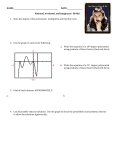
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
4.3.4 Marine Fish 4.3.4.1 Smalltooth sawfish Description and distribution The smalltooth sawfish is a tropical marine and estuarine elasmobranch (sharks, skates, and rays) that have a circumtropical distribution. In the western Atlantic Ocean, the smalltooth sawfish has been reported from Brazil through the Caribbean Sea and, the Gulf of Mexico, and the Atlantic coast of the United States (Bigelow and Schroeder, 1953). Historic capture records within the U.S. range from Texas to New York. Water temperatures no lower than 16-18 °C and the availability of appropriate coastal habitat appear to serve as the major environmental constraints limiting the northern movements of smalltooth sawfish in the western North Atlantic Ocean. As a result, most records of this species from areas north of Florida occur during spring and summer periods (May to August) when inshore waters reach warmer temperatures. These larger individuals (> 3.0 m) captured along the Atlantic coast north of Florida represent seasonal migrants from a core population in southern states rather than being members of a continuous, uniform-density population (Bigelow and Schroeder, 1953). It is likely that these individuals migrated southward toward Florida as water temperatures declined in the fall, as there is only one winter record from the Atlantic coast north of Florida. Reproduction Fertilization in smalltooth sawfish is internal. Development in sawfish is believed to be ovoviviparous. The embryos of smalltooth sawfish, while still bearing the large yolk sac, already resemble adults relative to the position of their fins and absence of the lower caudal lobe. During embryonic development the rostral blade is soft and flexible. The rostral teeth are also encapsulated or enclosed in a sheath until birth. Shortly after birth, the teeth become exposed and attain their full size proportionate to the size of the saw. The size at birth is approximately 80 cm, with the smallest free-living specimens reported during field studies in Florida being 77 - 84 cm (Simpfendorfer, unpublished data). Bigelow and Schroeder (1953) reported gravid females carry 15-20 embryos. Simpfendorfer (2000) estimated age at maturity between 10 and 20 years. Studies of largetooth sawfish in Lake Nicaragua (Thorson 1976) report brood sizes of 1-13 individuals, with a mean of 7.3 individuals. The gestation period for largetooth sawfish is approximately 5 months and females likely produce litters every second year. Although there are no such studies on smalltooth sawfish, its similarity in size and habitat to the largetooth sawfish implies that their reproductive biology may be similar. Development, growth and movement patterns To date, no formal studies on the age and growth of wild smalltooth sawfish have been conducted. Individuals have been maintained in public aquaria for up to 20 years (Cerkleski, pers. comm., 2000) which can provide some information on growth. For example, Bohoroquez (2001) reported three specimens in Columbia grew at an average rate of 19.6 cm/year (one animal grew from 84 cm to 320 cm in 12 years). In a more comprehensive study, Clark et al. (2004) reported an average growth rate of 13.9 cm/year for16 captive smalltooth sawfish (size from 80 cm to 412 cm) in North American. Based on their growth parameters Clark et al. (2004) estimated an age at maturity of 19 yr for males and 33 yr for females while Simpfendorfer (2000) estimated age at maturity between 10 and 20 years, and a maximum age of 30 to 60 yr. Sawfish in general inhabit the shallow coastal waters but recent data suggests adults can be found to much greater depths. Simpfendorfer and Wiley (2005) reported close associations between encounters and mangroves, seagrasses, and the shoreline than expected at random. Encounter data have also demonstrated that smaller smalltooth sawfish occur in shallower water and larger sawfish occur regularly at depths greater than 10 meters. Poulakis and Seitz (2004) reported that almost half of all sawfish < 3 meters in length were found in water less than 10 meters deep and 46% of encounters with adult sawfish (> 3 meters) in Florida bay and the Florida Keys occurred at depths between 70 to 122 meters. Simpfendorfer and Wiley (2005) also reported a substantial number of larger sawfish in depths greater than 10 meters. Encounter data bases have also identified river mouths as areas where many people observe sawfish. Seitz and Poulakis (2002) noted that many of the encounters occurred at or near river mouths in southwest Florida. Simpfendorfer and Wiley (2005) reported a similar pattern of distribution along the entire west coast of Florida. It is unclear whether this observation represents a preference for river mouths because of physical characteristics. Acoustic tracking results for very small smalltooth sawfish indicate depth and red mangrove root systems are potentially important in helping them avoid predators (Simpfendorfer 2003). Small juveniles have many of the same habitat use characteristics seen in the very small sawfish. However, their association with very shallow water (< 1 ft deep) is weaker, possibly because they are better suited to predator avoidance due to their larger size and greater experience. They do, however, still have a preference for shallow water, remaining in depths mostly less than three feet (90 cm). Information on the habitat use of adult smalltooth sawfish suggests that adult sawfish occur from shallow coastal waters to deeper shelf waters. Abundance and status of stocks There are few long-term abundance data sets that include smalltooth sawfish. Catch data from shrimp trawlers off Louisiana from the late 1940‟s to the 1970‟s suggests a rapid decline in the species from the period 1950-1964. However, this data set has not been validated nor subjected to statistical analysis to correct for factors unrelated to abundance. The Everglades National Park has established a fisheries monitoring program based on sport fisher dock-side interviews since 1972 (Schmidt et al. 2000). Analysis of these data using a lognormal generalized linear model to correct for factors unrelated to abundance (e.g., change in fishing practices) indicate a slight increasing trend in abundance for smalltooth sawfish in the park (Carlson et al. 2007). From 1989-2002, smalltooth sawfish relative abundance has increased by about 5% per year. Using a demographic approach and life history data for smalltooth sawfish and similar species from the literature, Simpfendorfer (2000) estimated intrinsic rates of natural population increase as 8-13% per year and population doubling times from 5.4 years to 8.5 years. Musick (1999) and Musick et al. (2000) noted that intrinsic rates of increase less than ten percent were low, and are particularly vulnerable to excessive mortalities and rapid population declines, after which recovery may take decades. As such, smalltooth sawfish populations will recover slowly from depletion, confounding recovery efforts. Simpfendorfer (2000) concluded that recovery was likely to take decades or longer depending on how effectively sawfish could be protected. Ecological relationships Unknown. Listing Designations The National Marine Fisheries Service (NMFS) added the smalltooth sawfish to its list of candidate species in 1991 (56 FR 26797). Smalltooth sawfish were removed from the list in 1997 (62 FR 37561), but returned on a revised list published June 23, 1999 (64 FR 33466). On November 30, 1999, NMFS received a petition from the Center for Marine Conservation requesting listing of the North American population of smalltooth sawfish as endangered under the Endangered Species Act. Subsequently, NMFS announced the initiation of a smalltooth sawfish formal status review (65 FR 12959) and published a proposed rule (66 FR 19414) to list the U.S. population of smalltooth sawfish as endangered under the Endangered Species Act. On April 1, 2003, NMFS listed the U.S. population of smalltooth sawfish as an endangered species (50 CFR 224). A recent proposal by the United States of America to list all sawfish under Appendix 1 of the Convention on International Trade in Endangered Species of Wild Fauna and Flora banning international trade was approved. The convention made an exception for one species of sawfish, listing them on Appendix II, allowing Australia to continue trade in live animals if it does not hinder the conservation of the species. 4.3.4.2 Shortnose sturgeon Description and Distribution The shortnose sturgeon (Acipenser brevirostrum) is a federally-listed endangered species, for which a Recovery Plan has been prepared (NMFS 1998). The species occurs in South Atlantic rivers, and on occasion in marine waters. There are apparently no documented records from the EEZ in the South Atlantic (Mark Collins, SC Department of Natural Resources, personal communication to R.W. Laney); however, the species is included here since the Council also must consider the impacts of its plans on federally listed protected species. It is an anadromous species which historically, with the Atlantic sturgeon, formed the basis for a valuable commercial fishery. Information in this account is taken largely from the December 1998 Final Recovery Plan for the species (NMFS 1998), with supplementation from other sources as cited. The shortnose sturgeon occurs in large coastal rivers of eastern North America, historically from the St. John River in New Brunswick, Canada to the Indian River, Florida (NMFS 1998). NMFS currently recognizes 19 distinct population segments of shortnose sturgeon inhabiting 25 river systems ranging from the Saint John River in New Brunswick, Canada, to the St. Johns River, Florida. Genetic analysis of 11 of these population segments indicate most rivers and estuaries currently harboring shortnose sturgeon and identified as population segments in the NMFS 1998 Recovery Plan contain genetically distinct populations. Of the 11 population segments tested, at least 9 genetically distinct population segments of shortnose sturgeon were identified. Most comparisons among collections in the southeast showed significant genetic discontinuities between adjoining rivers, despite geographic proximity of their river mouths (20-100 km) (Wirgin et al. 2006). Shortnose sturgeon historically occurred in most rivers of the four South Atlantic states from the Albemarle Sound system in NC through the Indian River system in FL. One relatively recent capture (1998) of a shortnose sturgeon in the Albemarle Sound system has been documented (Wayne Starnes, North Carolina Museum of Natural Sciences, personal communication). There have been no recent documented captures in the Pamlico Sound and its tributaries (Tar, Neuse Rivers). However, there are two recent reports of sturgeon from the Pamlico Sound identified by commercial fishery observers that were allegedly shortnose, but no photographs or tissue were taken for confirmation by professional fishery biologists or geneticists (Wilson Laney, USFWS, personal communication). There is currently a small population of shortnose in the Cape Fear River, North Carolina. At least one population presently exists in the Winyah Bay system (Waccamaw, Pee Dee and Black Rivers), as well as the Santee River, the Cooper River, the ACE Basin (Ashepoo, Combahee and Edisto Rivers) and the Savannah River. Georgia populations occur in the Savannah, Ogeechee, Altamaha, St. Marys and Satilla Rivers. Florida has shortnose presently only in the St. Johns River. The shortnose is a small species of sturgeon, reaching maturity at fork lengths of 45-50 cm (1820 in) and maximum size of approximately 120 cm (47 in) (Dadswell et al. 1984). It differs from juvenile Atlantic sturgeon in having a shorter nose, wider mouth, and no enlarged bony plates between the base of the anal fin and the lateral row of scutes (Gilbert 1989). Shortnose sturgeon are found in rivers, estuaries and the sea, but populations spend most of their time in their natal rivers and estuaries (NMFS 1998). In the southeast, the species is estuarine anadromous (i.e., spends most of the year in estuaries and ascends the freshwater portions of rivers to spawn in the spring). Adults in rivers in the south Atlantic forage at the interface of fresh tidal water and saline estuaries and migrate to the upper reaches of rivers to spawn during early spring (Savannah River: Hall et al. 1991; Altamaha River: Heidt and Gilbert 1979; Flouronoy et al. 1992, Rogers and Weber 1995a; Ogeechee River: Weber 1996). Reproduction Length and age at maturity Length at maturity (45-55 cm FL) is similar throughout the shortnose sturgeon‟s range, but because fish in southern rivers grow faster than those in northern rivers, southern fish mature at younger ages. Males spawn first at 2-3 years in Georgia and 3-5 years in South Carolina, whereas females first spawn at 6 years or less in the Savannah River (Dadswell et al. 1984). Most shortnose sturgeon probably survive spawning, although there is some post-spawning mortality (B. Kynard, United States Geological Survey, personal observation). Spawning periodicity Spawning periodicity is poorly understood, but males seem to spawn more frequently than females. At least some males and females in the Savannah River may spawn in consecutive years, but most apparently do not (Collins and Smith 1993). Males may spawn every 1-2 years, but the minimum duration between spawning events for females is generally 3 years (Dadswell 1979, Cooke et al. 2002). Fecundity and sex ratio Gonadal maturity and fecundity of females were characterized by Dadswell (1979) for the Saint John River, Canada. Just prior to spawning, egg diameter was 3.1 mm and the ovaries composed 25 percent of the body weight. The number of eggs released ranged from 27,000 to 208,000 (11,568 eggs/kg body weight). Males were most abundant in the sex ratio estimate for the Savannah River (3.5:1, Collins and Smith 1997). Spawning behavior The shortnose sturgeon spawning period is estimated to last from a few days to multiple weeks. Sturgeon in the Savannah River remained on the spawning grounds for 2-3 weeks (Hall et al. 1991). Altamaha River fish remained on suspected spawning grounds for as long as nine weeks (Rogers and Weber 1995a). Cooper River fish residenced the spawning site for 89 days, longer than reports from other populations. It is hypothesized that these individuals were actively searching for an upstream route past Pinopolis Dam, but to no avail (Cooke and Leach, 2004a). Males fertilize the female‟s eggs as the eggs are released close to the substrate. In captivity, males nuzzle the anal and head areas of females, suggesting that females attract males with a chemical attractant (B. Kynard, personal observation). Spawning habitat In populations that have free access to the total length of a river, (e.g., no dam within the species‟ range in the river) spawning areas are located at the most upstream reach of the river used by sturgeon (Saint John, Kennebec, and Altamaha rivers: Dadswell et al. 1984, Rogers and Weber 1995a; Savannah River: Hall et al. 1991). Characteristic channel spawning habitats vary slightly among rivers: in curves with gravel/sand/log substrate in the Savannah River (Hall et al. 1991) and areas near limestone bluffs with gravel to boulder substrate in the Altamaha River (Rogers and Weber 1995a). Timing of spawning and river conditions Spawning begins in freshwater from late winter/early spring in southern rivers when water temperatures increase to 8-9°C. Spawning usually ceases when water temperatures reach 12-15° C (Dadswell et al. 1984; Buckley and Kynard 1985b; Hall et al. 1991; O‟Herron et al 1993; Squiers et al. 1993; Kynard 1997). However, shortnose sturgeon may spawn at higher temperatures. For example, when high river flow conditions delayed spawning in the Connecticut River, shortnose sturgeon had the physiological flexibility to spawn successfully at 18° C (Kynard 1997). Spawning was detected from 11.5-19°C in the Cooper River, SC (Cooke and Leach, 2004a). In general, spawning occurs earlier in the year in southern rivers and at moderate river discharge levels (relative to northern rivers). For example, spawning occurs in early-February to mid-March in the Savannah River (Hall et al. 1991) and has been documented in late February to late March in the Cooper River (Cooke and Leach, 2004a). Shortnose spawning generally occurs earlier and at lower temperatures than Atlantic sturgeon. Dadswell et al. (1984) report that most shortnose spawn between 9 and 12°C. Physical factors affecting spawning success As observed in the Connecticut River, high river flows during the normal spawning period can cause unacceptably fast bottom water velocities and prevent females from spawning (Buckley and Kynard 1985b; Kynard 1997). Buckley and Kynard (1985b) speculated that the reproductive rhythm of females may be under endogenous control and suitable river conditions must be available or endogenous factors prevent females from spawning. Thus, reproductive success depends on suitable river conditions during the spawning season. Development, growth and movement patterns Habitat and Environmental Requirements Shortnose sturgeon in the South Atlantic portion of the range require the use of large coastal rivers from the estuarine portions to upstream spawning areas. Habitat and environmental requirements of shortnose sturgeon are reviewed in Gilbert (1989). Shortnose require large rivers unobstructed by dams, or in which the dams are above their preferred spawning areas, or at which fish passage has been provided. Shortnose are apparently able to maintain completely freshwater populations, such as in the Santee-Cooper system (Collins et al. 2003). Preferred temperature ranges and upper and lower lethal temperatures for shortnose are not currently known. Shortnose sturgeon are seldom found in shallow water where water temperatures exceed 22°C; however, in the Altamaha they were found at temperatures as high as 34°C. Temperatures at wintering sites ranged from 5-10°C in Winyah Bay. Shortnose sturgeon prefer waters of lower salinity than Atlantic sturgeon. The maximum salinity at which shortnose were found is 30-31 ppt, slightly less than sea water. In areas where shortnose sturgeon and Atlantic sturgeon co-occur, shortnose are typically found in waters less than 3 ppt. Adult sturgeon are typically found in areas with little or no current throughout their lives, especially when they are present in the lower portions of rivers and in the estuaries. Shortnose have been reported from shallower waters in the summer (2-10 m; 6.5-33 ft) and deeper water in the winter (10-30 m; 33-99 ft). They have been observed feeding in heavily vegetated, muddy backwater areas; however, in general submerged aquatic vegetation does not appear to be an important factor in their life history. Early life stages At hatching, shortnose sturgeon larvae are blackish-colored, 7-11 mm (0.3-0.4 in) long and resemble tadpoles (Dadswell et al. 1984). Larvae have a large yolk-sac, poorly developed eyes, mouth and fins, and are capable of only limited swimming. In laboratory experiments, 1 to 8day–old shortnose sturgeon were photonegative, actively sought cover under any available material, and swam along the bottom until cover was found (Richmond and Kynard 1995). It is likely that they hide under available cover at spawning sites. The yolk-sac is absorbed in 9-12 days, and larvae resemble miniature adults by about 20 mm (0.8 in) in length. They likely begin swimming downstream at this size. Larvae collected in the wild were in the deepest waters of the channel. Laboratory studies suggest that there is a two-stage downstream migration: a 2-day migration by larvae, followed by a residency period of young-of-the-year fish, then a resumption of migration by yearlings the second summer of life. Juveniles Juveniles occur in or at the saltwater/freshwater interface in most rivers (Savannah: Hall et al. 1991; Altamaha: Flournoy et al. 1992; Ogeechee River: Weber 1996). Juveniles in the Ogeechee River moved into more saline areas (0-16 ppt) and were most active when water temperature dropped below 16°C (Weber 1996). In the Savannah River, juveniles use sand/mud substrate in depths of 10-14 m (33-46 ft) (Hall et al. 1991). Warm summer temperatures above 28 degrees F may severely limit available juvenile nursery habitat in some southern rivers. Summering habitat in the Altamaha River was limited mainly to one cool, deepwater refuge (Flournoy et al. 1992). Likewise, a similar distribution was observed in the Ogeechee River (Rogers and Weber 1994; Rogers and Weber 1995b, Weber 1996). Adults Adults that occur in freshwater or tidal fresh reaches of rivers in summer and winter often occupy only a few short reaches of the total river length (Savannah River: Hall et al. 1991; Altamaha River: Flouronoy et al. 1992). In the Connecticut and Merrimack Rivers, the “concentration areas” used by fish were reaches where natural or artificial features cause a decrease in river flow, possibly creating suitable substrate conditions for freshwater mussels (Keiffer and Kynard 1993), a major prey item for adult sturgeon (Dadswell et al. 1984). Summer concentration areas in southern rivers are cool, deep, thermal refugia, where adults and juveniles congregate (Flouronoy et al. 1992; Rogers and Weber 1994; Rogers and Weber 1995b; Weber 1996). Growth Growth of juvenile shortnose sturgeon is fast throughout the species‟ range (Dadswell et al. 1984). YOY are 14-30 cm TL after the first year. Fish reach 50 cm after only 2-4 years in the southern part of the range. Fish grow faster in the South, but do not attain the large sizes of northern fish (Dadswell et al. 1984). Survival and Recruitment There is no information on survival of eggs or early life stages in the wild. Many eggs reared in captivity die of fungus infections (Dadswell et al. 1984). Richmond and Kynard (1995) maintain that the availability of spawning substrate with crevices is critical to survival of eggs and embryos. Year class strength of shortnose sturgeon populations is probably established early in life, perhaps in the initial few weeks. Although there is no commercial fishery for shortnose sturgeon (and thus, no fisheries recruitment information), some fisheries incidentally catch adult sturgeon and poaching impacts all populations to an unknown degree. Incidental capture of shortnose sturgeon also occurs in gill net fisheries in the southern portion of the shortnose sturgeon‟s range. Gill net fisheries for American shad (Alosa sapidissima) and trawl fisheries for shrimp (Penaeus spp.) in Georgia and South Carolina captured about 2% of a tagged sample of shortnose sturgeon (Collins et al. 1996). The gill net fishery was responsible for 83% of the total shortnose sturgeon captures. Moser and Ross (1993) reported that 4 of 7 telemetered adult sturgeon in the Cape Fear River were captured in the gill net fishery for American shad or striped bass (Morone saxatilis). In addition, apprehension of poachers operating in South Carolina indicates that illegal directed take of shortnose sturgeon in southern rivers may be a significant source of mortality (D. Cooke, personal communication). Natural mortality Estimates of total instantaneous mortality rates (Z) are available for several river systems, albeit mainly northern populations. Total mortality for the Pee Dee-Winyah River in South Carolina was estimated at 0.08 to 0.12 (Dadswell et al. 1984). This estimate and the estimates for the other populations were based on catch curves which were adjusted for gill net selectivity and effort. Annual egg production Annual egg production is determined by the fecundity of females and the number of spawning females. Estimates of egg production from the Saint John River indicated mean fecundity per female was 94,000 eggs (Dadswell 1979). Monitoring of spawner abundance in the Connecticut River indicated that abundance varies greatly from year to year (Kieffer and Kynard unpublished data). Smith et al. (1992) also suggested that spawner abundance in the Savannah River can fluctuate greatly from year to year. This information indicates that the number of eggs spawned annually varies greatly (possibly by several magnitudes) over the species‟ range and complicates estimation of annual egg production. Migration and Movements Movement patterns in shortnose sturgeon vary with fish size and home river location. Juvenile shortnose sturgeon generally move upstream in spring and summer and move back downstream in fall and winter; however, these movements usually occur in the region above the saltwater/freshwater interface (Dadswell et al. 1984; Hall et al. 1991). Adult shortnose sturgeon are generally estuarine anadromous in southern rivers (Keiffer and Kynard 1993). Spawning migrations are apparently triggered when water temperatures warm above 8°C (Dadswell et al. 1984). Consequently, spring spawning migrations occur earlier in southern systems than in northern ones: January-March (Altamaha River: Gilbert and Heidt 1979, Rogers and Weber 1995a; Savannah River: Hall et al. 1991; Pee-Dee/Waccamaw Rivers: Dadswell et al. 1984; Cape Fear River: Moser and Ross 1993). In the Altamaha River, Rogers and Weber (1995a) also documented upstream movement of most adults to suspected spawning grounds in autumn (late November-early December). A second spawning migration occurred in that system during mid-winter (late January-early February). A shortnose sturgeon spawning migration is characterized by rapid, directed and often extensive upstream movement. Hall et al. (1991) tracked adults during pre-spawning upstream migrations of up to 200 km in the Savannah River and Dadswell et al. (1984) noted that a migration of 193 km occurred in the Altamaha River. Telemetry studies have documented maximum ground speeds of 20-33 km d-1, although mean ground speeds during riverine spawning migrations were around 16 km d-1 (Buckley and Kynard 1985a; Hall et al. 1991; Moser and Ross 1993). Both Hall et al. (1991) and Moser and Ross (1993) observed that spawning migrations are easily interrupted by capture and handling or by dams. Non-spawning movements include rapid, directed post-spawning movements to downstream feeding areas in spring and localized, wandering movements in summer and winter (Dadswell et al. 1984; Buckley and Kynard 1985a; O‟Herron et al. 1993). Shortnose sturgeon usually leave the spawning grounds soon after spawning. Keiffer and Kynard (1993) reported that post-spawning migrations were correlated with increasing spring water temperature and river discharge. Post-spawning migration rates range from 3.5-36 km d-1 (Buckley and Kynard 1985a; Hall et al. 1991; Kieffer and Kynard 1993). During these movements shortnose sturgeon apparently move singly and “home” to very specific sites (Dadswell et al. 1984; Kieffer and Kynard 1993; Savoy and Shake 1992). Continuous tracking of shortnose sturgeon provides detailed information on their migratory behavior. Moser and Ross (1994) demonstrated that, in the Cape Fear River estuary, upstream spawning migration in saltwater was slower (10 km d-1) than migration in freshwater (15 km d1). This was due to the saltatory nature of movement in the estuary and faster swimming (0.8 body lengths (BL) s-1) in freshwater than in the estuary (0.6 BL s-1). Estimated swimming speed during summer, 0.07-0.37 BL s-1, is considerably slower than during spawning migrations (McCleave et al. 1977), while shortnose sturgeon are even less active in winter (Seibel 1993). Moser and Ross (1994) and McCleave et al. (1977) estimated swimming speed to be greatest when sturgeon oriented against rapid ebbing currents. Moser and Ross (1994) and McCleave et al. (1977) reported that shortnose sturgeon do not display any diel activity pattern, traveled in the upper part of the water column (within 2 m of the surface), and that their movement was apparently unaffected by temperature and salinity. Abundance and status of stocks As stated earlier, shortnose sturgeon historically occurred in most rivers of the four South Atlantic states from the Albemarle Sound system in NC through the Indian River system in FL. One relatively recent capture (1998) of a shortnose sturgeon in the Albemarle Sound system has been documented (Wayne Starnes, North Carolina Museum of Natural Sciences, personal communication). There have been no recent documented captures in the Pamlico Sound and its tributaries (Tar, Neuse Rivers). However, there are two recent reports of sturgeon from the Pamlico Sound identified by commercial fishery observers that were allegedly shortnose, but no photographs or tissue were taken for confirmation by professional fishery biologists or geneticists (Wilson Laney, USFWS, personal communication). There is currently a small population of shortnose in the Cape Fear River, North Carolina. At least one population presently exists in the Winyah Bay system (Waccamaw, Pee Dee and Black Rivers), as well as the Santee River, the Cooper River, the ACE Basin (Ashepoo, Combahee and Edisto Rivers) and the Savannah River. Georgia populations occur in the Savannah, Ogeechee, Altamaha, St. Marys and Satilla Rivers. Within the State of Florida, shortnose sturgeon are known only in the St. Johns River. Total population estimates are not available for the Cape Fear, Winyah Bay, Santee, Cooper, ACE Basin, Savannah, Satilla, St. Marys and St. Johns Rivers. For the Ogeechee River, the total population estimate using Modified Schnabel methodology was 361 (95% CI= 326-400) in 1993 and 147 (95% CI= 104-249) in 2000 (Rogers and Weber 1994; Fleming et al. 2003). For the Altamaha River, the total population estimate using Modified Schnabel methodology was 2,862 (95% CI= 1,069-4,226) in 1988, 798 (95% CI= 645-1,045) in 1990, and 468 (95% CI= 316-903) in 1993 (Rogers unpublished data). A more recent study conducted in the Altamaha system from 2004-2006 resulted in a Schnabel population estimate of 6048 (95% CI= 4526-9110) (Peterson unpublished data). Analysis of data for 1996, 1997, and 1998 resulted in estimates of 87 (95% CL 56-170), 193 (95% CL 123-319), and 301 (95% CL 150-659) adult fish, respectively residing in the Pinopolis Dam tailrace area during the spawning season (Cooke and Leach, 2004c). The population in the Cape Fear River is thought to be less than 50 fish (Moser and Ross 1995). Some of these population estimates should be viewed with caution as sampling biases may have violated the assumptions of the abundance models, effort may be limited therefore biasing data, or sampling periodicity may have been inadequate (NMFS 1998). Albemarle Sound/Roanoke and Chowan Rivers A historic record of a shortnose sturgeon in this area, confirmed by a museum specimen, was from Salmon Creek in the lower Chowan River, April, 1881 (USNM 64330, Vladykov and Greeley, 1963). More recently, an individual was caught in the north Batchelor Bay portion of Albemarle Sound (i.e., ca. mouth of Roanoke River) on 18 April 1998. It died in a net, was brought to the North Carolina Museum of Natural Sciences by Jim Armstrong of United States Fish and Wildlife Service, and is vouchered at the museum (NCSM 27062, Wayne Starnes, North Carolina Museum of Natural Sciences, personal communication). Additionally, an unconfirmed record from Oregon Inlet (Holland and Yelverton 1973) was also reported in Gruchy and Parker (1980), Dadswell et al. (1984), and Gilbert (1989). Pamlico Sound/Pamlico and Neuse River Yarrow (1877) reported that shortnose sturgeon were abundant in the North, New, and Neuse Rivers, but these records are doubtful due to their apparent basis in hearsay (Ross et al. 1988). Shortnose sturgeon were also reported from the Beaufort (Jordan 1886) and Neuse Rivers by Fowler (1945). Nearshore records of shortnose sturgeon in this area (Holland and Yelverton 1973) may be misidentifications (Ross et al. 1988). There are two recent reports of sturgeon from the Pamlico Sound identified by commercial fishery observers that were allegedly shortnose, but no photographs or tissue were taken for confirmation by professional fishery biologists or geneticists (Wilson Laney, USFWS, personal communication). Cape Fear River Since the first confirmed capture of shortnose sturgeon in the Cape Fear River (January 1987, Ross et al. 1988), an extensive sampling program has produced eight additional specimens (Moser and Ross 1993). All nine specimens captured were adults; no juveniles were collected. The river is dammed in the coastal plain, a short distance upstream of Wilmington, North Carolina. The river channel near the coast is channelized and heavy industries exist near the port. At least two additional records have been reported from fishermen (Fritz Rohde, North Carolina Division of Marine Fisheries, personal communication). Winyah Bay Drainages Shortnose sturgeon were documented in the Winyah Bay system during the late 1970‟s and early 1980‟s (Dadswell et al. 1984). Fed by the Waccamaw, Pee Dee, and Black Rivers, this coastal plain watershed produced over 100 collections of juveniles and adults during the study period. Within North Carolina, a vouchered specimen was collected from the North Carolina portion of the Pee Dee River in 1984 (Wayne Starnes, North Carolina Museum of Natural Sciences, personal communication), most likely the same individual that was collected below Blewett Falls Dam (Mark Collins, SCDNR-MRD, personal communication). In South Carolina, shortnose sturgeon have been located in the Waccamaw River, although it is possible that they were seasonal migrants from the Pee Dee River. Shortnose sturgeon from the Winyah Bay/Pee Dee River population are able to migrate upstream to the base of Lake Waccamaw (within North Carolina) if so inclined; however, there are currently no evidence/data to support the existence of a separate Waccamaw River shortnose sturgeon population. Shortnose sturgeon spawning has been verified in the Pee Dee River, South Carolina via egg collection (Mark Collins, SCDNRMRD, personal communication). Santee River Seven shortnose sturgeon were recorded from the Santee River drainage in 1978, and one fish was captured in a gillnet in 1992 (Collins and Smith 1997). In addition, 20 specimens were recovered from a fishkill in the Lower Rediversion Canal, St. Stephen Dam tailrace that occurred during a low dissolved oxygen event below the dam. During the period from 1979-1991, shortnose sturgeon were also recorded from Lake Marion, and in the Congaree and Wateree rivers above the dam (Collins and Smith 1997). These fish may represent an essentially landlocked population (Collins et al. 2003). However, during a very high water year, 15 of 16 (94%) Cooper River adult transmittered shortnose sturgeon that were released in Lake Moultrie were able to exit the Santee-Cooper Lakes (Lakes) (1 possible mortality within the Lakes), primarily to the Santee River, although one individual appears to have exited through the lock at Pinopolis Dam on the Cooper River. At least 14 of the 15 (93%) shortnose sturgeon that exited the Lakes are thought to have survived downstream passage (Cooke and Leach 2004b, Cooke and Leach 2004d). Of the 13 shortnose sturgeon that successfully survived downstream passage specifically into the Santee River, all appear to have exited the Santee River system. Eight were relocated in the Winyah Bay system, two in the Cooper River, two in the Intracoastal Waterway, and one disappeared altogether. This behavior appears to be similar to four shortnose sturgeon that were implanted with transmitters in the Santee River. Albeit a small sample size, one was relocated in the Savannah River, the other seemed to disappear from the Santee River only to reappear approximately a year and a half later, and two disappeared altogether (Cooke and Leach 2004d). Cooper River Shortnose sturgeon were documented in what is now the metro Charleston area during the late 1800‟s (Jordan and Evermann 1896). Shortnose sturgeon were collected in this heavily altered (dammed and urbanized) drainage in the 1980‟s during research on the American shad (Alosa sapidissima) fishery. A functionally landlocked segment may exist in Lake Marion (Collins et al. 2003) above the dam that blocks the system in the lower coastal plain (Pinopolis Dam). However, during a very high water year, 15 of 16 (94%) Cooper River adult transmittered shortnose sturgeon that were released in Lake Moultrie were able to exit the Santee-Cooper Lakes (1 possible mortality within the Lakes), primarily to the Santee River, although one individual appears to have exited through the lock at Pinopolis Dam on the Cooper River. At least 14 of the 15 (93%) shortnose sturgeon that exited the Lakes are thought to have survived downstream passage (Cooke and Leach 2004b, Cooke and Leach 2004d). 14 of 15 (93%) Cooper River adult transmittered shortnose sturgeon that were placed in Pinopolis Lock did not pass upstream into the Santee-Cooper Lakes (Lakes), indicating that upstream passage in the Cooper River into the Lakes is extremely limited (Cooke and Leach 2004b). Below Pinopolis Dam, the lowermost dam on the Cooper River, eleven sturgeon were taken in gillnets in the tailrace in February 1995 (Collins et al. 1996). Cooke and Leach (1999) found that shortnose sturgeon congregate and spawn below Pinopolis Dam. Successful egg fertilization and development is occurring, but only one newly hatched yolk sac larva and no juveniles have been documented (Duncan et al. 2004). ). Analysis of data for 1996, 1997, and 1998 resulted in estimates of 87 (95% CL 56-170), 193 (95% CL 123-319), and 301 (95% CL 150-659) adult fish, respectively residing in the Pinopolis Dam tailrace area during the spawning season (Cooke and Leach 2004c). Limited gillnet sampling in 2003 in the spawning area resulted in 8 of the 12 adults (67%) as recaptured fish from previous years (Cooke and Leach 2004a). Ashepoo, Combahee and Edisto Rivers (The “ACE” Basin) The Ashepoo, Combahee, and Edisto drainages form one of the most pristine coastal plain watersheds in the southeastern United States. Shortnose sturgeon were incidentally collected during American shad studies in the Ashepoo and Edisto Rivers in the 1970‟s and early 1980‟s (Collins and Smith 1997). Ripe adults, as well as YOY/age-1 fish, have been more recently collected by the South Carolina Department of Natural Resources-Marine Resources Division (SCDNR-MRD) from the Edisto River. It is possible that these are actually Savannah River fish that were stocked into the Savannah River and then strayed to colonize the Edisto River (Mark Collins, SCDNR-MRD, personal communication). Savannah River The Savannah River is a heavily industrialized and channelized drainage that forms the South Carolina/Georgia border. The river is dammed, but not below the fall line. Shortnose sturgeon were first documented in the system in the mid-1970‟s (Dadswell et al. 1984). During 19841992, over 600 adults were collected by shad fishermen and researchers using gillnets and trammel nets (Collins and Smith 1993). The ratio of adults to juveniles in this study was very high, indicating that recruitment is low in this river (Smith et al. 1992). Adult population estimates were calculated using Jolly Seber (96-1075) and Schnabel (1676) techniques, but were deemed unreliable as not all basic assumptions were met (M. Collins, South Carolina Department of Natural Resources, personal communication). During 1984-1992, approximately 97,000 shortnose sturgeon (19% tagged) of various sizes were stocked in the Savannah River to evaluate the potential for shortnose sturgeon stock enhancement (Smith and Jenkins 1991). Subsequent investigation showed that stocked fish were at large for an average of 416 days and comprised 41% of all juvenile sturgeon collected (Smith et al. 1995). It is thought that the Savannah River population and the Altamaha River population are the two largest populations in the Southeast (Wirgin et al. 2006). Ogeechee River The Ogeechee is primarily a coastal plain drainage with 5% of its watershed in the piedmont. The river is undammed, but water quality has changed (eutrophied) during the last 30 years (Weber 1996). Shortnose sturgeon were first documented in the system during the early 1970‟s (Dadswell et al. 1984). A survey of shortnose sturgeon occurrence, distribution, and abundance, including a 1994-1995 mark/recapture experiment, was conducted from 1993 to 1995 and then from 1999 to 2004 in the tidal portion of the drainage (Rogers and Weber 1994; Weber 1996). The size distribution of shortnose sturgeon sampled indicated that, as in the Cape Fear and Savannah Rivers, the Ogeechee population is dominated by adults. Mark/recapture analysis indicated that abundance is low and possibly declining in the Ogeechee system; the highest point estimate yielded less than 400 individuals from all age classes in 1993 (Weber 1996) and less than 200 individuals from all age classes in 2000 (Fleming et al. 2003). Size frequency, abundance, and catch rate data indicate that natural recruitment does occur in the Ogeechee River system but shortnose sturgeon may be experiencing higher juvenile mortality rates in this system than in the Altamaha (below), with evidence of total recruitment failure in some years. A portion of the shortnose sturgeon that were stocked in the Savannah system have migrated to the Ogeechee River and have contributed substantially to some year-classes in this system (Fleming et al. 2003). Coalescence-based migration estimates suggest that populations south of the Pee Dee River exchange between 1-9.8 individuals per generation, with the highest rate occurring between the Ogeechee and the Altamaha Rivers (Wirgin et al. 2005). Altamaha River The Altamaha River system drains the largest watershed east of the Mississippi River and comprises the confluence of the Ocmulgee and Oconee Rivers plus additional, smaller piedmont and coastal plain drainages. The system is moderately industrialized including two kraft process paper mills and a nuclear generating plant. The watershed landscape has been heavily altered by urbanization, suburban development, agriculture, and silviculture. The system is also dammed, but not below the fall line. Shortnose sturgeon were first documented in the Altamaha in the early 1970‟s (Dadswell et al. 1984), and, later, in a cursory study of spawning movements conducted in the late 1970‟s (Heidt and Gilbert 1979). A two-year study of population structure and dynamics was conducted during the early 1990‟s (Flournoy et al. 1992), building on three additional years of survey data from the late 1980‟s (B. T-A.Woodward, Georgia Department of Natural Resources, unpublished data). Over 650 individuals were collected during the five years of study, with samples heavily dominated by juveniles (90%). Subsequent analysis of tag/recapture data indicated that, during the two-year study period in the 1990‟s, abundance did not exceed 6,055 individuals for all size and age classes. However, under the more rigorous constraints imposed by the assumptions of the recapture model and (probably) met under the conditions experienced during the summer of 1990, the point estimate is 798 individuals with a 95% confidence interval (CI) of 645-1,045 fish. The next time that those conditions were met (during the late summer of 1993), a similar 95% CI of 316-903 individuals was generated with a point estimate of 468 fish. An estimate generated from 1988 data, which met the same criteria, yielded 2,862 fish (95% CI 1,069-4,226). A more recent study conducted in the Altamaha system from 2004-2006 resulted in a Schnabel population estimate of 6048 (95% CI=4526-9110) (Peterson unpublished data). Based on these data, the Altamaha population segment is likely the largest and most viable one south of Cape Hatteras, NC (Joel Fleming, GDNR, personal communication). Similarly, Wirgin et al. (2006) suggested that the Savannah River population and the Altamaha River population are the two largest populations in the Southeast (Wirgin et al. 2006). Satilla and St. Marys The Satilla and St. Marys Rivers are relatively small coastal plain drainages emptying into the Atlantic Ocean between the Altamaha River, GA and St. Johns River, FL. There are no dams and few human impacts beyond agriculture and timber management along the Satilla system. The St. Marys system (draining the eastern portion of the Okefenokee Swamp and forming a portion of the GA/FL border) is likewise undammed, but is heavily channelized in its estuary to support a small port and military installation. The estuary also receives effluents from three major forest product plants. Collections of shortnose sturgeon were made in the estuaries of both systems during the late 1980‟s and early 1990‟s during crustacean monitoring (G. Rogers, Georgia Department of Natural Resources, personal communication). Surveys for sturgeon in the St. Marys (1994 and 1995, 117 net hours) and in the Satilla (1995, 74 net hours) failed to yield any shortnose sturgeon (Rogers and Weber 1995b). St. Johns River The St. Johns River in FL is a heavily altered system flowing northward from the east-central portion of the state and emptying into the Atlantic Ocean near Jacksonville, FL. The system is dammed by Rodman Dam in the headwaters (although currently scheduled for removal), heavily industrialized and channelized near the sea, and affected by urbanization, suburban development, agriculture, and silviculture throughout portions of the basin. Shortnose sturgeon have been reported from the system since 1949 (Kilby et al. 1959). Five shortnose sturgeon were collected in the St. Johns in the late 1970‟s (Dadswell et al. 1984) and, in 1981, three sturgeon were collected and released by the Florida Game and Freshwater Fish Commission. A shortnose sturgeon tagged in Georgia by Georgia Department of Natural Resources near St. Simons Island in March 1996 was captured from the St. Johns River in August 2000 (Jay Holder, FLFWC, personal communication). Most recently, a shortnose sturgeon was collected by Florida Fish and Wildlife Commission personnel in 2002 on the south side of Federal Point near Palatka, Florida during a 2002-2003 study to determine presence/absence of shortnose sturgeon in the St. Johns River. This single shortnose sturgeon was collected in 4,493 hours of 100-m gillnet sets. Preliminary analysis by Joe Quattro at the University of South Carolina indicated that the sample was genetically similar to other shortnose sturgeon populations found in the southern United States (FLFWC 2005). Historically, few shortnose sturgeon in the St. John‟s River have been positively identified by biologists and commercial landings were relatively low compared to other southern states. However, there have been several verified incidental captures off the coast of Daytona Beach (Allan Brown, United States Fish and Wildlife Service, personal communication). No sturgeon reproduction has ever been documented in the St. Johns River and the spawning habitat that is currently accessible seems to be marginal (FLFWC 2005). Ecological relationships Feeding Shortnose sturgeon are benthic omnivores but have also been observed feeding off plant surfaces (Dadswell et al. 1984). Based on the high incidence of non-food items in juvenile shortnose sturgeon, Dadswell et al. (1984) concluded that juveniles randomly vacuum the bottom while adults are more selective feeders. Dadswell (1979) determined that adult shortnose sturgeon in the Saint John River, Canada are not opportunists and only switch to other prey when preferred food are unavailable. The presence of food in the gut during all times of day indicated that shortnose sturgeon are continuous feeders (Dadswell 1979). Shortnose sturgeon feed on crustaceans, insect larvae, worms, and mollusks; however, they apparently undergo ontogenetic shifts in preferred foods. Insect larvae (Hexagenia sp., Chaoborus sp., Chironomous sp.) and small crustaceans (Gammarus sp., Asellus sp., Cyathura polita) predominate in the diet of juveniles (Dadswell et al. 1984; Carlson and Simpson 1987) while adults feed primarily on small mollusks (Dadswell 1984; Hastings 1983). Molluscs ingested by adults captured in freshwater include Physa sp., Heliosoma sp., Corbicula manilensis, Amnicola limnosa, Valvata sp., Pisidium sp., and small Elliptio complanata (Dadswell et al. 1984). In saline areas molluscan prey include small Mya arenaria, and Macoma balthica (Dadswell 1979). Probable foraging activity in southern rivers has been described at the saltwater/freshwater interface during fall and winter in the Pee Dee and Savannah rivers (Dadswell et al. 1984; Hall et al. 1991) and just downstream of the saltwater/freshwater interface in the Altamaha and Ogeechee rivers (Rogers and Weber 1995a; Weber 1996). During summer, shortnose sturgeon in these southern systems appear to reduce activity, fast, and lose weight (Dadswell et al. 1984; Rogers et al. 1994). Predators, Parasites, and Diseases There is very little documentation of predation on any life stage of shortnose sturgeon. Youngof-the-year shortnose sturgeon (approximately 5 cm FL) were found in the stomachs of yellow perch (Perca flavescens) in the Androscoggin River, Maine (Dadswell et al. 1984). It is likely that sharks and seals may occasionally prey on shortnose sturgeon based on the occasional specimen lacking a tail (Dadswell et al. 1984). Parasites recorded from shortnose sturgeon (all data from northern populations) include Coelenterata (Polypodium sp.), Platyhelminthes (Diclybothrium armatum, Spirochis sp., Nitzschia sturionis), Nematoda (Capillospirura pseudoargumentosus), Acanthocephala (Fessesentis friedi, Echinorhynchus attenuatus), Hirudinea (Calliobdella vivida, Piscicola milneri, Piscicola punctata), Arthropoda (Argulus alosa), and Pisces (Petromyzon marinus). The degree of infestation has been reported as being quite low with the exception of Capillospirura sp. (Dadswell et al. 1984). Sturgeon do not appear to be harmed by these parasites. There have been no reported incidences of disease for shortnose sturgeon in the wild, although an epizootic of Columnaris sp. occurred at the FWS‟ Orangeburg Hatchery in South Carolina (Willie Booker, FWS, South Carolina, personal communication). Listing designations 4.3.5 Marine Plants (material from the 2002 Recovery Team Status Review) Description and distribution Description After many years of confusion over identification, Johnson‟s seagrass (Halophila johnsonii) was formally proposed as a separate species by Eiseman and McMillan (1980). Halophila johnsonii was previously referred to either as H. decipiens or H. baillonis Ascherson, but it most closely resembles H. ovalis (R. Brown) Hooker f., an Indo-Pacific species, both morphologically and genetically (McMillan and Williams 1980). Plant classification schemes based on anatomical (den Hartog 1970) and molecular phylogenetic (Les et al. 1997) methods both place the seagrass genus Halophila in the angiosperm family Hydrocharitaceae, along with two other seagrass genera, Thalassia and Enhalus. Morphologically, Johnson‟s seagrass is recognized by the presence of pairs of linearly shaped foliage leaves, each with a petiole formed on the node of a horizontally creeping rhizome (Figure 4.3-2)(NMFS 2001). Figure 4.3-2. Photograph of Halophila johnsonii showing the genet and individual ramets, the rhizome, a female flower, fruit, nodes, and lateral branching of rhizome The rhizome is located at or just below the sediment surface and is anchored to unconsolidated substrate by unbranched roots. The leaves are generally 2-5 cm long (including the petioles), and the rhizome internodes rarely exceed 3-5 cm in length, making this species appear diminutive relative to the larger seagrasses. Halophila johnsonii differs from H. decipiens in a number of morphological, reproductive, and genetic characteristics (Table 4.3-5). The diagnostic characteristics of H. johnsonii remain relatively unchanged when plants are cultured in artificial conditions; thus, differences between the two species are not due to phenoplasticity (NMFS 2001). Table 4.3-5. Morphological, reproductive, and genetic characteristics of H. johnsonii and H. decipiens. Illustrations adapted from Phillips and Menez, 1988. Distribution Johnson‟s seagrass is found only in southeastern Florida from near Sebastian Inlet (27.85°, 80.45°) to Virginia Key (27.74°, -80.14°) (Figure 4.3-3). Recently, however, the St. Johns River Water Management District (SJRWMD) observed H. johnsonii 3 km north of the Sebastian River mouth on the western shore of the lagoon (27.88°, -80.50°) – a discovery that slightly extends the species known northern range. Where it does occur, its distribution is patchy, both spatially and temporally (Virnstein and Morris 2007). Halophila johnsonii is a perennial species showing no consistent seasonal or year-to-year pattern in surveys of the Indian River Lagoon (IRL). Although perennial, it exhibited some winter decline. However, during exceptionally mild winters, as in the winter of 2004, H. johnsonii can maintain or even increase its abundance from summer to winter (Virnstein et al. 1997, Virnstein and Morris 2007). Depth of occurrence ranged from 0.03 to 2.5 m within transects monitored in the IRL (Virnstein et al. 1997, Virnstein and Morris 2007). When data from all transects were combined, there was no correlation of H. johnsonii abundance with depth. However, the deep edge at some transects was only 0.1 m; at other transects, it was 2.5 m. When all depths of occurrence were standardized (as percent of maximum depth of a transect), H. johnsonii was more abundant in the deeper parts of the transects. Most (78% or 574 out of 733) occurrences of H. johnsonii were at >70% of maximum transect depth; half were at >90% of maximum depth (Virnstein et al. 1997, Virnstein and Morris 2007). Figure 4.3-3. Geographic range of Halophila johnsonii: Sebastian Inlet to northern Virginia Key (Kenworthy 1997). Observations of its distribution and the results of limited experimental work suggest that H. johnsonii has a wider tolerance range for salinity, temperature, and optical water quality conditions than H. decipiens (Dawes et al. 1989; Gallegos and Kenworthy 1996; Durako et al. 2003; Kunzelman et al. 2005; Torquemada et al. 2005). Halophila johnsonii has been observed growing perennially near the mouths of freshwater discharge canals (Gallegos and Kenworthy 1996), in deeper turbid waters of the interior portion of the Indian River Lagoon (Kenworthy 2000; Virnstein and Morris 2007), and in clear water associated with the high energy environments and flood deltas inside ocean inlets (Kenworthy 1993, 1997; Virnstein et al. 1997; Heidelbaugh et al. 2000; Virnstein and Morris 2007). While H. johnsonii is negatively affected by both extreme hypo- and hyper-salinity conditions, it does tolerate hypersaline conditions better than hyposaline conditions. Most other seagrasses, conversely, are thought to be more sensitive to increased salinity (Ogata and Matsui 1965; Biebl and McRoy 1971; Zieman 1975; Adams and Bate 1994; Doering and Chamberlain 1998). Torquemada et al. (2005) and Dawes et al. (1989) concluded that H. johnsonii could be seriously affected by salinity variations produced by human activities, such as freshwater discharges through water management practices or brine discharges from seawater desalination plants. Interestingly, salinity changes do not seem to alter the tolerance of this species to other environmental factors, such as temperature or pH (Torquemada et al. 2005). Reproduction Like all other seagrasses, H. johnsonii is clonal, which refers to plants that have many semiindependent units (ramets) acting together as a single organism (Cook 1983). Reproduction is achieved primarily by asexual means. While all other species of seagrass reproduce sexually, there is still no evidence of sexual reproduction in H. johnsonii. All attempts to find seeds and seedlings have failed to detect any evidence of their occurrence (Jewitt-Smith et al. 1997; Hammerstom and Kenworthy 2003). Likewise, despite widespread sampling and surveys throughout the entire range of the species, no male flowers have ever been reported and confirmed. Female flowers, however, have been documented in both culture and nature (Eiseman and McMillan 1980; Heidelbaugh et al. 2000). They are common and often very abundant (Heidelbaugh et al. 2000). They have been observed throughout the entire range of the species during all times of the year, but no consistent patterns of spatial or temporal distribution have been observed or reported. Although male flowers have never been observed, it is not possible to completely rule out their existence and the potential for sexual reproduction. They may occur cryptically in isolation or in the vicinity of females. They may be extremely rare, or they may express themselves only at night, as was the case in a related species, Halophila hawaiiana (Herbert 1986). While sexual reproduction of H. johnsonii remains somewhat of a mystery, reproduction by asexual means and clonal growth is well understood. Asexual reproduction occurs when rhizome apical meristems divide and form new leaf pairs, flowers, or rhizome apices (Posluszny and Tomlinson 1990) (Figure 4.3-2). The divisions and subsequent differentiation of meristems (meristem dependence) into the various attributes of the ramets are the foundation of growth and productivity in all seagrasses (Tomlinson 1974). Halophila johnsonii grows by division of apical meristems on horizontal rhizomes which branch, forming leaf pairs, female flowers and new lateral branches (Figure 4.3-4). On average, new meristems are formed on rhizomes every 2 to 4 days (Kenworthy 1997, Bolen 1997) and meristem densities can reach hundreds to thousands per square meter (Kenworthy 1997, Heidelbaugh et al. 2000). Figure 4.3-4. Johnson‟s seagrass, Halophila johnsonii. Leaves are generally 2-5 cm long. Adapted from Eiseman and McMillan (1980). Development and growth The species spreads as clones expand in local space by rhizome extension and leaf pair formation, eventually forming high density “patches.” Rhizomes can elongate at rates approaching 0.5 cm per day (Bolen 1997, Kenworthy 1997), and when combined with prolific branching, individual patches (clones) can expand at extraordinary rates, ranging from 0.3 to 1.0 m-2 per month (Kenworthy 1997, 2003; Greening and Holland 2003). This expansion can lead to coalescence of adjacent patches and the formation of larger meadows. Widely spaced patches, usually on the order of 1-20 square meters in size, are the most commonly encountered feature of H. johnsonii meadows (Virnstein et al. 1997, Kenworthy 1997, 2000; Virnstein and Morris 2007; Kenworthy 2003). Although it is more commonly found in monotypic patches, H. johnsonii can also grow among low to moderate densities of Halodule wrightii and Syringodium filiforme, and in deeper water mixed with H. decipiens (Kenworthy 1993, 1997, 2000; Virnstein et al. 1997, Virnstein and Morris 2007). Patches can also disappear rapidly. Sometimes they will disappear for several years and then reestablish: a process referred to as “pulsating patches” (Heidelbaugh et al. 2000, Virnstein and Morris 2007). Mortality, or the disappearance of patches, can be caused by a number of processes, including burial from bioturbation and sediment deposition, erosion, herbivory, desiccation, and turbidity. Halophila johnsonii‟s canopy is only 2-5 cm tall and may be easily covered by sediments transported during storms or redistributed by macrofaunal bioturbation during the feeding activities of benthic organisms (Heidelbaugh et al. 2000). In the absence of sexual reproduction, one possible explanation for the pulsating patches is dispersal and re-establishment of vegetative fragments, a process which commonly occurs in aquatic plants and has been demonstrated in other seagrasses (DiCarlo et al.; 2005). While H. johnsonii is vulnerable to uprooting by wind waves, storm events, tidal currents, bioturbation, and motor vessels, these are also mechanisms capable of disturbing patches and creating clonal fragments for dispersal. Hall et al. (2006) showed that drifting fragments of H. johnsonii can remain viable for 4 to 8 days, after which time they can settle, root, and grow. Ecological relationships Seagrasses have recently received increasing attention from scientists and managers because of the valuable functional roles they play in coastal ecosystems (Costanza et al. 1997; Larkum et al. 2006). Seagrass beds are one of the primary nursery habitats because of their abundance of prey items as well as the protection they provide from predators (Zieman and Zieman 1989; Heck et al. 2003). Other functions associated with seagrasses include nutrient recycling, detrital production and export, and sediment stabilization. Very little work has been done on the functional value of H. johnsonii, therefore, the functional roles of its closest relative, H. ovalis, and other Halophila spp. are used a proxy in this discussion. The most well-known function of seagrasses is their role as habitat for numerous fishes and invertebrates. Some species spend their entire lives within seagrass beds and others utilize it only during certain stages of their life cycle (usually the postlarval and juvenile stages). Heidelbaugh (1999) conducted one of the only studies that examined benthic fauna associated with H. johnsonii. In this study, differences in benthic fauna among H. johnsonii, H. wrightii, and bare sand were compared on the flood tidal delta just inside Sebastian Inlet, Fl. Halophila johnsonii beds yielded a total of 126 species (69 epifauna and 57 infauna), while 117 species were collected from H. wrightii beds and 99 species from bare sand. The most abundant infaunal organisms belonged to Nematoda while the most abundant epifaunal species were amphipods and tanaids. The majority of macrofaunal organisms consisted of decapod crustaceans (Callinectes sapidus), fishes (Eucinostomus sp.), and some gastropods (especially Bursatella leachii). Three hundred and twenty macrofaunal organisms were collected from H. johnsonii beds compared to 690 from H. wrightii beds and 78 from bare sand. Rapid growth, high turnover rates, and labile tissues make Halophila spp. a good source of nutrition for several marine herbivores (Kenworthy et al. 1989, Lanyon 1991, Preen 1995, Bolen 1997). The Florida manatee (Trichechus manatus latirostris) has been observed grazing on H. johnsonii near a power plant in Palm Beach, Florida (J. Reid, Sirennia Project, U.S.G.S., Gainesville, FL, personal observation). Green turtles (Chelonia mydas) are known to eat several species of Halophila (Hasbun et al. 2000; Kannan and Rajagopalan 2004; Russell et al. 2003; Whiting and Miller 1998). Halophila species also provides nutrition for herbivorous fish. Through consumption, the stareye parrotfish (Calotomus carolinus) has the ability to control the abundance and distribution of short lived seagrass species such as H. stipulacea in Kenya (Mariani and Alcoverro 1999). Even invertebrates such as the queen conch (Strombus gigas) (Thayer et al. 1984) and various species of harpacticoid copepods (Shimode and Shirayama 2006) have been observed feeding on Halophila species. Seagrasses play an important role in nutrient cycling within systems and can act as both a source and sink for nutrients. Processes that lead to a loss of nutrients from the system include: exudation/leaching from living and dead plant material, export of sloughed leaves and leaf fragments, nutrient transfer by foraging animals, denitrification, and diffusion from sediment. Processes that result in an increase of nutrients include: nitrogen-fixation, sedimentation, and nutrient uptake by leaves. It is the fluctuation of these processes that leads to interannual variations in net losses or net gains of nutrients, and therefore, fluctuations in the productivity of seagrass meadows (Hemminga et al., 1991). Bacteria mediate the recycling of nutrients and may be important in regulating the flow of energy from seagrass detritus to consumer organisms (Robertson et al. 1982). Studies in the Salt River Submarine Canyon at St. Croix, US Virgin Islands show that H. decipiens is an important source of organic matter and detritus for the Canyon (Josselyn et al. 1983; Josselyn et al. 1986; Kenworthy et al. 1989). Despite its production being less than other seagrasses, H. decipiens has a fast turnover time and is a major source of primary production on the floor of the Canyon (Kenworthy et al. 1989). Disturbance and burial of plant material are important mechanisms influencing the disposition of organic matter (Williams et al. 1985; Josselyn et al. 1986). Burial of H. decipiens through wave action and animal activities increases the rate of detrital input and retains the detritus within the Canyon (Kenworthy et al. 1989). Seagrasses have long been recognized for their ability to stabilize sediments. It was once assumed, however, that due to its small size and sparse biomass, Halophila spp. were not capable of stabilization (den Hartog 1970). Fonseca (1989) proved this assumption incorrect using a surface-supplied, inverted seawater flume. He found the cumulative effect of H. decipiens in reducing sediment erosion was significantly greater than adjacent, unvegetated sand. Abundance and status Johnson‟s seagrass has only relatively recently been identified as a distinct species (Eiseman and McMillan 1980) and no historical information on the species‟ distribution is available. However, since 1994, the St. Johns River Water Management District has monitored 73 permanent transects in the Indian River Lagoon (IRL) in both summer (June-July) and winter (JanuaryFebruary) (Virnstein et al. 1997, Virnstein and Morris 2007). Despite extensive ground-truthing since 1986 and monitoring all 73 transects throughout the IRL beginning in the summer of 1994 (a total of about 25,000 quadrats), H. johnsonii has never been found more than 3 km north of the Sebastian Inlet area. Where it does occur, its distribution is patchy, both spatially and temporally. Halophila jonsonii is a perennial plant with no strong seasonal pattern, although it generally exhibits some winter decline. Monitoring in the IRL indicates that there is spatial and temporal variation in the abundance of H. johnsonii patches (Virnstein et al. 1997). Although the monitoring data are limited, no large distributional gaps have been detected in the IRL, and there has been no overall increase or decrease in abundance or geographic range over the period from summer 1994 to summer 1999. Halophila. johnsonii was listed as threatened under the Endangered Species Act on September 14, 1998 (63 FR 49035). NMFS concluded that Johnson‟s seagrass is rare, has a limited reproductive capacity, and is vulnerable to a number of anthropogenic and natural disturbances. It also exhibits the most limited geographic distribution of any seagrass. Within its small geographic range (lagoons on the east coast of Florida from Sebastian Inlet to central Biscayne Bay), it is one of the least abundant species. Because of its limited reproductive capacity and energy storage capacity, it is less likely to survive environmental perturbations and to be able to repopulate an area when lost. Finally, environmental degradation and habitat loss have continued despite existing federal and state conservation efforts. Completion of a report reviewing the current status of H. johnsonii is anticipated in the fall of 2007. Critical Habitat Ten areas in the geographic range of Johnson‟s seagrass were designated as critical habitat on April 5, 2000 (65 FR 17768). These areas and their approximate acreage include: a portion of the Indian River Lagoon, north of the Sebastian Inlet Channel (5.7); a portion of the Indian River Lagoon, south of the Sebastian Inlet Channel (2.0); a portion of the Indian River Lagoon near the Fort Pierce Inlet (4.3); a portion of the Indian River Lagoon, north of the St. Lucie Inlet (2770); a portion of Hobe Sound (900); a site on the south side of Jupiter Inlet (4.3); a site in central Lake Worth Lagoon (15.0); a site in Lake Worth Lagoon, Boynton Beach (95.5); a site in Lake Wyman, Boca Raton (20.0); and a portion of Biscayne Bay (18,757). This designated area accounts for approximately 22,574 acres or 9,139 hectares. 4.3.6 Marine Invertebrates (all information below from Acropora Status review) Description All Atlantic Acropora spp. are considered to be environmentally sensitive, requiring relatively clear, well-circulated water (Jaap et al. 1989). Atlantic Acropora spp. are almost entirely dependent upon sunlight for nourishment compared to massive, boulder-shaped species in the region (Porter 1976; Lewis 1977), with these latter types of corals more dependent on zooplankton. Thus, Atlantic Acropora spp. are much more susceptible to increases in water turbidity than some other coral species. Dredging or pollution activities that reduce long-term water clarity can also reduce the coral photosynthetic to respiration ratio (P/R ratio) below unity. Therefore, Acropora spp. may not be able to compensate with an alternate food source, such as zooplankton and suspended particulate matter, like other corals. Optimal water temperatures for A. palmata range from 25 to 29°C, although colonies in the U.S.V.I. have been known to tolerate short-term temperatures around 30°C without obvious bleaching (loss of zooxanthellae). Jaap (1979) and Roberts et al. (1982) note an upper temperature tolerance of 35.8°C for A. palmata. All Atlantic acroporids are susceptible to bleaching due to adverse environmental conditions (Ghiold and Smith 1990; Williams and Bunkley-Williams 1990). Major mortality of A. palmata and A. cervicornis occurred in the Dry Tortugas, Florida, in 1977 due to a winter cold front that depressed surface water temperatures to 14 to 16°C. Some reduction in growth rates of A. cervicornis was reported in Florida when temperatures dropped to less than 26°C (Shinn 1966). All Acropora spp. require near oceanic salinities (34 to 37 ppt). Staghorn Coral Historically, A. cervicornis (Figure 4.3-5) was reported from depths ranging from <1 to 60 m (Goreau and Goreau 1973). It is believed that 60 m is an extreme situation and that the coral is relatively rare below 20 m depth. The common depth range is currently observed at 5 to 15 m. In southeastern Florida, this species historically occurred on the outer reef platform (16 to 20 m) (Goldberg 1973), on spur and groove bank reefs and transitional reefs (Jaap 1984; Wheaton and Jaap 1988), and on octocoral-dominated hardbottom (Davis 1982). In the Florida Keys A. cervicornis can occur from 1 to 34 m depths (Wells 1933; Davis 1982; Jaap 1984; Jaap and Wheaton 1988; Jaap et al. 1989). Colonies may also be common in back- and patch-reef habitats (Gilmore and Hall 1976; Cairns 1982). Although A. cervicornis colonies are sometimes found interspersed among colonies of A. palmata, they are generally in more protected, deeper water or seaward of the A. palmata zone and hence, protected from waves. Historically, A. cervicornis was the primary constructor of mid-depth (10 to 15 m) reef terraces in the western Caribbean, including Jamaica, the Cayman Islands, Belize, and some reefs along the eastern Yucatan peninsula (Adey 1978). Figure 4.3-5a. Staghorn coral, Acropora cervicornis (Source: Walt Jaap). As depth increases, A. cervicornis colonies tend to be less compacted, have longer branches, and branching tends to be at greater intervals. Gladfelter (1982) demonstrated that infilling occurs as the branch elongates. Thus, at the tip, the porosity of the axial calyx is >90% and the wall is 60%, while at 60 cm from the tip, the porosity of the axial calyx is dead and the porosity of the wall is about 20%. This strengthens the branch as it elongates and the momentum of the branch increases. At depths of 20 to 40 m, where currents and wave force are minimal, branch diameter is thinner, being approximately half the diameter of a colony in the shallow surge zone. The porosity of the skeletons of A. cervicornis ranges from 35 to 65% by volume, with the mechanical strength of the skeleton proportional to the porosity (Schumacher and Plewka 1981). Because the skeleton is quite porous, it breaks readily in strong wave forces. Elkhorn Coral The maximum range in depth reported for A. palmata Figure 4.3-5b is <1 m to 30 m, but the optimal depth range for this coral is considered to be 1 to 5 m depth (Goreau and Wells 1967). Figure 4.3-5b. Elkhorn coral, Acropora palmata (Source: W. Jaap). Currently, the deepest known colonies of A. palmata occur at 21 m in the Flower Garden Banks National Marine Sanctuary (Hickerson, pers. comm.) and at Navassa National Wildlife Refuge (Miller, pers. comm.). The preferred habitat of A. palmata is the seaward face of a reef (turbulent shallow water), including the reef crest, and shallow spur and groove zone (Shinn 1963; Cairns 1982; Rogers et al. 1982). At low tide, colonies are sometimes exposed. Colonies of A. palmata often grow in nearly mono-specific, dense stands and form interlocking framework known as thickets in fringing and barrier reefs (Jaap 1984; Tomascik and Sander 1987; Wheaton and Jaap 1988). Storm-generated fragments are often found occupying back reef areas immediately landward of the reef flat/reef crest, while colonies are rare on lagoonal patch reefs (Dunne 1979). Acropora palmata formed extensive barrier-reef structures in Belize (Cairns 1982), the greater and lesser Corn Islands, Nicaragua (Gladfelter 1982; Lighty et al. 1982), and Roatan, Honduras, and built extensive fringing reef structures throughout much of the Caribbean (Adey 1978). Colonies generally do not form a thicket below 5 m depth, with maximum water depths of framework construction ranging from 3 m to 12 m (see Table 1 in Lighty et al. 1982). Distribution and Abundance When discussing historic distribution and abundance, it is important to briefly mention the environmental setting of the wider Caribbean region (tropical western Atlantic, CaribbeanAtlantic province), insofar as environmental differences across the region influence the extent to which Acropora spp. have been able to build extensive reef structures. Specifically, although both A. cervicornis and A. palmtata are found throughout the Caribbean Sea, their historical abundance patterns are not necessarily similar and there is ample evidence to suggest that many reef systems were constructed without significant contributions by acroporids. Early reviews of western Atlantic reefs and coral species, as well as discussions of reef geomorphology in the western Atlantic, are provided elsewhere (e.g., Glynn 1973, Milliman 1973, Adey 1977; 1978), but provide context to the historical patterns of these corals. The entire Caribbean-Atlantic province is characterized as microtidal and is impacted by largely unidirectional trade winds and waves subject, in part, to strong ocean flows. The most northern reefs in the province (i.e. Florida, northwestern Bahamas, and Bermuda) are cyclically stressed by the occasional effect of polar air during winter months and thus have limited reef development by Acropora spp. or lack these species altogether (Bermuda). Throughout the Caribbean, wave energy influences the degree to which crustose coralline algae and Acropora spp. dominate as reef-building elements (Adey 1977; Geister 1977). For example, large swells from the Atlantic Ocean limit acroporid reef development in the Windward Islands (eastern Caribbean) and the eastern flanks of the Bahamas (Roberts et al. 1992). In the Lesser Antilles, neither A. cervicornis nor A. palmata are significant agents of reef framework construction, due principally to higher wind strength, easterly consistencies, and longer fetch; this area is also subjected to longperiod swells or rollers during the winter months that further limits shallow and middepth reef construction (Adey 1977). In the southwestern Caribbean (e.g., Panama), reef terraces are present that are potentially conducive to acroporid-reef development, but seasonally rough seas batter the area resulting in wave-swept pavements (Glynn 1973). In contrast, the northwestern Caribbean (e.g. Cuba, Cayman Islands, Jamaica, eastern Yucatan, Belize) is characterized by relatively low winds of medium to high easterly consistencies, that allows extensive acroporid growth at shallow and mid-depth (10 to 25 m). For example, the Belize Barrier Reef, the largest barrier reef in the province, appears to be based upon an A. cervicornis framework (Adey 1977). The current range for both A. cervicornis and A. palmata remains unchanged from the historical (Figure 4.3-6) as far as data are available; there is a paucity of quantitative data for many locations throughout the wider Caribbean. Historically most data collected has been from a few specific reef sites that may or may not represent the regional condition of the acroporids or coral reefs in general. In contrast, there are many qualitative data/observations indicating drastic declines in abundance of both A. palmata and A. cervicornis throughout their geographic range (e.g., Aronson and Precht 2001a). Figure 4.3-6. Approximate range of Acropora spp. (highlighted), including the Gulf of Mexico, Atlantic Ocean and Caribbean Sea. The highlighted areas are not specific locations of the corals, rather reflect general distribution. Recently, there have been two publications that have summarized status (abundance and distribution) of A. cervicornis and A. palmata. The Status of Coral Reefs in the western Atlantic: Results of initial Surveys, Atlantic and Gulf Rapid Reef Assessment (AGRRA) Program (Lang 2003) provides results (1997–2004) of a regional systematic survey of corals, including Acropora spp., from many locations throughout the Caribbean. While data from this survey represent a snapshot in time of the reef, the geographic scope of the survey is great; targeted areas are visited by data collectors of varying expertise, and data represent a single survey. AGRRA data (1997-2004) indicate that the historic range of both species remains intact, that A. cervicornis is rarely found throughout the range (including areas of previously known occurrence) and a moderate occurrence of A. palmata. An AGRRA bio-area index for A. palmata was recently developed to summarize data for nearly 300 sites throughout the wider Caribbean (Garza-Perez and Ginsburg pers. comm.). This bioarea index utilizes maximum diameter and partial mortality values of A. palmata colonies per site (total area of living tissue/10 transects) and is presented in Figures 4.3-7a and 4.3-7b. Results from the spatial analysis are as follows: 1. most (n=61) bio-areas (Figure 4.3-7a) ranked as moderate to high (100 to 500 m2/10 transects) are concentrated in Andros Barrier Reef (Bahamas) and the northern Caribbean (Cuba and Belize); 2. 195 sites (Figure 4.3-7b) distributed throughout the geographic area were ranked as low bio-areas (from 0.01 to 100 m2/10 transects) 3. standing dead colonies of A. palmata were found throughout the geographic range (Figure 4.3-7b). It is important to note that the data for the Andros Barrier Reef AGRRA surveys were conducted prior to the Caribbean-wide 1998 coral die-off and the site has not been resurveyed since 1997. Furthermore, status of A. palmata has not been updated following the 2004 hurricane season where Hurricanes Charley passed over Cuba, and Hurricanes Frances and Jeanne passed over the Bahamas. Figure 4.3-7a. Locations of reefs indexed with moderate or high (circles) Acropora palmata bio-area as reported from 1997- 2004 AGRRA surveys. Map provided courtesy Garza-Perez and Ginsburg. Figure 4.3-7b. Locations of reefs indexed with low (flag) Acropora palmata bio-areas as reported from 1997-2004 AGRRA surveys. Locations of standing-dead A. palmata colonies are indicated by a cross. Map provided courtesy Garza-Perez and Ginsburg. Prior to the AGRRA summary, Bruckner (2002) provided a comprehensive summary of the bestknown quantitative and qualitative data on the status of the Atlantic acroporids resulting from a NOAA-sponsored workshop wherein participants compiled data and summarized conditions throughout the range. Much of the data from the Bruckner (2002) report are summarized below, and some are updated and included in the case studies at the end of this section. Abundance and distribution (historic and current) of Acropora cervicornis Historically, A. cervicornis so dominated the reef within the 7 to 15 m depth that the area became known as the staghorn zone (Figure 4.3-8) throughout much of the Caribbean. In many other reef systems in the wider Caribbean, most notably the western Caribbean areas of Jamaica, Cayman Islands, Belize and eastern Yucatan (Adey 1977), A. cervicornis was a major mid-depth (10 to 25 m) reef-builder. Principally due to wind conditions and rough seas, A. cervicornis has not been known to build extensive reef structures in the Lesser Antilles and southwestern Caribbean. Acropora cervicornis has also been documented in deeper water (16 to 30 m) (Goldberg 1973) and as far north as Palm Beach, Florida (26° 3‟N). It is also distributed further south and west throughout the coral and hardbottom habitats of the Florida Keys (Antonius et al. 1978; Burns 1985; Dustan 1988; Dustan and Halas 1987; Glynn et al. 1989; Jaap 1984; Jaap and Wheaton 1975; Jaap et al. 1988; Wheaton and Jaap 1988), and Dry Tortugas (Vaughan 1915; Davis 1982; Dustan 1985, 1988; Jaap et al. 1989). In Biscayne National Park (upper Florida Keys), A. cervicornis was more abundant on reefs further from tidal passes (e.g., Ajax and Long Reefs) than those nearby, with historical coverage ranging from 0.1% to 2.7% in the 1980s (Burns 1985). Because Florida is one of the few areas where multi-year quantitative data are available for A. cervicornis at more than a single location (Carysfort Reef; Dustan and Halas 1987, Looe Key Reef; Wheaton and Jaap 1988, Dry Tortugas; Davis 1982 and Porter et al. 1982) (Figure 4.3-9), those data are further analyzed and presented as a case study in section 4.5.3 of the Acropora Status Review document. Figure 4.3-8. Reef zonation schematic example modified from several reef zonation-descriptive studies (Goreau 1959; Kinzie 1973; Bak 1977). Figure 4.3-9. Map of the Florida Keys National Marine Sanctuary and reefs of the Florida Keys. Historical and current distribution and abundance of Acropora palmata Throughout much of the Caribbean, A. palmata historically occupied the 1 to 8 m depth range (reef flat, wave zone, reef crest) known as the „elkhorn zone‟ (Figure 4.3-8). Acropora palmata occupied this zone in Jamaica (Goreau 1959), Alacran Reef, Yucatan peninsula (Kornicker and Boyd 1962), Abaco Island, Bahamas (Storr 1964), the southwestern Gulf of Mexico, Bonaire (Scatterday 1974), and the Florida Keys (Jaap 1984; Dustan and Halas 1987). The predominance of A. palmata in shallow reef zones is related to the degree of wave energy. In areas with strong wave energy conditions only isolated colonies may occur, while thickets may develop in areas of intermediate wave energy (Geister 1977). Although considered a turbulent water species, A. palmata is sensitive to breakage by wave action, and is often replaced by coralline algae in heavy surf zones throughout the province (Adey 1977). While A. cervicornis has been documented further north along the Florida east coast, the northern extension of A. palmata is at Fowey Rocks offshore the Miami area (25° 37‟ N) (Porter 1987). This area technically begins the Florida Reef Tract where all of the major reefbuilding corals appear in shallow water in the southeastern U.S. (Burns 1985). Surveys in the early 1970s north of Miami (e.g., Palm Beach) did not note the occurrence of A. palmata (Goldberg 1973). Between Fowey Rocks and Carysfort Reef, A. palmata has been historically rare as the significant reef development or framework construction by A. palmata begins further south at Carysfort Reef (25° 20‟N), extending discontinuously southwestward to the Dry Tortugas (Jaap 1984). Notably, recent surveys have reported a few colonies of A. palmata off Pompano Beach, Broward County, FL (Figure 4.3-10). The status of these few northern-most colonies presently is unknown. Figure 4.3-10. Acropora palmata off of Pompano Beach, Broward County, Florida, in 2003; (Photo credit: J. Sprung). Distribution records of A. palmata for the southeast Florida coast include: upper Florida Keys (Burns 1985; Dustan 1985, 1988; Dustan and Halas 1987; Jaap et al. 1988), lower Florida Keys (Jaap and Wheaton 1975; Antonius et al. 1978; Jaap 1979; Wheaton and Jaap 1988), and Dry Tortugas (Davis 1982; Jaap et al. 1989). Offshore reefs built primarily by A. palmata are situated along the outer margin of an arc-shaped limestone plateau (south Florida shelf). “Flourishing” A. palmata reefs (i.e., those with a shallow or emergent reef flat) are limited to the northern seaward half of Key Largo where an Acropora zone (reef flat/reef crest) was present. At Molasses Reef (see Figure 4.3-9) living A. palmata was almost absent in 195960 (Shinn 1963) and it was later suggested that conditions necessary for the growth of this coral changed since the coralline spurs were originally accreted (Shinn et al. 1981). Numerous other studies describing A. palmata abundance and distribution in the Florida Keys are available. When possible, data (e.g., Chiappone and Sullivan 1997) were further analyzed and presented as a case study in section 4.5.3 of the Acropora Status Review document. Other data from short-term projects throughout the Florida Keys are summarized below: From 1984 to 1991, a decline in A. palmata abundance at shallow depths (4 to 6 m) but not deeper were noted in coral communities on six reefs (including sites in both the upper Keys and lower Keys) (Porter and Meier 1992). These changes were attributed to disease and the demise of the long-spined sea urchin (Diadema antillarum). Living and dead assemblages of corals on two offshore sites and two patch reefs were compared and significant differences in taxonomic composition between live and dead coral assemblages were found between reef types (Greenstein and Pandolfi 1997). While both the patch and offshore reefs historically had more A. cervicornis and A. palmata, they were now dominated by other corals (Porites astreoides and Siderastrea siderea). Interestingly, massive growth forms were under-represented in the dead assemblage, while branching growth forms (Acropora spp.) were underrepresented in the live coral assemblage. Reproduction The distribution and abundance of Atlantic Acropora spp., like other coral species, reflects patterns of larval recruitment, asexual reproduction via fragmentation, mortality, regenerative capabilities, and aggressive interactions (Richmond and Hunter 1990). Interspecific differences in the mechanisms of recruitment, dispersal, and mortality are likely important in determining the species composition of reef corals in different environments. These differences reflect the differential allocation of energy to the basic life history functions of growth (rate and rigidity of the skeleton), reproduction (fecundity, mode of larval dispersal, recruitment success), and colony maintenance (intra- and interspecific interactions, competitive ability, regeneration) (Connell 1973; Lang 1973; Bak and Engel 1979; Szmant 1986). Populations of Atlantic Acropora spp. are dependent upon sexual recruits for recovery after catastrophic disturbance, but can locally dominate hardbottom and coral reef habitats when colonies fragment and propagate across the bottom. Extensive research has been conducted on the diverse reproductive strategies employed by scleractinian corals (Fadlallah 1983; Szmant 1986; Richmond and Hunter 1990). Atlantic Acropora spp., like many stony coral species, employ both sexual and asexual reproductive propagation. Sexual reproduction in corals includes gametogenesis (i.e., development of gametes) within the polyps near the base of the mesenteries. Some coral species have separate sexes, while others such as the Atlantic Acropora spp., are hermaphroditic. Acropora cervicornis and A. palmata in particular do not differ substantially in their sexual reproductive biology. Both species are spawners, meaning that coral larvae develop externally to the parental colonies (Szmant 1986) and both species are simultaneous hermaphrodites, meaning that a given colony will contain both female and male reproductive parts during the spawning season. Gametes (eggs and sperm) are located in different mesenteries of the same polyp (Soong 1991). The development period is longer for eggs than sperm, lasting approximately 10 months (Szmant 1986). The spawning season for A. cervicornis and A. palmata is relatively short; with gametes released only a few nights during July, August, and/or September. In some populations, spawning is synchronous after the full moon during any of these three months. Annual egg production in A. cervicornis and A. palmata populations studied in Puerto Rico was estimated to be 600 to 800 eggs per cm2 of living coral tissue (Szmant 1986). Eggs from both corals are ~300 μm in diameter. Colonies of A. cervicornis studied on the Caribbean coast of Panama during 1987-88 produced eggs 0.3 to 1.0 mm in length along the long axis that were elliptical in shape (Soong 1991). Spermaries were present during July and August and not during other times of the year. In the same study, A. palmata eggs were 0.2 to 1.0 mm in length along the long axis and shaped as irregular ellipses. Spermaries were present during July, August, and September. In a subsequent study, Soong and Lang (1992) observed that large axial polyps and basal tissues (1.0 to 4.5 cm from the colony base) in A. cervicornis were infertile, whereas gonads located within 2 to 6 cm of the branch tips always had smaller eggs than those in the mid-region of the branches. In A. palmata, small eggs were found in the whole colony, while infertile areas were observed in the encrusting base and along the growing edges of branches (Soong and Lang 1992). Upper surfaces of A. palmata colonies had significantly greater fecundity (more fertile polyps per unit surface area) and larger numbers of eggs within fertile polyps. Colonies of A. cervicornis and A. palmata studied on the Caribbean coast of Panama indicated that larger colonies of both species (as measured by surface area of the live colony) have higher fertility rates (Soong and Lang 1992). For A. palmata, no colonies with a surface area between 4 and 15 cm2 (n=4) or between 15 to 60 cm2 (n=9) were fertile, while 7% of those 60 to 250 cm2 in tissue surface area were fertile (n=14). Over 30% of the colonies between 250 and 1000 cm2 in tissue surface area were fertile (n=16), 43% of colonies between 1000 and 4000 cm2 (n=7), and 88% of colonies larger than 4000 cm2 (n=33). In the same study, only colonies of A. cervicornis with a branch length larger than 9 cm were fertile, with 38% fertility for those 9 to 13 cm in branch length (n=13), 59% for 13 to 17 cm (n=17), and 89% for colonies with branches longer than 17 cm (n=18). Estimated size at puberty for A. palmata was 1600 cm2 (n=84 colonies sampled) and for A. cervicornis was 17 cm in branch length (n=52 colonies sampled). The smallest reproductive colony of A. palmata was 16 x 8 cm2 and for A. cervicornis was 9 cm in branch length (Soong and Lang 1992). In corals, fertilization can occur internally or externally, but in Atlantic Acropora spp., fertilization and development are exclusively external. Embryonic development culminates with the development of planktonic larvae called planulae. Little is known concerning the settlement patterns of planula larvae of Atlantic Acropora spp. (Bak et al. 1977 ; Sammarco 1980 ; Rylaarsdam 1983). In general, upon proper stimulation, coral larvae, whether released from parental colonies or developed in the water column external to the parental colonies, settle and metamorphose on appropriate substrates. Unlike most other coral larvae, A. palmata planulae appear to prefer to settle on upper, exposed surfaces, rather than in dark or cryptic ones (Szmant and Miller, accepted), at least in a laboratory setting. Initial calcification ensues with the forming of the basal plate and the initial protosepta, followed by the theca or polyp wall and axial skeletal members. Buds that form on the initial corallite develop into daughter corallites. Both externally and internally produced coral planula larvae presumably experience considerable mortality (up to 90% or more) from predation or other factors prior to settlement and metamorphosis (Goreau et al. 1981). Once larvae are able to settle onto appropriate hard substrates, metabolic energy is diverted to colony growth and maintenance. Because newly settled corals barely protrude above the substrate, juveniles need to reach a certain size to reduce damage or mortality from impacts such as grazing, sediment burial, and algal overgrowth (Bak and Elgershuizen 1976; Birkeland 1977; Sammarco 1985). Recent studies examining early survivorship of lab cultured A. palmata settled onto experimental limestone plates and placed in the field indicate that survivorship is substantially higher than for Montastraea faveolata, another spawner, and similar to brooding species over the first nine months after settlement (Szmant and Miller, accepted). This pattern corresponds to the size of planulae; A. palmata eggs and larvae are much larger than those of Montastraea spp. Overall, older recruits (i.e., after they have survived to a size they are visible to the human eye, probably 1 to 2 yrs after settlement (Figure 4.3-11) appear to have similar growth and post-settlement mortality rates across species (Van Moorsel 1988). Figure 4.3-11. Acropora palmata sexual recruit, St. John, U.S.V.I. Photo credit: C. Rogers. Spatial and temporal patterns of coral recruitment have been intensively studied on Caribbean reefs (Birkeland 1977; Bak and Engel 1979; Rogers et al. 1984; Baggett and Bright 1985; Chiappone and Sullivan 1996). Biological and physical factors that have been shown to affect spatial and temporal patterns of coral recruitment include substrate availability and community structure (Birkeland 1977), grazing pressure (Rogers et al. 1984; Sammarco 1985), fecundity, mode and timing of reproduction (Harriot 1985; Richmond and Hunter 1990), behavior of larvae (Lewis 1974; Goreau et al. 1981), hurricane disturbance (Hughes and Jackson 1985), physical oceanography (Baggett and Bright 1985; Fisk and Harriot 1990), the structure of established coral assemblages (Lewis 1974; Harriot 1985), and chemical cues (Morse et al. 1988). Relatively few studies, however, have examined variation in Caribbean coral recruitment over larger spatial scales (10 to 100 km) or among different structural types of reefs (Wallace and Bull 1981; Harriot and Fisk 1987; Fisk and Harriot 1990). Many studies of Caribbean reefs have used the quantification of juvenile coral densities as a proxy to measure recruitment success, with juvenile corals defined as metamorphosed corals visible underwater to the unaided eye ranging up to 4 cm in maximum diameter (Bak and Engel 1979). Newly settled corals are visible in the field at approximately 5 to 10 mm in diameter, and colonies approaching 4 cm in diameter are approximately 1 to 3 years old (Van Moorsel 1988). Studies of Acropora spp. from across the Caribbean confirm two overall patterns of sexual recruitment: (1) Low juvenile densities relative to other coral species and (2) low juvenile densities relative to the commonness of adults (Porter 1987). This pattern suggests that the composition of the adult population is dependent upon variable recruitment. It also likely reflects the dominance of asexual reproduction by fragmentation for these species (for example, surviving fragments are usually larger than 4 cm and thus never undergo a “juvenile” stage by this definition). In both Curaçao and Bonaire in the 1970s, densities of juvenile A. palmata reached 0.13 per m2, while no A. cervicornis juveniles were found (Bak and Engel 1979). On the north coast of Jamaica, juvenile A. cervicornis densities were as high as 4.3 per m2 at 11 m depth on barren substrate (Rylaarsdam 1983). However, phototransects revealed no Acropora spp. recruitment in 1976 or 1980 at <20 m depth, with smaller colonies presumably originated from the larger colonies via fragmentation (Porter et al. 1981). In Salt River, St. Croix, A. palmata juveniles occurred at densities of 0.1 to 0.3 per m2 at 3 to 9 m depth, while densities of juvenile A. cervicornis ranged from 0.01 to 0.30 per m2 at 9 m depth (Rogers et al. 1984). Similar results were obtained in the Florida Keys (Dustan 1977; Porter and Meier 1992; Chiappone and Sullivan 1996). Surveys of nine sites representing three different offshore reef types from 3 to 15 m depth yielded no juveniles of Acropora spp. from 450, 1-m2 quadrats (Chiappone and Sullivan 1996). To date, however, the settlement rates (number of larvae settling per unit area) of Atlantic Acropora spp. have still not been quantified; the juvenile density measurements cited above represent larvae that have not only settled, but metamorphosed (i.e., excreted a calcium carbonate skeleton) and survived to a specific size visible to surveyors underwater. Anecdotal evidence and observations in the Caribbean indicate that both A. cervicornis and A. palmata sexually recruit onto reefs, and in several instances, populations that have experienced major declines (>90%) are showing signs of recovery in terms of newly settled sexual recruits (Bruckner 2002). Besides sexual reproduction, most coral species, including Atlantic acroporids, also reproduce asexually. Asexual reproduction involves fragmentation, wherein colony pieces or fragments are dislodged from larger colonies to form new colonies (Highsmith 1982). The budding of new polyps within a colony can also be considered asexual reproduction. Fragmentation can occur during storms (Porter et al. 1981; Tunnicliffe 1981; Highsmith 1982), with susceptibility to mechanical breakage of colony branches influenced by the boring activities of sponges and lithophagus bivalves. Fragmentation is a common means of propagation in many species of branching corals and historically has been considered to be the most common means of forming new colonies in Atlantic Acropora spp. (Gilmore and Hall 1976; Davis 1977; Tunnicliffe 1981; Bak and Criens 1982; Hughes 1985). The perception of the dominance of fragmentation as a reproductive mode for A. palmata and A. cervicornis implies that colonies derived from fragmentation can be distinguished from those derived from larvae. However, this may not always be the case. Recently developed genetic tools can detect colonies with the same genotype (implying one was fragmented from the other). Application of these tools in the field to a population of A. palmata (where individual small colonies were scored by field experts as larval or asexual recruits) indicated very poor correlation (Miller et al. in review). It appears that the reliability of assessing the contribution of sexual versus asexual reproduction in Acropora populations by field survey is limited, but is an area of study that warrants further investigation. Asexual reproduction can play a major role in maintaining local populations when sexual recruitment is very limited. Fragmentation, followed by stabilization, survivorship, and growth can provide a mechanism for maintaining and expanding Atlantic Acropora spp. populations. However, region-wide declines have increased the reliance of Acropora spp. on sexual recruitment as a means of establishing and sustaining populations (Bruckner 2002). Atlantic Acropora spp. may require a certain storm frequency to maintain and expand populations through asexual reproduction, principally by fragmentation, when sexual recruitment is limited (Bruckner 2002). Frequent occurrence of storms or a single intense storm, however, may negatively impact colony survival, since a fragment may become abraded during the storm or may not encounter suitable substrate to reattach after the storm passes. Development and growth Staghorn Coral The growth rate for A. cervicornis has been reported to range from 3 to 11.5 cm/yr (Table 4.3-6). This growth rate is relatively fast in comparison to that of other corals and historically enabled the species to construct significant bioherms (reef structures) in several locations throughout the Caribbean (Adey 1978). Table 4.3-6. The annual growth rate for Acropora cervicornis as reported from several sources. Gladfelter (1982, 1983a) used a scanning electron microscope to describe the growth process in A. cervicornis. She reported that crystals are initially deposited randomly on the distal margin of the axial corallite. Subsequently, needle-like crystals attach and grow outward from the surface of the crystals. The needle-like crystals in contact with the calicoblastic epithelial cells grow and fuse together generating the skeletal foundation or septotheca. During daylight, calcium carbonate accretion occurs on all of the skeletal elements; at night the activity is limited to fusiform crystal formation. Gladfelter (1983b) reported daily tissue growth of 300 μm in the region of the axial polyp. “A. cervicornis exhibits a daily rhythm in calcification capacity, with daily maxima at sunrise and sunset. Daily minima occur shortly after sunrise and sunset” (Chalker 1977; Chalker and Taylor 1978; Gladfelter 1983b). Contrasting growth of in situ and laboratory-reared specimens revealed differences in the basal extension; however, other measurements (e.g., CaCO3 accretion and vertical extension) were equivalent (Becker and Mueller 2001). Growth in A. cervicornis is also expressed in expansion, occurring as a result of fragmenting and forming new centers of growth (Bak and Criens 1982; Tunnicliffe 1981). A broken off branch may be carried by waves and currents to a distant location or may land in close proximity to the original colony. If the location is favorable, branches grow into a new colony, expanding and occupying additional area. Fragmenting and expansion, coupled with a relatively fast growth rate, facilitates potential spatial competitive superiority for A. cervicornis relative to other corals and other benthic organisms (Shinn 1976; Neigel and Advise 1983; Jaap et al. 1989). Fragments that contained the axial corallite were found to have lower mortality than fragments that came from the inner portions of a colony and did not have axial corallites (Bowden-Kerby 2001a). There was up to a six-fold difference in growth rate over 12 months based on the fragment‟s origin (Bowden-Kerby 2001b). Elkhorn Coral The growth rate of A. palmata, expressed as the linear extension of branches, is reported to range from 4 to 11 cm annually (Table 4.3-7) (Vaughan 1915, Jaap 1974). The 4-cm annual growth rate cited by Vaughan (1915) undoubtedly underestimates growth. Annual linear extension was estimated to be 8.8 cm; basal extension was 2.3 mm/month, and tissue growth was 200 cm2 per month at Quintana Roo, Puerto Morelos, Mexico (Padilla and Lara 1996). A colony two meters in height could theoretically be 20 to 29 years old based on a 7- to 10-cm annual growth rate. The theoretical age of a much larger colony(4 m) is 40 to 57 years old. Linear extension and tissue growth were dependent on the size of the colony; however, basal extension was independent of colony size (Padilla and Lara 1996). Colonies of A. palmata in the field had a greater calcification rate and rate of extension relative to specimens grown in an experimental tank (Becker and Mueller 2001). Wells (1933) reported from observations in 1932 that colonies of A. palmata were eight feet high (2.4 m) and 15 feet (4.5 m) in diameter at Bird Key Reef, Dry Tortugas; this is probably the maximum size that this species can attain. Settled larvae typically create a small crust or patch with tubular corallites oriented at approximately 90 degrees from the plane of attachment. One or more protuberances develop and grow outward to form proto-branches. Table 4.3-7. Acropora palmata growth rates reported from several sources. The range of growth forms in A. palmata includes the iconic broad frond, with symmetrical colonies that are two or more meters across. Branches are up to 50 cm across and range in thickness from 4 to 5 cm, tapering toward the branch terminal; these colonies are most typical of the spur and groove formations where water circulation is omni-directional. In areas where wind and waves are predominantly from a single direction, the branches tend to grow in to the direction of the waves. The series of branches look like a medieval fortification (palisade); this growth form is typical of the barrier reef habitat. As depth increases, the branches are oriented in a more vertical orientation (Wainwright 1976; Graus et al. 1977). This compensates for hydraulic bending forces, but the thickening of the base also helps to counteract the hydrodynamic forces (Schumacher and Plewka 1981). Acropora palmata porosity ranges from approximately 35 to 45% by volume (Schumacher and Plewka 1981). Acropora palmata can rapidly monopolize large areas by fragment propagation. A branch of A. palmata may be carried by waves and currents away from the mother colony to distances that range from 0.1 to 100 m, but usually less than 30 m (Baums et al., unpublished data). Fragments cleaved from the colony may grow into new colonies (Highsmith et al. 1980; Bak and Criens 1982; Highsmith 1982; Rogers et al. 1982). Fragmentation during storm events is a significant means of generating new colonies, as documented during several storms: Hurricanes Hattie (Stoddart 1962, 1969), Edith (Glynn et al. 1964), Gerta (Highsmith et al. 1980), Allen (Woodley et al. 1981), David and Frederic (Rogers et al. 1982), Hugo (Bythell et al. 1991), Joan (Geister 1992, Zea et al. 1998), Gilbert (Kobluk and Lysenko 1992; Jordan-Dahlgren and Rodriguez-Martinez 1998), Andrew (Lirman and Fong 1996, 1997, Jaap pers. observ.), Georges and Charley (2004) (Jaap pers. observ.), as well as after Storms Bret (Van Veghel and Hoetjes 1995) and Gordon (Lirman and Fong 1997). Lirman and Fong (1997) reported that A. palmata fragment wounds healed rapidly (1.59 cm of linear growth/month). Nine months after Tropical Storm Gordon, 157 of 218 fragments had fused to the sea floor, and protobranches on the fragments grew rapidly. Population dynamics Atlantic Acropora spp. are generally considered intermediate along the continuum from rselected (rapid colonizers, fast growth, early maturation, but small maximum size and thus limited contribution to reef growth; generally brooding corals) to k-selected (slow growing, generally spawning, but attaining large colony size via indeterminate growth) life history strategies. Acropora palmata and A. cervicornis tend to be fast growing, have rapid wound healing, high rates of survival of asexually produced fragments, and the ability of broken branches to grow into new colonies (Gladfelter et al. 1978; Bak and Criens 1982; Highsmith 1982). Their level of aggression (i.e., ability to extend their mesenterial digestive filaments onto neighboring species and digest away living tissue) is relatively low compared to many other Caribbean corals (Lang 1973). However, Atlantic Acropora spp. have superior overgrowth capabilities. These life history characteristics, supported by documentation of recent trends in populations across the Caribbean, illustrate that once Acropora spp. experience local (reef-scale) reductions in colony numbers and size, recovery may not occur for decades. There are several implications of the current low population sizes of Acropora spp. throughout much of the Caribbean. First, the number of sexual recruits to a population will be most influenced by larval availability, recruitment, and early juvenile mortality. Because corals cannot move and are dependent upon external fertilization in order to produce larvae, fertilization success declines greatly as adult density declines; this is termed an Allee effect (Levitan 1991). To compound the impact, Acropora spp., although hermaphroditic, do not effectively selffertilize; gametes must be outcrossed with a different genotype to form viable offspring. Thus, in populations where fragmentation is prevalent, the effective density (of genetically distinct adults) will be even lower than colony density. It is highly likely that this type of recruitment limitation (Allee effect) is occurring in some local A. palmata and A. cervicornis populations, given their state of drastically reduced abundance/density. Simultaneously, when adult abundances of A. palmata and A. cervicornis are reduced, the source for fragments (to provide for asexual recruitment) is also compromised. These conditions imply that once a threshold level of population decline has been reached (i.e., a density where fertilization success becomes negligible) the chances for recovery are low. Population Genetics Understanding the population structure of A. cervicornis and A. palmata is complicated by the fact that both corals undergo both sexual and asexual (clonal) reproduction (see Reproduction section above) and the relative contribution of each is not readily discernable in the field (Miller et al. in review). Two aspects of population structure are of critical importance in assessing extinction risk in widespread clonal species: (1) The degree of genotypic diversity (within populations and overall); and (2) the degree of genetic exchange between populations. The levels of genotypic diversity in A. palmata and A. cervicornis are of particular concern given their presumed dominant asexual reproductive mode (Highsmith 1982) and rapid range-wide decline (see Description and Distribution section above). That is, while quantitative field surveys may provide abundance estimates based on number of colonies or percent cover, it is conceivable that the genotypic diversity in either species might be drastically lower. The degree of genetic connectivity among populations is important in understanding the potential adaptation of local populations to specific environmental conditions and the potential for re-colonization from neighboring or distant reefs in areas of extirpation. Immunological self-recognition (fusion versus rejection response when two individuals are placed in contact) was used in an early study to investigate clonal structure in A. cervicornis (Neigel and Avise 1983). This approach indicated that ramets of individual genets occurred at up to 20 m distance and individual genets occupied up to 10 m2 in Jamaica and St. Croix, U.S.V.I. (Neigel and Avise 1983). However, there has been subsequent questioning of the genetic basis of the self-recognition response, as electrophoretically distinct individuals have been shown to fuse (e.g., Heyward and Stoddart 1985). Molecular genetic analysis seems to be necessary to reliably evaluate clonal structure. Common molecular approaches to study genetic population structure such as mitochondrial DNA markers have yielded low levels of intraspecific variation in anthozoans in general and corals in particular and, hence, are of limited use in coral population genetic studies (Shearer et al. 2002). The presence of intracellular symbionts in coral tissue greatly complicates the application of highly polymorphic, anonymous DNA markers since it is difficult to distinguish between coral and symbiont DNA. Previous efforts at developing coral-specific microsatellite markers for Acropora spp. also met with little success (Marquez et al. 2000). Nonetheless, molecular genetic tools have recently become available to address questions of population genetic structure and gene flow in A. cervicornis (Vollmer and Palumbi in prep) and A. palmata (Baums et al. in press a). These tools are summarized below, based upon manuscripts in development or under scientific review. A recent study examined genetic exchange and clonal population structure in A. palmata by sampling and genotyping colonies from eleven locations throughout its geographic range using microsatellite markers (Baums et al. in press a). Results (Baums et al. in press b) indicate that populations in the eastern Caribbean (St. Vincent and the Grenadines, U.S.V.I., Curaçao, and Bonaire) have experienced little or no genetic exchange with populations in the western Caribbean (Bahamas, Florida, Mexico,Panama, Navassa, and Mona Island). Puerto Rico is an area of mixing where populations show genetic contribution from both regions, though it is more closely connected with the western Caribbean. Within these regions, the degree of larval exchange appears to be asymmetrical with some locations being entirely selfrecruiting and some receiving immigrants from other locations within their region (Baums et al. in press b). The clonal structure of individual A. palmata populations was found to be highly variable, ranging from completely sexual where each colony represents a different genet to completely asexual, where all colonies comprise a single genet (Baums et al. in prep). The overall rangewide average, expressed as Ng/N (the number of genotypes found divided by the total number of colonies sampled) was about 0.5. Interestingly, clonal structure appeared to vary between the eastern and western Caribbean, with eastern populations being denser and more genotypically diverse (i.e., greater contribution by sexual recruitment) than western populations (Baums et al. in prep). In fact, four out of five populations sampled in the Florida Keys were monoclonal, indicating they were derived from fragmentation of a single larval recruit (Baums et al. in press (a) and unpublished data). This lack of genotypic diversity in several A. palmata populations implies that sexual reproduction may be completely lost and is thus a basis for concern for the long-term persistence of this species. Measures of genetic diversity such as heterozygosity are unknown for either species and are not likely to be revealed from the current genetic approaches. Vollmer and Palumbi (in prep.) used DNA sequences of specific nuclear and mitochondrial genes, to analyze connectivity of A. cervicornis populations on a Caribbean-wide scale. Their results indicate a much finer scale of geographic differentiation (i.e., less connectivity across large areas) than the microsatellite results for A. palmata (Baums et al. in press b). They report that larval exchange between A. cervicornis populations as close as 2 to 15 km is extremely limited, implying that larval sources need to be conserved on a very small spatial scale. Little is known regarding clonal structure of A. cervicornis populations throughout their geographic range, although Vollmer and Palumbi (in prep.) indicate that approximately 60% of the colonies they sampled (purposely sampling colonies distant from each other) from areas throughout the Caribbean represented distinct genotypes. As in A. palmata, populations of A. cervicornis in southeast Florida (Broward County, probably the most abundant extant stands anywhere) appear to have low genotypic diversity as each of the large thickets sampled to date is monoclonal (Baums and Vargas unpubl. data). Ecological relationships Coral reefs serve a number of functional roles in subtropical and tropical environments of the Caribbean, including, but not limited to primary production, recycling of nutrients in relatively oligotrophic seas, calcium carbonate deposition yielding reef construction, refuge and foraging base for other organisms, and modification of near-field or local water circulation patterns (De Freese 1991). Coral reefs also protect shorelines, serving to buffer inshore subtidal (e.g., seagrass) and intertidal (e.g., mangroves) communities from otherwise high wave energy conditions in certain localities. Coral reefs are host to a multitude of species of algae, invertebrates, and fishes. Reef environments are characterized by an incredible diversity of species packed into a relatively small spatial dimension (m2 to km2) defined by high benthic diversity (Connell 1978; Richards and Lindeman 1987). Organisms essential in the construction of tropical reefs are hermatypic (reef-building) corals and coralline algae. Through reef construction, these organisms provide habitat for sedentary and mobile species (Lewis 1981). The functional roles discussed below are presented for Acropora spp. where information specific to acroporids are available, and otherwise for coral reefs in general. This generalization to function in coral reef systems as a whole is appropriate in evaluating the role of Acropora spp. given their status as constructional or “foundation” species in Caribbean coral reef ecosystems as described below. Acropora spp. were important shallow and mid-depth reef builders in the wider Caribbean Acropora palmata and A. cervicornis are two of the major reef-building corals in the wider Caribbean. Historically, both of these species formed dense thickets at shallow (<5 m) and intermediate (10 to 15 m) depths in many reef systems, including some locations in the Florida Keys, western Caribbean (e.g., Jamaica, Cayman Islands, Caribbean Mexico, Belize), and eastern Caribbean. In the Florida Keys, for example, A. palmata was the primary builder of constructional spur and groove reefs along much of the Florida reef tract, with coralline spurs up to several meters in height and up to 15 m in length (Shinn 1963; Shinn et al. 1981). Early descriptions of Florida Keys reefs referred to reef zones, of which the elkhorn (A. palmata) zone was described for many shallow-water reefs (Figure 4.3-8) (Jaap 1984; Dustan 1985; Dustan and Halas 1987). Interestingly, Shinn et al. (1977) noted that in southeastern Florida, some reefs were able to form and keep pace with sea level rise without the “help” of reef construction of A. palmata. As summarized in Bruckner (2002), however, the structural and ecological roles of Atlantic Acropora spp. in the Caribbean are unique and cannot be filled by other reef-building corals in terms of accretion rates and the formation of structurally complex reefs. Coral reefs influence water circulation patterns An important characteristic of coral reefs is their ability to modify the surrounding physicalchemical environment (Ginsburg and Lowenstam 1958). The reef framework controls the accumulation of sediments on and adjacent to the reef, as well as local circulation patterns (Jaap 1984). Barrier reefs are the best example of the ability of organic communities to affect circulation patterns that in turn influence benthic community distribution and sedimentation. Barrier reefs provide shelter for the back reef lagoon, allowing for benthic communities adapted to low-wave energy conditions, such as seagrass beds, to persist and flourish. Several studies have noted the differences in sediment and habitat characteristics between inshore and offshore environments (Enos 1977; Szmant and Forrester 1996) and associated differences in sediment nutrient characteristics. Sediments in the back reef (inner shelf margin) consist of finer grain particles with greater nutrient pools relative to sediments directly associated with reefs, such as large skeletal fragments. Benthic community distribution also differs considerably between nearshore and offshore. Seagrasses and other soft-sediment communities dominate the inner shelf margin, while reefs and bare sand slope areas dominate the outer shelf margin. Coral reefs serve important refuge and foraging functions Coral reefs, including hard substrate and associated sediments, afford organisms an incredible array of refuges (Jaap 1984). Epifauna are organisms living on the reef surface, and include mobile epifauna (crustaceans, echinoderms, mollusks, and fishes) and sessile epifauna (e.g., sponges, corals, gorgonians, and bryozoans). Infauna are those animals which burrow into hard substrate, such as polychaete and sipunculid worms, sponges, and mollusks, while minute meiofauna are associated with reef sediments. Holes and crevices in the reef structure provide shelter for echinoderms, mollusks, polychaetes, crustaceans, other invertebrate groups, and fishes. In a single coral colony, for example, Grassle (1973) counted 1,441 polychaetes representing 103 species. In several coral colonies, McClosky (1970) counted 1,517 individuals representing 37 different invertebrate species. Gastropods, crustaceans, echinoderms, and fishes consume benthic algae associated with the reef structure (i.e., coral-produced substrate); these herbivores, in turn, fuel the production of higher trophic levels such as invertivores and carnivores. While no comprehensive quantitative inventories have been made of all of the flora and fauna associated with coral reefs (Lewis 1981), probably the best information illustrating the diversity associated with these structures is for fishes. In western Atlantic reef environments, the number of fish species directly or indirectly associated with the reef system can exceed 400 species (Starck and Davis 1967; Jones and Thompson 1978; Bohnsack et al. 1987). The high taxonomic diversity of reef fishes indicates that many species are highly evolved, with several families entirely restricted to the reef environment, among them: Chaetodontidae (butterflyfishes), Scaridae (parrotfishes), Acanthuridae (surgeonfishes), Labridae (wrasses), Holocentridae (squirrelfishes), Balistidae (triggerfishes), and Pomacentridae (damselfishes) (Sale 1977; Longhurst and Pauly 1987). Many reef fishes are highly sedentary, with some species (e.g., damselfishes) actively defending territories. Even the spatial distribution of larger predatory species tends to be very reef-specific, with individuals rarely traveling more than 5 km from a home site after post-settlement, except for spawning purposes (Longhurst and Pauly 1987). In addition to the important functions of reef building and reef maintenance provided by Atlantic Acropora spp., these species serve as fish habitat (Ogden and Ehrlich 1977; Appeldoorn et al. 1996), including essential fish habitat (CFMC 1998), for species of economic and ecologic importance. Loss of Acropora spp. from the Caribbean would have substantial impacts on many coral reef species and by extension on the composition of reef communities. Assessments of reef fish abundances and diversity have been conducted in the Caribbean and the Florida Keys over the last four to five decades. Invariably, these studies have quantified fish populations relative to geomorphic strata or reef zonation (Ehrlich 1975; Sale 1980; McGehee 1994; Lindeman 1997; Kendall at al. 2003), or relative to substrate characteristics such as rugosity (Luckhurst and Luckhurst 1978), complexity (Nunez Lara and Arias Gonzalez 1998), or refuge (hole) size (Hixon and Beets 1989, 1993). A number of long-term sampling efforts may have data that can be used to infer habitat use or value, but these analyses have either not been published or are limited in spatial scope. However in St. Croix, U.S.V.I., heterotypic schools of juvenile French and white grunts (Haemulon flavolineatum and H. plumieri) were found to transfer substantial amounts of nitrogen and phosphorous in the nutrient-poor waters of a coral reef; water nearby A. palmata with grunts had ammonia (NH4+) concentrations up to 0.7 μM (micromolar) greater compared to a nearby colony without fish (Meyer et al. 1983). While direct connections between reef fishes and Atlantic Acropora spp. have not been well reported (with the exceptions below), several studies have found a positive relationship between substrate complexity and fish densities and diversity. Unfortunately, few of these studies provide data on the use of certain coral species or growth forms by particular fish species. One exception to this pattern is the study by Lirman (1999) who reported significantly higher abundances of grunts (Haemulidae), snappers (Lutjanidae), and sweepers (Pempheridae) in hightopography areas with coverage by A. palmata compared to lower topography or lower coral cover sites. Comparisons between sites where A. palmata was absent and present suggested that fish schools, comprised primarily of grunts and snappers, use A. palmata colonies preferentially. Settlement habitats of the white grunt, Haemulon plumieri, in another study were examined in southwest Puerto Rico in a Thalassia-Acropora cervicornis back-reef lagoon (Hill 2001). Although this site might nominally be classed as a seagrass bed, A. cervicornis was the primary focus of newly settling grunts. Neither the Thalassia (seagrass) nor other available coral sites (boulder or brain corals, gorgonians, algal covered corals) attracted or maintained significant numbers of juveniles during the study. Hill (2001) indicated that A. cervicornis thickets were the preferred settlement habitat for grunts that became saturated during high recruitment seasons, yielding greater usage of supposed suboptimal habitats nearby (e.g., seagrass or gorgonians). Numerous reef studies have described the relationship between increased habitat complexity, and increased species richness, abundance and diversity of fishes. Habitat selection is viewed as a trade-off between refuge from predation and access to feeding resources (Werner and Gillliam 1984). Settlement and juvenile habitats typically are thought to reduce exposure to predators (Shulman 1984). Hixon and Beets (1989, 1993) showed that appropriately sized refuges could moderate predation effects and thus alter reef fish distribution patterns. At a larger scale, complete absence of particular habitats has been shown to affect fish assemblage composition if species are not able to use alternate habitats (Nagelkerken et al. 2000). Loss of the complex habitats provided by A. cervicornis and A. palmata could result in increased rates of predation on juvenile snappers and grunts, with likely reductions of habitat-specifics like H. plumieri. It is important to note that A. palmata and A. cervicornis are the only large, branching coral species in Caribbean reef systems capable of creating large amounts of complex reef habitat. Though “standing dead” coral skeletons (especially A. palmata, as A. cervicornis tends to crumble into rubble) can still serve as habitat for fishes, subsequent storms and bioerosion will eventually destroy this habitat if none is being constructed to replace it. In the current situation, with low abundance of Acropora spp. on most Caribbean reefs, very little new complex reef habitat is being created and, hence, its availability to ecologically and economically important reef fishes is likely to continue to decline in the coming years. Competition Coral reefs are described as space-limited systems and thus it is believed that competition for space is an important structuring factor. Because of their fast growth rates and canopy-forming morphology, A. palmata and A. cervicornis are known to be competitive dominants within coral communities, in terms of their ability to overgrow other stony and soft corals. However, other types of reef benthic organisms (i.e., algae) have higher growth rates and, hence, expected greater competitive ability than Acropora spp. Since the 1980s, many Caribbean reef areas have undergone a shift in benthic community structure involving reduced cover by stony corals and increased coverage by macroalgae. This shift is generally attributed to the greater persistence of macroalgae under reduced grazing regimes due to human overexploitation of herbivorous fishes (Hughes 1994) and the regional mass mortality of the long-spined sea urchin in 1983-84. Impacts to water quality (principally nutrient input) are also believed to enhance macroalgal productivity. Aronson and Precht (2001) emphasize, however, that these Caribbean-wide changes in benthic assemblages were precipitated by massive coral mortality events (namely the loss of Acropora spp. from White Band Disease) as macroalgae are generally unable to actively overgrow and kill live corals. In other words, the coral-dominated Caribbean reef system was resistant to reduced herbivory regimes for a period of time as long as corals maintained their occupation of space. However, when coral mortality occurred, macroalgae were able to pre-empt that space (especially following the loss of grazing by Diadema) and were subsequently resistant to coral re-colonization (Hughes and Connell 1999). Thus the described shifts have been persistent on a decadal scale. The noted exception is in areas where the grazing sea urchins (Diadema antillarum) have recently recovered and removed the macroalgal dominants, thereby clearing space to allow enhanced coral recruitment (Edmunds and Carpenter 2001). In summary, macroalgae are now the major space-occupiers on many Caribbean reefs. Their dominant occupation of reef surfaces impedes the recruitment of new corals (McCook et al. 2001) and hence, recovery by sexual recruits of Acropora spp. It is unlikely, however, that macroalgae have major impacts as direct competitors with healthy adult colonies. Other encrusting invertebrates may also pose a direct overgrowth threat to small colonies or bases of Acropora spp., but the extent of such interactions is not well documented. Predation Acropora spp. are subject to invertebrate (e.g., polychaete, mollusk, echinoderm) and vertebrate (fish) predation, but “plagues” of coral predators such as the Indo-Pacific crown-of-thorns outbreaks (Acanthaster planci) have not been described in the Atlantic. Predation may directly cause mortality or injuries that lead to invasion of other biota (e.g., algae, boring sponges). The most important predators on Atlantic Acropora spp. are the fireworm, Hermodice carunculata, and the muricid snail, Coralliophila abbreviata. Both these predators will feed on a wide range of cnidarian prey, but may prefer Acropora spp. Hermodice are commonly found enveloping the long branch tips of A. cervicornis that are subsequently left devoid of tissue (Marsden 1962; Lizama and Blanquet 1975; Dustan 1977). Hermodice also feeds on branch tips or protuberances of A. palmata, where the predation scars appear as white patches (Porter 1987). Vargas-Angel et al. (2003) report a density between 86 and ~618 Hermodice ha-1 in A. cervicornis thickets in southeast Florida with predation scars affecting <0.2% of the A. cervicornis cover. There are few other data on the prevalence or impact of Hermodice on Acropora spp. populations. Although these predators rarely kill entire colonies, there are several possible mechanisms of indirect impact. Because they prey on the growing tips (including the apical polyps), especially of A. cervicornis, growth of the colony may be arrested for prolonged periods of time. Additionally, Hermodice carunculata from the Mediterranean Sea has been shown to serve as a vector for a bacterial bleaching pathogen in laboratory experiments (Sussman et al. 2003). The other important predator of Atlantic Acropora spp. is the gastropod, Coralliophila abbreviata. This predator also feeds on a wide range of corals, but seems to be particularly damaging to Acropora spp. (Baums et al. 2003b). Prevalence data from throughout the Caribbean indicates that approximately 10% to 20% of Acropora spp. colonies harbor snails (Baums et al. 2003a). The rate of consumption by Coralliophila is highly variable, but may reach 6.5 cm2 of coral tissue per snail per day (Bruckner et al. 1997) and probably averages ~1.5 cm2 of coral tissue per snail per day (Baums et al. 2003b). Given that the mean snail density on infested A. palmata colonies is reported at over three snails per colony (Bruckner et al. 1997; Baums et al. 2003a) with a maximum of at least 23 snails per colony (Baums et al. 2003a), snail predation clearly represents a significant potential source of tissue loss. There is evidence that these predators concentrate on remnant Acropora populations following host coral decline (Knowlton et al. 1990; Baums et al. 2003a). For example, after Hurricane Allen struck the north coast of Jamaica in 1980 and greatly reduced the acroporid population, C. abbreviata continued to feed on remnant A. cervicornis colonies, reducing the population further (Knowlton et al. 1981). It should be noted, however, that Coralliophila seem to be extremely rare or absent on Acropora spp. in certain areas (e.g., Bocas del Toro, Panama, Baums pers. comm.; Dry Tortugas, Miller pers. observ.). The three-spot damselfish (Pomacentrus planifrons) and other species in the genus establish algal nursery gardens within branching Acropora spp. when available and on other coral species when acroporids are rare (Thresher 1976; Brawley and Adey 1977; Kaufman 1977; Itzkowitz 1978; Williams 1978; Sammarco and Carleton 1982). Although not predators in the strict sense, the damselfishes nip off living coral tissue, thus denuding the skeleton to make a place for their algal gardens. Again, it is likely that P. planifrons impacts are proportionally greater when the abundance of Acropora is reduced. Observations in several areas (e.g., Dry Tortugas, Navassa) suggest that isolated small colonies, particularly of A. cervicornis, have a very high prevalence of damselfish occupation. Other predators also consume Acropora tissue to a lesser degree. Although not widely documented, the Caribbean long-spined sea urchin (Diadema antillarum) is known to feed upon live Acropora coral tissue (Bak and van Eys 1975, Sammarco 1980). Laboratory experiments confirmed that this sea urchin will feed on coral tissue when starved, but may also do so when feeding on turf algae when sea urchin population numbers are relatively high (e.g., >4 individuals/m2) (Porter 1987). More recent studies indicate that besides damselfishes, parrotfishes, such as the stoplight parrotfish (Sparisoma viride), may also incidentally feed upon Acropora tissue. Very little is known concerning the extent of parrotfish grazing on Atlantic Acropora spp., but monitoring in the Florida Keys indicates that these scars usually heal in a matter of weeks to months (Williams pers. comm.). Overall, predators can have important direct and indirect impacts on A. palmata and A. cervicornis. Predation impacts are greater in the current scenario of low coral abundance as coral predators have not been subject to the same degrees of disturbance mortality and their broad diet breadth has allowed them to persist at high levels despite decreases in acroporid prey. However, predation impacts on Acropora spp. appear to be much lower in certain geographic areas.