Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Complement system wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Monoclonal antibody wikipedia , lookup
DNA vaccination wikipedia , lookup
Innate immune system wikipedia , lookup
Immune system wikipedia , lookup
Antimicrobial peptides wikipedia , lookup
Adaptive immune system wikipedia , lookup
Human leukocyte antigen wikipedia , lookup
Molecular mimicry wikipedia , lookup
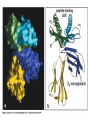
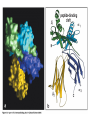
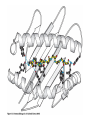
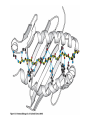
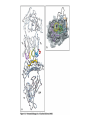
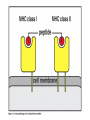
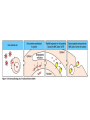
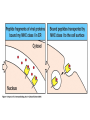
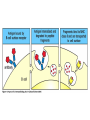
The Major Histocompatibility Complex (MHC) • In all vertebrates there is a genetic region that has a major influence on graft survival • This region is referred to as the Major Histocompatibility Complex (MHC) • Individuals identical for this region can exchange grafts more successfully than MHC non-identical combinations • Unlike minor histocompatibility antigens, the MHC products play an important role in antigen recognition by T cells Structure of MHC proteins • The MHC genes and their products are grouped into 2 classes on the basis of their chemical structure and biological properties • The two MHC proteins have a similar secondary and tertiary structure with subtle functional differences Structure of MHC proteins Structure of MHC proteins • Class I molecules are made up of one heavy chain (45 kD) and a light chain called ß2microglobulin (12 KD) that contributes to the overall structure of the protein Figure 3-20 Figure 3-20 part 1 of 2 Structure of MHC proteins • Class II molecules do not contain ß2microglobulin and consist of two (alpha and ß) chains of similar size (34 and 30 kD) • Both classes of MHC molecule fold up to produce very similar 3-D structures Figure 3-21 Figure 3-21 part 1 of 2 Structure of MHC proteins • Each has 2 MHC-unique domains which fold together to form a peptide binding platform • This structure forms a cleft or groove which accommodates a peptide • In both classes the peptide binding "MHC superdomain" is supported by a pair of immunoglobulin-like (IgSF) domains • The differences between the 2 classes are the linear connectivity of the polypeptide chains and the dimensions of the peptide-binding groove which accommodates 8-9 amino acids in class I but is openended for class II Expression of MHC molecules • MHC class I molecules are widely expressed, though the level varies between different cell types • MHC class II molecules are constitutively expressed only by certain cells involved in immune responses Figure 3-19 MHC Molecules MHC Loci • In man and mouse, as in most species, each class of MHC is represented by more than one locus (polygeny), in man these are called HLA for Human Leucocyte Antigen • The class I loci are HLA-A,-B and -C • The class II loci HLA-DR, -DQ and –DP • All the MHC genes map within a single region of the chromosome (hence the term Complex) MHC Molecules MHC Function • The products of the MHC play a fundamental role in regulating immune responses • T cells must recognise antigen as a complex with MHC molecules • This requires antigen to be processed by unfolding and proteolytic digestion before it complexes with the MHC molecule • Once formed the complex of antigenic peptide and MHC are generally very stable (half life ~ 24hrs) MHC Molecules MHC Function • Thus the biological role of MHC proteins is to bind small peptides and to "present" these at the cell surface for the inspection of T cell antigen receptors • The allelic variation of MHC molecules is functionally reflected in the selection of peptides which can bind Figure 3-20 part 2 of 2 Figure 3-21 part 2 of 2 MHC Molecules- T Cell Receptors • T cells requires MHC antigens MHC Molecules Peptide Binding to MHC • Each allelic product has a unique set of peptides which it can bind with high affinity (though rarely particular peptides may bind to more than one MHC allele) • In a normal cell the majority of MHC molecules will be complexed with self peptides, "empty" MHC molecules are less stable especially in the case of class I products • There are 50,000 - 100,000 MHC molecules on a typical cell • Most 'normal' MHC molecules are occupied by self peptides • The requirements for binding to a particular allele are met by ~1/1000 1/10000 random peptides • This would lead to the population of any given MHC allele on a single cell displaying a very large number of peptides each at only a few copies per cell • But there is a restriction on binding to tightly • This would make it easier for small pathogens to escape the immune response by having no peptides which bind to a given host's MHC molecules MHC Molecules Peptide Binding to MHC • This stringency has to make a balance between allowing too many peptides to bind • The typical population of ~100,000 MHC class I molecules of a single allotype on a normal cell displays >1000 different peptides • Individual peptide-MHC complexes are represented in widely different amounts from 1 - 5000 molecules/cell (mean~100) • T cells vary in the threshold for activation from a few (1?) complexes/cell to a few thousand, depending on the affinity, activation state etc. of the T cell and on the antigen presenting cell . Figure 3-22 Figure 3-23 Figure 3-25 Figure 3-27 Figure 3-28 MHC Molecules Pathways for antigen processing • The 2 classes of MHC molecule are specialised to present different sources of antigen • MHC class I molecules present endogenously synthesised antigens, e.g. viral proteins • MHC class II molecules present exogenously derived proteins, e.g. bacterial products or viral capsid proteins • The cell biology and expression patterns of each class of MHC are tailored to meet these distinct roles • MHC class I molecules are very unstable in the absence of peptide. They bind peptides in the Endoplasmic reticulum (ER) • Peptides are generated continuously in the cytoplasm by the degradation of proteins, predominantly by the proteasome • Peptides of suitable length (~8-18 amino acids) are specifically transported across the ER membrane by a heterodimeric transporter made up of the TAP1 and TAP2 molecules Figure 1-27 Figure 1-28 Figure 1-28 part 1 of 2 Figure 1-28 part 2 of 2 MHC Molecules Pathways for antigen processing • MHC class II molecules bind to a third polypeptide in the ER called invariant chain or Ii .The invariant chain serves two purposes. It blocks the binding of peptides to the class II molecule and it targets the class II molecule to a specialised endosomal compartment (MIIC). Exogenous antigens enter the cell in membrane vesicles, either by fluid phase pincytosis or receptor mediated endocytosis. These vesicles fuse with the MIIC compartment. The MIIC compartment has an acid pH and contains proteases, this combination unfolds and degrades both the antigen and the invariant chain causing the generation of antigenic peptides and the release of class II molecules to bind those peptides with appropriate sequence motifs .The class II molecules, peptide complexed or "empty", then traffic to the plasma membrane . Figure 1-29 Figure 1-29 part 1 of 2 Figure 1-29 part 2 of 2