Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
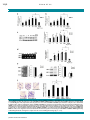
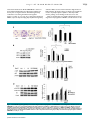
ORIGINAL RESEARCH ARTICLE Journal of Ang II–AT1R Increases Cell Migration Through PI3K/AKT and NF-kB Pathways in Breast Cancer Cellular Physiology YANBIN ZHAO,1 HONGBIN WANG,2 XIULI LI,1 MENGRU CAO,1 HAILING LU,1 QINGWEI MENG,1 HUI PANG,1 HAILIN LI,3 CHRISTINA NADOLNY,4 XIAOQUN DONG,4** 1 AND LI CAI * 1 Department of Internal Medical Oncology, Harbin Medical University Cancer Hospital, Harbin, Heilongjiang Province, China 2 Department of Surgical Oncology, Harbin Medical University Cancer Hospital, Harbin, Heilongjiang Province, China 3 Hongqi Hospital, Mudanjiang, Heilongjiang Province, China 4 Department of Biomedical and Pharmaceutical Sciences, College of Pharmacy, The University of Rhode Island, Kingston, Rhode Island Angiotensin II (Ang II), a biologically active peptide of the renin–angiotensin system (RAS), plays an important role in promoting cell migration via Angiotensin II type 1 receptor (AT1R). In this study, we examined the mechanisms by which Ang II affected cell migration in AT1R-positive MDA-MB-231 human breast cancer cells. Ang II increased cell migration and expression of matrix metalloproteinase (MMP)-2,-9 in a dose-dependent manner. Ang II-mediated cell migration was reduced by specific blocking of MMP-2 and MMP-9, as well as with pretreatment with inhibitors of AT1R, phosphatidylinositol 3-kinase (PI3K), Akt, and NF-kB. Similarly, Ang II-mediated expression of MMP-2,-9 was downregulated by pretreatment with inhibitors of AT1R and PI3K. In addition, Ang II treatment significantly induced phosphorylation of PI3K, Akt, and resulted in increased NF-kB activity. These findings suggest that Ang II activates the AT1R/PI3K/Akt pathway, which further activates IKKa/b and NF-kB, resulting in enhanced expression of MMP-2,-9 and migration in human breast cancer cells. Therefore, targeting Ang II/AT1R/PI3K/Akt/NF-kB signaling could be a novel anti-metastatic therapy for breast cancer. J. Cell. Physiol. 229: 1855–1862, 2014. © 2014 Wiley Periodicals, Inc. Angiotensin II (Ang II), a biologically active peptide of the renin–angiotensin system (RAS), plays a fundamental role in controlling blood pressure, tissue remodeling, and renal homeostasis (Dzau, 1988). Recently, Ang II has been reported to play critical roles in stimulating cell proliferation and angiogenesis of various tumors, including gastric cancer (Kinoshita et al., 2009), breast cancer (Chen et al., 2013), ovarian cancer (Suganuma et al., 2005), prostate (Uemura et al., 2011), and bladder cancer (Shirotake et al., 2011). Ang II transmits biological signals to target cells through specific G-protein-coupled Angiotensin II type 1 receptor (AT1R). Expression of AT1R has been detected in a variety of tumors, and is associated with a poor prognosis (Ino et al., 2006; Arrieta et al., 2008; De Ronde et al., 2013). Moreover, previous literature has shown that Ang II can modulate tumor cell migration and invasion (Luo et al., 2011; Dominska et al., 2012). Likewise, AT1R blockers (ARBs) can efficiently reduce tumor growth, angiogenesis, and metastasis in mouse experimental models (Deshayes and Nahmias, 2005; George et al., 2010; Wasa et al., 2011). Breast cancer is one of the most common malignances and the leading cause of cancer death in women worldwide, with an estimated 1.4 million new cases and 458,000 deaths in 2008 (Ferlay et al., 2008). The mortality for female breast cancer has increased by 20.1% during the last decade in China (Guo et al., 2012). The occurrence of distant metastasis leads to a reduced survival of patients with breast cancer. Cancer metastasis is a multiple-step process, where cancer cells first detach from the primary tumor, invade surrounding tissues and intravasate into blood and/or lymphatic systems, extravasate from the vasculature, and subsequently settle and colonize at the target organs. Matrix metalloproteinases (MMPs) belong to a zincdependent endopeptidases family and play a key role in tumor metastasis. MMPs act as enzymes that degrade extracellular matrix (ECM) and destroy the basement of membranes (Zitka © 2 0 1 4 W I L E Y P E R I O D I C A L S , I N C . et al., 2010). So far, most investigators have focused on the expression profile of MMP-2 (gelatinase A) and MMP-9 (gelatinase B). These MMPs are upregulated in various malignancies, and high levels are linked to metastasis (Kohrmann et al., 2009). Generally, MMP-2 is overexpressed constitutively in highly metastatic tumors, whereas MMP-9 is induced by stimulating factors such as inflammatory cytokines and epidermal growth factor (EGF) (Turpeenniemi- Abbreviations: Ang II, Angiotensin II; AT1R, Angiotensin II type 1 receptor; ARB, AT1R blocker; ECM, extracellular matrix; MMP, matrix metalloproteinase; PI3K, phosphatidylinositol 3-kinase; RAS, renin–angiotensin system. Yanbin Zhao and Hongbin Wang contributed equally to this work. The authors declared that they have no conflicts of interest. Contract grant sponsor: Educational Commission of Heilongjiang Province of China; Contract grant number: 12521246. *Correspondence to: Li Cai, Department of Internal Medical Oncology, Harbin Medical University Cancer Hospital, Harbin 150040, Heilongjiang Province, China. E-mail: [email protected] **Correspondence to: Xiaoqun Dong, Department of Biomedical and Pharmaceutical Sciences, College of Pharmacy, The University of Rhode Island, Pharmacy Building, 7 Greenhouse Road, Kingston, RI 02881. E-mail: [email protected] Manuscript Received: 5 January 2014 Manuscript Accepted: 28 March 2014 Accepted manuscript online in Wiley Online Library (wileyonlinelibrary.com): 1 April 2014. DOI: 10.1002/jcp.24639 1855 1856 ZHAO ET AL. Hujanen, 2005; Rydlova et al., 2008; Roy et al., 2009). Therefore, MMP-2 and MMP-9 may play a more important role in cancer cell migration and invasion. MMPs expression is regulated by transcriptional factors (like NF-kB) through upstream pathways including PI3K–Akt pathways (Westermarck and Kahari, 1999), which can regulate a variety of cellular functions, including proliferation, apoptosis, and migration (Castaneda et al., 2010). Recently, studies have shown that Ang II may act directly on breast cancer cells to accelerate the development of metastasis in vivo. Microarray studies revealed an increase in MMP-2, MMP9, and ICAM-1 expression levels in response to Ang II (Rodrigues-Ferreira et al., 2012). Ang II can stimulate the expression of MMP-2 via activation of AT1R, but not MMP-9, in B16F10 melanoma cells (Akhavan et al., 2011). These results suggest that Ang II facilitates migration and metastasis in tumor cells, and MMP-2 or MMP-9 is involved in Ang II-induced migration. However, the mechanisms of MMP-2,-9 induced by Ang II in human breast cancer cells are not well defined. In this study, we demonstrate that Ang II/AT1R signaling promotes migration and enhanced expression of MMP-2 and MMP-9 in breast cancer cells. In addition, we found that phosphatidylinositol 3-kinase (PI3K), Akt, IKKa/b, and NF-kB signaling pathways are involved in Ang II-mediated migration. These data suggest that Ang II/AT1R signaling through activation of the PI3K/Akt/NF-kB pathway plays a critical role in migration of breast cancer, and may guide AT1R-targeted therapy to treat human breast cancer. Materials and Methods Reagents and antibodies Human Ang II was purchased from AnaSpec (San Jose). AT1R antagonist losartan (DuP 753) was purchased from DuPont Merck (Hangzhou, China). AT2R antagonist PD123319 and NF-kB inhibitor pyrrolidine dithiocarbamate (PDTC) were purchased from Sigma–Aldrich (St. Louis). LY294002, p-p85, Akt, p-AktThr308, p-Akt-Ser473, IkB, p-IkB, IKKa/b antibodies were purchased from Cell Signaling Technology (Boston). Akt inhibitor, MK-2206, was purchased from Selleck Chemicals (Houston). MMP-2 siRNA, MMP-9 siRNA, IKKa-siRNA and IKKb-siRNA were purchased from Santa Cruz (CA). Cell lines and culture The MDA-MB-231 cell line was a generous gift from Heilongjiang Cancer Institute (Harbin, China). Cells were maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum and 50 U/ml penicillin/50 mg/ml streptomycin, then incubated at 37°C in a humidified atmosphere of 5% CO2. Experimental cells were grown for an additional 24 h with either serum-free medium (vehicle), Ang II (1–1,000 nM) alone, or a combination of Ang II with Losartan (105 M), PD123319 (105 M), LY294002 (50 mM), Akt inhibitor (0.5 mM). All dilutions were made in serum-free medium. These cells were then used for subsequent assays. Cell migration assays Cell migration was evaluated by a Boyden chamber assay. Prior to migration assay, cells were pretreated for 30 min with inhibitors including Losartan, LY294002, Akt inhibitor, PDTC, or vehicle control (0.1% DMSO). These concentrations of inhibitors did not affect the survival of MDA-MB-231 cells as shown by cell viability assay (data not shown). Approximately 1 104 cells in 200 ml of serum-free medium were placed in the upper chamber, and 300 ml of the same medium containing Ang II (1–1,000 nM) was placed in the lower chamber. The cells were incubated for 24 h at 37°C in 5% CO2, then fixed in methanol for 15 min and stained with 0.05% crystal violet in PBS for 15 min. Cells on the upper side of the filters were removed with cotton-tipped swabs, and the filters were JOURNAL OF CELLULAR PHYSIOLOGY washed with PBS. Cells on the underside of the filters were examined and counted under a microscope. Each experiment was repeated at least three times. The number of invading cells in each experiment was adjusted by the cell viability assay to correct for proliferation effects of Ang II treatment (corrected migrating cell number ¼ counted migrating cell number/percentage of viable cells) (Pilarsky et al., 1998). Zymography analysis To analyze metalloprotease enzymatic activity, supernatants collected from MDA-MB-231 cell cultures were mixed with sample buffer without reducing agent or heating. Samples were loaded onto 10% SDS–PAGE gel containing 1 mg/ml gelatin and electrophoresed under constant voltage. Afterward, the gel was washed with 2.5% Triton X-100 to remove SDS, rinsed with 50 mM Tris–HCl, pH 7.5, and then incubated overnight at room temperature with developing buffer (50 mM Tris–HCl, pH 7.5, 5 mM CaCl2, 1 mM ZnCl2, 0.02% thimerosal, and 1% Triton X-100). Zymographic activity was revealed by staining with 1% Coomassie Blue. Real-time RT-PCR Total RNA was extracted from cells with TRIzol reagent (Invitrogen, San Diego, CA) according to the manufacturer’s instructions, and quantified by measuring the absorbance at 260 nm. Gene expressions were detected by quantitative real-time RT-PCR (qRT-PCR) using the standard SYBR Green RT-PCR kit (Takara, Dalian, China) according to the manufacturer’s instructions. The cDNA was synthesized using the Revert Aid First-Strand cDNA Synthesis kit (Takara) according to the manufacturer’s instructions. Primer sequences were designed with Origene Technologies, Inc. (Beijing, China). The primers used were as follows: MMP-2—sense: 50 -TCTTCAAGGACCGGTTCATTTG-30 ; antisense: 50 -GATGCTTCCAAACTTCACGCTC-30 ; MMP-9—sense: 50 -ACCCTGCCAGTTTCCATTCATC-30 ; antisense: 50 -CCGGCACTGAGGAATGATCTAAG-30 ; b-actin—sense: 50 -CTACAATGAGCTGCGTGTGGC-30 antisense: 50 -CAGGTCCAGACGCAGGATGGC-30 ; PCR conditions consisted of pre-denaturation at 94°C for 2 min, followed by 35 cycles of denaturation at 94°C for 30 sec, annealing at 60°C for 30 sec and extension at 72°C for 1 min and a final extension at 72°C for 7 min. Data were analyzed by 244Ct. SiRNA transfection The siRNA against human MMP-2 (sc-29398), MMP-9 (sc-29400), IKKa (sc-29365), IKKb (sc-35644), and control siRNA (sc-37007) were purchased from Santa Cruz Biotechnology (USA). For experiments using targeted siRNA transfection, each consisted of a scrambled sequence that did not lead to the specific degradation of any known cellular mRNA. Cells were transfected with siRNAs (100 nM) using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions. Western blot analysis MDA-MB-231 cells were lysed in a lysis buffer consisting of 20 mM Tris–HCl (pH 7.5), 2 mM EDTA, 150 mM NaCl, 1% Triton X-100, and protease inhibitors. After centrifugation at 12,000g for 5 min at 4°C, the supernatant was obtained. The supernatant was used as a total cell lysate and analyzed for protein concentration by the Bradford method (BioRad, Hercules, CA). Equal amounts of cellular proteins (30 mg/lane) were separated by SDS–PAGE and transferred to a nitrocellulose membrane. After blocking with 1% skimmed milk in Tris-buffered saline/Tween-20 overnight at 4°C, the blots Ang II–AT1R AND BREAST CANCER were incubated with rabbit anti-human p-p85, p85, Akt, p-AktThr308, p-Akt-Ser473, IkB, p-IkB, IKKa/b, p65, or p-p65 at a dilution of 1:600 for 2 h at room temperature. The blots were subsequently washed three times (10 min for each wash) with Tris-buffered saline/Tween-20 and then treated with the appropriate alkaline phosphatase-conjugated anti-rabbit secondary antibody (dilution 1:5,000; Promega, Madison) for 1 h at room temperature. The bands were visualized using BCIP/ NBT (Promega) coloration method. Statistics Data are expressed as the mean SD. Statistical analysis was carried out using the Student’s t-test or ANOVA. A P-value < 0.05 was considered statistically significant. Each experiment was repeated three times with similar results. Results Ang II-induced MDA-MB-231 cell migration is mediated through AT1R In order to determine the dose-dependent effects of Ang II on cell migration, MDA-MB-231 cells were treated with Ang II (1–1,000 nM) for 24 h using the Transwell assay. Ang IIinduced cell migration in a dose-dependent manner, with a maximal response obtained at 100 nM Ang II at 24 h (172.76 13.24%, P < 0.01, Fig. 1a). In order to determine if Ang II receptors were involved in Ang II-induced cell migration, MDA-MB-231 cells were treated with Losartan (105 M, an AT1R antagonist) or PD123319 (105 M, an AT2R antagonist) for 30 min prior to Ang II treatment. Losartan significantly inhibited Ang II-induced cell migration effects (125.06 8.52%, P < 0.01), whereas PD123319 had no effect (169.51 18.17%, P > 0.05, Fig. 1b). These data suggest that Ang II may induce cancer cell migration via AT1R. Therefore, cells were not pretreated with PD123319 in subsequent experiments. Involvement of MMP-2,-9 in Ang II-induced MDA-MB-231 cell migration MMP-2 and MMP-9 have been shown to play important roles in cancer cell migration. To explore the involvement of MMP-2,-9 in Ang II-induced breast cancer cell migration, qPCR, Western blot, and zymography assays were carried out to analyze expression and activity. Treatment of cells with Ang II (1–1,000 nM) significantly increased transcriptional expression of MMP-2,-9 in a dose-dependent manner, peaking at 100 nM Ang II at 24 h measured by qPCR (Fig. 2a,b, MMP-2: 100 nM, 2.8 0.16%, P < 0.05; MMP-9: 100 nM, 2.95 0.12%, P < 0.05). Ang II also increased protein expression of MMP-2,-9 in MDA-MB-231 cells in a dose-dependent manner, peaking at 100 nM Ang II at 24 h (Fig. 2c). Expression and enzyme activity of MMP-2,-9 were also increased in the supernatant (Fig. 2d). Furthermore, MMP-2,-9 expression and enzyme activity were abolished by Losartan (105 M, an AT1R antagonist) (Fig. 2a–d). To further confirm that Ang II-induced MDA-MB-231 cell migration is associated with upregulation of MMP-2 and MMP-9, cells were transfected with siRNAs against MMP-2 or MMP-9. RT-PCR, Western blot confirmed the efficiency of the MMP-2 or MMP-9 siRNA (Fig. 2e). The Transwell migration assay demonstrated that Ang II-induced migration was significantly attenuated by knockdown of MMP-2 or MMP-9 compared to control cells (MMP-2: 131.27 10.85%, P < 0.01; MMP-9: 138.61 15.48%, P < 0.01, Fig. 2f). These results confirm that Ang II may regulate the expression of MMP-2,-9 via AT1R, and MMP-2,-9 may be involved in Ang II-induced MDA-MB-231 cell migration. PI3K/Akt signaling pathway is involved in Ang II-mediated MMP-2,-9 up-regulation and migration of breast cancer cells To further study the mechanisms underlying the migrationpromoting effects of Ang II, we investigated the signaling Fig. 1. Ang II enhanced migration of MDA-MB-231 cells. MDA-MB-231 cells were incubated with Ang II (1–1,000 nM), and in vitro migration activity was measured using the Transwell system. a: Ang II-induced cell migration was dose-dependent. b: Cells were pretreated with Losartan (105 mM, an AT1R antagonist) or PD123319 (105 M, an AT2R antagonist) for 30 min followed by stimulation with Ang II (100 nM). The in vitro migration activity measured after 24 h showed that Losartan inhibited cell migration. Results were expressed as the mean SE for three replicate wells (*P < 0.01). JOURNAL OF CELLULAR PHYSIOLOGY 1857 1858 ZHAO ET AL. Fig. 2. Ang II increased the expression and enzymatic activity of MMP2 and MMP9 in MDA-MB-231 cells. a,b: Cells were incubated with 1– 1,000 nM Ang II for 24 h; cell lysates were collected; MMP2 and MMP9 mRNA levels were determined using qPCR. c: Cells were incubated with 1–1,000 nM Ang II for 24 h; supernatants and cell lysates were then collected. MMP2 and MMP9 protein levels in cell lysates were determined by Western blot analysis. d: Enzymatic activity of MMP2 and MMP9 in cell lysates and supernatants was measured using zymography. The mRNA, protein levels, and enzymatic activity increased in a dose-dependent manner. Cells were pretreated with Losartan (105 mM, an AT1R antagonist) for 30 min followed by stimulation with Ang II (100 nM) for 24 h. Losartan significantly reduced the expression at both mRNA and protein levels, as well as inhibited enzymatic activity of MMP2 and MMP9. e,f: Cells were transfected with siRNAs specific blocking MMP2 and MMP9 for 24 h; in vitro migration was measured with the Transwell assay after 24 h. Transfection with MMP2 and MMP9 siRNAs significantly reduced Ang II-induced cell migration compared to the control and Ang II-treated group (*P < 0.01). JOURNAL OF CELLULAR PHYSIOLOGY Ang II–AT1R AND BREAST CANCER molecules downstream of AT1R. MDA-MB-231 cells were treated with LY294002 (50 mM, a PI3-kinase inhibitor) for 30 min prior to Ang II treatment. LY294002 significantly reduced the level of Ang II-mediated cell migration (124.91 10.79%, P < 0.01, Fig. 3a), as well as Ang II-induced MMP-2,-9 protein expression (Fig. 3b). As p85 is the regulatory subunit of PI3K, we then examined whether Ang II-induced PI3K activation. As shown in Figure 3c, Ang II led to a significant increase in phosphorylation of p85 in a dose-dependent manner, peaking at 100 nM Ang II in breast cancer cells. Next, we examined the role of Akt in Ang II-induced cancer migration and upregulation of MMP-2,-9. Pretreatment of cells Fig. 3. The PI3K/Akt signaling pathway is involved in Ang II-mediated upregulation of MMP2/MMP9 and cell migration of breast cancer. a,b: MDA-MB-231 cells were treated with LY294002 (50 mM, a PI3-kinase inhibitor) or Akt inhibitor for 30 min prior to the Ang II treatment. LY294002 significantly inhibited Ang II-mediated cell migration as well as reduced the expression of MMP2 and MMP9. c: Ang II led to a significant increase in phosphorylation of p85 in a dose-dependent manner peaking at 100 nM Ang II in breast cancer cells. d: Ang II stimulated the phosphorylation of Akt-Thr308 and Akt-Ser473 in a dose-dependent manner. Ang II (100 nM) caused maximal phosphorylation of Akt in MDA-MB-231 cells (*P < 0.01). JOURNAL OF CELLULAR PHYSIOLOGY 1859 1860 ZHAO ET AL. with Akt inhibitor (0.5 mM) antagonized Ang II-induced migration and expression of MMP-2,-9 in cells (129.93 14.39%, P < 0.01, Fig. 3a,b). We also investigated whether Akt-Ser473 and Akt-Thr308 were phosphorylated in response to Ang II treatment. We found Ang II stimulated the phosphorylation of both Akt-Thr308 and Akt-Ser473 in a dosedependent manner. Ang II (100 nM) caused maximal phosphorylation of Akt in MDA-MB-231 cells (Fig. 3d). Based on these results, Ang II may act through the AT1R/ PI3K/Akt signaling pathway to enhance MMP-2,-9 expression and cell migration in human breast cancer cells. NF-kB signaling is involved in Ang II-mediated MMP-2,-9 upregulation and cell migration The promoter region of MMP2 contains binding sites for NF-kB, which is a pathway that can be activated by PI3K/AKT (Kane et al., 1999; Westermarck and Kahari, 1999). To determine if NF-kB activation is involved in Ang II-induced cell migration and MMP-2,-9 upregulation in breast cancer, we used the NF-kB inhibitor pyrrolidine dithiocarbamate (PDTC). Figure 4a shows that pretreatment with PDTC (10 mM) inhibited Ang II-induced cell migration (139.23 17.40%, Fig. 4. NF-kB signaling is involved in Ang II-mediated MMP-2,-9 upregulation and cell migration. a: Pretreatment with PDTC (10 mM), IKKa, or IKKb siRNA inhibited Ang II-induced cell migration (*P < 0.05). b: Treatment of cells with PDTC also antagonized Ang II-induced expression of MMP-2,9. c: Ang II-induced IKKa/b phosphorylation in a dose-dependent manner. d: Ang II also caused IkB phosphorylation in a dose-dependent manner. e: Treatment of cells with Ang II for various time intervals resulted in p65 Ser536 phosphorylation. f: Hypothesized mechanism of Ang II-induced cell migration in breast cancer (*P < 0.01). JOURNAL OF CELLULAR PHYSIOLOGY Ang II–AT1R AND BREAST CANCER P < 0.01). Treatment of cells with PDTC also antagonized Ang II-induced expression of MMP-2,-9 (Fig. 4b). We further analyzed whether Ang II induces MMP-2, MMP-9 expression through activation of NF-kB upstream molecules IKKab and p65 through phosphorylation. Phosphorylation of IKKa/b and p65 was examined by Western blot. Stimulation of cells with Ang II-induced IKKa/b phosphorylation in a dose-dependent manner (Fig. 4c). To further confirm that Ang II-induced MDA-MB-231 cell migration is associated with upregulation of IKKa and IKKb, cells were transfected with siRNAs against IKKa and IKKb. RT-PCR, Western blot confirmed the efficiency of the IKKa or IKKb siRNA (Fig. 4a). Transfection with IKKa–siRNA or IKKb–siRNA significantly inhibited Ang II-induced cancer cell migration (IKKa–siRNA: 141.2 13.79%, P < 0.01; IKKb–siRNA: 137.5 16.95%, P < 0.01, Fig. 4a). The data suggest that IKKa/b activation is involved in Ang II-induced cell migration of breast cancer. Ang II also caused IkB phosphorylation in a dose-dependent manner (Fig. 4d). Furthermore, it has previously been shown that p65 Ser536 phosphorylation increases NF-kB transactivation. Treatment of cells with various doses of Ang II resulted in p65 Ser536 phosphorylation (Fig. 4e). These results indicate that NF-kB activation is important for Ang II-induced cancer cell migration and the expression of MMP-2,-9. Discussion Previous studies have shown that local RAS exists in various malignant tumor tissues, especially in breast cancer. Ang II promotes tumor cell proliferation, anti-apoptosis, migration, and angiogenesis. Ang II promotes the migration of androgenindependent prostate cancer cells, but not androgendependent cells (Dominska et al., 2012). Ang II also plays an important role in intrahepatic cholangiocarcinoma (ICC) cell migration by mediating epithelial–mesenchymal transition (EMT) (Okamoto et al., 2012). Ang II enhances migratory capability and vimentin expression, reduces E-cadherin expression, and stimulates translocation of b-catenin from cytoplasm into the nucleus in ICC cells (Okamoto et al., 2012). In breast cancer, Ang II can upregulate integrin b1 expression (Berry et al., 2000). An in vivo model of metastasis in breast cancer cells (D3H2LN) revealed that Ang II accelerates the formation of metastatic foci at secondary sites (RodriguesFerreira et al., 2012). It is also reported that Ang II reduces cell adhesion through AT1R by downregulation of integrins (a3 and b1) (Puddefoot et al., 2006). In this study, we proposed that the activation of the AT1R/PI3K/Akt/NF-kB cascade is involved in Ang II-induced cell migration of breast cancer. First, we demonstrated that Ang II significantly enhanced migration of AT1R-positive MDA-MB-231 cells in a dosedependent manner. Moreover, to determine whether the receptor is responsible for Ang II-mediated motility, we used receptor-type-specific inhibitors. We found the AT1R antagonist, but not AT2R antagonist, completely suppressed Ang II-enhanced migration of MDA-MB-231 cells. These findings suggest that Ang II-mediated tumor cell migration requires the activation of AT1R; thus, selective AT1R blockade therapy could efficiently inhibit the migration of AT1R expressing tumor cells in response to Ang II. We next investigated potential pathways that participate in Ang II-enhanced cell migration. Enzymatic degradation of ECM is one of the crucial steps in cancer invasion and metastasis. MMPs, a family of more than 20 extracellular enzymes, play an important role in metastasis because of their proteolytic activities, which assist in degradation of ECM and the basement membrane (Egeblad and Werb, 2002; Kerkela and SaarialhoKere, 2003; Tsai et al., 2010). Cytokines, growth factors, chemokines, and MMPs regulate tumor cell migration and invasion through autocrine and paracrine pathways JOURNAL OF CELLULAR PHYSIOLOGY (Woodhouse et al., 1997). Ang II can induce MMP2 expression in gastric cancer cells (Huang et al., 2009). Ang II can stimulate the expression of MMP-2 and MMP-13 through activation of AT1R, but not MMP-9, in B16F10 melanoma cells (Akhavan et al., 2011). In breast tumor D3H2LN cells, Ang II upregulates MMP-2/MMP-9 and ICAM1, which are involved in cell adhesion, migration, and invasion (Castaneda et al., 2010). In this study, we found that Ang II enhanced the expression and enzymatic activity of MMP-2 and MMP-9 in human MDA-MB-231 cells. In addition, specific blocking of MMP-2 and MMP-9 by siRNA significantly suppressed Ang II-induced cell migration. Therefore, MMP-2 and MMP-9 are proposed to be Ang IIresponsive mediators that can cause the degradation of ECM and lead to subsequent events of cancer migration and metastasis. Stimulation of AT1R has been shown to trigger the activation of PI3k/Akt (Takahashi et al., 1999), which plays a major role in a wide range of cellular processes, including motility, cell-cycle progression, anti-apoptosis, and angiogenesis. PI3K phosphorylates the 30 -OH group of the inositol ring in inositol phospholipids after activation by a variety of extracellular stimuli. Previous findings support a dosedependent effect of Ang II on cancer cell proliferation and anti-apoptosis through the PI3K/Akt pathway (Zhao et al., 2008, 2010). Furthermore, Ang II mediated cell migration in choriocarcinoma cells can be abolished by a selective AT1R antagonist and a PI3K inhibitor (Ishimatsu et al., 2006). In MDA-MB-231 cells, pretreatment with PI3K inhibitor LY294002 antagonized Ang II-induced cell migration and MMP-2,-9 upregulation. Moreover, Ang II increased phosphorylation of the p85 subunit in a dose-dependent manner. The most important downstream effector of PI3K is Akt. The induction of Akt activity is primarily under the control of phosphoinositide products of PI3K, PIP2, and PIP3, which bind to the pleckstrin homology domain of membrane-associated Akt. Subsequently, full activation of Akt requires phosphorylation of two amino acids in Akt, one within the activation loop (Thr308) and one at the C-terminus hydrophobic domain (Ser473) (Datta et al., 1999; Kandel and Hay, 1999). In this study, Ang II increased the protein levels of p-Thr308 and p-Ser473 in MDA-MB-231 cells in a dosedependent manner. Moreover, Akt inhibitor blocked Ang IImediated cell migration and expression of MMP-2,-9. Our data indicate that PI3K/Akt may play an important role in upregulating MMP-2,-9 and promoting cell migration of human breast cancer. The PI3K/AKT signaling pathway influences NF-kB activity by promoting IKK activation as well as subsequent phosphorylation and degradation of IkB, which activates the RelA/p65 subunit of NF-kB in a number of systems (Yin et al., 2006; Kloo et al., 2011). NF-kB regulates genes that are responsible for cell growth, anti-apoptosis, and migration. Ang II has been reported to prevent cisplatin-induced apoptosis through NF-kB activation in pancreatic cancer cells (Amaya et al., 2004). In this study, we demonstrated that Ang II-induced migration was inhibited by PDTC, an NF-kB inhibitor, indicating that activation of NF-kB might be involved in the induction of cell migration stimulated by Ang II. We also found that exposure of MDA-MB-231 cells to Ang II resulted in increased phosphorylation of IKK, IkB, and p65. The promoter region of MMP2 contains binding sites for transcription factors (NF-kB, AP-1, and SP-1) (Sato and Takino, 2010). We found that NF-kB inhibitors, as well as IKKa and IKKb siRNA, reduced Ang II-induced cell migration and expression of MMP-2,-9. Therefore, the NF-kB-binding sites may be critical for Ang II-induced MMP-2,-9 expression and cell migration in human breast cancer. However, we did not examine the role of other transcriptional factors, including CREB and p53, which also bind to the MMP2 promoter region. Whether other 1861 1862 ZHAO ET AL. transcription factors are involved in Ang II-induced MMP-2 expression in breast cancer needs further examination. Because the prognosis of breast cancer patients with distant metastasis is very poor, detection and prevention of metastasis is essential to improve clinical outcome of those patients. Our study revealed that Ang II increased expression of MMP-2,-9 and enzymatic activities as well as enhanced migration of human breast cancer cells via the AT1R/PI3K/Akt/NF-kB pathway. These findings will help us to understand the mechanisms of human breast cancer metastasis and may lead to the development of effective therapies in the future. Literature Cited Akhavan MM, Karimi M, Ghodrati M, Falahtpishe H. 2011. AT1 receptors activation enhances the expression of MMP-2, MMP-13 and VEGF but not MMP-9 in B16F10 melanoma cells. Pak J Biol Sci 14:821–830. Amaya K, Ohta T, Kitagawa H, Kayahara M, Takamura H, Fujimura T, Nishimura G, Shimizu K, Miwa K. 2004. Angiotensin II activates MAP kinase and NF-kappaB through angiotensin II type I receptor in human pancreatic cancer cells. Int J Oncol 25:849–856. Arrieta O, Pineda-Olvera B, Guevara-Salazar P, Hernandez-Pedro N, Morales-Espinosa D, Cer on-Lizarraga TL, Gonzalez-De la Rosa CH, Rembao D, Segura-Pacheco B, Sotelo J. 2008. Expression of AT1 and AT2 angiotensin receptors in astrocytomas is associated with poor prognosis. Br J Cancer 99:160–166. Berry MG, Goode AW, Puddefoot JR, Vinson GP, Carpenter R. 2000. Integrin beta1 upregulation in MCF-7 breast cancer cells by angiotensin II. Eur J Surg Oncol 26:25–29. Castaneda CA, Cortes-Funes H, Gomez HL, Ciruelos EM. 2010. The phosphatidyl inositol 3kinase/AKT signaling pathway in breast cancer. Cancer Metastasis Rev 29:751–759. Chen X, Meng Q, Zhao Y, Liu M, Li D, Yang Y, Sun L, Sui G, Cai L, Dong X. 2013. Angiotensin II type 1 receptor antagonists inhibit cell proliferation and angiogenesis in breast cancer. Cancer Lett 328:318–324. Datta SR, Brunet A, Greenberg ME. 1999. Cellular survival: A play in three Akts. Genes Dev 13:2905–2927. De Ronde JJ, Lips EH, Mulder L, Vincent AD, Wesseling J, Nieuwland M, Kerkhoven R, Vrancken Peeters MJ, Sonke GS, Rodenhuis S, Wessels LF. 2013. SERPINA6, BEX1, AGTR1, SLC26A3, and LAPTM4B are markers of resistance to neoadjuvant chemotherapy in HER2-negative breast cancer. Breast Cancer Res Treat 137:213–223. Deshayes F, Nahmias C. 2005. Angiotensin receptors: A new role in cancer? Trends Endocrinol Metab 16:293–299. Dominska K, Piastowska-Ciesielska AW, Lachowicz-Ochedalska A, Lachowicz-Ochędalska A, Ochędalski T. 2012. Similarities and differences between effects of angiotensin III and angiotensin II on human prostate cancer cell migration and proliferation. Peptides 37:200– 206. Dzau VJ. 1988. Circulating versus local renin-angiotensin system in cardiovascular homeostasis. Circulation 77:I-4–I-13. Egeblad M, Werb Z. 2002. New functions for the matrix metalloproteinases in cancer progression. Nat Rev Cancer 2:161–174. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. 2010. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer 127:2893–2917. George AJ, Thomas WG, Hannan RD. 2010. The renin-angiotensin system and cancer: Old dog, new tricks. Nat Rev Cancer 10:745–759. Guo P, Huang ZL, Yu P, Li K. 2012. Trends in cancer mortality in China: An update. Ann Oncol 23:2755–2762. Huang W, Yu LF, Zhong J, Wu W, Zhu JY, Jiang FX, Wu YL. 2009. Stat3 is involved in angiotensin II-induced expression of MMP2 in gastric cancer cells. Dig Dis Sci 54:2056– 2062. Ino K, Shibata K, Kajiyama H, Yamamoto E, Nagasaka T, Nawa A, Nomura S, Kikkawa F. 2006. Angiotensin II type 1 receptor expression in ovarian cancer and its correlation with tumour angiogenesis and patient survival. Br J Cancer 94:552–560. Ishimatsu S, Itakura A, Okada M, Kotani T, Iwase A, Kajiyama H, Ino K, Kikkawa F. 2006. Angiotensin II augmented migration and invasion of choriocarcinoma cells involves PI3K activation through the AT1 receptor. Placenta 27:587–591. Kandel ES, Hay N. 1999. The regulation and activities of the multifunctional serine/threonine kinase Akt/PKB. Exp Cell Res 253:210–229. Kane LP, Shapiro VS, Stokoe D, Weiss A. 1999. Induction of NFkappaB by the Akt/PKB kinase. Curr Biol 9:601–604. JOURNAL OF CELLULAR PHYSIOLOGY Kerkela E, Saarialho-Kere U. 2003. Matrix metalloproteinases in tumor progression: focus on basal and squamous cell skin cancer. Exp Dermatol 12:109–125. Kinoshita J, Fushida S, Harada S, Yagi Y, Fujita H, Kinami S, Ninomiya I, Fujimura T, Kayahara M, Yashiro M, Hirakawa K, Ohta T. 2009. Local angiotensin II-generation in human gastric cancer: Correlation with tumor progression through the activation of ERK1/2, NF-kappaB and survivin. Int J Oncol 34:1573–1582. Kloo B, Nagel D, Pfeifer M, Grau M, Düwel M, Vincendeau M, Dörken B, Lenz P, Lenz G, Krappmann D. 2011. Critical role of PI3K signaling for NF-kappaB-dependent survival in a subset of activated B-cell-like diffuse large B-cell lymphoma cells. Proc Natl Acad Sci USA 108:272–277. Kohrmann A, Kammerer U, Kapp M, Dietl J, Anacker J. 2009. Expression of matrix metalloproteinases (MMPs) in primary human breast cancer and breast cancer cell lines: New findings and review of the literature. BMC Cancer 9:188. Luo Y, Ohmori H, Shimomoto T, Fujii K, Sasahira T, Chihara Y, Kuniyasu H. 2011. Antiangiotensin and hypoglycemic treatments suppress liver metastasis of colon cancer cells. Pathobiology 78:285–290. Okamoto K, Tajima H, Nakanuma S, Sakai S, Makino I, Kinoshita J, Hayashi H, Nakamura K, Oyama K, Nakagawara H, Fujita H, Takamura H, Ninomiya I, Kitagawa H, Fushida S, Fujimura T, Harada S, Wakayama T, Iseki S, Ohta T. 2012. Angiotensin II enhances epithelial-to-mesenchymal transition through the interaction between activated hepatic stellate cells and the stromal cell-derived factor-1/CXCR4 axis in intrahepatic cholangiocarcinoma. Int J Oncol 41:573–582. Pilarsky CP, Schmidt U, Eissrich C, Stade J, Froschermaier SE, Haase M, Faller G, Kirchner TW, Wirth MP. 1998. Expression of the extracellular matrix signaling molecule Cyr61 is downregulated in prostate cancer. Prostate 36:85–91. Puddefoot JR, Udeozo UK, Barker S, Vinson GP. 2006. The role of angiotensin II in the regulation of breast cancer cell adhesion and invasion. Endocr Relat Cancer 13:895–903. Rodrigues-Ferreira S, Abdelkarim M, Dillenburg-Pilla P, Luissint AC, di-Tommaso A, Deshayes F, Pontes CL, Molina A, Cagnard N, Letourneur F, Morel M, Reis RI, Casarini DE, Terris B, Couraud PO, Costa-Neto CM, Di Benedetto M, Nahmias C. 2012. Angiotensin II facilitates breast cancer cell migration and metastasis. PLoS ONE 7:e35667. Roy R, Yang J, Moses MA. 2009. Matrix metalloproteinases as novel biomarkers and potential therapeutic targets in human cancer. J Clin Oncol 27:5287–5297. Rydlova M, Holubec L , Jr. , Ludvikova M , Jr. , Kalfert D, Franekova J, Povysil C, Ludvikova M. 2008. Biological activity and clinical implications of the matrix metalloproteinases. Anticancer Res 28:1389–1397. Sato H, Takino T. 2010. Coordinate action of membrane-type matrix metalloproteinase-1 (MT1-MMP) and MMP-2 enhances pericellular proteolysis and invasion. Cancer Sci 101:843–847. Shirotake S, Miyajima A, Kosaka T, Tanaka N, Maeda T, Kikuchi E, Oya M. 2011. Angiotensin II type 1 receptor expression and microvessel density in human bladder cancer. Urology 77:1009.e19–1009.e25. Suganuma T, Ino K, Shibata K, Kajiyama H, Nagasaka T, Mizutani S, Kikkawa F. 2005. Functional expression of the angiotensin II type 1 receptor in human ovarian carcinoma cells and its blockade therapy resulting in suppression of tumor invasion, angiogenesis, and peritoneal dissemination. Clin Cancer Res 11:2686–2694. Takahashi T, Taniguchi T, Konishi H, Kikkawa U, Ishikawa Y, Yokoyama M. 1999. Activation of Akt/protein kinase B after stimulation with angiotensin II in vascular smooth muscle cells. Am J Physiol 276:H1927–H1934. Tsai YY, Chiang CC, Yeh KT, Lee H, Cheng YW. 2010. Effect of TIMP-1 and MMP in pterygium invasion. Invest Ophthalmol Vis Sci 51:3462–3467. Turpeenniemi-Hujanen T. 2005. Gelatinases (MMP-2 and -9) and their natural inhibitors as prognostic indicators in solid cancers. Biochimie 87:287–297. Uemura H, Hoshino K, Kubota Y. 2011. Engagement of renin-angiotensin system in prostate cancer. Curr Cancer Drug Targets 11:442–450. Wasa J, Sugiura H, Kozawa E, Kohyama K, Yamada K, Taguchi O. 2011. The tumor suppressive effect of angiotensin II type 1 receptor antagonist in a murine osteosarcoma model. Anticancer Res 31:123–127. Westermarck J, Kahari VM. 1999. Regulation of matrix metalloproteinase expression in tumor invasion. FASEB J 13:781–792. Woodhouse EC, Chuaqui RF, Liotta LA. 1997. General mechanisms of metastasis. Cancer 80:1529–1537. Yin D, Woodruff M, Zhang Y, Whaley S, Miao J, Ferslew K, Zhao J, Stuart C. 2006. Morphine promotes Jurkat cell apoptosis through pro-apoptotic FADD/P53 and anti-apoptotic PI3K/Akt/NF-kappaB pathways. J Neuroimmunol 174:101–107. Zhao Y, Chen X, Cai L, Yang Y, Sui G, Wu J. 2008. Angiotensin II suppresses adriamycininduced apoptosis through activation of phosphatidylinositol 3-kinase/Akt signaling in human breast cancer cells. Acta Biochim Biophys Sin (Shanghai) 40:304–310. Zhao Y, Chen X, Cai L, Yang Y, Sui G, Fu S. 2010. Angiotensin II/angiotensin II type I receptor (AT1R) signaling promotes MCF-7 breast cancer cells survival via PI3-kinase/Akt pathway. J Cell Physiol 225:168–173. Zitka O, Kukacka J, Krizkova S, Huska D, Adam V, Masarik M, Prusa R, Kizek R. 2010. Matrix metalloproteinases. Curr Med Chem 17:3751–3768.