Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
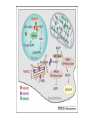
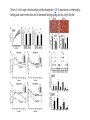
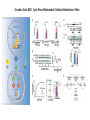
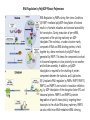
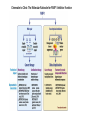
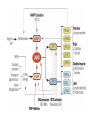
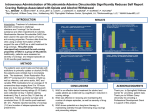
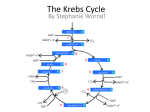
Originally defined as a molecular fraction (“cozymase”) that accelerated fermentation in yeast extracts The medical importance of NAD+ was established early with the discovery of pellagra, a disease characterized by four “Ds”: dermatitis, diarrhea, dementia, and death. A heat-stable dietary factor (known as pellagra-preventing factor) that cured pellagra was determined to be a NAD+ precursor called niacin. This provided the first evidence of a therapeutic role for what is now vitamin B3 (1). decreased cellular NAD+ concentrations occur under defined conditions, including aging, and supplementation with NAD+ precursors may be useful against aging and its chronic diseases. NAD+ biosynthesis, degradation, and salvage NAD+ is an important cosubstrate for three classes of enzymes: (i) the sirtuins (SIRTs), (ii) the adenosine diphosphate (ADP)–ribose transferases (ARTs) and poly(ADP-ribose) polymerases (PARPs), and (iii) the cyclic ADP-ribose (cADPR) synthases (CD38 and CD157). NAD+ biosynthetic pathways nicotinic acid phosphoribosyltransf erase (NAPRT) NAMN transferase (NMNAT) NAD+ synthase (NADS) A key enzyme in this pathway is nicotinamide mononucleotide adenylyl transferase (NMNAT), which transforms nicotinic acid mononucleotide (NAMN) into nicotinic acid adenine dinucleotide (NAAD) in the presence of adenosine triphosphate (ATP). Three forms of the enzyme have distinct subcellular localizations: NMNAT1 in the nucleus, NMNAT2 in the cytosol and Golgi, and NMNAT3 in the cytosol and mitochondria NAD+ as an enzyme cosubstrate Nicotinamide adenine dinucleotide is a critical cofactor for other enzymes, including the sirtuin protein deacetylases, the ADP-ribose transferases and PARP, and the cADPR synthases (CD38 and CD157). The sirtuin protein deacylases These proteins are conserved from bacteria to humans. They remove acyl groups from lysine residues on proteins in a NAD-dependent manner. NAD+ is cleaved between nicotinamide and ADPribose, and the latter serves as an acyl acceptor, generating acyl-ADP-ribose Requirement of NAD and SIR2 for Life-Span Extension by Calorie Restriction in Saccharomyces cerevisiae Calorie restriction extends life-span in a wide variety of organisms. Although it has been suggested that calorie restriction may work by reducing the levels of reactive oxygen species produced during respiration, the mechanism by which this regimen slows aging is uncertain. Here, we mimicked calorie restriction in yeast by physiological or genetic means and showed a substantial extension in life-span. This extension was not observed in strains mutant forSIR2 (which encodes the silencing protein Sir2p) orNPT1 (a gene in a pathway in the synthesis of NAD, the oxidized form of nicotinamide adenine dinucleotide). These findings suggest that the increased longevity induced by calorie restriction requires the activation of Sir2p by NAD. Mitochondrial sirtuins: regulators of protein acylation and metabolism Yeast sirtuin Sir2 was identified as a gene controlling aging in yeast, and this function was confirmed in worms and Drosophila melanogaster The sirtuin SIRT6 regulates lifespan in male mice, nature 2012 Alterations in the IGF1–AKT pathway in Sirt6transgenic males. Sirt1 extends life span and delays aging in mice through the regulation of Nk2 homeobox 1 in the DMH and LH. There are seven sirtuins in mammals in different subcellular compartments: nuclear for SIRT1, SIRT6, and SIRT7; cytoplasmic for SIRT2; and mitochondrial for SIRT3, SIRT4, and SIRT5 The protein deacetylase of SIRT1 functions as an epigenetic regulator by targeting specific histone-acetylated residues (e.g., H3K9, H3K14, and H4K16) but also regulates transcription by deacetylating transcription factors (such as TP53, NF-κB, PGC-1α, and FOXO3a) SIRT6, which is also localized in the nucleus, is linked to aging by regulating telomere stability and inflammation through NF-κB signaling. Deacetylation of histone H3K9 appears to be the modification connecting SIRT6 activity with these aging pathways. Loss of SIRT6 leads to progeria, whereas gain of function extends life span in male mice by 15% Genomic Instability and Aging-like Phenotype in the Absence of Mammalian SIRT6 we demonstrate that SIRT6 is a nuclear, chromatin-associated protein that promotes resistance to DNA damage and suppresses genomic instability in mouse cells, in association with a role in base excision repair (BER). SIRT6 links histone H3 lysine 9 deacetylation to NFkappaB-dependent gene expression and organismal life span. SIRT6 attenuates NF-kappaB signaling via H3K9 deacetylation at chromatin, and hyperactive NF-kappaB signaling may contribute to premature and normal aging. Sirtuin 3 is the major mitochondrial protein deacetylase (28). Its expression is enhanced by fasting and calorie restriction and is decreased during aging and by a high-fat diet Circadian Clock NAD+ Cycle Drives Mitochondrial Oxidative Metabolism in Mice Poly(ADP-ribose) polymerases Activated PARP1 and PARP2 catalyze the transfer of multiple ADP-ribose moieties from NAD+ to protein acceptors, generating long poly(ADP-ribose) (PAR) chains This DNA-dependent nuclear PARP is strongly activated by DNA damage, leading to consumption of a large amount of cellular NAD+. In fact, DNA damage leads to a decrease (up to 80%) in cellular NAD+ concentrations. PARP1 is important in DNA damage detection and repair, as well as in a cell’s decision to repair itself or die after a genotoxic insult RNA Regulation by Poly(ADP-Ribose) Polymerases RNA Regulation by PARPs during Non-stress Conditions (A) PARP1-mediated poly(ADP-ribosyl)ation of histones results in chromatin relaxation and increased accessibility for transcription. During maturation of pre-mRNA, components of the splicing machinery are ADPribosylated. The nucleolus, a nuclear structure mainly composed of RNA and RNA binding proteins, is held together by a dense meshwork of poly(ADP-ribose) generated by PARP1. This keeps the components involved in ribosome biogenesis in close proximity to one another and facilitates assembly. In addition, poly(ADPribosyl)ation is required for the shuttling of protein components between the nucleolus and Cajal bodies. (B) Cytoplasmic RNA regulation by PARPs. PARP7, PARP10, PARP12, and PARP13 are involved in translation inhibition, e.g., by ADP-ribosylation of the elongation factor EF2 and ribosomal proteins. PARP13 and PARP14 promote degradation of specific transcripts by targeting these transcripts to the cellular RNA decay machinery. PARP13 can also inhibit microRNA-mediated mRNA silencing. RNA Regulation by PARPs during Stress Conditions (A) Cytoplasmic stress induces stress granule assembly mediated by PARP5a, PARP12, PARP13, and PARP14. MicroRNA-mediated silencing is relieved upon cytoplasmic stress or viral infection, a process which requires PARP13 function. PARP7, PARP12, PARP13, and PARP14 directly inhibit translation in response to stress or upon viral infection. Viral RNA is additionally targeted to the RNA decay machinery by PARP13. (B) During heat shock PARP1 regulates splicing by recruiting hnRNPs to poly(ADP-ribosyl)ated proteins. This results in the dissociation of the hnRNPs from their target mRNAs. Heat shock-activated PARP1 also mediates poly(ADP-ribosyl)ation of poly(A)polymerase (PAP), resulting in decreased polyadenylation activity of the protein. RNAs lacking poly(A) tails fail to be exported to the cytoplasm and are degraded. During DNA damage, PARP2 binds to accumulated rRNA through its SAP domain, activating its enzymatic activity. Chromatin to Clinic: The Molecular Rationale for PARP1 Inhibitor Function NAD+ as an enzyme cofactor Why do concentrations of NAD+ decrease during the aging process? The fact that supplementation with NMN (a product of NAMPT) corrects defects associated with aging may indicate that the NAD salvage pathway is deficient in aging. Decreased NAMPT expression occurs in several tissues (e.g., pancreas, white adipose tissue, and skeletal muscle) during aging (89), resulting from defective circadian rhythm regulation by CLOCK and BMAL (22) or from the oxidative stress and chronic inflammation associated with aging (7)