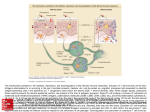
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
From www.bloodjournal.org by guest on August 9, 2017. For personal use only. Review in translational hematology Manipulating dendritic cell biology for the active immunotherapy of cancer David W. O’Neill, Sylvia Adams, and Nina Bhardwaj Dendritic cells (DCs) are specialized antigen-presenting cells (APCs) that have an unequaled capacity to initiate primary immune responses, including tolerogenic responses. Because of the importance of DCs in the induction and control of immu- nity, an understanding of their biology is central to the development of potent immunotherapies for cancer, chronic infections, autoimmune disease, and induction of transplantation tolerance. This review discusses recent advances in DC research and the application of this knowledge toward new strategies for the clinical manipulation of DCs for cancer immunotherapy. (Blood. 2004;104:2235-2246) © 2004 by The American Society of Hematology Introduction The concept of tumor “immunosurveillance,” whereby the host immune system is thought to protect against the development of primary cancers, has been debated for decades and has been recently resurrected.1 Evidence in support of tumor immunosurveillance includes observations in mice that lymphocytes and molecules essential for immune function, such as interferon-␥ (IFN␥) and perforin, collaborate to protect against the development of certain cancers. Additional corroboration has come from identification of numerous human tumor-associated or tumor-specific antigens recognized by T cells and from isolation of tumor antigenspecific T cells from metastatic lesions. Furthermore, infiltration of certain human cancers by T cells may correlate with dramatically improved survival.2 The accumulating evidence in favor of tumor immunosurveillance indicates that immunotherapies or “vaccines” may prove effective for the treatment of cancer. Indeed, numerous published reports have shown that vaccination of cancer patients with killed tumor cells, tumor cell lysates or tumor antigen proteins, peptides or DNA administered with cytokines or adjuvants can produce immunologic and clinical responses. However, the immune responses to these vaccines are often weak, and clinical responses are rarely complete and long lasting.3-5 Dendritic cells (DCs) are bone marrow–derived antigenpresenting cells (APCs) that play a critical role in the induction and regulation of immune responses. It has been proposed that the manipulation of DCs as a “natural” vaccine adjuvant may prove to be a particularly effective way to stimulate antitumor immunity.6,7 This hypothesis has been supported by experiments in mice. However, published reports of DC-based vaccine trials in humans have yet to demonstrate improved potency of DC vaccines over more traditional vaccine preparations.5,8,9 In this review we discuss the pitfalls of current DC vaccine approaches in the context of recent advances in DC biology and how improved understanding of DC biology can be applied to develop more effective immunotherapies for cancer. From the New York University (NYU) Cancer Institute Tumor Vaccine Center, NYU School of Medicine, New York, NY. Submitted December 24, 2003; accepted June 2, 2004. Prepublished online as Blood First Edition Paper, July 1, 2004; DOI 10.1182/blood-2003-12-4392. Supported by grants from the National Institutes of Health (CA-84512, AI-44628), the Cancer Research Institute, the Burroughs Wellcome Fund, and the Doris Duke Charitable Foundation. N.B. is an Elizabeth Glaser Scientist of BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 DC biology DC differentiation and subtypes DCs are a heterogeneous population of cells produced in the bone marrow in response to growth and differentiation factors fms-like tyrosine kinase-3 ligand (Flt3L) and granulocyte-macrophage colony-stimulating factor (GM-CSF). There are 3 generally accepted stages of differentiation for all DC subtypes: DC precursors, immature DCs, and mature DCs.10 In human blood, immature DCs and DC precursors are lineage-negative (CD3⫺CD14⫺CD19⫺CD56⫺) HLA-DR⫹ mononuclear cells6 and are traditionally divided into 2 populations by staining with antibodies to CD11c and CD123 (interleukin 3 receptor ␣ [IL-3R␣]). CD11c⫹CD123lo DCs have a monocytoid appearance and are called “myeloid DCs” (MDCs), whereas CD11c⫺CD123hi DCs have morphologic features similar to plasma cells and are thus called “plasmacytoid DCs” (PDCs). Although commonly used, this nomenclature is somewhat misleading. Experiments in mice indicate that both DC populations can be derived from Flt3-expressing myeloid and lymphoid progenitors.11,12 PDCs and MDCs differ in many ways, including their tissue distribution, cytokine production, and growth requirements. PDCs are important in innate antiviral immunity, are found primarily in blood and lymphoid organs, and are the principal interferon ␣ (IFN␣)–producing cells in the body. PDCs can activate antitumor and antiviral antigen responses,13,14 but their potential as immunotherapeutic adjuvants is largely unexplored because they are difficult to obtain in large quantities. MDCs, the focus of this review, are found in many tissues, where they may be classified into 2 principal subtypes: Langerhans cells (which express the C-type lectin Langerin, have unique intracellular organelles called Birbeck granules, and are found in the epidermis and oral, respiratory, and genital mucosa), and so-called interstitial, dermal, or submucosal DCs (variously named according to their anatomic location).15 the Elizabeth Glaser Pediatric AIDS Foundation. D.W.O. is supported in part by an NYU Cancer Institute Translational Research Program Award. S.A. is supported in part by an American Society of Clinical Oncology Young Investigator Award. Reprints: Nina Bhardwaj, New York University School of Medicine, 550 1st Ave, MSB 507, New York, NY 10016; e-mail: [email protected]. © 2004 by The American Society of Hematology 2235 From www.bloodjournal.org by guest on August 9, 2017. For personal use only. 2236 BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 O’NEILL et al Antigen uptake, processing, and presentation DC maturation DCs capture bacteria, viruses, dead or dying cells, proteins, and immune complexes through phagocytosis, endocytosis, and pinocytosis. They have an array of cell surface receptors for antigen uptake, many of which also function in signaling or cell-cell interactions (Table 1). DCs process captured proteins into peptides that are loaded onto major histocompatibility complex class I and II (MHC I and II) molecules, and these peptide-MHC complexes (pMHC I and II) are transported to the cell surface for recognition by antigen-specific T cells (pMHC I and pMHC II are recognized by CD8⫹ and CD4⫹ T cells, respectively).22 Antigens acquired endogenously (ie, synthesized within the DC cytosol) are typically processed and loaded onto MHC I, whereas antigens acquired exogenously (from the extracellular environment) are processed onto MHC II. Processing of endogenous proteins onto MHC I is through a cytosolic pathway that involves ubiquitination, degradation by proteasomes, and transport by TAP (transporters for antigen presentation) into the endoplasmic reticulum. In contrast, exogenously acquired proteins are typically degraded in endosomes/ lysosomes, where the peptides are loaded onto MHC II following degradation of the MHC II–associated invariant chain (Ii, which blocks access to the peptide-binding pocket of MHC II).22 An alternative pathway also exists whereby DCs process exogenous antigens onto MHC I. This pathway, called “crosspresentation,” permits DCs to elicit CD8⫹ as well as CD4⫹ T-cell responses to exogenous antigens such as apoptotic or necrotic tumor cells, virus-infected cells, and immune complexes.23-25 Cross-presentation is linked to specific DC antigen uptake receptors (Table 1), which may be targeted in strategies to load exogenous antigens onto both MHC I and II.24 Lipid and glycolipid antigens expressed on pathogens or self tissues are presented by DCs to T cells on CD1 molecules (CD1a-d), which are structurally similar to MHC I but specialized to bind lipids instead of peptides.26,27 Processing of lipid antigens onto CD1 molecules is carried out in specialized intracellular compartments, much like antigen processing onto MHC II. CD1 molecules present lipid antigens to a variety of lymphocytes, including T cells with substantial T-cell receptor diversity as well as relatively invariant natural killer T (NKT) cells. Maturation is a terminal differentiation process that transforms DCs from cells specialized for antigen capture into cells specialized for T-cell stimulation. DC maturation is induced by components of pathogens or by host molecules associated with inflammation or tissue injury. These stimuli are often collectively referred to as “danger signals.”28 Maturation is characterized by reduced phagocytic uptake, the development of cytoplasmic extensions or “veils” (Figure 1), migration to lymphoid tissues, and enhanced T-cell activation potential. Mature DCs express a number of characteristic markers, including CD83, a cell surface molecule involved in CD4⫹ T-cell development and cell-cell interactions,29,30 and DCLAMP, a DC-specific lysosomal protein. Maturation signals act on DCs through receptors that trigger intracellular signaling, including receptors for host-derived inflammatory molecules such as CD40L, TNF␣, IL-1, and IFN␣ (Table 2). Microbial products and molecules released by damaged host tissues transmit maturation signals through Toll-like receptors (TLRs), trans-membrane receptors expressed on DCs and other cell types related to Drosophila Toll protein.40 There are 11 known TLRs thus far, each with different expression patterns and each recognizing different sets of molecules. In humans, MDCs express TLRs 1 through 5 and, depending on the MDC subset, TLR 7 and/or 8. Human PDCs express TLRs 1, 7, and 9.41-43 Some TLRs act at the cell surface, whereas others such as TLRs 3, 7, 8, and 9 are found within endosomes and are presumably activated following capture and internalization of pathogens or their products. TLRs signal through the adapter molecule MyD88, which recruits other signaling molecules in a pathway that activates NF-B and mitogen-activated protein (MAP) kinases, inducing the transcription of genes encoding inflammatory mediators such as TNF␣, IL-1, and IL-6.44 Stimulation of some TLRs can trigger additional, MyD88-independent, signaling pathways.44 In DCs, the distinct signaling pathways triggered can influence the direction of the resulting T-cell response.45 TLR agonists, therefore, can be used to target DC subsets to induce desired T-cell responses. On maturation, DCs develop an enhanced ability to form pMHC II46 and pMHC I,22 and some maturation stimuli can also induce Table 1. DC antigen uptake receptors Receptor class C-type lectin-like Examples DC-SIGN (CD209) Ligands Notes Viruses (HIV, Dengue, Ebola), DC-SIGN also binds adhesion molecules (ICAM-2 and -3) that are important receptors16 Fc receptors mycobacteria MMR (MRC1) Mannosylated molecules DEC-205 (LY75) ? BDCA-2 (CLECSF11) ? Langerin ? Dectin-1 -glucan Fc␥RI (CD32) Immune complexes and Fc␥RII (CD64) Integrins in DC trafficking and cell-cell interactions. — opsonized cells ␣V5 Apoptotic cells ␣M2 (CD11b/CD18, CR3) Opsonized antigens (via ␣X2 (CD11c/CD18, CR4) Opsonized antigens (via iC3b) CD36 Apoptotic cells LOX-1 (OLR1) Hsp-peptide complexes CD9119 Hsp-peptide complexes Aquaporins Fluids CR3 binds ICAM-1, found on activated lymphocytes and endothelial cells. iC3b), bacteria Scavenger receptors17 Heat shock proteins (Hsps) are highly conserved molecules that are released by dying cells and that can mature DCs. Their normal function is to chaperone peptides between subcellular compartments.17,18 Other — DC-SIGN indicates dendritic cell-specific ICAM-3-grabbing nonintegrin; ICAM-2, intracellular adhesion molecule 2; MMR, macrophage mannose receptor; BDCA, blood dendritic cell antigen; CR, complement receptor; LOX-1, low density lipoprotein, oxidized, receptor 1; and CLECSF, C-type lectin superfamily. Antigen uptake by way of DEC-205,20 Fc␥ receptors,135 ␣V5 integrin,21 CD36,21 LOX-1,18 and CD9119 have all been associated with cross-presentation. From www.bloodjournal.org by guest on August 9, 2017. For personal use only. BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 MANIPULATING DC BIOLOGY FOR IMMUNOTHERAPY 2237 Figure 1. DC morphology. (A) Immature monocyte-derived DC. (B) Monocyte-derived DC matured with IL-1, IL-6, tumor necrosis factor ␣ (TNF␣), and prostaglandin E2 (PGE2). Dif-Quick–stained cytocentrifuge preparations are shown. Viewed at 1000⫻ magnification using an Olympus BX51 microscope with an Olympus U Plan Fluorite 100⫻/1.30 NA oil immersion objective. Images were captured using an Optronics MicroFire digital camera and MicroFire image acquisition software and processed with Adobe Photoshop. cross-presentation.47-49 Maturation also results in increased expression of adhesion and costimulatory molecules involved in the formation of the immunologic synapse (Figure 2) and induces DCs to secrete cytokines that are critical in determining the nature of the ensuing immune response (Figure 3). Another important effect of maturation is the induced secretion of chemokines that recruit monocytes, DCs, and specific subsets of T cells into the local environment (Table 3). Finally, maturation imparts on peripheral DCs the ability to migrate from the tissues to T-cell zones of lymph nodes. This is mediated, at least in part, through differential regulation of DC chemokine receptors such as CCR1, CCR5, and CCR7 (Table 3). DC interactions with lymphocytes DCs initiate or “prime” T-cell responses in secondary lymphoid organs such as lymph nodes, spleen, or mucosal lymphoid tissues.62-64 Effective priming of naive T cells is manifested by their Table 2. DC maturation stimuli Classes TNF family molecules TLR agonists Examples Receptor on DC TNF␣ (TNF) TNFR (TNFRSF1A) CD40L (CD154, TNFSF5) CD40 (TNFSFR5) FasL (TNFSF6) Fas (TNFRSF6) TRANCE (TNFSF11) RANK (TNFRSF11A) Notes TNF family molecules are found on a variety of activated immune cells. LIGHT (TNFSF14) HVEM (TNFRSF14) Bacterial lipopeptides TLR1 Pathogen-associated peptidoglycans, lipoproteins, glycolipids, TLR2 breakdown products of host extracellular dsRNA, polyl:C TLR3 antimicrobial peptides. Imiquimod and LPS, Hsp60, Hsp70, oligosaccharides of hyaluronan, -defensins TLR4 R848 are synthetic antiviral compounds Flagellin TLR5 (imidazoquinolines). Imiquimod TLR7 R-848, ssRNA31,32 TLR7, TLR8 Oligosaccharides of hyaluronan are matrix. -defensins are host-derived Hsp70 CpG DNA, HSV DNA33 TLR9 IL-1 IL-1R IL-6 IL-6R Growth factors TSLP IL-7R␣/TSLPR heterodimer Interferons IFN␣35 IFNAR1 Adhesion molecules Agonistic antibody CEACAM-1 (CD66a) — Costimulatory Agonistic antibody B7-DC36 — Immune complexes Opsonized antigens Fc receptors Microbes Viruses (Influenza, HIV) Cytokines — Expressed by inflamed epithelial cells and stimulates Th2 responses.34 — molecules Triggers signaling via Syk kinase37 — — — — Bacteria Activated lymphocytes CD4⫹ and CD8⫹ T cells NK cells NKT cells V␦1⫹ ␥:␦ T cells Other Uric acid38 ? Necrotic cells Agonistic antibody Uric acid is released from dying cells. Stimulation of TREM2 on DCs triggers TREM2 signaling via DAP12.39 TNF indicates tumor necrosis factor; TNFRSF, TNF receptor superfamily; CD40L, CD40 ligand; TNFSF, TNF ligand superfamily; TRANCE, TNF-related activation-induced cytokine; RANK, receptor activator of NF-B (nuclear factor-B); LIGHT, homologous to lymphotoxins, exhibits inducible expression, and competes with HSV glycoprotein D for HVEM, a receptor expressed by T lymphocytes; HVEM, Herpes virus entry mediator; TLR, Toll-like receptor; Hsp, heat shock protein; dsRNA, double-stranded viral RNA; LPS, bacterial lipopolysaccharide; ssRNA, single-stranded RNA; CpG DNA, bacterial unmethylated CpG motif DNA; HSV, Herpes simplex virus; TSLP, thymic stromal lymphopoietin; Th2, T helper cell type 2; IFN, interferon; CEACAM-1, CEA (carcinoembryonic antigen)—related cell adhesion molecule 1; TREM, triggering receptor expressed on myeloid cells; DAP12, DNAX activation protein 12. From www.bloodjournal.org by guest on August 9, 2017. For personal use only. 2238 O’NEILL et al BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 including OX40 and 4-1BB, which may be critical for both initiating and sustaining long-lived T-cell immunity (Figure 2).71-75 DCs also interact directly with B cells and lymphocytes of the innate immune system. Activated MDCs can directly induce B-cell proliferation, immunoglobulin isotype switching, and plasma cell differentiation through the production of the B-cell activation and survival molecules BAFF (B-cell–activating factor belonging to the TNF family) and APRIL (a proliferation-inducing ligand),76-78 and activated PDCs can induce the differentiation of CD40activated B cells into plasma cells through the secretion of IFN␣/ and IL-6.79 DCs can also activate and induce the expansion of resting NK cells by mechanisms that are just beginning to be understood. Requirements for direct cell contact or soluble factors have both been described.80 Activated NK cells can kill immature, but not mature, DCs and can stimulate DCs to induce protective CD8⫹ T-cell responses.80,81 Finally, DCs presenting the synthetic glycolipid ␣-galactosylceramide (␣-GalCer) on CD1d can activate NKT cells to produce IFN␥ and promote resistance to tumors.82 Activated NKT cells can in turn rapidly induce the full maturation Figure 2. Molecules involved in the immunologic synapse between DCs and T cells. Molecules expressed on DCs are listed on the left, with their corresponding T-cell ligands listed on the right. In the right-hand column, stimulatory interactions are indicated in black text and inhibitory interactions are indicated in red. Signaling from T cells to DCs also occurs but is not shown here. Initial DC–T-cell interactions are mediated by adhesion molecules and semaphorins such as neuropilin-1.50 Following engagement of the T-cell receptor by pMHC complexes (signal 1) and engagement of CD28 by B7-1 and B7-2 (signal 2), additional molecules are up-regulated on both cell types that determine the nature of the ensuing T-cell response. Up-regulated molecules include semaphorins such as SEM4-A and members of the B7, CD28, TNF, and TNFR families of costimulatory molecules. Bidirectional signaling between these molecules results in either further T-cell activation or in attenuation of the T-cell response, depending on the molecules involved. Both B7-H1 and B7-DC interact with PD-1 to inhibit activated T cells, but B7-DC can also work synergistically with B7-1 and B7-2 to enhance T-cell activation through an unknown receptor.51,52 B7x transmits an inhibitory signal by way of BTLA (B and T lymphocyte attenuator),53 and B7-H3 can also transmit an inhibitory signal but through an unknown receptor.54 Thromboxane A2 (TXA2) secreted by the DCs also attenuates the DC–T-cell interaction by way of the thromboxane receptor (TP) on the T cell.55 Inhibitory molecules are thought to prevent excessive inflammation and autoimmunity. Alternative names for B7 family members are CD80 (B7-1), CD86 (B7-2), PD-L1 (B7-H1), PD-L2 (B7-DC), B7-H2 or ICOSL (B7RP-1), and B7-H4 (B7x). clonal expansion and differentiation into memory cells and cytokinesecreting effector cells. The strength of the T-cell response is dependent on many factors, including the concentration of antigen on the DC, the affinity of the T-cell receptor for the corresponding pMHC, the state of DC maturation, and the type of maturation stimulus.65 For example, T-cell stimulation by immature DCs leads to initial T-cell proliferation but only short-term survival (“abortive proliferation”), whereas stimulation by mature DCs results in long-term T-cell survival and differentiation into memory and effector T cells.65 Enhanced survival following priming, referred to as T-cell “fitness,” is characterized by resistance to cell death in the absence of cytokines and by responsiveness to the “homeostatic” cytokines IL-7 and IL-15, which promote T-cell survival in the absence of antigen.65,66 Importantly, recent observations indicate that CD4⫹ T-cell help at the time of priming is required to generate CD8⫹ T-cell memory.67-69 This effect is thought to be mediated by CD40CD40L interactions between CD4⫹ T cells and DCs.70 Other T-cell surface molecules are also involved in the generation of long-lived T-cell responses and T-cell memory and have corresponding ligands that are up-regulated on activated APCs such as DCs.71 Examples include members of the TNF receptor superfamily, Figure 3. DC plasticity in response to different maturation stimuli directs Th polarization. DCs can direct the fate of naive CD4⫹ T cells, depending on the type of DC maturation stimulus. Following priming, CD4⫹ T cells may differentiate toward T-helper 1 (Th1) cells, which produce IFN␥ and support CD8⫹ cytotoxic T lymphocyte (CTL) responses, or toward T-helper 2 (Th2) cells, which produce IL-4, IL-5, and IL-13, support humoral immunity, and down-regulate Th1 responses. The direction of Th polarization is determined by the secreted cytokine profile of the stimulating DCs, which in turn depends on the DC subtype, the anatomic location of the DCs, and the type of maturation stimulus.45,56 These factors control other characteristics of the T-cell response as well, such as tolerance induction57 or T-cell homing.58,59 Th1polarizing stimuli such as LPS or flagellin direct a DC differentiation program that causes the DCs to secrete IL-12p70, which together with IFN␥ potently induce CD4⫹ T cells to differentiate into IFN␥-secreting Th1 effector cells. This T-cell program is mediated largely by the transcription factors signal transducer and activator of transcription 4 (Stat4) and T-bet.60,61 Th1 polarization can also be induced in the absence of IL-12p70 by mechanisms that are not entirely known but may be due in part to IL-12–related cytokines such as IL-27. Other DC maturation stimuli such as cholera toxin or schistosome eggs can differentiate DCs that do not produce IL-12p70 and that, in the presence of IL-4, induce naive CD4⫹ T cells to differentiate into IL-4–secreting Th2 effector cells. It is not clear whether Th2 polarization is induced by specific DC cytokines or is rather a default program carried out in the absence of a Th1 polarization signal from the DCs. However, DC secretion of chemokines such as thymus and activation-regulated chemokine (TARC) and MDC can act to potentiate a Th2 response by preferentially attracting Th2 cells. The Th2 program in CD4⫹ T cells is dependent on transcription factors GATA-3 and c-Maf.60,61 From www.bloodjournal.org by guest on August 9, 2017. For personal use only. BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 MANIPULATING DC BIOLOGY FOR IMMUNOTHERAPY 2239 Table 3. DC chemokine receptors and chemokines Receptors CCR1 and CCR5 Receptor expression on DCs Immature DCs Ligands CCL3 (MIP-1␣) Ligand expression by DCs Mature DCs (some stimuli only) CCL4 (MIP-1) Notes CCL3, 4, and 5 are also secreted by activated inflammatory cells and activated endothelium CCL5 (RANTES) CCR2 Immature DCs CCL2 (MCP-1) Not expressed CCL2 is secreted by activated monocytes, CCR4 Immature DCs CCL17 (TARC) Mature DCs (some stimuli only) CCR4 is preferentially expressed on Th2 cells, and CCR6 Immature DCs (some subsets only) CCL20 (MIP-3␣) Not expressed CCL20 is expressed by activated monocytes and CCR7 Mature DCs CCL19 (MIP-3) Not expressed CCL19 and 21 are expressed in T-cell zones of lymph macrophages and endothelium CCL22 (MDC) TARC and MDC can promote Th2 responses. lymphocytes CCL21 (SLC) nodes and spleen, lymphatic endothelium, and lymph node high endothelial venules CXCR1 and CXCR2 Immature DCs CXCL8 (IL-8) Mature DCs IL-8 is a mediator of neutrophil recruitment CXCR3 Not expressed CXCL10 (IP-10) Mature DCs (some stimuli only) IP-10 attracts IFN␥-producing T cells, which express CXCR4 Mature ⬎ Immature CXCL12 (SDF-1) Not expressed SDF-1 is a chemoattractant for lymphocytes and CXCR3, and can promote Th1 responses. monocytes. CXCR4 is also a coreceptor for T-cell-trophic strains of HIV CCR indicates CC motif chemokine receptor; CCL, CC motif chemokine ligand; MIP, macrophage inflammatory protein; RANTES, regulated on activation, normally T-expressed, and presumably secreted; MCP, monocyte chemoattractant protein; TARC, thymus and activation-regulated chemokine; MDC, macrophage-derived chemokine; SLC, secondary lymphoid chemokine; CXCR, CXC motif chemokine receptor; CXCL, CXC motif chemokine ligand; IP-10, IFN␥-inducible 10-kDa protein; SDF-1, stromal cell-derived factor 1; HIV, human immunodeficiency virus. of DCs and can directly interact with DCs to enhance both CD4⫹ and CD8⫹ T-cell responses.82,83 DC induction of immune tolerance Antigen presentation by immature DCs is considered to be an important pathway by which tolerance to self-antigens is maintained. Antigens targeted to immature DCs in vivo can induce tolerance through abortive proliferation and anergy of antigenspecific T cells, whereas simultaneous delivery of a DC maturation stimulus induces a full effector T-cell response (Figure 4).20,84,96 Immature DCs can also induce tolerance through the induction of CD4⫹ and CD8⫹ regulatory T (Tr) cells that suppress immune responses by way of secretion of cytokines such as IL-10 and TGF (Figure 4).97-99 This is in contrast to “naturally occurring” CD4⫹ Tr cells produced in the thymus, which constitutively express CD25 (IL-2R␣), CTLA-4, and Foxp3, and exert their immunosuppressive effect in a cell contact-dependent manner.84-86,97,100,101 Mature DCs can inhibit naturally occurring Tr cells Figure 4. Tolerogenic DCs. There are a number of pathways by which immature MDCs can be rendered tolerogenic. Some of these mechanisms may overlap. The 5 mechanisms summarized here (from left to right) include antigen presentation by resting (steady-state) DCs; exposure of DCs to “modulating” cytokines such as IL-10 and transforming growth factor- (TGF) or to other modulating substances such as corticosteroids and vitamin D3; targeted inhibition of the RelB transcription factor (which controls CD40 expression) or direct inhibition of CD40; DC exposure to CD8⫹CD28⫺ regulatory T (Tr) cells (which have been associated with graft tolerance in patients who received transplants); and the induction of indoleamine 2,3-dioxygenase (IDO)–expressing DCs (IDO-DCs) by ligating B7-1 and B7-2 molecules on the DC with a cytotoxic T-lymphocyte–associated antigen 4 (CTLA-4)–immunoglobulin fusion protein or by CD4⫹CD25⫹ Tr cells. Immature DCs (left) can induce tolerance through the induction of abortive proliferation and anergy, as well as through the induction of IL-10–producing Tr cells.84,85,86 DCs that have been modulated by factors such as IL-10 or TGF (second from left) may also lead to the inhibition of effector T-cell expansion and the induction of IL-10–secreting Tr cells.87,88 Stat3 signaling within the DC appears to be critical for this effect.89 IL-10–producing Tr cells are also induced by DCs that are deficient in RelB or CD4090 (center). CD8⫹CD28⫺ Tr cells induce tolerogenic DCs by up-regulating inhibitory receptors immunoglobulin-like transcript 3 (ILT3) and ILT4 on the DC surface, which ultimately leads to decreased DC expression of B7-1 and B7-2 and T-cell anergy91 (second from right). Finally, IDO-DCs (right) inhibit T-cell expansion and induce T-cell apoptosis by way of IDO-mediated tryptophan catabolism within the DCs.92-94 Both MDCs and PDCs can also be rendered tolerogenic by factors secreted by malignant tumors (not shown).95 From www.bloodjournal.org by guest on August 9, 2017. For personal use only. 2240 O’NEILL et al through the production of IL-6.102 DC expression of CD40 may be an important factor in determining whether T-cell priming will result in immunity or Tr cell–mediated immune suppression. Antigen-exposed mouse DCs which lack CD40 prevent T-cell priming, suppress previously primed immune responses, and induce IL-10–secreting CD4⫹ Tr cells.90 DCs may actively be rendered tolerogenic by a number of mechanisms. In humans, a subset of monocyte-derived DCs has been described that expresses indoleamine 2,3-dioxygenase (IDO), inhibits T-cell proliferation, and induces T-cell death.92 IDO can be induced in DCs by ligation of their B7 molecules with CTLA-493,94 (Figure 4). Large numbers of “IDO DCs” can be found in tumor-draining lymph nodes, suggesting that they may be involved in the immunologic unresponsiveness seen in cancer patients.92 DCs may also be rendered tolerogenic by naturally occurring CD8⫹CD28⫺ Tr cells, which up-regulate inhibitory receptors on DCs and disrupt CD40-induced B7-1 and B7-2 expression91 (Figure 4). Finally, DCs can be rendered tolerogenic in culture by the presence of IL-10, TGF, vitamin D3, or corticosteroids (Figure 4).87 DC Stat3 activity may be critical to the induction of antigen-specific T-cell tolerance. Stat3 is activated by tyrosine phosphorylation following DC exposure to IL-10 and other factors produced by tumor cells, and forced expression of activated Stat3 in DCs can result in impaired antigen-specific T-cell responses.89 Manipulation of DCs for cancer immunotherapy Current approaches to DC vaccine design The most common approach to using DCs for vaccines is to prepare large numbers of autologous mature MDCs ex vivo, load them with antigens, and inject them back into the subject (Figure 5).103,104 BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 Three general methods have been described, involving, respectively (1) differentiating DCs from leukapheresis-derived monocytes with GM-CSF and IL-4105,106 (the most popular approach; IL-13 has been used by some groups in place of IL-4), (2) GM-CSF and TNF␣–mediated differentiation of CD34⫹ hematopoietic progenitor cells into mixtures of interstitial DCs and Langerhans cells107 (Flt3L or stem cell factor may be added to expand DC progenitors, and differentiation may be skewed toward Langerhans cells by adding TGF to the culture108), or (3) directly isolating DCs from leukapheresis products by density gradient centrifugation109 or with commercially available closed systems that use immunomagnetic beads. The yields of both plasmacytoid and classic myeloid-type DCs purified from blood can be significantly enhanced by stimulating patients with Flt3L prior to leukapheresis,110 although pharmaceutical-grade Flt3L is not currently available. All 3 types of DC preparations can stimulate antigen-specific T-cell responses in human subjects and have been associated with clinical responses in cancer patients. No direct comparisons have been performed in clinical trials, although one such trial is currently in progress. DCs are frequently matured in culture prior to injection. Currently, many laboratories using monocyte-derived DCs induce maturation by the addition of a “cocktail” of IL-1, IL-6, TNF␣, and PGE2.111 Several groups have observed that DCs matured in this manner do not secrete detectable bioactive IL-12p70, but still express CCR7 and induce Th1 and CD8⫹ T-cell responses.112,113 How the DCs induce these T-cell responses is currently under investigation. The choice of tumor antigen is important to consider (Table 4). Because vaccines may select for tumor cells that escape immune detection by loss of target antigen expression, antigens critical to tumor growth are preferred. MHC-restricted peptide antigens are frequently used, including altered or enhanced peptides that boost Figure 5. Clinical DC vaccines. There are many alternative approaches for the preparation and use of DC vaccines to treat cancer. DCs may be prepared ex vivo following 3 general methods (lower left), each of which results in a different mixture of cells. DCs may be matured and loaded with antigens using a variety of techniques. Some of these techniques include the addition of DC survival factors, the use of substances that induce crosspresentation, or the use of stimulators of other innate immune cells such as NKT cells. DCs loaded with RNA or DNA can take advantage of sequences encoding cytokines, chemokines, or DC survival factors. Numerous variables such as antigen dose, cell dose, and route of administration also need to be optimized. In addition, less costly and labor-intensive approaches that stimulate and load DCs with antigen in situ are being explored. There are many unresolved issues regarding the monitoring of immune responses and in correlating these responses with clinical outcome. DC vaccines may prove most effective in the adjuvant setting or in combination with other treatments. From www.bloodjournal.org by guest on August 9, 2017. For personal use only. BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 MANIPULATING DC BIOLOGY FOR IMMUNOTHERAPY 2241 Table 4. Classes of tumor antigens Category Cancer-testis (CT) antigens Examples MAGE-1 (MAGEA1) Notes Expressed in germ cells, germinal tissues, tumor cells. BAGE GAGE-1 (GAGE1) NY-ESO-1 (CTAG1) Lineage-specific antigens Melanocyte antigens: Expressed in specific tissues or cells. -Tyrosinase (TYR) -Melan-A/MART-1 (MLANA) -gp100/Pmel17 (SILV) Tumor-specific altered gene HER-2/neu (ERBB2) Associated with a wide variety of tumors. KRAS2 is mutated in 30% to 40% of products (amplified, p53 (TP53) colorectal cancers, and p53 is mutated in up to 70% of all human cancers. Altered aberrantly expressed, Ras genes (KRAS2, HRAS, NRAS) MUC1 glycosylation is seen in a variety of adenocarcinomas, and these altered overexpressed or Mucin 1 (MUC1) glycopeptides can be presented by DCs to T cells.114 Myeloid leukemia cells can mutated genes, splice Beta-catenin (CTNNB1) potentially be differentiated into DCs to vaccinate against endogenously expressed variants, gene fusion MUM1 (IRF4) leukemia-specific antigens.115 products, etc) CDK4 BCR-ABL fusion products N-acteylglucosaminyltranferase V (MGAT5) Survivin (BIRC5) TERT CEA AFP Immunoglobulin idiotypes Multiple myeloma B-cell lymphoma Viral antigens HPV E6 and E7 proteins EBV (HHV4) LMP1 and LMP2 proteins Unique, tumor-specific idiotypes because of clonal rearrangements of immunoglobulin genes. Associated with B-cell malignancies. May be used for tumors such as cervical cancer that are induced by oncogenic viruses. A useful web site with links to current cancer antigen databases may be found at: http://www.cancerimmunity.org/statics/databases.htm. TERT indicates telomerase reverse transcriptase; AFP, alpha-fetoprotein; HPV, human papillomavirus; EBV, Epstein-Barr virus; LMP1, latent membrane protein 1. immunity to less immunogenic self-antigens or that improve antigen presentation or T-cell receptor affinity.3,104,107,110,116,117 A disadvantage to using peptides is that they must be compatible with the HLA type of the patient, often restricting peptide vaccination studies to individuals with common HLA types. In addition, the half-life of pMHC complexes may be short, and competition may prevent priming to lower-affinity epitopes when mixtures of peptides are used. DCs may be loaded with purified or recombinant proteins, transduced with nonreplicating recombinant viral vectors, or transfected with RNA or, less commonly, plasmid vectors encoding tumor-associated antigens.118-122 All of these approaches allow the host’s MHC molecules to select epitopes from an antigen’s entire amino acid sequence (Figure 5). Immunogenicity may be enhanced by using antigens coupled to or expressing other more immunogenic molecules such as foreign proteins (eg, keyhole limpet hemocyanin [KLH]), cytokines (IL-12, IL-15),118,123 costimulatory molecules (B7-2, CD40L), or chemokines (CCL21). DCs may also be loaded with whole tumor cells or tumor cell lysates or be transfected with whole tumor RNA, which permit vaccination with the complete antigenic content of the tumor.103,124-126 Studies to compare routes and frequency of injection, DC dose, and DC subset will be essential to optimize DC immunotherapy. DC vaccines may be stored frozen prior to vaccination103,104 and are typically injected intradermally, subcutaneously, or intravenously in numbers ranging from 2 to 100 million cells. Route of administration may directly affect the nature of T-cell priming. Skin injections may be required to induce immunity to cutaneous tumors, whereas intravenous injections may be less effective at Th1 induction but more effective at induction of humoral immunity.127,128 Injection into lymph nodes or lymphatics has also been attempted, because only 5% or fewer DCs may migrate to draining nodes following subcutaneous injection. Direct injection into tumors is also being investigated. Lessons learned from early DC vaccine trials DC vaccines have minimal side effects and have induced antigenspecific cytotoxic T lymphocyte (CTL) and Th1 responses in healthy volunteers and in patients with a variety of advanced cancers.5,9 Most of the trials in cancer patients have focused on the safety and immunogenicity of DC vaccines and were not designed to evaluate clinical responses. Larger controlled trials are now under way to objectively assess clinical efficacy by documenting responses following standardized criteria such as World Health Organization (WHO) or Response Evaluation Criteria in Solid Tumors Group (RECIST) guidelines.129 Some fundamental lessons have been learned from the smaller, published DC vaccine trials, although they have not led to a consensus on optimal antigen source or dose, DC dose, DC subset or frequency, or route of administration. Most investigators now avoid intravenous administration, as studies have suggested that subcutaneous or intradermal vaccination leads to improved DC migration to lymph nodes127 and enhanced Th1 polarization.128 Importantly, several studies indicate that DCs need to be matured to effectively generate antigen-specific immune responses in humans. Injection of healthy volunteers with antigen-loaded immature DCs has been associated with tolerogenic responses,99 and a randomized trial in patients with metastatic melanoma comparing peptidepulsed immature DCs with peptides administered with adjuvant and GM-CSF demonstrated significantly lower immunogenicity in patients receiving the DC vaccine.130 In addition, a direct comparison of peptide-loaded immature and mature DCs in patients with metastatic melanoma showed that only mature DCs induced antigen-specific CTL responses.98 The availability of sensitive and specific techniques to monitor the induction of antigen-specific T-cell responses has provided insight into the capacity of DCs to induce primary responses to tumor antigens. For instance, it is clear that DC immunization can From www.bloodjournal.org by guest on August 9, 2017. For personal use only. 2242 O’NEILL et al elicit Th1 and CD8⫹ T-cell responses specific to the immunizing antigens (as measured by enzyme-linked immunospot [ELISPOT], lymphocyte proliferation, cytolytic assays, and peptide-MHC tetramer staining), with a suggestion of epitope or antigen spreading in some cases.131,132 However, immune responses in most studies have been weak or undetectable, and durability has not been clearly established. Correlation with tumor regression or disease stabilization has been variable and needs to be established in larger trials. Furthermore, a general lack of standardization makes results difficult to assess or compare, especially when we have little concept of what magnitude of response correlates with protective immunity. The results of DC-based clinical trials have been extensively reviewed5,9; therefore, our comments are limited to more recent studies. Although it is difficult to compare the results of the DC vaccine trials published to date, in our opinion the most impressive objective clinical responses have been associated with the use of whole proteins, killed tumor cells, or tumor lysates. This may be because these are exogenous antigen sources that target MHC II to generate CD4⫹ T-cell help and also target MHC I by way of cross-presentation to generate CD8⫹ CTLs. Using tumor-specific idiotype immunoglobulin-pulsed DCs in patients with follicular lymphoma, Timmerman et al118 reported 2 long-lasting complete responses (CRs) and 1 partial response (PR) among 10 patients with measurable disease in the pilot phase of the study. An additional 25 patients were vaccinated after their best clinical response was achieved by chemotherapy, and objective tumor regression was seen in 4 of 18 patients with residual disease. Holtl et al125 reported a trial of 35 patients with metastatic renal cell carcinoma who received monthly injections of autologous, mature monocyte-derived DCs loaded with tumor lysates. Of 27 evaluable patients, 2 had objective CR (as per WHO), 1 had a PR, and 7 had stable disease. Objective responses and disease stabilization were long lasting, ranging from 6 months to 3 years. Durable CRs were also reported by O’Rourke et al126 in a trial of 17 patients with metastatic melanoma who received mature monocyte-derived DCs loaded with autologous irradiated tumor cells. By WHO criteria there were 3 CRs (with durable remissions of over 3 years) and 3 PRs among 12 patients who completed the vaccinations. One patient with progressive disease was vaccinated every 6 weeks for more than 3 years, indicating that maintenance vaccinations may be useful even for patients with slowly progressive disease. Finally, another promising trial using autologous tumor lysate pulsed DCs showed objective responses in patients with refractory cutaneous T-cell lymphoma after intranodal vaccination.133 Larger studies will be important to confirm the results of the these trials. New vaccine strategies that exploit DC biology Tumors can evade immunity by a number of mechanisms, including mutations in genes encoding target antigens, loss of antigen expression, or immunosuppressive maneuvers such as secretion of TGF.1 This may be particularly true of large or metastatic tumors. Thus, DC vaccines may be most effective in the adjuvant setting for patients in remission but with a high risk of recurrence. However, DC biology may be exploited in many ways to generate more effective immunotherapies, and multimodality approaches may be used to enhance the effectiveness of these vaccines. Below we discuss some novel applications studied in murine models or in preclinical studies using human cells. BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 Provision of CD4ⴙ T-cell help for CD8ⴙ T cells Vaccination studies in mice using MHC II–deficient DCs,134 as well as experiments that demonstrate the importance of CD4⫹ T cell help to generate CD8⫹ T-cell memory,67-69 call into question vaccine strategies that target only CD8⫹ T-cell responses. Peptideloaded DC vaccines should incorporate antigens targeting both CD4⫹ and CD8⫹ T cells, and a polyvalent approach should be considered. Peptide-loaded dendritic cells can clearly prime CD4⫹ T-cell responses,131 but a more practical approach that circumvents the problems of HLA-restricted peptides may be to target crosspresentation. For example, targeting antigens to Fc receptors on DCs using antibody-antigen complexes has been shown to activate both CD4 and CD8 effector responses and tumor immunity in mice.135 Coating myeloma cells with anti–syndecan-1 antibody similarly promotes cross-presentation.136 Pharmaceutical-grade antibodies already in use to treat human cancer (eg, anti-CD20, anti-HER-2/neu) may act in part through this mechanism and could be used in the preparation of DC vaccines. Cross-presentation can also be enhanced by targeting DC surface receptors such as DEC-205,20 loading DCs with killed cells or cell lysates or by stimulating DCs with TLR agonists that induce cross presentation49 (Figure 5). Transfected RNA, which primarily targets MHC I, may also be targeted to MHC II by incubating the transfected DCs with antisense oligonucleotides to the MHC II–associated Ii protein137 or by using fusion constructs carrying an endosomal/lysosomal sorting signal.138 Strategies to recruit, mature, and load DCs in situ Existing DC vaccine methods require expensive facilities and labor-intensive cell processing. To avoid this, alternative approaches that simultaneously recruit, mature, and pulse DCs with antigens in vivo are being explored (Figure 5). To recruit DCs, locally implanted chemokines such as MIP-3 may be used to condition the injection site prior to vaccination.139 To mature DCs, simple vaccines that take advantage of CpG motif DNA (a TLR9 agonist) coinjected with or conjugated to a protein antigen have been used.140,141 Vaccination with heat shock protein-peptide complexes can similarly mature DCs in situ and may induce immunologic and clinical responses in melanoma patients.142 Another in situ approach uses CpG motif-containing DNA vaccines that encode tumor antigens. These vaccines can be engineered to carry xenogeneic antigens143 or to include DCspecific promoters to specifically target antigen expression to DCs. DNA vaccines may also be designed to drive the expression of survival factors such as Bcl-xL144 or to encode DC maturation signals145 or immunostimulatory cytokines.146 Certain microbes directly induce MDC or PDC maturation, even in nonreplicating form, and are being tested as recombinant vaccine vectors.3 One advantage of some viral vectors is that IFN␣ generated from virus-stimulated DCs may promote cross-priming of CD8⫹ T-cell responses.35 Microbial vectors may also be engineered to express adhesion molecules, costimulatory molecules, or cytokines that direct Th polarization, promote T-cell activation and longevity, and promote DC survival (Figure 5). Ex vivo–derived DCs can also be matured in situ by preconditioning the injection site with TLR agonists,147 and DC migration can be enhanced by preconditioning the injection site with cytokines or with DCs themselves.7,148,149 This approach may be preferable to ex vivo maturation, because DC cytokines such as IL-12 are often expressed only briefly after exposure to many From www.bloodjournal.org by guest on August 9, 2017. For personal use only. BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 MANIPULATING DC BIOLOGY FOR IMMUNOTHERAPY maturation stimuli, and local production of cytokines and chemokines induced by local application of TLR agonists may also promote DC viability and migration to draining lymph nodes. Strategies to activate NKT cells Vaccination with melanoma cells in adjuvant can activate CD1drestricted NKT cells that recognize tumor-associated gangliosides,150 and intravenous delivery of a soluble antigen together with the synthetic CD1d-binding glycolipid ␣-GalCer can lead to in vivo activation of NKT cells and induction of antitumor T-cell immunity.83 Trials to test the activating potential of ␣-GalCer– pulsed DCs are under way in cancer patients. Inhibition of immune tolerance One way to enhance cancer vaccines is to simultaneously block inhibitory costimulatory molecules or Tr cells. For example, administering an inhibitory antibody to CTLA-4 in previously vaccinated cancer patients can result in effective antitumor immunity.151 Possible synergy of CTLA-4 blockade and concomitant tumor antigen vaccination has been observed in patients with metastatic melanoma.152 In this study, tumor regressions were accompanied by significant toxicity, including severe or lifethreatening autoimmunity. Nevertheless, this approach is worth addressing in conjunction with DC vaccines, using different dosages or schedules to alleviate toxicity. In mice, blockade of the inhibitory costimulatory molecule B7-H1 has also been shown to improve DC-mediated antitumor T-cell responses.51 The activity of cancer vaccines may be enhanced through depletion or inhibition of Tr cells through the use of cytotoxic anti-CD25 antibodies or IL-2 coupled to cytotoxic molecules. In mice with poorly immunogenic tumors, depletion of Tr cells alone can slow tumor growth but does not efficiently reject the tumor.153 However, immune responses induced by antigen-pulsed mature DCs are significantly enhanced in CD25-depleted mice.154 The use of both of CTLA-4 blockade and CD25⫹ cell depletion may further potentiate the effectiveness of vaccines.155 Combination therapies Multimodality approaches incorporating tumor vaccination have also shown promise in animal models, although it may prove difficult to translate some of these approaches into clinical use. For example, in one study the combination of vaccination with adoptively transferred T cells and administration of IL-2 resulted in tumor regression and long-term cures.156 Using another approach, Cui et al157 showed that transducing hematopoietic progenitor cells with a model tumor antigen and transplanting these cells into irradiated recipient mice resulted in expression of the antigen in 2243 donor-derived DCs in the host’s lymphoid organs. When combined with systemic agents that generate and activate DCs and adoptive transfer of donor T cells, this treatment resulted in expansion of antigen-specific T cells and successful treatment of the antigenbearing tumor. Antitumor vaccination in combination with therapies that target the tumor’s vascular supply have also shown promise in mouse models,158 as has vaccination during lymphoid recovery following bone marrow transplantation.159 Use of “regulatory DCs” for the induction of transplantation tolerance DC-based immunotherapy may also prove to be a highly selective way to induce graft tolerance in organ or hematopoietic stem cell transplantation or to induce tolerance in patients with autoimmune disease. Studies in mice and humans have shown that tolerogenic or “regulatory DCs” (rDCs) may be induced ex vivo by culturing immature DCs in modulating cytokines or growth factors such as IL-10 and TGF. In a mouse model for the treatment of leukemia, rDCs have been used to treat acute graft-versus-host disease and leukemia relapse in conjunction with allogeneic bone marrow transplantation.88 Conclusion DC-based immunotherapy is still in its infancy. Two-arm trials are needed to assess the efficacy of DC vaccines compared with other immunotherapies and to optimize the use of DCs for vaccines. Until then, it is not truly meaningful to compare DC immunotherapy with standard cancer therapies in large randomized trials. The greatest clinical benefit of DC immunotherapy for cancer may be found in the adjuvant setting, although it is hoped that patients with advanced cancer will also benefit, at the very least, through disease stabilization. Eventually, it is possible that the most effective DC therapies may not necessarily involve the ex vivo manipulation of DCs. Multimodality approaches that include novel biologic agents may also help achieve effective, durable antitumor immune responses. With greater understanding of DC biology and of mechanisms to enhance DC immunogenicity, the answers will begin to come. Acknowledgments We thank Teresita O’Neill for assistance with artwork; Stephen Schachterle for preparation of dendritic cells; and Giorgio Inghirami, Marie Larsson, Anne-Sophie Beignon, and Mojca Skoberne for helpful advice. References 1. Dunn GP, Bruce AT, Ikeda H, Old LJ, Schreiber RD. Cancer immunoediting: from immunosurveillance to tumor escape. Nat Immunol. 2002;3:991998. 2. Zhang L, Conejo-Garcia JR, Katsaros D, et al. Intratumoral T cells, recurrence, and survival in epithelial ovarian cancer. N Engl J Med. 2003; 348:203-213. approaches to human cancer immunotherapy. J Leukoc Biol. 2003;73:3-29. 10. Shortman K, Liu YJ. Mouse and human dendritic cell subtypes. Nat Rev Immunol. 2002;2:151-161. 6. Banchereau J, Briere F, Caux C, et al. Immunobiology of dendritic cells. Annu Rev Immunol. 2000; 18:767-811. 11. Karsunky H, Merad M, Cozzio A, Weissman IL, Manz MG. Flt3 ligand regulates dendritic cell development from Flt3⫹ lymphoid and myeloidcommitted progenitors to Flt3⫹ dendritic cells in vivo. J Exp Med. 2003;198:305-313. 7. Ardavin C, Amigorena S, Reis e Sousa C. Dendritic cells: immunobiology and cancer immunotherapy. Immunity. 2004;20:17-23. 3. Berzofsky JA, Ahlers JD, Belyakov IM. Strategies for designing and optimizing new generation vaccines. Nat Rev Immunol. 2001;1:209-219. 8. Timmerman JM, Levy R. Dendritic cell vaccines for cancer immunotherapy. Annu Rev Med. 1999; 50:507-529. 12. D’Amico A, Wu L. The early progenitors of mouse dendritic cells and plasmacytoid predendritic cells are within the bone marrow hemopoietic precursors expressing Flt3. J Exp Med. 2003;198:293-303. 4. Finn OJ. Cancer vaccines: between the idea and the reality. Nat Rev Immunol. 2003;3:630-641. 9. Schuler G, Schuler-Thurner B, Steinman RM. The use of dendritic cells in cancer immunotherapy. Curr Opin Immunol. 2003;15:138-147. 13. Fonteneau JF, Gilliet M, Larsson M, et al. Activation of influenza virus-specific CD4⫹ and CD8⫹ T cells: a new role for plasmacytoid dendritic cells 5. Davis ID, Jefford M, Parente P, Cebon J. Rational From www.bloodjournal.org by guest on August 9, 2017. For personal use only. 2244 BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 O’NEILL et al in adaptive immunity. Blood. 2003;101:35203526. 14. Salio M, Cella M, Vermi W, et al. Plasmacytoid dendritic cells prime IFN-gamma-secreting melanoma-specific CD8 lymphocytes and are found in primary melanoma lesions. Eur J Immunol. 2003; 33:1052-1062. 15. Ebner S, Ehammer Z, Holzmann S, et al. Expression of C-type lectin receptors by subsets of dendritic cells in human skin. Int Immunol. 2004;16: 877-887. 16. Cambi A, Figdor CG. Dual function of C-type lectin-like receptors in the immune system. Curr Opin Cell Biol. 2003;15:539-546. 17. Peiser L, Mukhopadhyay S, Gordon S. Scavenger receptors in innate immunity. Curr Opin Immunol. 2002;14:123-128. 18. Delneste Y, Magistrelli G, Gauchat J, et al. Involvement of LOX-1 in dendritic cell-mediated antigen cross-presentation. Immunity. 2002;17: 353-362. 19. Basu S, Binder RJ, Ramalingam T, Srivastava PK. CD91 is a common receptor for heat shock proteins gp96, hsp90, hsp70, and calreticulin. Immunity. 2001;14:303-313. 20. Bonifaz L, Bonnyay D, Mahnke K, Rivera M, Nussenzweig MC, Steinman RM. Efficient targeting of protein antigen to the dendritic cell receptor DEC205 in the steady state leads to antigen presentation on major histocompatibility complex class I products and peripheral CD8⫹ T cell tolerance. J Exp Med. 2002;196:1627-1638. 21. Albert ML, Pearce SF, Francisco LM, et al. Immature dendritic cells phagocytose apoptotic cells via alphavbeta5 and CD36, and cross-present antigens to cytotoxic T lymphocytes. J Exp Med. 1998;188:1359-1368. 22. Guermonprez P, Valladeau J, Zitvogel L, Thery C, Amigorena S. Antigen presentation and T cell stimulation by dendritic cells. Annu Rev Immunol. 2002;20:621-667. 23. Guermonprez P, Saveanu L, Kleijmeer M, Davoust J, Van Endert P, Amigorena S. ERphagosome fusion defines an MHC class I crosspresentation compartment in dendritic cells. Nature. 2003;425:397-402. 24. Fonteneau JF, Larsson M, Bhardwaj N. Interactions between dead cells and dendritic cells in the induction of antiviral CTL responses. Curr Opin Immunol. 2002;14:471-477. 25. Ackerman AL, Kyritsis C, Tampe R, Cresswell P. Early phagosomes in dendritic cells form a cellular compartment sufficient for cross presentation of exogenous antigens. Proc Natl Acad Sci U S A. 2003;100:12889-12894. 26. Moody DB, Porcelli SA. Intracellular pathways of CD1 antigen presentation. Nat Rev Immunol. 2003;3:11-22. 27. Joyce S, Van Kaer L. CD1-restricted antigen presentation: an oily matter. Curr Opin Immunol. 2003;15:95-104. 28. Matzinger P. The danger model: a renewed sense of self. Science. 2002;296:301-305. 29. Fujimoto Y, Tu L, Miller AS, et al. CD83 expression influences CD4⫹ T cell development in the thymus. Cell. 2002;108:755-767. 30. Lechmann M, Berchtold S, Hauber J, Steinkasserer A. CD83 on dendritic cells: more than just a marker for maturation. Trends Immunol. 2002;23:273-275. 31. Diebold SS, Kaisho T, Hemmi H, Akira S, Reis ESC. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science. 2004;303:1529-1531. 32. Heil F, Hemmi H, Hochrein H, et al. Species-specific recognition of single-stranded RNA via tolllike receptor 7 and 8. Science. 2004;303:15261529. 33. Lund J, Sato A, Akira S, Medzhitov R, Iwasaki A. Toll-like receptor 9-mediated recognition of Her- pes simplex virus-2 by plasmacytoid dendritic cells. J Exp Med. 2003;198:513-520. CD4⫹ T cells independent of the PD-1 receptor. J Exp Med. 2003;198:31-38. 34. Soumelis V, Reche PA, Kanzler H, et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat Immunol. 2002;3:673-680. 53. Watanabe N, Gavrieli M, Sedy JR, et al. BTLA is a lymphocyte inhibitory receptor with similarities to CTLA-4 and PD-1. Nat Immunol. 2003;4:670679. 35. Le Bon A, Etchart N, Rossmann C, et al. Crosspriming of CD8⫹ T cells stimulated by virus-induced type I interferon. Nat Immunol. 2003;4: 1009-1015. 54. Suh WK, Gajewska BU, Okada H, et al. The B7 family member B7-H3 preferentially down-regulates T helper type 1-mediated immune responses. Nat Immunol. 2003;4:899-906. 36. Nguyen LT, Radhakrishnan S, Ciric B, et al. Cross-linking the B7 family molecule B7-DC directly activates immune functions of dendritic cells. J Exp Med. 2002;196:1393-1398. 55. Kabashima K, Murata T, Tanaka H, et al. Thromboxane A2 modulates interaction of dendritic cells and T cells and regulates acquired immunity. Nat Immunol. 2003;4:694-701. 37. Sedlik C, Orbach D, Veron P, et al. A critical role for Syk protein tyrosine kinase in Fc receptormediated antigen presentation and induction of dendritic cell maturation. J Immunol. 2003;170: 846-852. 56. Lanzavecchia A, Sallusto F. The instructive role of dendritic cells on T cell responses: lineages, plasticity and kinetics. Curr Opin Immunol. 2001;13: 291-298. 38. Shi Y, Evans JE, Rock KL. Molecular identification of a danger signal that alerts the immune system to dying cells. Nature. 2003;425:516-521. 39. Bouchon A, Hernandez-Munain C, Cella M, Colonna M. A DAP12-mediated pathway regulates expression of CC chemokine receptor 7 and maturation of human dendritic cells. J Exp Med. 2001;194:1111-1122. 40. Takeda K, Kaisho T, Akira A. Toll-like receptors. Annu Rev Immunol. 2003;21:335-376. 41. Kadowaki N, Ho S, Antonenko S, et al. Subsets of human dendritic cell precursors express different toll-like receptors and respond to different microbial antigens. J Exp Med. 2001;194:863-869. 42. Hornung V, Rothenfusser S, Britsch S, et al. Quantitative expression of toll-like receptor 1-10 mRNA in cellular subsets of human peripheral blood mononuclear cells and sensitivity to CpG oligodeoxynucleotides. J Immunol. 2002;168: 4531-4537. 43. Jarrossay D, Napolitani G, Colonna M, Sallusto F, Lanzavecchia A. Specialization and complementarity in microbial molecule recognition by human myeloid and plasmacytoid dendritic cells. Eur J Immunol. 2001;31:3388-3393. 57. Akbari O, DeKruyff RH, Umetsu DT. Pulmonary dendritic cells producing IL-10 mediate tolerance induced by respiratory exposure to antigen. Nat Immunol. 2001;2:725-731. 58. Mora JR, Bono MR, Manjunath N, et al. Selective imprinting of gut-homing T cells by Peyer’s patch dendritic cells. Nature. 2003;424:88-93. 59. Kim CH, Nagata K, Butcher EC. Dendritic cells support sequential reprogramming of chemoattractant receptor profiles during naive to effector T cell differentiation. J Immunol. 2003;171:152158. 60. Usui T, Nishikomori R, Kitani A, Strober W. GATA-3 suppresses Th1 development by downregulation of Stat4 and not through effects on IL12Rbeta2 chain or T-bet. Immunity. 2003;18:415428. 61. Murphy KM, Reiner SL. The lineage decisions of helper T cells. Nat Rev Immunol. 2002;2:933-944. 62. Stoll S, Delon J, Brotz TM, Germain RN. Dynamic imaging of T cell-dendritic cell interactions in lymph nodes. Science. 2002;296:1873-1876. 63. Miller MJ, Wei SH, Parker I, Cahalan MD. Twophoton imaging of lymphocyte motility and antigen response in intact lymph node. Science. 2002;296:1869-1873. 44. Kopp E, Medzhitov R. Recognition of microbial infection by Toll-like receptors. Curr Opin Immunol. 2003;15:396-401. 64. Bousso P, Robey E. Dynamics of CD8⫹ T cell priming by dendritic cells in intact lymph nodes. Nat Immunol. 2003;4:579-585. 45. Agrawal S, Agrawal A, Doughty B, et al. Cutting edge: different toll-like receptor agonists instruct dendritic cells to induce distinct Th responses via differential modulation of extracellular signalregulated kinase-mitogen-activated protein kinase and c-Fos. J Immunol. 2003;171:49844989. 65. Gett AV, Sallusto F, Lanzavecchia A, Geginat J. T cell fitness determined by signal strength. Nat Immunol. 2003;4:355-360. 46. Trombetta ES, Ebersold M, Garrett W, Pypaert M, Mellman I. Activation of lysosomal function during dendritic cell maturation. Science. 2003;299: 1400-1403. 47. Larsson M, Fonteneau JF, Somersan S, et al. Efficiency of cross presentation of vaccinia virusderived antigens by human dendritic cells. Eur J Immunol. 2001;31:3432-3442. 48. Delamarre L, Holcombe H, Mellman I. Presentation of exogenous antigens on major histocompatibility complex (MHC) class I and MHC class II molecules is differentially regulated during dendritic cell maturation. J Exp Med. 2003;198:111122. 49. Datta SK, Redecke V, Prilliman KR, et al. A subset of Toll-like receptor ligands induces crosspresentation by bone marrow-derived dendritic cells. J Immunol. 2003;170:4102-4110. 50. Kikutani H, Kumanogoh A. Semaphorins in interactions between T cells and antigen-presenting cells. Nat Rev Immunol. 2003;3:159-167. 51. Curiel TJ, Wei S, Dong H, et al. Blockade of B7-H1 improves myeloid dendritic cell-mediated antitumor immunity. Nat Med. 2003;9:562-567. 52. Shin T, Kennedy G, Gorski K, et al. Cooperative B7–1/2 (CD80/CD86) and B7-DC costimulation of 66. van Stipdonk MJ, Hardenberg G, Bijker MS, et al. Dynamic programming of CD8⫹ T lymphocyte responses. Nat Immunol. 2003;4:361-365. 67. Shedlock DJ, Shen H. Requirement for CD4 T cell help in generating functional CD8 T cell memory. Science. 2003;300:337-339. 68. Sun JC, Bevan MJ. Defective CD8 T cell memory following acute infection without CD4 T cell help. Science. 2003;300:339-342. 69. Janssen EM, Lemmens EE, Wolfe T, Christen U, von Herrath MG, Schoenberger SP. CD4⫹ T cells are required for secondary expansion and memory in CD8⫹ T lymphocytes. Nature. 2003; 421:852-856. 70. Schoenberger SP, Toes RE, van der Voort EI, Offringa R, Melief CJ. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40L interactions. Nature. 1998;393:480-483. 71. Croft M. Co-stimulatory members of the TNFR family: keys to effective T-cell immunity? Nat Rev Immunol. 2003;3:609-620. 72. Rogers PR, Song J, Gramaglia I, Killeen N, Croft M. OX40 promotes Bcl-xL and Bcl-2 expression and is essential for long-term survival of CD4 T cells. Immunity. 2001;15:445-455. 73. Lee PK, Chang CJ, Lin CM. Lipopolysaccharide preferentially induces 4-1BB ligand expression on human monocyte-derived dendritic cells. Immunol Lett. 2003;90:215-221. 74. Bukczynski J, Wen T, Ellefsen K, Gauldie J, Watts From www.bloodjournal.org by guest on August 9, 2017. For personal use only. BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 TH. Costimulatory ligand 4-1BBL (CD137L) as an efficient adjuvant for human antiviral cytotoxic T cell responses. Proc Natl Acad Sci U S A. 2004; 101:1291-1296. 75. Wiethe C, Dittmar K, Doan T, Lindenmaier W, Tindle R. Provision of 4-1BB ligand enhances effector and memory CTL responses generated by immunization with dendritic cells expressing a human tumor-associated antigen. J Immunol. 2003;170:2912-2922. 76. Litinskiy MB, Nardelli B, Hilbert DM, et al. DCs induce CD40-independent immunoglobulin class switching through BLyS and APRIL. Nat Immunol. 2002;3:822-829. 77. Mackay F, Schneider P, Rennert P, Browning J. BAFF and APRIL: a tutorial on B cell survival. Annu Rev Immunol. 2003;21:231-264. 78. Balazs M, Martin F, Zhou T, Kearney J. Blood dendritic cells interact with splenic marginal zone B cells to initiate T-independent immune responses. Immunity. 2002;17:341-352. 79. Jego G, Palucka AK, Blanck JP, Chalouni C, Pascual V, Banchereau J. Plasmacytoid dendritic cells induce plasma cell differentiation through type I interferon and interleukin 6. Immunity. 2003;19:225-234. 80. Ferlazzo G, Munz C. NK cell compartments and their activation by dendritic cells. J Immunol. 2004;172:1333-1339. 81. Mocikat R, Braumuller H, Gumy A, et al. Natural killer cells activated by MHC class I(low) targets prime dendritic cells to induce protective CD8 T cell responses. Immunity. 2003;19:561-569. 82. Fujii S, Shimizu K, Smith C, Bonifaz L, Steinman RM. Activation of natural killer T cells by alphagalactosylceramide rapidly induces the full maturation of dendritic cells in vivo and thereby acts as an adjuvant for combined CD4 and CD8 T cell immunity to a coadministered protein. J Exp Med. 2003;198:267-279. 83. Hermans IF, Silk JD, Gileadi U, et al. NKT cells enhance CD4⫹ and CD8⫹ T cell responses to soluble antigen in vivo through direct interaction with dendritic cells. J Immunol. 2003;171:51405147. 84. Probst HC, Lagnel J, Kollias G, van den Broek M. Inducible transgenic mice reveal resting dendritic cells as potent inducers of CD8⫹ T cell tolerance. Immunity. 2003;18:713-720. 85. Jonuleit H, Schmitt E, Schuler G, Knop J, Enk AH. Induction of interleukin 10-producing, nonproliferating CD4(⫹) T cells with regulatory properties by repetitive stimulation with allogeneic immature human dendritic cells. J Exp Med. 2000; 192:1213-1222. 86. Roncarolo MG, Levings MK, Traversari C. Differentiation of T regulatory cells by immature dendritic cells. J Exp Med. 2001;193:F5–9. 87. Wakkach A, Fournier N, Brun V, Breittmayer JP, Cottrez F, Groux H. Characterization of dendritic cells that induce tolerance and T regulatory 1 cell differentiation in vivo. Immunity. 2003;18:605617. 88. Sato K, Yamashita N, Baba M, Matsuyama T. Regulatory dendritic cells protect mice from murine acute graft-versus-host disease and leukemia relapse. Immunity. 2003;18:367-379. 89. Cheng F, Wang HW, Cuenca A, et al. A critical role for Stat3 signaling in immune tolerance. Immunity. 2003;19:425-436. 90. Martin E, O’Sullivan B, Low P, Thomas R. Antigen-specific suppression of a primed immune response by dendritic cells mediated by regulatory T cells secreting interleukin-10. Immunity. 2003;18:155-167. 91. Chang CC, Ciubotariu R, Manavalan JS, et al. Tolerization of dendritic cells by T(S) cells: the crucial role of inhibitory receptors ILT3 and ILT4. Nat Immunol. 2002;3:237-243. 92. Munn DH, Sharma MD, Lee JR, et al. Potential regulatory function of human dendritic cells ex- MANIPULATING DC BIOLOGY FOR IMMUNOTHERAPY pressing indoleamine 2,3-dioxygenase. Science. 2002;297:1867-1870. 2245 93. Fallarino F, Grohmann U, Hwang KW, et al. Modulation of tryptophan catabolism by regulatory T cells. Nat Immunol. 2003;4:1206-1212. 111. Jonuleit H, Kuhn U, Muller G, et al. Pro-inflammatory cytokines and prostaglandins induce maturation of potent immunostimulatory dendritic cells under fetal calf serum-free conditions. Eur J Immunol. 1997;27:3135-3142. 94. Mellor AL, Baban B, Chandler P, et al. Cutting edge: induced indoleamine 2,3 dioxygenase expression in dendritic cell subsets suppresses T cell clonal expansion. J Immunol. 2003;171:16521655. 112. Kalinski P, Vieira PL, Schuitemaker JH, de Jong EC, Kapsenberg ML. Prostaglandin E(2) is a selective inducer of interleukin-12 p40 (IL-12p40) production and an inhibitor of bioactive IL-12p70 heterodimer. Blood. 2001;97:3466-3469. 95. Zou W, Machelon V, Coulomb-L’Hermin A, et al. Stromal-derived factor-1 in human tumors recruits and alters the function of plasmacytoid precursor dendritic cells. Nat Med. 2001;7:13391346. 113. Lee AW, Truong T, Bickham K, et al. A clinical grade cocktail of cytokines and PGE(2) results in uniform maturation of human monocyte-derived dendritic cells: implications for immunotherapy. Vaccine. 2002;20(Suppl 4):A8-A22. 97. Jonuleit H, Schmitt E. The regulatory T cell family: distinct subsets and their interrelations. J Immunol. 2003;171:6323-6327. 114. Vlad AM, Muller S, Cudic M, et al. Complex carbohydrates are not removed during processing of glycoproteins by dendritic cells: processing of tumor antigen MUC1 glycopeptides for presentation to major histocompatibility complex class IIrestricted T cells. J Exp Med. 2002;196:14351446. 98. Jonuleit H, Giesecke-Tuettenberg A, Tuting T, et al. A comparison of two types of dendritic cell as adjuvants for the induction of melanoma-specific T-cell responses in humans following intranodal injection. Int J Cancer. 2001;93:243-251. 115. Choudhury A, Gajewski JL, Liang JC, et al. Use of leukemic dendritic cells for the generation of antileukemic cellular cytotoxicity against Philadelphia chromosome-positive chronic myelogenous leukemia. Blood. 1997;89:1133-1142. 99. Dhodapkar MV, Steinman RM, Krasovsky J, Munz C, Bhardwaj N. Antigen-specific inhibition of effector T cell function in humans after injection of immature dendritic cells. J Exp Med. 2001;193: 233-238. 116. Dhodapkar MV, Steinman RM, Sapp M, et al. Rapid generation of broad T-cell immunity in humans after a single injection of mature dendritic cells. J Clin Invest. 1999;104:173-180. 96. Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenic dendritic cells. Annu Rev Immunol. 2003;21:685-711. 100. Francois Bach J. Regulatory T cells under scrutiny. Nat Rev Immunol. 2003;3:189-198. 117. Wang RF, Wang HY. Enhancement of antitumor immunity by prolonging antigen presentation on dendritic cells. Nat Biotechnol. 2002;20:149-154. 101. Sakaguchi S. Control of immune responses by naturally arising CD4⫹ regulatory T cells that express toll-like receptors. J Exp Med. 2003;197: 397-401. 118. Timmerman JM, Czerwinski DK, Davis TA, et al. Idiotype-pulsed dendritic cell vaccination for Bcell lymphoma: clinical and immune responses in 35 patients. Blood. 2002;99:1517-1526. 102. Pasare C, Medzhitov R. Toll pathway-dependent blockade of CD4⫹CD25⫹ T cell-mediated suppression by dendritic cells. Science. 2003;299: 1033-1036. 119. Heiser A, Coleman D, Dannull J, et al. Autologous dendritic cells transfected with prostate-specific antigen RNA stimulate CTL responses against metastatic prostate tumors. J Clin Invest. 2002; 109:409-417. 103. Thumann P, Moc I, Humrich J, et al. Antigen loading of dendritic cells with whole tumor cell preparations. J Immunol Methods. 2003;277:1-16. 104. Schuler-Thurner B, Schultz ES, Berger TG, et al. Rapid induction of tumor-specific type 1 T helper cells in metastatic melanoma patients by vaccination with mature, cryopreserved, peptide-loaded monocyte-derived dendritic cells. J Exp Med. 2002;195:1279-1288. 105. Thurner B, Roder C, Dieckmann D, et al. Generation of large numbers of fully mature and stable dendritic cells from leukapheresis products for clinical application. J Immunol Methods. 1999; 223:1-15. 106. Bender A, Sapp M, Schuler G, Steinman RM, Bhardwaj N. Improved methods for the generation of dendritic cells from nonproliferating progenitors in human blood. J Immunol Methods. 1996;196:121-135. 107. Banchereau J, Palucka AK, Dhodapkar M, et al. Immune and clinical responses in patients with metastatic melanoma to CD34(⫹) progenitorderived dendritic cell vaccine. Cancer Res. 2001; 61:6451-6458. 108. Strobl H, Bello-Fernandez C, Riedl E, et al. flt3 ligand in cooperation with transforming growth factor-beta1 potentiates in vitro development of Langerhans-type dendritic cells and allows single-cell dendritic cell cluster formation under serum-free conditions. Blood. 1997;90:14251434. 109. Hsu FJ, Benike C, Fagnoni F, et al. Vaccination of patients with B-cell lymphoma using autologous antigen-pulsed dendritic cells. Nat Med. 1996;2: 52-58. 110. Fong L, Hou Y, Rivas A, et al. Altered peptide ligand vaccination with Flt3 ligand expanded dendritic cells for tumor immunotherapy. Proc Natl Acad Sci U S A. 2001;98:8809-8814. 120. Engelmayer J, Larsson M, Lee A, et al. Mature dendritic cells infected with canarypox virus elicit strong anti-human immunodeficiency virus CD8⫹ and CD4⫹ T-cell responses from chronically infected individuals. J Virol. 2001;75:2142-2153. 121. Klein C, Bueler H, Mulligan RC. Comparative analysis of genetically modified dendritic cells and tumor cells as therapeutic cancer vaccines. J Exp Med. 2000;191:1699-1708. 122. Larregina AT, Morelli AE, Tkacheva O, et al. Highly efficient expression of transgenic proteins by naked DNA-transfected dendritic cells through terminal differentiation. Blood. 2004;103:811-819. 123. Fong L, Brockstedt D, Benike C, et al. Dendritic cell-based xenoantigen vaccination for prostate cancer immunotherapy. J Immunol. 2001;167: 7150-7156. 124. Su Z, Dannull J, Heiser A, et al. Immunological and clinical responses in metastatic renal cancer patients vaccinated with tumor RNA-transfected dendritic cells. Cancer Res. 2003;63:2127-2133. 125. Holtl L, Zelle-Rieser C, Gander H, et al. Immunotherapy of metastatic renal cell carcinoma with tumor lysate-pulsed autologous dendritic cells. Clin Cancer Res. 2002;8:3369-3376. 126. O’Rourke MG, Johnson M, Lanagan C, et al. Durable complete clinical responses in a phase I/II trial using an autologous melanoma cell/dendritic cell vaccine. Cancer Immunol Immunother. 2003; 52:387-395. 127. Mullins DW, Sheasley SL, Ream RM, et al. Route of immunization with peptide-pulsed dendritic cells controls the distribution of memory and effector T cells in lymphoid tissues and determines the pattern of regional tumor control. J Exp Med. 2003;198:1023-1034. 128. Fong L, Brockstedt D, Benike C, Wu L, Engleman EG. Dendritic cells injected via different routes From www.bloodjournal.org by guest on August 9, 2017. For personal use only. 2246 O’NEILL et al induce immunity in cancer patients. J Immunol. 2001;166:4254-4259. 129. Therasse P, Arbuck SG, Eisenhauer EA, et al. New guidelines to evaluate the response to treatment in solid tumors. European Organization for Research and Treatment of Cancer, National Cancer Institute of the United States, National Cancer Institute of Canada. J Natl Cancer Inst. 2000;92:205-216. 130. Slingluff C, Petroni G, Yamshchikov G, et al. Clinical and immunologic results of a randomized phase II trial of vaccination using four melanoma peptides either administered in granulocyte-macrophage colony-stimulating factor in adjuvant or pulsed on dendritic cells. J Clin Oncol. 2003;21: 4016-4026. 131. Schultz ES, Schuler-Thurner B, Stroobant V, et al. Functional analysis of tumor-specific Th cell responses detected in melanoma patients after dendritic cell-based immunotherapy. J Immunol. 2004;172:1304-1310. 132. Brossart P, Wirths S, Stuhler G, Reichardt VL, Kanz L, Brugger W. Induction of cytotoxic T-lymphocyte responses in vivo after vaccinations with peptide-pulsed dendritic cells. Blood. 2000;96: 3102-3108. 133. Maier T, Tun-Kyi A, Tassis A, et al. Vaccination of patients with cutaneous T-cell lymphoma using intranodal injection of autologous tumor-lysatepulsed dendritic cells. Blood. 2003;102:23382344. 134. Schnell S, Young JW, Houghton AN, Sadelain M. Retrovirally transduced mouse dendritic cells require CD4⫹ T cell help to elicit antitumor immunity: implications for the clinical use of dendritic cells. J Immunol. 2000;164:1243-1250. 135. Rafiq K, Bergtold A, Clynes R. Immune complexmediated antigen presentation induces tumor immunity. J Clin Invest. 2002;110:71-79. 136. Dhodapkar KM, Krasovsky J, Williamson B, Dhodapkar MV. Antitumor monoclonal antibodies enhance cross-presentation of cellular antigens and the generation of myeloma-specific killer T cells by dendritic cells. J Exp Med. 2002;195:125133. 137. Zhao Y, Boczkowski D, Nair SK, Gilboa E. Inhibition of invariant chain expression in dendritic cells presenting endogenous antigens stimulates CD4⫹ T-cell responses and tumor immunity. Blood. 2003;102:4137-4142. BLOOD, 15 OCTOBER 2004 䡠 VOLUME 104, NUMBER 8 138. Su Z, Vieweg J, Weizer AZ, et al. Enhanced induction of telomerase-specific CD4(⫹) T cells using dendritic cells transfected with RNA encoding a chimeric gene product. Cancer Res. 2002; 62:5041-5048. 139. Kumamoto T, Huang EK, Paek HJ, et al. Induction of tumor-specific protective immunity by in situ Langerhans cell vaccine. Nat Biotechnol. 2002;20:64-69. 140. Merad M, Sugie T, Engleman EG, Fong L. In vivo manipulation of dendritic cells to induce therapeutic immunity. Blood. 2002;99:1676-1682. 141. Cho HJ, Takabayashi K, Cheng PM, et al. Immunostimulatory DNA-based vaccines induce cytotoxic lymphocyte activity by a T-helper cell-independent mechanism. Nat Biotechnol. 2000;18: 509-514. 142. Belli F, Testori A, Rivoltini L, et al. Vaccination of metastatic melanoma patients with autologous tumor-derived heat shock protein gp96-peptide complexes: clinical and immunologic findings. J Clin Oncol. 2002;20:4169-4180. 143. Gold JS, Ferrone CR, Guevara-Patino JA, et al. A single heteroclitic epitope determines cancer immunity after xenogeneic DNA immunization against a tumor differentiation antigen. J Immunol. 2003;170:5188-5194. 144. Kim TW, Hung CF, Ling M, et al. Enhancing DNA vaccine potency by coadministration of DNA encoding antiapoptotic proteins. J Clin Invest. 2003; 112:109-117. 145. Leitner WW, Hwang LN, deVeer MJ, et al. Alphavirus-based DNA vaccine breaks immunological tolerance by activating innate antiviral pathways. Nat Med. 2003;9:33-39. 146. Barouch DH, Craiu A, Kuroda MJ, et al. Augmentation of immune responses to HIV-1 and simian immunodeficiency virus DNA vaccines by IL-2/Ig plasmid administration in rhesus monkeys. Proc Natl Acad Sci U S A. 2000;97:4192-4197. 147. Nair S, McLaughlin C, Weizer A, et al. Injection of immature dendritic cells into adjuvant-treated skin obviates the need for ex vivo maturation. J Immunol. 2003;171:6275-6282. 148. Tirapu I, Rodriguez-Calvillo M, Qian C, et al. Cytokine gene transfer into dendritic cells for cancer treatment. Curr Gene Ther. 2002;2:79-89. 149. Martin-Fontecha A, Sebastiani S, Hopken UE, et al. Regulation of dendritic cell migration to the draining lymph node: impact on T lymphocyte traffic and priming. J Exp Med. 2003;198:615621. 150. Wu DY, Segal NH, Sidobre S, Kronenberg M, Chapman PB. Cross-presentation of disialoganglioside GD3 to natural killer T cells. J Exp Med. 2003;198:173-181. 151. Hodi FS, Mihm MC, Soiffer RJ, et al. Biologic activity of cytotoxic T lymphocyte-associated antigen 4 antibody blockade in previously vaccinated metastatic melanoma and ovarian carcinoma patients. Proc Natl Acad Sci U S A. 2003;100:47124717. 152. Phan GQ, Yang JC, Sherry RM, et al. Cancer regression and autoimmunity induced by cytotoxic T lymphocyte-associated antigen 4 blockade in patients with metastatic melanoma. Proc Natl Acad Sci U S A. 2003;100:8372-8377. 153. Tanaka H, Tanaka J, Kjaergaard J, Shu S. Depletion of CD4⫹ CD25⫹ regulatory cells augments the generation of specific immune T cells in tumor-draining lymph nodes. J Immunother. 2002; 25:207-217. 154. Oldenhove G, de Heusch M, Urbain-Vansanten G, et al. CD4⫹ CD25⫹ regulatory T cells control T helper cell type 1 responses to foreign antigens induced by mature dendritic cells in vivo. J Exp Med. 2003;198:259-266. 155. Sutmuller RP, van Duivenvoorde LM, van Elsas A, et al. Synergism of cytotoxic T lymphocyteassociated antigen 4 blockade and depletion of CD25(⫹) regulatory T cells in antitumor therapy reveals alternative pathways for suppression of autoreactive cytotoxic T lymphocyte responses. J Exp Med. 2001;194:823-832. 156. Overwijk WW, Theoret MR, Finkelstein SE, et al. Tumor regression and autoimmunity after reversal of a functionally tolerant state of self-reactive CD8⫹ T cells. J Exp Med. 2003;198:569-580. 157. Cui Y, Kelleher E, Straley E, et al. Immunotherapy of established tumors using bone marrow transplantation with antigen gene—modified hematopoietic stem cells. Nat Med. 2003;9:952-958. 158. Nair S, Boczkowski D, Moeller B, Dewhirst M, Vieweg J, Gilboa E. Synergy between tumor immunotherapy and antiangiogenic therapy. Blood. 2003;102:964-971. 159. Asavaroengchai W, Kotera Y, Mule JJ. Tumor lysate-pulsed dendritic cells can elicit an effective antitumor immune response during early lymphoid recovery. Proc Natl Acad Sci U S A. 2002; 99:931-936. From www.bloodjournal.org by guest on August 9, 2017. For personal use only. 2004 104: 2235-2246 doi:10.1182/blood-2003-12-4392 originally published online July 1, 2004 Manipulating dendritic cell biology for the active immunotherapy of cancer David W. O'Neill, Sylvia Adams and Nina Bhardwaj Updated information and services can be found at: http://www.bloodjournal.org/content/104/8/2235.full.html Articles on similar topics can be found in the following Blood collections Immunobiology and Immunotherapy (5500 articles) Immunotherapy (577 articles) Neoplasia (4182 articles) Review Articles (712 articles) Information about reproducing this article in parts or in its entirety may be found online at: http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requests Information about ordering reprints may be found online at: http://www.bloodjournal.org/site/misc/rights.xhtml#reprints Information about subscriptions and ASH membership may be found online at: http://www.bloodjournal.org/site/subscriptions/index.xhtml Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036. Copyright 2011 by The American Society of Hematology; all rights reserved.