proceedings of the national academy of sciences, 103: 17042-17047 Bewley JD, Bradford K, Hilhorst HW (2013) Seeds: physiology of development, germination and dormancy. Springer Bhuiyan SI, Abedin M.Z, Singh VP,Hardy B (2004) Rice Research and Development in the Flood-Prone Ecosystem. IRRI Blum A (2011) Drought resistance -is it really a complex trait? Functional plant biology 38: 753-757 Bouman BAM, Humphreys E, Tuong TP, Barker R (2007) Rice and water. Advances in agronomy 92:187-237 Bovy A, de Vos R, Kemper M, et al. (2002) High-flavonol tomatoes resulting from the heterologous expression of the maize transcription factor genes LC and C1. The plant cell 14:2509-2526 Bradley JM, Davies KM, Deroles SC, Bloor SJ, Lewis DH (1998) The maize Lc regulatory gene up-regulates the flavonoid biosynthetic pathway of Petunia. The plant journal 13:381-392 Breusegum FV, Bailey-Serres J, Mittler R (2008) Unraveling the tapestry of networks involving reactive oxygen species in plants. Plant physiology 147:978-984 Brooks SA, Yan W, Jackson AK, Deren CW (2008) A natural mutation in rc reverts white-rice-pericarp to red and results in a new, dominant, wild-type allele: Rc-g. Theoretical and applied genetics 117: 575-580 Cai HW, Morishima H (2000) Diversity of rice varieties and cropping system in Bangladesh deepwater areas. Japan agricultural research quarterly 34:225-231 Caliskan M, Cuming AC (1998) Spatial specificity of H2O2 -generating oxalate oxidase gene expression during wheat embryo germination. Plant journal 15: 165-171 141 Carey CC, Strahle JT, Selinger DA, Chandler VL (2004) Mutations in the pale aleurone color1 regulatory gene of the Zea mays anthocyanin pathway have distinct phenotypes relative to the functionally similar TRANSPARENT TESTA GLABRA1 gene in Arabidopsis thaliana. Plant cell 16:450-464 Catling HD (1992) Rice in Deepwater. MacMillan Press, London. Catling HD, Puckridge DW, Hille Ris Lambers D (1988) The environment of Asian deepwater rice. In: Proceedings of the 1987 International Deepwater Rice Workshop. Manila, International Rice Research Institute, Manila Chang TT, Bardenas EA (1965) The morphology and varietal characteristics of the rice plant. IRRI Tech Bull 4, p 40 Chen H, Chen W, Zhou J, He H, Chen L, Chen H, Deng XW (2012) Basic leucine zipper transcription factor OsbZIP16 positively regulates drought resistance in rice. Plant science 193:8-17 Chen W, Provart NJ, Glazebrook J, Katagiri F, Chang HS, Eulgern T, Mauch F, Luan S, Zou G, Whitham SA, et al (2002) Expression profilematrix of Arabidopsis transcription factor genes suggests their putative functions in response to environmental stresses. Plant cell 14:559-574 Chen Y, Wang M, Ouwerkerk PB (2012) Molecular and environmental factors determining grain quality in rice. Food and energy security 1:111-132 Chinnusamy V, Schumaker K, Zhu JK (2004) Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. Journal of experimental botany 55: 225-36 Chinnusamy V, Zhu JK (2009) Epigenetic regulation of stress responses in plants. Current opinion in plant biology 12:133-139 Chono M, Honda I, Shinoda S, Kushiro T, Kamiya Y, Nambara E, Kawakami N, Kaneko S, Watanabe Y (2006) Field studies on the regulation of abscisic acid content and germinability during grain development of barley: molecular and chemical analysis of pre-harvest sprouting. Journal of experimental botany 57: 2421-2434 142 Clark LG, Zhang W, Wendel JF (1995) A phylogeny of the grass family (Poaceae) based on ndhF gene sequence data. Systematic botany 20:436-460 Dafni A , Maués MM (1998) A rapid and simple procedure to determine stigma receptivity. Sexual Plant Reproduction 11:177-180 Dai S, Petruccelli S, Ordiz MI, Zhang Z, Chen S, Beachy RN (2003) Functional analysis of RF2a, a rice transcription factor. Journal of biological chemistry 278: 36396-36402 Dai S, Zhang Z, Chen S, Beachy RN (2004) RF2b, a rice bZIP transcription activator, interacts with RF2a and is involved in symptom development of rice tungro disease. Proceedings of the national academy of sciences of USA 101: 687-692 Dalin P, Ågren J, Björkman C, Huttunen P, Kärkkäinen K (2008) Leaf trichome formation and plant resistance to herbivory. In Induced plant resistance to herbivory. Springer Netherlands pp. 89-105 Dasgupta S, Huq M, Khan ZH, Masud MS, Ahmed MMZ, Mukherjee N, Pandey K (2011) Climate Proofing Infrastructure in Bangladesh: The Incremental Cost of Limiting Future Flood Damage. The journal of environment & development 20:167-190 Datta SD, Malabuyoc JA, Aragon EL (1982) Evaluation of rices for drought tolerance for deepwater areas. In: International Deepwater Rice Workshop, Bangkok (Thailand), 2-6 Nov 1981, International Rice Research, Los Baños Deng B, Jin X (2013) Riboflavin spraying impairs the antioxidant defense system but induces waterlogging tolerance in tobacco. Acta Physiologiae Plantarum 35:2769-2776 Doebley J, Lukens L (1998) Transcriptional regulators and the evolution of plant form. Plant cell 10: 1075-1082 Doebley JF, Gaut BS, Smith BD (2006) The molecular genetics of crop domestication. Cell 127: 1309-1301 143 Dong H, Beer SV (2000) Riboflavin induces disease resistance in plants by activating a novel signal transduction pathway. Phytopathology 90:801-811 El-Maarouf-Bouteau H , Bailly C (2008) Oxidative signaling in seed germination and dormancy. Plant signaling & behavior 3: 175-182 Finch-Savage WE, Leubner-Metzger G (2006) Seed dormancy and the control of germination. New phytologist 171: 501-523 Finkelstein R, Reeves W, Ariizumi T, Steber C (2008) Molecular aspects of seed dormancy. Annual review plant biology 59: 387-415 Foreman J , Demidchik V , Bothwell, JH , Mylona, P , Miedema H , Torres MA , Linstead, P , Costa S , Brownlee C , Jones JD , Davies, JM , and Dolan L (2003) . Reactive oxygen species produced by NADPH oxidase regulate Plant cell growth . Nature 422:442 - 446 Foyer CH, Noctor G (2005) Oxidant and antioxidant signalling in plants: a reevaluation of the concept of oxidative stress in a physiological context. Plant, cell & environment 28: 1056-1071 Fukao T, Bailey-Serres J (2008) Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proceedings of the national academy of sciences 105: 16814-16819 Fukao T, Yeung E, Bailey-Serres J (2011) The submergence tolerance regulator SUB1A mediates crosstalk between submergence and drought tolerance in rice. The plant cell 23: 412-427 Fukazawa J, Sakai T, Ishida S, Yamaguchi I, Kamiya Y, Takahashi Y (2000) Repression of shoot growth, a bZIP transcriptional activator, regulates cell elongation by controlling the level of gibberellins. The plant cell 12: 901-15 Glaszmann JC (1987) Isozymes and classification of Asian rice varieties. Theoretical and applied genetics 74: 21-30 Gomosta R, Vergara BS (1983) Photoperiod sensitivity of Rayada rices. IRRN 8:6 144 Gouws LM, Kossmann J (2009) The molecular analysis of the effects of lumichrome as a plant growth promoting substance. Doctoral dissertation, Stellenbosch: University Stellenbosch Gowda VR, Henry A, Vadez V, Shashidhar HE, Serraj R (2012) Water uptake dynamics under progressive drought stress in diverse accessions of the OryzaSNP panel of rice (Oryza sativa). Functional plant biology 39:402-411 Grad LI, Lemire BD (2006) Riboflavin enhances the assembly of mitochondrial cytochrome c oxidase in C. elegans NADH-ubiquinone oxidoreductase mutants. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 1757:115-122 Graeber K, Nakabayashi K, Miatton E, Leubner‐Metzger G Erhard Soppe WJ (2012) Molecular mechanisms of seed dormancy. Plant, Cell & Environment 35: 1769-1786 Gubler F, Millar AA, Jacobsen JV (2005) Dormancy release, ABA and preharvest sprouting. Current opinion in plant biology 8: 183-187 Gutierrez L, Van Wuytswinkel O, Castelain M, Bellini C (2007) Combined networks regulating seed maturation. Trends in Plant science 12: 294-300 Haque (1974) Physiological behavior of deep-water rice. In: Proceedings of the International Seminar on Deepwater Rice, August 21-26, 1974. No. 15. Bangladesh Rice Research Institute, 1975. Pp.184-190 Hasanuzzaman SM (1974) Cultivation of deepwater rice in Bangladesh. In: Proceedings of the International Seminar on Deepwater Rice, August 21-26, 1974. No. 15. Bangladesh Rice Research Institute, 1975, pp 137-147 Hatier JHB, Gould KS (2009) Anthocyanin function in vegetative organs. In Anthocyanins. Springer New York. pp. 1-19 Hattori Y, Nagai K, Furukawa S, Song XJ, Kawano R, Sakakibara H, Wu J, Matsumoto T, Yoshimura A, Kitano H, Matsuoka M, Mori H, Ashikari M (2009) The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deepwater. Nature 460: 1026-1031 145 Heim MA, Jakoby M, Werber M, Martin C, Weisshaar B, Bailey PC(2003) The basic helix-loop-helix transcription factor family in plants: a genome-wide study of protein structure and functional diversity. Molecular biology and evolution 20:735-747 Henry A, Gowda VR, Torres RO, McNally KL, Serraj R (2011) Variation in root system architecture and drought response in rice (Oryza sativa): Phenotyping of the OryzaSNP panel in rainfed lowland fields. Field crop research 120:205-214 Hichri I, Barrieu F, Bogs J, Kappel C, Delrot S, Lauvergeat V (2011) Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. Journal of experimental botany 62: 2465-2483 Hirano HY, Hirai A, Sano Y, Sasaki T (2008) Rice Biology in the Genomics Era, Biotechnology in Agriculture and Forestry. Vol. 62. Springer Hite DR, Auh C, Scandalios JG (1999) Catalase activity and hydrogen peroxide levels are inversely correlated in maize scutella during seed germination. Redox report 4: 29-34 Hobo T, Kowyama Y, Hattori T(1999) A bZIP factor, TRAB1, interacts with VP1 and mediates abscisic acid-induced transcription. Proceedings of the national academy of sciences 96: 15348-15353 Hoffmann-Benning S, Kende H (1992) On the role of abscisic acid and gibberellin in the regulation of growth in rice. Plant physiology 99: 11561161 Holdsworth MJ, Bentsink L, Soppe WJ (2008) Molecular networks regulating Arabidopsis seed maturation, after‐ripening, dormancy and germination. New phytologist 179: 33-54 Hossain M, Jaim WMH (2012) Diversity, spatial distribution, and the process of adoption of improved rice varieties in Bangladesh. In: Adoption and diffusion of modern rice varieties in Bangladesh and Eastern India. International Rice Research Institute, Los Baños 146 Hossain MA, Cho JI, Han M, Ahn CH, Jeon JS, An G, Park PB (2010a) The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. Journal of plant physiology 167 : 1512-1520 Hossain MA, Lee Y, Cho JI, Ahn CH, Lee SK, Jeon JS, Kang H, Lee CH, An G, Park PB (2010b) The bZIP transcription factor OsABF1 is an ABA responsive element binding factor that enhances abiotic stress signaling in rice. Plant molecular biology 72:557-566 Hu X, Zhang A, Zhang J, Jiang M (2006) Abscisic Acid is a Key Inducer of Hydrogen Peroxide Production in Leaves of Maize Plants Exposed to Water Stress. Journal of experimental botany 47: 1484-1495 Huang X, Kurata N, Wei X, Wang ZX, Wang A, Zhao Q, et al(2012) A map of rice genome variation reveals the origin of cultivated rice. Nature 490: 497501 Huke RE, Huke EH (1997) Rice area by type of culture: South, Southeast, and East Asia. IRRI, Los Baños, Philippines Ichikawa H, Ichiyanagi T, Xu B, Yoshii Y, Nakajima M, Konishi T (2001) Antioxidant activity of anthocyanin extract from purple black rice. Journal of medicinal food 4: 211-218 Ishibashi Y, Tawaratsumida T, Kondo K, Kasa S, Sakamoto M, Aoki N, ... & Iwaya-Inoue M (2012) Reactive oxygen species are involved in gibberellin/abscisic acid signaling in barley aleurone cells. Plant physiology 158: 1705-1714 Ismail AM, Ella ES, Vergara GV, Mackill DJ (2009) Mechanisms associated with tolerance to flooding during germination and early seedling growth in rice (Oryza sativa). Annals of botany 103:197-209 Ismail AM, Johnson DE, Ella ES, Vergara GV, Baltazar AM (2012) Adaptation to flooding during emergence and seedling growth in rice and weeds, and implications for crop establishment. AoB PLANTS 2012: pls019; doi:10.1093/aobpla/pls019 147 Izawa T, Foster R, Nakajima M, Shimamoto K, Chua NH (1994) The rice bZIP transcriptional activator RITA-1 is highly expressed during seed development. Plant cell 6: 1277-1287 Jackson MB, Pearce DM, Summers JE (1996) Tolerance to submergence and anaerobiosis: rice seedlings have something still to learn. In Physiology of Stress Tolerance in Rice: Proceedings of the international conference on stress physiology or rice, 28 Feb-5 March 1994, Lucknow, India . Manila, IRRI, pp31-43 Jakoby M, Weisshaar B, Droge-Laser W, Vicente-Carbajosa J, Tiedemann J, Kroj T, Parcy F (2002) bZIP transcription factors in Arabidopsis. Trends in plant science 7: 106-11 Jeon JS, Lee S, Jung KH, Jun SH, Jeong DH, Lee J, Kim C, Jang S, Yang K, Nam J, et al(2000) T-DNA insertional mutagenesis for functional genomics in rice. The plant journal 22: 561-70 Jia L, Wu Q, Ye N, Liu R, Shi L, Xu W, Zhi H, Rahman ANMRB, Xia Y, Zhang J (2012) Proanthocyanidins inhibit seed germination by maintaining a high level of abscisic acid in Arabidopsis thaliana (F). Journal of integrative plant biology 54: 663-73 Joo JH, Bae YS, Lee JS (2001) Role of auxin-induced reactive oxygen species in root gravitropism. Plant physiology 126: 1055-1060 Juliano BO (1993) Rice in human nutrition. Rome, Italy: Food and Agriculture Organization of the United Nations (FAO) Juliano BO, Bechtel DB (1985) The rice grain and its gross composition. In Rice: Chemistry and Technology; American Association of Cereal Chemists: St. Paul, MN Kato H, Namai H (1987) Floral characteristics and environmental factors for increasing natural outcrossing rate for F1 hybrid seed production of rice Oryza sativa L. Japanese Journal of Breeding 37:318-330 148 Keith KA, Raskin I, Kende H (1986) A comparison of the submergence response of deepwater and non-deepwater rice. Plant physiology 80: 479-482 Kendall SL, Hellwege A, Marriot P, Whalley C, Graham IA, Penfield S (2011) Induction of dormancy in Arabidopsis summer annuals requires parallel regulation of DOG1 and hormone metabolism by low temperature and CBF transcription factors. The plant cell 23: 2568-2580 Kende H, van der Knaap E, Cho HT (1998) Deepwater rice: A model plant to study stem elongation. Plant physiology 118:1105-1110 Khush G (1997) Origin, dispersal, cultivation and variation of rice. Plant molecular biology 35:25-34 Khush GS, Virk PS (2005) IR varieties and their impact. Los Baños (Philippines): International Rice Research Institute. 163 p Kim H, Hwang H, Hong JW, Lee YN, Ahn IP, Yoon IS, Kim BG (2012) A rice orthologue of the ABA receptor, OsPYL/RCAR5, is a positive regulator of the ABA signal transduction pathway in seed germination and early seedling growth. Journal of experimental botany 63:1013-1024 Kitahata N, Saito S, Miyazawa Y, et al. (2005) Chemical regulation of abscisic acid catabolism in plants by cytochrome P450 inhibitors. Bioorganic and Medicinal Chemistry 13:4491-4498 Kobata T, Uemuki N (2004) High temperatures during the grain-filling period do not reduce the potential grain dry matter increase of rice. Agronomy Journal 96:406-414 Kobayashi Y, Yamamoto S, Minami H, Kagaya Y and Hattori T (2004) Differential activation of the rice sucrose nonfermenting1- related protein kinase2 family by hyperosmotic stress and abscisic acid. Plant cell 16:11631177 Koes R, Verweij W, Quattrocchio F (2005) Flavonoids: a colorful model for the regulation and evolution of biochemical pathways. Trends in Plant science10: 236-242 149 Kondo M, Terasaka A, Umino M (1942) Experimental studies on photoperiodism in rice.III. [translation] Nogaku Kenkyu 33:l-24 Kucera B, Cohn MA, Leubner-Metzger G (2005) Plant hormone interactions during seed dormancy release and germination. Seed Science Research 15: 281-307 Kushiro T, Okamoto M, Nakabayashi K, Yamagishi K, Kitamura S, Asami T, Hirai N, Koshiba T, Kamiya Y, Nambara E (2004) The Arabidopsis cytochrome P450 CYP707A encodes ABA 8’-hydroxylases: key enzymes in ABA catabolism. EMBO Journal 23: 1647-1656 Lao NT, Long D, Kiang S, Coupland G, Shoue DA, Carpita NC, Kavanagh TA (2003) Mutation of a family 8 glycosyltransferase gene alters cell wall carbohydrate composition and causes a humidity-sensitive semi-sterile dwarf phenotype in Arabidopsis. Plant molecular biology 53: 687-701 Lee KH, Piao HL, Kim HY, Choi SM, Jiang F, Hartung W, ... & Hwang I (2006) Activation of glucosidase via stress-induced polymerization rapidly increases active pools of abscisic acid. Cell 126: 1109-1120 Lee YC, Lu, Chen CA, Casaretto PW, Yu SM (2003) An ABA-responsive bZIP protein, OsBZ8, mediates sugar repression of alpha-amylase gene expression. Plant physiology 119: 78-86 Leymarie J, Vitkauskaité G, Hoang HH, Gendreau E, Chazoule V, Meimoun P, ... Bailly C (2012) Role of reactive oxygen species in the regulation of Arabidopsis seed dormancy. Plant and cell physiology 53: 96-106 Li M, Xu W, Yang W, Kong Z, Xue Y (2007) Genome-wide gene expression profiling reveals conserved and novel molecular functions of the stigma in rice. Plant physiology 144: 1797-1812 Li Y, Zhang T, Shen ZW, Xu Y, Li JY (2013) Overexpression of maize anthocyanin regulatory gene Lc affects rice fertility. Biotechnology letters 35: 115-119 150 Li ZK, Zhang F (2013) Rice breeding in the post-genomics era: from concept to practice. Current opinion in plant biology 16: 261-269 Li ZX, Septiningsih EM, Quilloy-Mercado SM, McNally KL, Mackill DJ (2011) Identification of SUB1A alleles from wild rice Oryza rufipogon Griff. Genetic resources and crop evolution 58: 1237-1242 Lin CC., Kao CH (1995) NaCl stress in rice seedlings: starch mobilization and the influence of gibberellic acid on seedling growth. Botanical Bulletin of Academia Sinica 36:169-173 Liu C, Mao B, Ou S, Wang W, Liu L, Wu Y, ... Wang X (2013) OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant molecular biology 1-18 Liu C, Wu Y, Wang X (2012) bZIP transcription factor OsbZIP52/RISBZ5: a potential negative regulator of cold and drought stress response in rice. Planta 235: 1157-1169 Liu Y, Ye N, Liu R, Chen M, Zhang J (2010) H2O2 mediates the regulation of ABA catabolism and GA biosynthesis in Arabidopsis seed dormancy and germination . Journal of experimental botany 61: 2979-2990 Lloyd AM, Walbot V, Davis RW(1992)Arabidopsis and Nicotiana anthocyanin production activated by maize regulators R and C1. Science 258:1773-1775 Lloyd AM. et al. (1994) Epidermal cell fate determination in Arabidopsis: patterns defined by a steroid-inducible regulator. Science 266:436-439 Londo JP, Chiang YC, Hung KH, Chiang TY, Schaal BA (2006) Phylogeography of Asian wild rice, Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. Proceedings of the national academy of sciences of USA 103: 9578-83 Lowry DB (2012) Ecotypes and the controversy over stages in the formation of new species. Biological journal of the Linnean society 106: 241-257 Lu G, Gao C, Zheng X, Han B(2009) Identification of OsbZIP72 as a positive regulator of ABA response and drought tolerance in rice. Planta 229:605-615 151 Lu SJ, Yang ZT, Sun L, Sun L, Song ZT, Liu JX (2012) Conservation of IRE1regulated bZIP74 mRNA unconventional splicing in rice (Oryza sativa L.) involved in ER stress responses. Molecular plant 5: 504-514 Maclean JL, Dawe D, Hardy B, Hettel GP (2002) Rice almanac. CABI Publishing, Wallingford UK Magneschi L, Perata P (2009) Rice germination and seedling growth in the absence of oxygen. Annals of botany 103:181-196. Matiru VN, Dakora FD (2005) The rhizosphere signal molecule lumichrome alters seedling development in both legumes and cereals. New phytologist 166:439-444 McCouch SR, Kovach MJ, Sweeney M, Jiang H, Semon M (2012) The Dynamics of Rice Domestication: A Balance between Gene Flow and Genetic Isolation. Gepts P, Famula TR, Bettinger RL, Brush SB, Damania AB (Eds.) In : Biodiversity in Agriculture: Domestication, Evolution, and Sustainability, Cambridge University Press McInnis SM, Emery DC, Porter R, Desikan R, Hancock JT, Hiscock SJ (2006) The role of stigma peroxidases in flowering plants: insights from further characterization of a stigma-specific peroxidase (SSP) from Senecio squalidus (Asteraceae). Journal of experimental botany 57: 1835-1846 McNally KL, Childs KL, Bohnert R, Davidson RM, Zhao K, Ulat VJ, Zeller G, Clark RM, Hoen DR, Bureau TE, Stokowski R, Ballinger DG, Frazer KA, Cox DR, Padhukasahasram B, Bustamante CD, Weigel D, Mackill DJ, Bruskiewich RM, Ratsch G, Buell CR, Leung H, Leach JE (2009) Genomewide SNP variation reveals relationships among landraces and modern varieties of rice. Proceedings of the national academy of sciences of USA 106:12273-12278 Millar AA, Jacobsen JV, Ross JJ, Helliwell CA, Poole AT, Scofield G,Reid JB , Gubler F (2006) Seed dormancy and ABA metabolism in Arabidopsis and barley: the role of ABA 8¢-hydroxylase. Plant journal 45: 942- 954 152 Miro B, Ismail AM (2013) Tolerance of anaerobic conditions caused by flooding during germination and early growth in rice (Oryza sativa L.). Frontiers in Plant science, 4:269. doi: 10.3389/fpls.2013.00269 Miyata M, Yamamoto T, Komori T , Nitta N (2007) Marker assisted selection and evaluation of the QTL for stigma exsertion under japonica rice genetic background. Theoretical and applied genetics114: 539-548 Mohanty B, Herath V, Wijaya E, Yeo HC, Reyes BG, Lee DY (2012) Patterns of cis-element enrichment reveal potential regulatory modules involved in the transcriptional regulation of anoxia response of japonica rice. Gene 511: 235242 Molina J, Sikora M, Garud N, Flowers JM, Rubinstein S, Reynolds A, Huang P, Jackson S, Schaal BA, Bustamante CD, Boyko AR, Purugganan MD (2011) Molecular evidence for a single evolutionary origin of domesticated rice. Proceedings of the national academy of sciences of USA 108: 8351-8356 Moyer RA, Hummer KE, Finn CE, Frei B, Wrolstad RE (2002) Anthocyanins, phenolics, and antioxidant capacity in diverse small fruits: Vaccinium, Rubus, and Ribes. Journal of agricultural and food chemistry 50:519-525 Nagai K, Kuroha T, Ayano M, Kurokawa Y, Angeles-Shim RB, Shim JH, Yasui H, Yoshimura A, Ashikari M (2012) Two novel QTLs regulate internode elongation in deepwater rice during the early vegetative stage. Breeding science 62: 178 Nakabayashi K, Bartsch M, Xiang Y, Miatton E, Pellengahr S, Yano R, ... & Soppe WJ (2012) The time required for dormancy release in Arabidopsis is determined by DELAY OF GERMINATION1 protein levels in freshly harvested seeds. The plant cell 24: 2826-2838 Nakagawa H, Ohmiya K, Hattori T (1996) A rice bZIP protein, designated OSBZ8, is rapidly induced by abscisic acid. Plant journal 9: 217-227 Nakashima K, Yamaguchi-Shinozaki K (2013) ABA signaling in stress-response and seed development. Plant cell reports 32: 959-970 153 Nambara E, Marion-Poll A (2005) Abscisic acid biosynthesis and catabolism. Annual review plant biology 56: 165-185 Nambara E, Okamoto M, Tatematsu K, Yano R, Seo M, Kamiya Y (2010) Abscisic acid and the control of seed dormancy and germination. Seed science research 20: 55-67 Nantel A, Quatrano RS (1996) Characterization of three rice basic/leucine zipper factors, including two inhibitors of EmBP-1 DNA binding activity. Journal of biological chemistry 271: 31296-31305 Nasiruddin M, Hassan MS (2009) Subsector: rice , Agricultural Research Priority : Vision 2030 and Beyond. Bangladesh Agricultural Research Council, Dhaka Neill S, Desikan R, Hancock J (2002) Hydrogen peroxide signalling. Current opinion in plant biology 5:388-395 Nijhawan A, Jain M, Tyagi AK, Khurana JP(2008) Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice. Plant physiology 146: 333-50 Ogawa K, Iwabuchi M(2001) A mechanism for promoting the germination of Zinnia elegans seeds by hydrogen peroxide. Plant and cell physiology 42: 286-291 Oh SJ, Kim YS, Kwon CW, Park HK, Jeong JS, Kim JK (2009) Overexpression of the transcription factor AP37 in rice improves grain yield under drought conditions. Plant physiology 150:1368- 1379 Oka HI, Chang WT (1960) Survey of variations in photoperiodic response in wild Oryza species. Bot Bull Acad Sinica 1: 1-14 Okamoto M, Kuwahara A, Seo M, Kushiro T, Asami T, Hirai N, Kamiya Y, Koshiba T, Nambara E (2006) CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant physiology 141: 97-107 Okamoto M, Tanaka Y, Abrams SR, Kamiya Y, Seki M, Nambara E(2009) High humidity induces abscisic acid 8′-hydroxylase in stomata and vasculature to 154 regulate local and systemic abscisic acid responses in Arabidopsis. Plant physiology 149: 825-834 Okamoto M, Tatematsu K, Matsui A, Morosawa T, Ishida J, Tanaka M , et al. (2010) Genome‐wide analysis of endogenous abscisic acid‐mediated transcription in dry and imbibed seeds of Arabidopsis using tiling arrays. The plant journal 62:39-51 Oliver MJ, Guo L, Alexander DC, Ryals JA, Wone BW, Cushman JC (2011) A sister group contrast using untargeted global metabolomic analysis delineates the biochemical regulation underlying desiccation tolerance in Sporobolus stapfianus. The plant cell 23:1231-1248 Onodera Y, Suzuki A, Wu CY, Washida H, Takaiwa F (2001) A rice functional transcriptional activator, RISBZ1, responsible for endospermspecific expression of storage protein genes through GCN4 motif. Journal of biological chemistry 276: 14139-14152 Onofri A (2007) Routine statistical analyses of field experiments by using an Excel extension.In: Proceedings 6th National Conference Italian Biometric Society: "La statistica nelle scienze della vita e dell’ambiente", Pisa, 20-22 June 2007, 93-96 Oracz K, Bouteau HEM, Farrant JM, Cooper K, Belghazi M, Job C, ... Bailly C (2007) ROS production and protein oxidation as a novel mechanism for seed dormancy alleviation. The plant journal 50: 452-465 O'Toole JC, Cruz RT (1980) Response of leaf water potential, stomatal resistance, and leaf rolling to water stress. Plant physiology 65: 428-432 Passardi F, Longet D, Penel C, Dunand C (2004) The class III peroxidase multigene family in rice and its evolution in land plants. Phytochemistry 65: 1879-1893 Patra B, Pattanaik S, Yuan L (2013) Ubiquitin protein ligase 3 mediates the proteasomal degradation of GLABROUS 3 and ENHANCER OF GLABROUS 3, regulators of trichome development and flavonoid biosynthesis in Arabidopsis. The plant journal 74:435-447 155 Perez AT, Nasiruddin M (1974) Field notes on the Rayadas: a flood-tolerant deepwater rice of Bangladesh. In: Proceedings of the International Seminar on Deepwater Rice, August 21-26, 1974. No. 15. Bangladesh Rice Research Institute, 1975, pp 87-91 Phillips DA, Joseph CM, Yang GP, Martínez-Romero E, Sanborn JR, Volpin H (1999) Identification of lumichrome as a Sinorhizobium enhancer of alfalfa root respiration and shoot growth. Proceedings of the national academy of sciences 96: 12275-12280 Potocky M , Jones MA , Bezvoda R , Smirnoff N , Zarsky V (2007) . Reactive oxygen species produced by NADPH oxidase are involved in pollen tube growth. New phytologist 174: 742 - 751 Puntarulo S, Sa´nchez RA, Boveris A (1988) Hydrogen peroxide metabolism in soybean embryonic axes at the onset of germination. Plant physiology 86: 626-630 Qin X, Zeevaart JAD (1999) The 9-cis-epoxycarotenoid cleavage reaction is the key regulatory step of abscisic acid biosynthesis in water-stressed bean. Proceedings of the national academy of sciences 96: 15354-15361 Rabbani MA, Maruyama K, Abe H, Khan MA, Katsura K, Ito Y, Yoshiwara K, Seki M, Shinozaki K, Yamaguchi-Shinozaki K (2003) Monitoring expression profiles of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA microarray and RNA gel-blot analyses. Plant physiology 133:1755-1767 Raghavendra AS, Gonugunta VK, Christmann A, Grill E (2010) ABA perception and signalling. Trends in plant science 15: 395-401 Ramsay NA, Glover BJ (2005) MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends in plant science, 10: 63-70 Rapisarda P, Fanella F, Maccarone E (2000) Reliability of analytical methods for determining anthocyanins in blood orange juices. Journal of agricultural and food chemistry 48:2249-2252 156 Ray H, Yu M, Auser P, Blahut-Beatty L, McKersie B, Bowley S, Westcott N, Coulman B, Lloyd A, Gruber MY(2003) Expression of anthocyanins and proanthocyanidins after transformation of alfalfa with maize Lc. Plant physiology132:1448-1463 Raz V, Bergervoet JHW, Koornneef M (2001) Sequential steps for developmental arrest in Arabidopsis seeds. Development 128: 243-252 Ren H, Gao Z, Chen L, Wei K, Liu J, Fan Y, Davies WJ, Jia W, Zhang J(2007) Dynamic analysis of ABA accumulation in relation to the rate of ABA catabolism in maize tissues under water deficit. Journal of experimental botany 58: 211-9 Rio LAD, Puppo A (2009) Reactive oxygen species in plant signaling. Springer Rodriguez AA, Grunberg K, Taleisnik EL (2002) Reactive oxygen species in the elongation zone of maize leaves are necessary for leaf expansion. Plant physiology 129: 1627-1632 Saika H, Okamoto M, Miyoshi K, Kushiro T, Shinoda S, Jikumaru Y, Fujimoto M, Arikawa T, Takahashi H, Ando M, et al (2007) Ethylene promotes submergence-induced expression of OsABA8ox1, a gene that encodes ABA 8'-hydroxylase in rice. Plant and cell physiology 48: 287-298 Saito S, Hirai N, Matsumoto C, Ohigashi H, Ohta D, Sakata K, Mizutani M(2004) Arabidopsis CYP707As encode (+)-abscisic acid 8′-hydroxylase, a key enzyme in the oxidative catabolism of abscisic acid. Plant physiology 134:1439-1449 Sang T, Ge S (2013) Understanding rice domestication and implications for cultivar improvement. Current opinion plant biology 16: 1-8 Schoft VK, Chumak N, Choi Y., Hannon M., Garcia-Aguilar M, Machlicova A, ... Tamaru H (2011) Function of the DEMETER DNA glycosylase in the Arabidopsis thaliana male gametophyte. Proceedings of the National Academy of Sciences108:8042-8047 157 Schwartz SH, Zeevaart JA (2010) Abscisic acid biosynthesis and metabolism. In Plant Hormones (pp. 137-155). Springer Netherlands. Seki M, Kamei A, Yamaguchi-Shinozaki K, Shinozaki K (2003) Molecular responses to drought, salinity and frost: common and different paths for plant protection. Current opinion in biotechnology 14:194-199 Seki M, Umezawa T, Urano K, Shinozaki K (2007) Regulatory metabolic networks in drought stress responses. Current opinion in plant biology 10: 296-302 Septiningsih EM, Ignacio JCI, Sendon PM, Sanchez DL, Ismail AM, Mackill DJ (2013) QTL mapping and confirmation for tolerance of anaerobic conditions during germination derived from the rice landrace Ma-Zhan Red. Theoretical and applied genetics, 1-10 Sharp RE, Davies WJ (1989) Regulation of growth and development of plants growing with a restricted supply of water. In: Jones HG, Flowers TL, Jones MB, eds. Plants under stress. Cambridge: Cambridge University Press, 71-93 Shen SQ, Shu QY, Bao JS, Wu DX, Cui HR , Xia YW (2004) Development of a greenable leaf colour mutant Baifeng A and its application in hybrid rice production. Chinese J Rice Sci 18: 34-38. (In Chinese with English abstract) Shigeoka S, Ishikawa T, Tamoi M, Miyagawa Y, Takeda T, Yabuta Y, Yoshimura K (2002) Regulation and function of ascorbate peroxidase isoenzymes. Journal of experimental botany 53: 1305-1319 Shimizu H, Sato K, Berberich T, Miyazaki A, Ozaki R, Imai R, Kusano T (2005) LIP19, a basic region leucine zipper protein, is a Fos-like molecular switch in the cold signaling of rice plants. Plant and cell physiology 46: 1623-1634 Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks involved in drought stress response and tolerance. Journal of experimental botany 58:221-227 Sohail Q, Inoue T, Tanaka H, Eltayeb AE, Matsuoka Y, Tsujimoto H (2011) Applicability of Aegilops tauschii drought tolerance traits to breeding of hexaploid wheat. Breeding science 61: 347 158 Speranza A, Crinelli R, Scoccianti V, Geitmann A (2012) Reactive oxygen species are involved in pollen tube initiation in kiwifruit. Plant biology 14: 64-76 Sreenivasulu N, Radchuk V, Alawady A, Borisjuk L, Weier D, Staroske et al. (2010) De‐regulation of abscisic acid contents causes abnormal endosperm development in the barley mutant seg8. The plant journal 64: 589-603 Steffens B, Geske T, Sauter M (2011) Aerenchyma formation in the rice stem and its promotion by H2O2. New phytologist 190: 369-378 Steffens B, Sauter M (2009) Epidermal cell death in rice is confined to cells with a distinct molecular identity and is mediated by ethylene and H2O2 through an autoamplified signal pathway. The plant cell 21:184-196 Steyn WJ (2009) Prevalence and functions of anthocyanins in fruits. In Anthocyanins (pp. 86-105). Springer New York Sugimoto K, Takeuchi Y, Ebana K, Miyao A, Hirochika H, Hara N, ... & Yano, M (2010) Molecular cloning of Sdr4, a regulator involved in seed dormancy and domestication of rice. Proceedings of the national academy of sciences 107: 5792-5797 Sun C, Wang X, Yoshimura A, Doi K (2002) Genetic differentiation for nuclear, mitochondrial and chloroplast genomes in common wild rice (Oryza rufipogon Griff.) and cultivated rice (Oryza sativa L.). Theoretical and applied genetics104:1335-1345 Suslow T, Thomas B, Bradford K (2002) Biotechnology provides new tools for plant breeding. Agricultural Biotechnology in California Series, Publication 8043, Seed Biotechnology Center, UC Davis Taheri P, Tarighi S (2010) Riboflavin induces resistance in rice against Rhizoctonia solani via jasmonate-mediated priming of phenylpropanoid pathway. Journal of plant physiology 167: 201-208 159 Takahashi H, Kawakatsu T, Wakasa Y, Hayashi S, Takaiwa F (2012) A rice transmembrane bZIP transcription factor, OsbZIP39, regulates the endoplasmic reticulum stress response. Plant and cell physiology 53:144-153 Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Molecular biology and evolution 28: 2731-2739 Tang H, Sezen U, Paterson AH (2010) Domestication and plant genomes. Current opinion in plant biology 13:160-166 Tominaga-Wada R, Iwata M, Nukumizu Y, Sano R, Wada T (2012) A full-length R-like basic-helix-loop-helix transcription factor is required for anthocyanin up-regulation whereas the N-terminal region regulates epidermal hair formation. Plant science 183:115-122 Tsugama D, Liu S, Takano T (2012) A bZIP protein, VIP1, is a regulator of osmosensory signaling in Arabidopsis. Plant physiology 159: 144-155 Tzfira T, Vaidya M, Citovsky V(2001) VIP1, an Arabidopsis protein that interacts with Agrobacterium VirE2, is involved in VirE2 nuclear import and Agrobacterium infectivity. The EMBO journal 20: 3596-607 Uga Y, Siangliw M, Nagamine T, Ohsawa R, Fujimura T , Fukuta Y (2010) Comparative mapping of QTLs determining glume, pistil and stamen sizes in cultivated rice (Oryza sativa L.). Plant breeding 129: 657-669 UgaY, Fukuta Y , Cai HW, Iwata H, Ohsawa R, Morishima H ,Fujimura T (2003) Mapping QTLs influencing rice floral morphology using recombinant inbred lines derived from a cross between Oryza sativa L. and Oryza rufipogon Griff. Theoretical and applied genetics 107: 218-226 Umezawa T, Nakashima K, Miyakawa T, Kuromori T, Tanokura M, Shinozaki K, Yamaguchi-Shinozaki K (2010) Molecular basis of the core regulatory network in ABA responses: sensing, signaling and transport. Plant and cell physiology 51: 1821-1839 160 Umezawa T, Okamoto M, Kushiro T, Nambara E, Oono Y, Seki M, Kobayashi M, Koshiba T, Kamiya Y, Shinozaki K(2006) CYP707A3, a major ABA 8′hydroxylase involved in dehydration and rehydration response in Arabidopsis thaliana. Plant journal 46: 171-182 Van Breusegem F and Dat JF (2006) Reactive oxygen species in Plant cell death. Plant physiology 141: 384-390 Varshney RK , Koebner RMD (2006) Model Plants and Crop Improvement. CRC Press Inc, Florida, USA Vaughan DA (1994) The wild relatives of rice: a genetic resources handbook. . International Rice Research Institute, Los Baños Veasey EA, Karasawa MG, Santos PP, Rosa MS, Mamani E, Oliveira GCX (2004) Variation in the loss of seed dormancy during after-ripening of wild and cultivated rice species. Annals of botany 94:875-882 Vergara BS, Chang T T (1985) The flowering response of the rice plant to photoperiod: A review of the literature, 4th edn, International Rice Research, Los Baños Villareal CP, Juliano BO (1989) Variability in contents of thiamine and riboflavin in brown rice, crude oil in brown rice and bran-polish, and silicon in hull of IR rices. Plant foods for human nutrition 39: 287-297 Voesenek LACJ, Bailey-Serres J (2009) Genetics of high-rise rice. Nature 460: 959-960 Volpin H, Phillips DA (1998) Respiratory eclicitors from Rhizobium meliloti affect intact alfalfa roots. Plant physiology 116: 777-783 Walker AR, Davison PA, olognesi-Winfield AC, James CM, Srinivasan N, Blundell TL, Esch JJ, Marks MD, Gray JC (1999) The TRANSPARENT TESTA GLABRA1 locus, which regulates trichome differentiation and anthocyanin biosynthesis in Arabidopsis, encodes a WD40 repeat protein. Plant cell 11:1337-1350 161 Wang JC, Xu H, Zhu Y, Liu QQ, Cai XL (2013) OsbZIP58, a basic leucine zipper transcription factor, regulates starch biosynthesis in rice endosperm. Journal of experimental botany 64: 3453-3466 Wang M, Zhu Z, Tan L, Liu F, Fu Y, Sun C, Cai H (2013) Complexity of indicajaponica varietal differentiation in Bangladesh rice landraces revealed by microsatellite markers. Breeding science 63: 227 Watt G (1891) A Dictionary of the Economic Products of India. Vol. V. Cosmo Publications, Delhi (Reprint 1972) Weerakoon WMW, Maruyama A, Ohba K (2008) Impact of Humidity on Temperature‐Induced Grain Sterility in Rice (Oryza sativa L). Journal of agronomy and crop science 194: 135-140 Weitbrecht K, Müller K, Leubner-Metzger G. 2011. First off the mark: early seed germination. Journal of experimental botany 62: 3089-3309 Wood JP, Mammone T, Chidlow G, Greenwell T, Casson RJ (2012) Mitochondrial inhibition in rat retinal cell cultures as a model of metabolic compromise: mechanisms of injury and neuroprotection. Investigative Ophthalmology & Visual Science 53:4897-4909 Xiang Y, Tang N, Du H, Ye H, Xiong L (2008) Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant physiology 148: 1938-1952 Xiong Y, Liu T, Tian C, Sun S, Li J, Chen M (2005) Transcription factors in rice: a genome-wide comparative analysis between monocots and eudicots. Plant molecular biology 59: 191-203 Xu K, Xu X, Fukao T, Canlas P, Maghirang-Rodriquez R, Huer S, Ismail AM, Bailey-Serres J, Ronald PC, Mackill DJ (2006) Sub1A is an ethyleneresponse-factor-like gene that confers submergence tolerance to rice. Nature 442: 705-708 162 Xu ZY, Lee KH, Dong T, Jeong JC, Jin JB, Kanno Y, & Hwang I (2012) A vacuolar β-glucosidase homolog that possesses glucose-conjugated abscisic acid hydrolyzing activity plays an important role in osmotic stress responses in Arabidopsis. The plant cell 24: 2184-2199 Yamakawa H, Hirose T, Kuroda M, Yamaguchi T (2007) Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant physiology 144:258-277 Yamamoto MP, Onodera Y, Touno SM, Takaiwa F (2006) Synergism between RPBF Dof and RISBZ1 bZIP activators in the regulation of rice seed expression genes. Plant physiology 141: 1694-1707 Yamamoto T, Takemori N, Sue N ,Nitta N (2003) QTL analysis of stigma exsertion in rice. Rice Genet Newsl 20: 33-34 Yamauchi M, Aguilar AM, Vaughan DA, Seshu DV (1993) Rice (Oryza sativa L.) germplasm suitable for direct sowing under flooded soil surface. Euphytica 67: 177-184 Yan WG, Li Y, Agrama HA, Luo D, Gao ,Lu X, Ren G (2009) Association mapping of stigma and spikelet characteristics in rice (Oryza sativa L.). Molecular Breeding 24: 277-292 Yanagita T, Foster JW (1956) A bacterial riboflavin hydrolase. Journal of Biological Chemistry 221: 593-607 Yang J, Zhang J (2010) Grain-filling problem in ‘super’rice. Journal of experimental botany 61:1-5 Yang D, Wu L, Hwang YS, Chen L, Huang N (2001) Expression of the REB transcriptional activator in rice grains improves the yield of recombinant proteins whose genes are controlled by a Reb-responsive promoter. Proceedings of the national academy of sciences of USA 98: 11438-11443 Yang J, Zhang J (2006) Grain filling of cereals under soil drying. New phytologist 169: 223-236 163 Yang SH, Zeevaart JA (2006) Expression of ABA 8′-hydroxylases in relation to leaf water relations and seed development in bean. The plant journal 47: 675686 Yang SH, Choi D (2006) Characterization of genes encoding ABA 8′-hydroxylase in ethylene-induced stem growth of deepwater rice. Biochemical and biophysical research communications 350: 685-690 Ye N, Jia L, Zhang J (2012b) ABA signal in rice under stress conditions. Rice 5: 1 Ye N, Zhu G, Liu Y, Li Y, Zhang J (2011) ABA controls H2O2 accumulation through the induction of OsCATB in rice leaves under water stress. Plant and cell physiology 52:689-698 Ye N, Zhu G, Liu Y, Zhang A, Li Y, Liu R, ... & Zhang J (2012a) Ascorbic acid and reactive oxygen species are involved in the inhibition of seed germination by abscisic acid in rice seeds. Journal of experimental botany 63: 1809-1822 Yuan Y, Chiu LW, Li L (2009). Transcriptional regulation of anthocyanin biosynthesis in red cabbage. Planta 230:1141-1153 Yue B, Cui K, Yu S, Xue W, Luo L, Xing Y (2006) Molecular marker-assisted dissection of quantitative trait loci for seven morphological traits in rice (Oryza sativa L.). Euphytica 150: 131-139 Zaman SMH (1977) Progress of deep-water rice research in Bangladesh. In: Proceedings of the workshop on deep-water rice 1976. International Rice Research Institute, Los Los Baños, pp 127-134 Zetian H, Yanrong W (2009) Advances in japonica hybrid rice breeding. Accelerating Hybrid Rice Development, IRRI, Manila, Philippines, pp 139149 Zhang H, Jin JP, Tang L, Zhao Y, Gu XC, Gao G, Luo JC (2011) Plant TFDB 2.0: update and improvement of the comprehensive plant transcription factor database. Nucleic acids research 39: D1114-D1117 164 Zhang X, Zhang L, Dong F, Gao J, Galbraith DW, Song CP (2001) Hydrogen peroxide is involved in abscisic acid-induced stomatal closure in Vicia faba. Plant physiology 126: 1438-1448 Zhu G, Ye N, Zhang J (2009) Glucose-induced delay of seed germination in rice is mediated by the suppression of ABA catabolism rather than an enhancement of ABA biosynthesis. Plant and cell physiology 50: 644-651 Zhu Y, Nomura T, Xu Y, Zhang Y, Peng Y, et al. (2006). ELONGATED UPPERMOST INTERNODE encodes a cytochrome P450 monooxygenase that epoxidizes gibberellins in a novel deactivation reaction in rice. Plant cell 18:442-56 Zou M, Guan Y, Ren H, Zhang F, Chen F (2008) A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant molecular biology 66: 675-68 165 CURRICULUM VITAE Academic qualifications of the thesis author, Mr. BIN RAHMAN A N M Rubaiyath: Received the degree of Bachelor of Science (Honours) from University of Dhaka, January 1997. Received the degree of Master of Science from University of Dhaka, January 1999. November 2013 166