Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
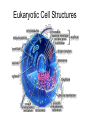
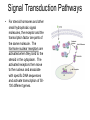
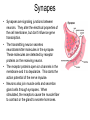
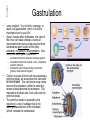
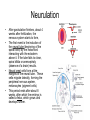
Cells and Development Domains of Life • • • • • Current classification scheme has the largest division into 3 groups: Bacteria, Archaea, and Eukarya. Based on 16S ribosomal RNA sequence similarities. Bacteria (also called Eubacteria) and Archaea are prokaryotes. Eukarya are eukaryotes. Prokaryote: no membrane-bound nucleus containing the cell’s DNA. Eukaryotes are defined by having a membrane-bound nucleus that holds the DNA. Eukaryotes also have other membrane bound organelles, and eukaryotes have linear chromosomes. Prokaryotes don’t have membrane bound organelles, although some prokaryotes have some internal membranes. Most prokaryotes have circular chromosomes, but some have linear chromosomes or even a mixture of circular an linear chromosomes. Eukaryotic Cell Structures Eukaryotic Cell Structures • nucleus: holds the chromosomes, surrounded by the double membrane nuclear envelope, which has nuclear pores in it--traffic is controlled, but ribosomes (big) can get out for example. The nucleolus is an area of the nucleus where ribosomal RNA is made in large quantities. Other structures in the nucleus have also been defined, including area for transcription and for RNA processing. • mitochondria: makes most of the ATP by aerobic respiration: Krebs cycle and electron transport/oxidative phosphorylation. Two membranes separate 2 different regions of the mitochondria. Mitochondria have their own circular DNA with about 30 genes on it: derived from bacterial DNA (endosymbiont hypothesis). Eukaryotic Cell Structures • endoplasmic reticulum (ER): series of membrane-bound channels and vesicles in the cell. The rough ER is studded with ribosomes: for translation of proteins that get secreted or get inserted into the cell membrane. The smooth ER is where sugars are added to the proteins (glycosylation); membrane lipids are also synthesized in the smooth ER. • Golgi apparatus: takes proteins from the ER and packages them for secretion from the cell. Movement between the ER, the Golgi, and the plasma membrane occurs in small membrane-bound bodies called vesicles. • Lysosomes: intracellular digestion. Low internal pH and full of various hydrolytic enzymes. Vesicles full of extracellular material get transported from plasma membrane to the lysosomes; also involved in apoptosis (programmed cell death). • Peroxisomes: use superoxide and peroxide (very toxic to the cell) to oxidize various compounds. Eukaryotic Cell Structures • Plasma membrane: the outer surface of the cell. Composed of a phospholipid bilayer: by itself only lets oxygen, carbon dioxide, water, as few other small molecules in or out. All other molecules are transported down the electrochemical gradient by channel proteins, or pumped up the gradient by ATP-driven pumps. Also, plasma membrane has adhesion proteins that connect to other cells or extracellular matrix, and receptors for hormones and other signaling molecules. • Cytoskeleton: microtubules, microfilaments, intermediate filaments: structure and transport within the cell. – microfilaments are made of actin, which interacts with various forms of myosin to provide cell movement and changes in cell shape. – microtubules are made of tubulin monomers. Mitotic spindle is made of microtubules. Also movement of organelles occurs with the interaction of microtubules and motor proteins. Cilia and flagella are made of microtubules as well. – intermediate filaments are structural only. Eukaryotic Cells • Size is limited by the need for oxygen, nutrients, control signals, etc., to diffuse from one end of the cell to the other. For more-or-less round cells, 10-30 µm is typical. Most bacterial cells are 1-2 µm. • Many different cell types in humans. All have same basic DNA, with minor changes due to random mutations plus a few cell types (e.g. immune system cells) have major DNA changes. • Most human cells are diploid (2n). However: – the gametes, sperm and egg, are haploid (1n). – some are polyploid due to endomitosis, DNA replication without cell division. Examples: hepatocytes (liver cells) range from 2n to 8n, and bone marrow megakaryocytes range from 16n to 64n. – some cells, notably red blood cells, have no nucleus. – some cells fuse together to form a multinuclear syncytium, notably muscle cells. • In most animals, the germ line cells, cells which become the sperm and egg, are separated from the somatic cells (all other cells) early in development. Tissues • • • • A tissue is a group of interconnected cells that have similar structures and functions. The cells in a tissue interact with each other through cell adhesion (sticking together) and various cell junctions, which have additional functions beyond just sticking cells together. Tissues often contain extracellular matrix, composed of large carbohydrates and proteins that are secreted by the cells. Cell adhesion molecules are transmembrane proteins, often with carbohydrates attached. Various types either interact with similar molecules on the surfaces of other cells (homophilic interaction), or with different molecules on the other cell (or in the extracellular matrix): heterophilic interactions. Cells from different tissues are held together by different sub-classes of cell adhesion molecule. Cell Junctions • • • Tight junctions are used to block movement of fluids around cells, to separate one body compartment from another. For instance, cells that line the gut are joined together with tight junctions, so molecules in the gut lumen must pass through a cell to get into the body itself. Gap junctions permit the movement of small molecules between cells (1000 daltons or less). They allow nerve cells to be electrically coupled and also allow direct communication between cells Anchoring junctions couple cells mechanically, by attaching cytoskeleton inside one cell to those inside the neighboring cell. Some types bind to extracellular matrix elements instead of other cells. Desmosomes and adherens junctions. Extracellular Matrix • • Extracellular matrix (ECM) is an array of protein fibers embedded in a gel composed of large carbohydrates called glycosamino glycans. Glycosaminoglycans are also called mucopolysaccharides. They are often very hydrophilic, so much of the structure of the ECM in many tissues is water. – Common glycosaminoglycans: chondroitin sulfate, hyaluronic acid, heparin, keratan sulfate – Most of these are covalently linked to proteins – Synthesized inside cells and secreted • • • ECM functions as a matrix for the cells of bone and cartilage, tendons, and other tissues. Cell migration during development is controlled by altering the properties of the ECM to allow cells to move in one direction only. Some ECM molecules can bind and store growth factors and other regulatory molecules. • • Epithelium is sheets of cells connected tightly with each other and to a thin, tough layer of ECM called the basal lamina (or basement membrane). It is used to line body tubes and cover the surfaces of organs Connective tissue consists of glycosamino glycans and fibers with some cells (mostly fibroblasts) embedded in it. It provides the main structural framework for the body. It holds organs in place and provides cushioning. – Fibers are mostly collagen (up to 25% of body protein!) with some elastin (stretchable) fibers as well. – Loose connective tissue contains few fibers. Adipose tissue (fat storage), skin hypodermis, blood (which as no fibrous components). – Dense connective tissue: closely packed fibers with less glycosaminoglycans. Tendons, ligaments, bones. • • Muscle. Contractile fibers (myosin and actin). Three basic types: skeletal, smooth, and cardiac. Nervous tissue. Neurons and their support cells (glia) Four Basic Tissue Types Cell Signaling • In multicellular organisms, cells need to communicate with each other, and also to respond to changes in their environment. • The cell responds to the signal by altering gene activity, changing which genes are being transcribed. • The "signal" is a molecule called a ligand, and it binds to a receptor on the receiving cell. – The usual case: the ligand is a small molecule and the receptor is a transmembrane protein. – In some cases, a ligand that is small and hydrophobic can enter the cell by itself and bind to receptors in the cytoplasm or even in the nucleus. Steroid hormones, retinoids, and nitric oxide (NO) do this. – In other cases, the ligand is attached to the surface of one cell, and it binds to a receptor on the surface of the receiving cell. • Between the receptor and the transcription factor are various proteins and small molecules that make up the signal transduction pathway. Transcription Factors • • • • • Transcription factors are proteins that bind to the DNA near a gene and stimulate its transcription. All cell signaling starts with a ligand binding to a receptor and ends with a transcription factor on the DNA. There are many different transcription factors, each of which binds to a different set of genes. Most transcription factors consist of a DNA-binding domain and an activation domain. – DNA-binding domain determines which DNA sequences the factor can bind to – activation domain stimulates transcription by interacting with transcription complex that contains RNA polymerase. Several common DNA-binding motifs: helix-turn-helix, helix-loop-helix, leucine zipper, zinc finger. Signal Transduction Pathways • For steroid hormones and other small hydrophobic signal molecules, the receptor and the transcription factor are parts of the same molecule. The hormone nuclear receptors are activated when they bind to the steroid in the cytoplasm. The activated receptors then move to the nucleus and associate with specific DNA sequences and activate transcription of 50100 different genes. Transduction Pathways • Many signal transduction pathways use transmembrane receptors attached to the primary cilium. – The primary cilium is not involved in locomotion. – It is composed of microtubule bundles like other cilia and flagella, but its function seems to be a sensory antenna. – In addition to receptors for many signaling pathways, the primary cilium is sensitive to bending and other mechanical stimuli. – Nearly all vertebrate cells have a single primary cilium on the surface of each cell. – The receptor complex in the rod cells of the retina is a primary cilium • Two general approaches to transmitting the signal from the cell surface receptor to the nucleus: kinase cascades and second messengers. Kinase Cascades • • A kinase is an enzyme that attaches a phosphate (PO4) group to another molecule. Some enzymes are activated by phosphorylating them. – A kinase that activates another kinase by phosphorylating it is called a "kinase kinase". A kinase that activates a kinase kinase is a "kinase kinase kinase". Etc. • • • These pathways start with a receptor binding to its ligand. This changes the conformation of the receptor and turns it into an active kinase. The receptor kinase then activates an intracellular kinase. Further kinases may intervene. This system allows for amplification of the signal, and for interactions between pathways. Eventually, a transcription factor is activated by being phosphorylated, and it alters transcription at target genes. • • • • • • • • G Protein Signaling Signaling pathways often involve a second messenger, which is small molecule that transmits the signal within the cell. Examples: cyclic AMP, Ca2+ Many second messenger systems use G protein signaling. The G-protein-coupled receptor is a “serpentine” protein that passes through the membrane 7 times. The ligand binding portion in on the outside, and the effector region is on the inside of the cell. In addition to working with peptide hormones, serpentine receptors also are used for the olfactory system, which is the largest gene family in humans: over 900 genes. When the ligand binds to the receptor, it changes conformation and interacts with the G protein itself. A G protein is a trimer of alpha, beta, and gamma subunits, bound to the cytoplasmic face of the membrane by covalently attached fatty acids. The G protein is usually in an inactive state. It is activated by an interaction with the receptor, and it spontaneously reverts to inactive state after a very short time. The G protein is thus thought to be the critical link that rapidly responds to changes in environmental conditions. More G Protein Signalling • • • • Alpha binds GDP in the normal, unactivated state. When the receptor interacts with the G protein, it causes the G protein alpha subunit to release the GDP and bind a GTP (the concentration of GTP is much higher than that of GDP in the cell). Binding the GTP causes the G protein trimer to dissociate into two parts: alpha+GTP, and beta-gamma. Alpha slowly hydrolyzes the GTP back to GDP, which causes the trimer to reform. – • • In some cases, other proteins stimulate the GTP-> GDP hydrolysis While the subunits are separate they are active. In some cases, alpha interacts with the next protein in the signaling pathway, and in other cases betagamma interacts with the next protein. More G Protein Signaling • In the case of adenylate cyclase, activation is accomplished by the alpha+GTP subunit. Adenylate cyclase then creates cAMP, an intracellular second messenger that activates certain kinases. • In other cases, the G protein alpha subunit activates phospholipase C, which cleaves phosphatidyl inositol bisphosphate (PIP2), a phospholipid into diacyl glycerol and inositol triphosphate (IP3). – Diacyl glycerol mobilizes Ca2+ ions from the endoplasmic reticulum, by opening a calcium ion channel. Calcium bound to calmodulin in turn activates a kinase cascade. – IP3 activates a protein kinase which in turn activates other kinases in a cascade that ends up activating transcription factors in the nucleus. Synapes • Synapses are signaling junctions between neurons. They alter the electrical properties of the cell membrane, but don't influence gene transcription. • The transmitting neuron secretes neurotransmitter molecules in the synapse. These molecules are detected by receptor proteins on the receiving neuron. • The receptor proteins open ion channels in the membrane and it to depolarize. This starts the action potential of the nerve impulse. • Neurons also join muscle cells and secretion gland cells through synapses. When stimulated, the receptors cause the muscle fiber to contract or the gland to secrete hormones. Cell Proliferation • Cells proliferate by going through the cell cycle. – Cells that are terminally differentiated stay in a modified form of the G1 stage, called G0. • 3 cell cycle check points: G1->S, G2>M, metaphase->anaphase – Cell checks for errors and only proceeds if none are found. • • • Controlled by cyclin proteins, which are synthesized at different times of the cell cycle. They bind to and activate cyclindependent kinases, which are transcription factors. Check for proper conditions and errors: DNA damage, chromosomes not attached to spindle, etc. Mitogens are external signal molecules that overcome blocks to G1->S Cell Death • Cell senescence: natural loss of ability to divide. Telomere shortening is probably the cause • Programmed cell death. Several forms, of which apoptosis is the best known. Kills cells with DNA damage, virally infected, developmental things like excess neurons and area between fingers. – The mechanism of programmed cell death is quite different from senescence. • Apoptosis is performed by several different caspase proteins. Synthesized with N-terminal “pro-domains”: short peptide region that must be removed by a protease to activate the protein. Some caspases can autoactivate, and then activate other caspases. • There are external signals and receptors that trigger apoptosis, as well as survival factors and receptors that prevent it. Differentiation • The zygote is the source of all cells in an organism. However, as cell division occurs, cells become increasingly channeled into a specific pathway that allows them to become only a single cell type in the mature organism. • The process of becoming increasingly committed to forming a single type of cell is called differentiation. • In general, once a cell has committed to becoming a certain cell type, it can’t reverse itself and become a different cell type. – However, modern techniques allow reversal of differentiation in some cases. Differentiation Potential • • The zygote and the cells of the first few divisions are totipotent: they can become any cell type. This makes identical twins possible. Once the embryo has differentiated into a blastocyst (about 100 cells), the cells of the inner cell mass will become the embryo, and they are no longer capable of forming the extra-embryonic membranes. – Inner cell mass cell are pluripotent: any of them can form any cell type in the embryo. • With further cell division and differentiation, cells become multipotent: capable of developing into several different, related cell types. A good example: the hemapoietic stem cells, which can become any of the various types of blood cell. Stem Cells • • • Most cells in an adult are terminally differentiated: they have a specific function and are not capable of dividing. Some cell types have precursor cells that divide and produce only cells of that type. The names of these cells usually ends in –blast, like osteoblasts produce bone cells and myoblasts produce muscle cells. A few cells remain multipotent in adults: tissue stem cells (also called adult stem cells). They remain capable of dividing, and they do not become terminally differentiated. – – • When a stem cell divides, one daughter cell remains a stem cell and the other one goes on to terminal differentiation. This is very common in epithelia: the cells adjacent to the basement membrane divide so one daughter cell is touching the membrane and the other isn’t. In normal tissue, only the cell attached to the basement membrane can divide. In cervical cancer (and pre-cancerous conditions), cells above the membrane can still divide: basis of the Pap smear. Stem cells are rare and hard to find: they don’t have any obvious biochemical or morphological markers on their surfaces. – For example, only about 1 cell in 10,000 in the bone marrow is a hemopoeitic stem cell. More Stem Cells • Stem cells are usually found in specific locations: stem cell niches. These areas presumably have microclimates that keep the stem cells alive and active an undifferentiated. – It is quite difficult to maintain stem cells in an undifferentiated state when they are isolated in tissue culture. • Adult stem cells are multipotent: they can differentiate into several different (related) cell types. Embryonic stem cells are pluripotent: they can generate any cell type in the body. – This makes embryonic stem cells potentially vey valuable for medicine. – However, embryonic stem cells are derived from fertilized eggs generated as part of the in vitro fertilization (IVF) process. Often IVF produces more embryos than the mother wants to carry. Extras are either frozen away (and almost never used), or donated to produce embryonic stem cells. There is a big moral issue with this. Cell Culture • How to study cell behavior. Can do it in whole organisms, sometimes called in situ studies. This can have lots of complicating factors as many tissues and organs interact. Also, can’t see or access many cells. • Tissue explants: cut out a piece, culture it in a nutrient medium • Primary cell culture: dissociate a tissue into individual cells and grow in nutrient medium. Problem: cells are mortal, after about 60 divisions they stop dividing. • Permanent cell line: Easy to grow and maintain. No limit to cell divisions, immortalized (transformed) by mutations equivalent to cancer induction. Can be done spontaneously, or with known mutagens, or by transfection with DNA containing oncogenes (mutated genes which cause cancer). – an important resource is cell lines from interesting patients. To keep a stable source of their genetic material, lymphoblastoid cells can be immortalized by Epstein-Barr virus, which remains separate from the chromosomal DNA. Principles of Development • How to get from a single cell, the zygote, to a multicellular organism. • General events: – cell division and growth – differentiation: cells develop different phenotypes – pattern formation: overall development of body axes, the general body plan, and structure of individual organs – morphogenesis: changes in shape Canalization of Development • • • • Cells become increasingly specialized during development. Their range of possible fates (final cell type) decreases. This is called the canalization of development. Initially, cells of the embryo are totipotent: can develop into any embryonic cell. After a while, embryo is divided into a trophoblast and an inner cell mass. Inner cell mass become the embryo while trophoblast becomes outer membranes and placenta. Cells in ICM can become any embryonic tissue, but they can’t become trophoblast cells: these cells are pluripotent. As development proceeds, embryonic cells become increasingly specialized and can no longer become any final cell type: they become multipotent and finally unipotent when they can only become one final cell type. At some point, a cell is determined to be a particular cell type. Determination is followed by differentiation, changes of form and function, into that cell type. decisions about determination are caused by a cell’s lineage: previous decisions, by its position in the embryo, and by signals passed between the cells. Stem Cells • • • Stem cells are self-renewing cells that differentiate into a variety of cell types. After a stem cell divides, one daughter cell typically remains a stem cell, while the other one starts to differentiate into a final cell type: this is called asymmetric cell division. There are many types of stem cell in adults, and they are generally rare and hard to find. Some differentiate into a single cell type, while others can have multiple fates. Embryonic stem cells are the pluripotent cells of the inner cell mass, which can become any kind of embryonic cell. Pattern Formation • How does the basic body plan get formed – axes: dorsal-ventral (back-front), cranial-caudal (head-tail), left-right. – position within an organ: e.g. how to get 5 different fingers on a hand • axis development: based partly an uneven distribution of components in the egg and partly on external events. – sperm entry point determines boundary between trophoblast and inner cell mass, which in turn determines the dorsal-ventral axis – cranial-caudal axis probably determined by position of second polar body exit relative to sperm entry point • morphogen gradients. Certain cells secrete chemicals that act as morphogens: signals that allow other cells in that tissue to determine their position in the tissue. The farther a cell is from the morphogen secretion site, the lower the concentration of the morphogen. – a well known example is the zone of polarizing activity, which occurs in limb buds. Cells nearest the ZPA become the little finger or toe, while those farthest away become the thumb or big toe. Hox genes • • • Animal development along an axis, from Drosophila to humans, is largely determined by clusters of homeobox (Hox) genes. Different members of the Hox clusters are activated in different parts of the morphogen gradient, in an overlapping pattern The Hox gene products then stimulate the activity of other genes that cause the cells to differentiate into the proper type. Fertilization • • • • • • Egg is surrounded by two layers of extracellular matrix, the vitelline membrane and the zona pellucida. The sperm cells must dissolve their way through these layers to get to the egg. Sperm contain an acrosome at their tips that contains the necessary enzymes. When a sperm reaches the egg membrane, the membranes fuse, putting the sperm nucleus inside the egg. Egg membrane then depolarizes and cortical granules release their contests to push all other sperm cells away. Meiosis 2 occurs and the second polar body exits opposite the sperm entry point. the male and female pronuclei then undergo mitosis together, and the resulting nuclei fuse. Fertilization occurs in the Fallopian tubes. After fertilization, the embryo takes about a week to reach the uterus. Early Development • • • • • The early cell divisions of the embryo occur without any overall growth. These divisions, the cleavage divisions, result in the morula, a ball of 16 or more cells. Each cell is called a blastomere. After a few more divisions, cells on the outside of the morula flatten out, and the inside develops into a hollow ball, the blastocyst. On one side of the blastocyst a clump of cells, the inner cell mass, forms. The inner cell mass develops into the embryo and the amnion, the inner membrane. The other cells of the blastocyst are called the trophoblast (trophoectoderm), The trophoblast forms the chorion, the outer membrane of the embryo, and the embryonic part of the placenta (which is also composed of maternal tissues). At about 5 days afer fertilization, the blastocyst hatches by releasing itself from the zona pellucida that surrounded the egg, Then implantation into the uterine wall occurs, about 6 days post-fertilization. Gastrulation • • Lewis Wolpert: “It is not birth, marriage, or death, but gastrulation, which is truly the important event in your life.” About 3 weeks after fertilization, the cells of the inner cell mass undergo a series of movements that end up producing the three fundamental germ layers of the body: ectoderm, mesoderm, and endoderm. Also, body orientation gets established. – – – • • ectoderm turns into skin and nervous system mesoderm turns into muscle, bone, circulatory system, kidneys endoderm turns into gut lining, endocrine glands, most internal organs Cells in one area of inner cell mass develop a primitive streak, an area where the cells start to move inward. The cells that end up inside become the endoderm, while the cells that remain outside become the ectoderm. The mesoderm develops last, from cells near the primitive streak. The primitive streak is replaced by the notochord, a rod of cartilage that is the defining characteristic of the chordates (which includes the vertebrates). Neurulation • • • • After gastrulation finishes, about 4 weeks after fertilization, the nervous system starts to form, The first event is the induction of the neural tube (beginning of the spinal cord) by the notochord interacting with the ectoderm above it. If the tube fails to close, spina bifida or anencephaly (absence of a brain) results. Neural crest cells form at the margins of the neural tube. These cells migrate laterally, forming the peripheral nervous system, melanocytes (pigment cells). This period ends after about 8 weeks, after which the embryo is called a fetus, which grows and develops further. Twinning • • • • 2 basic types: dizygotic (fraternal): develop from 2 separate eggs fertilized by separate sperm. Nothing more than siblings who happen to share a womb. monozygotic (identical): develop from one fertilized egg, with the embryo splitting into 2 early in development. Cause is unknown. Cells up to the 4 cell stage are pluripotent: any single cell can develop into a whole person. This limits identical siblings to quadruplets. splitting the embryo after about 12 days of development can be incomplete, resulting in conjoined twins. The joined region can include almost any area of the body and any degree of completeness. – A split before 4 days gives separate placentas – 4-8 day gives a shared placenta but separate amniotic sacs – 8-12 day split gives shared amniotic sacs, but still two separate individuals – 13-15 day splits are usually incomplete, resulting in conjoined twins. • A competing theory says that you start with a fertilized egg, the embryo splits completely, and then stem cells seeking similar cells cause them to re-fuse.