Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
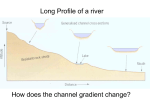
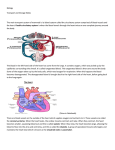
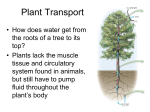
Physiology of Organisms Discuss how physiological constraints effect transport of water in different organisms The primary physiological constraint that effects transport of water is resistance, both resistance to movement of water across cell membranes, and resistance to movement of water through vessels in osmosis and bulk flow respectively. This resistance must be overcome by various means in order for water to be transported. Resistance to movement across membranes by osmosis is overcome by the mechanisms that create water potential gradients. This follows Fick’s first law of diffusion, which states that flux goes from regions of high concentration to regions of lower concentration, with a magnitude proportional to the concentration gradient. In both plants and animals water is transported between cells and from the intracellular fluid into the cells by osmosis; the movement of water from an area of high water potential, to area of lower water potential across partially permeable membrane. Water potential w = s + p, where s is the solute potential, and p is the pressure potential. Movement of water down its potential gradient involves transition to a position of lower free energy. The creation of water potential gradients by active loading of is solutes used to induce movement of water across membranes, overcoming the resistances to this; for example isosmotic fluid reabsorption in the kidneys, which occurs in the proximal tubule. Water is reabsorbed as Na+ is pumped over the basolateral membrane of the cells by Na+/K+ ATPases, causing Na+ to move from the lumen of the tubule into the epithelial cells down its concentration gradient. The solute reabsorption results in a higher osmotic pressure (the pressure that has to be applied to a concentrated solution in order to stop solvent moving into it from a more dilute solution), or lower water potential, in the peritubular fluid, which causes water to move by osmosis through leaky tight junctions, and through the epithelial cells via aquaporin-1 channel proteins. Water is moved by a similar mechanism involving pumping of solutes in plants. In the phloem, active sucrose transporters create high concentrations of sucrose in the phloem. This produces a lower water potential, therefore water follows the sucrose into the phloem to create a pressure gradient that is used for transport of the phloem’s contents. Pressure gradients are also utilised in overcoming resistance to the movement of water on a larger scale than across cell membranes. Bulk flow is a way in which water, and other fluids, are transported over large distances in plants and animals, usually through specialised vessels. In bulk flow water moves down its mechanical energy gradient. From Darcy’s law, flow of water is defined by the equation: Flow = Δ Pressure/Resistance, meaning that the rate of flow is proportional to the pressure gradient but inversely proportional to resistance. The resistance is a physiological constraint. Resistance to flow through vessels is described by the Poisseuille equation: Resistance=(length of vessel x viscosity of fluid)/(internal radius of vessel)4. An additional constraint is friction with the walls of the xylem and blood vessels. In both plants and animals movement of water through the xylem and blood vessels respectively is described by Darcy’s law and the resistance to this flow is described by the Poisseuille equation. In animals, in order for blood containing Physiology of Organisms water to flow, a hydrostatic pressure gradient must be generated. In higher animals such as mammals, birds, and fish this gradient is generated by the pumping of the heart muscle. The flow of blood to organs through the blood vessels is driven by the difference in pressure between the arteries that supply the organ, a higher pressure created by contraction of the heart, and the veins that drain it, which have a lower pressure. This is a positive pressure gradient that moves water-containing blood through the vessels. In contrast to in the blood, the driving force behind the movement of water through the xylem to overcome resistance is negative pressure. In plants the hydrostatic pressure gradient is generated by transpiration. Water evaporates and leaves the leaves through the stomata, resulting in water present in the mesophyll cells inside the leaf evaporating into the air spaces. The water in the xylem vessels are subject to very low water potential, as a result of cohesive forces from hydrogen bonds between water molecules in the vessel and those in the leaves, causing transpiration tension. Additionally the adhesive forces between the water molecules and the lignin lining the walls of the xylem contributes to the negative tension in the plant. This tension draws water from the roots to the leaves through the xylem, overcoming the resistance described in the Poisseuille equation, in addition to gravitational forces and friction. In conclusion, the physiological constraint of resistance to movement of water across cell membranes by osmosis and through vessels by bulk flow results in mechanisms to overcome these resistances and enable transport of water. These mechanisms include the generation of water potential gradients from active transport of solutes, and the generation of pressure potential gradients by the pumping of the heart muscle in animals, and transpiration in plants. The mechanisms of water transport into and out of cells in plants and animals by osmosis are very similar, whereas bulk flow is more different; for example transport of water in the blood uses a positive potential gradient, while in the xylem a negative pressure potential it utilised.