Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
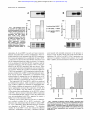
From www.bloodjournal.org by guest on August 3, 2017. For personal use only. Production of Chemokines, Interleukin-8 and Monocyte Chemoattractant Protein-l, During Monocyte:Endothelial Cell Interactions By Nicholas W. Lukacs, Robert M. Strieter, Victor Elner, Holly L. Evanoff, Marie D. Burdick, and Steven L. Kunkel The extravasationof leukocytesfrom the lumen of the vessel to.a site of inflammation requires specific binding events. The interaction of leukocytes with endothelium, via specific receptors, may provide intracellular signals that activate extravasating cells. In thepresent study, we have investigated the production of chemokines, interleukin-8 (IL-8). and monocyte chemoattractant protein-l (MCP-1) during monocyte:endothelial cell interactions. Both unstimulated and interferon-? (IFN-?)-prestimulated human umbilical vein endothelial cells (HUVEC) produced low constitutive levels of IL-8 and MCP-1. The addition of enriched monocytes with unstimulated HUVEC resulted in synergistic increases in production of both IL-8 and MCP-1. Monocytes cultured with IFN-y-preactivated HUVECs demonstrated little additional increase in IL-8 and MCP-1 production in coculture assays compared with unstimulated HUVEC. Northern blot analysis paralleled the protein data, demonstrating upregulated expression of IL-8 andMCP-1 mRNAin stimulated and unstimulated coculture assays. Culture of enrichedmonocytes and endothelial cells in transwells demonstrated no increases in IL-8 or MCP-1, indicating the necessity for cellular contact for chemokine production. In previous investigations, we have demonstrated that increased monocyte-derived MIP-la production was induced by intracellular adhesion molecule-l (ICAM-1) interactions on activated HUVECs. In contrast, addition of anti-CAM-l monoclonal antibodies (MoAbs) did notdiminish the production of IL-8 and MCP-1 in the present study. Furthermore,neither antibodies t o IL1 nor tumor necrosisfactor (TNF) diminished the production of either IL-8 or MCP-1. However, when soluble matrix proteins were added to the coculture t o block cellular interactions, the chemokine protein and mRNA levels were significantly decreased.IL-8 production was decreased by both soluble collagen and fibronectin, whereas MCP-1 was decreased by only soluble collagen, suggestingdifferential activation pathways. These results indicate that IL-8 andMCP1 production are increasedduring monocyte and endothelial cell interactions in part due t o matrix protein binding mechanisms. This mechanism may servea role in cell activation, production of chemokines, as well as extravasation and recruitment of additional leukocytes during inflammatory responses. 0 1995 by The American Societyof Hematology. T strated that monocyte interaction with intracellular adhesion molecule-l (ICAM-l), but not vascular cell adhesion molecule-l (VCAM-l), on the surface of activated endothelial cells, induced the expression of macrophage inflammatory protein-la (MIP-~cI).’~ These results support the concept that adhesion events not only provide a method of cellular interaction, but also may aid in the activation of the cells and induce the production of chemokines. In the present study, we have determined that monocyte:human umbilical vein endothelial cells (HUVEC) interactions resulted in the increased expression of MCP-1 and IL-8. This increased expression was neither dependent on the early response cytokine cascades (IL-l, tumor necrosis factor [TNF]) nor expression of adhesion molecules, ICAM1 or VCAM-1, which were upregulated during endothelial cell activation. However, chemokine expression was significantly reduced during the cell-to-cell interactions by addition of soluble matrix proteins before the coculture. These results suggest that once leukocytes adhere they may be activated HE INTERACTION of leukocytes with vascular endothelium is a primary event that occurs during cellular extravasation into inflamed tissue.’.’This interaction requires the expression of adhesion molecules on the endothelium to initiate the leukocyte adherence e ~ e n t .The ~ . ~currently held concept is that adhered leukocytes undergo migration into the tissue through a series of detachmentheadherence events typified by the polar expression of integrins specific for the adhesion molecules on the surface of mesenchymal-derived cells.’ The movement of cells into the tissue is mediated via chemotactic gradients with the highest concentration at the site of inflammation. Specific chemotactic factors have been isolated and classified into a supergene family of chemotactic cytokines (chemokines). The chemokines have been divided into two subgroups based on the juxtaposition of the first two cysteine residues in their amino acid sequences. The CX-C family, typified by interleukin-8 (IL-8), is primarily chemotactic for neutrophils, while the C-C chemokine family, typified by monocyte chemoattractant protein-l (MCPl), is primarily chemotactic for mononuclear leukocytes.’ The movement of specific populations of leukocytes into tissue is thought to be mediated by concentration gradients of chemokines that appear to be chemotactic for specific groups of leukocytes. Therefore, the extravasation of leukocytes into inflamed tissue relies on the coordination of endothelial expressed adhesion molecules, leukocyte adherence, and specific chemotactic gradients. The expression of chemokines can be induced by a variety of cytokines in a variety of immune and nonimmune cells. These cell types include fibroblasts, epithelial, endothelial, and smooth muscle cells, as well as mononuclear leukocytes and gran~locytes.~ In addition, recent studies have demonstrated the expression of cytokine mRNAon adhesion of monocytes to plastic or tissue matrix, thus indicating that adherence events alone were sufficient for the activation of cytokine Recently our laboratory has demonBlood, Vol 86,No 7 (October l ) , 1995:pp 2767-2773 From the Departments of Pathology and Internal Medicine, the Division of Pulmonary and Critical Care, Medicine, University of Michigan Medical School, Ann Arbor. Submitted March 6, 1995; accepted May 31, 1995. Supported in part by National Institutes of Health Grants No. HL02401, HL31963, and HL35276, and the American Heart Association of Michigan. Address reprint requests to Nicholas W . Lukacs, PhD, University of Michigan Medical School, Department of Pathology, 1301 Catherine, Ann Arbor, MI 48109-0602. The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 0 I995 by The American Society of Hematology. 0006-4971/95/8607-~20$3.00/0 2767 From www.bloodjournal.org by guest on August 3, 2017. For personal use only. 2768 LUKACSET AL by mechanisms in conjunction with extravasation, resulting in increased production of IL-8 and MCP- 1. MATERIALS AND METHODS Mononuclear cell isolation. Peripheral bloodwas drawn into a heparinized syringe from healthy volunteers, diluted 1:l in normal saline and mononuclear cells separated by density gradient centrifugation. The recovered cells were washed three times withRPM1 1640. The PBMs were then layered onto a density gradient (1.068 g/mL) for the enrichment of monocytes (Fico-lite, Atlanta Biologics, Atlanta, GA). The isolated cells were then washed, cytospun onto and difa glass slide, stained with Diff-Quik (Baxter, McGaw, L), ferentially counted. The purity of the monocytes from the gradient was consistently between 75% and 80% monocytes with the remainder lymphocytes. HUVEC culture. HUVEC were isolated and established in culture as previously described? Briefly, primary cultures were passaged and maintained in endothelial cell growth medium (modified MCDB 131; Clonetics, San Diego, CA). For experiments, the cells were subcultured (passage 3) into 6-well plates (Costar, Kennebunkport, ME), grown to confluency, and used in monocyte adherence assays. HUVECtmonocyte cocultures. Enriched monocyte populations (5 X lo5cells/mL) were layered onto unstimulated or 24 hour stimulated (IFN-y, 1.000 UlmL) HUVEC monolayers in 6-well plates. Before the addition of the monocytes, HUVECs were gently washed free of IFN-7. The monocyte enriched cells were then added to the activated HUVECs in a total of 1 mL of media in the 60-mm culture dishes. Culture supernatants were collected at specific times (4, 12, 24, and 48 hours) after coculture. Peak MCP-1 and IL-8 production was detected at 24 hours of coculture. To detect the source of the chemokines, monocytes were separated from the endothelial cells after 4 hours of incubation using cold (4°C) Ca", Mgz'-free saline containing EDTA (3-minute incubation). In addition, monocytes were added to transwells (Costar) in the HUVEC cultures to detect whether cell contact was obligatory for chemokine production. Monoclonal antibodies to either ICAM-l, or VCA"1 (R 8r D Systems, Minneapolis, MN) were added at a concentration of 5 pg/ mL to the HUVECmonocyte interactions. This concentration has been shown in previous studies to be optimal for inhibiting monocyte:endothelial cell interaction^.'^ Northern blot analysis. Total RNA from either monolayer of cells or from homogenized tissue was isolated using a modification of the method of Chirgwin" andJonas.16Briefly, samples were dispersed in a solution of 25 mmoVL Tris Ph 8.0 containing 4.1 m o m guanidine isothiocyanate, 0.5% sarkosyl, and 0.1 m o m beta mercaptoethanol. The RNA was further extracted with chloroformphenol and then alcohol precipitated. The separated RNA was transblotted to nitrocellulose, baked, prehybridized, and hybridized with '*P-5' end-labeled oligonucleotide antisense probes for IL-8,and MCP-l. In addition, a labeled oligonucleotide for actin served as a control. Autoradiographs were quantitated using a computer image analysis system. Equivalent amounts of RNA/gel were monitored by assessing 28s and 18s rRNA or beta actin mRNA. Blockage of cellular interactions. Previous results have indicated a role of matrix protein binding for activation of cytokine expression in monocytes. To interfere with the interaction of monocytes with endothelial cells, the addition of soluble matrix proteins, fibronectin (Chemicon, Temecula, CA) from human serum or basement membrane component collagen type 1(30 pg/mL), to the coculture assays was used toelucidate the role of these interactions during cell-to-cell contact. Monocytes were preincubated with the matrix proteins for 30 minutes at 37°C before the addition to the HUVEC monolayers. Culture supematants were harvested at various time Table 1. Production of IL-8 and MCP-1 During Monocyte:EndothelialCell Interaction IL-8 Production (ng/mL) Group Nonstimulated HUVEC Nonstimulated monocytes Nonstimulated HUVEC + monocytes IFN-stimulated HUVEC IFN-stimulated monocytes IFN-stimulated HUVEC + monocytes 2.5 1.5 28.7 4.3 1.0 37.5 i 0.72 -t 0.20 i 4.4 1.10 2 0.02 i 4.1 MCP-1 Production (nglmL) 1.4 2 0.16 1.01 5.9 2 0.66 2.7 2 0.62 1.01 6.2 i 0.38 points and assayed for the production of IL-8and MCP- 1 and compared with an intraassay control culture. IL-8 and MCP-I enzyme-linked immunosorbent assay. Extracellular immunoreactive L-8 and MCP-l was quantitated using a modification of a double-ligand method as previously described." Briefly, flat-bottomed 96-well microtiter plates were coated with 50 pL/well of rabbit anticytokine antibodies (1 pg/mL in 0.6 mom NaCl, 0.26 moVL H'BO,, and 0.08 N NaOH, pH 9.6) for 16 hours at 4°C and then washed with phosphate-buffered saline (PBS) pH 7.5, 0.05% Tween 20 (wash buffer). Nonspecific binding sites on microtiter plates were blocked with 2% bovine serum albumin (BSA) in PBS. Plates were rinsed with wash buffer, and samples added, followed by incubation for 1 hour at 37°C. Plates were washed, then 50 pL/ well of biotinylated rabbit anti-IL-l0 was added and incubated for 30 minutes at 37°C. After washing of plates, chromogen substrate was added. The plates were incubated at room temperature, and the reaction terminated with 50 pL/well of 3 m o m H3S04solution, and read at 490 nm in an enzyme-linked immunosorbent assay (ELISA) reader. Standards were '/* log dilutions of recombinant IL-8 and MCP-l from 1 pg/mL to 100 ng/mL. This ELISA method consistently detected concentrations above 10 pglmL and did not crossreact with MIP-lP, RANTES, IL-la/P, TNFa, ENA-78 (epithelialderived neutrophil activating protein-78), MIP-la, growth-related oncogene a (GROa), GROP, GRO-y, or neutrophil activating protein-2 (NAP-2). Statistical analysis. Data are expressed as means ? standard error ofmean (SEM). Data that appeared statistically significant were compared by analysis of variance (ANOVA) for comparing the means of multiple groups and considered significant if P values were less than .05. RESULTS Production of IL-8 and MCP-l during monocyte:HUVEC interactions. To determine whether HUVEC:monocyte interactions induced increased production of the chemokines, supernatants from a time course of cocultures were assayed. Both HUVEC and monocytes individually produced E-8. However, when monocytes were added onto monolayers of HUVEC cultures, a synergistic increase in IL-8 production was observed (P < .OOl). Additional experiments demonstrated that 12 to 18 hours' preactivation of HUVEC monolayers with IFN-y did not significantly increase the level of IL-8 in either the HUVEC alone or during coculture conditions (Table 1). MCP-1 was produced by unstimulated cultures of HUVEC, but not by monocytes. On HUVEC:monocyte coculture, there was an increase in MCP-1 production ( P < .001). The preactivation of HUVEC monolayers significantlyincreased MCP-l production in HUVEC alone, but not in monocyte cocultures (Table 1). The addition of enrichedmonocytesontoHUVECmonolayersreleased From www.bloodjournal.org by guest on August 3, 2017. For personal use only. 2769 PRODUCTION OF CHEMOKINES 1-mstimalated HWEC Fig l.TheexpressionofIL-8 (A) and MCP-1 (B) mRNA during monocyte:endothelial cell interactions. Enriched monocytes (5 x 105/mLIwere plated onto unstimulated and IFN-y-stimulated (1.000 UlmL) HUVEC monolayer cultures. Total cellular RNA was isolated from 8hour cultures (time of peak mRNA expression). Northern blot analysis was performed using 11-8and MCP-1specific P’*labeled oligo-probes. Repeat exp e r i m e n t ~showed ~ similar results. 2-IFW-HWEC + 3-unsti mutated H W E C + t l o no:, .c i W 0 1 2 1 3 2 3 18s chemokines, IL-8, and MCP- 1 as early as 4 hours’ postcombination and peaked after 24 hours. Furthermore, when enriched monocytes were incubated with HUVEC monolayers for 4 hours and separated for an additional 24 hours, MCP1 production was derived from the HUVEC monolayers and not the separated monocytes (data not shown). In contrast, IL-8 was produced by both the HUVEC monolayer and the separated monocytes, with the majority (70% to 75%) produced by the HUVEC monolayer (data not shown). These results indicate that both cell populations were activated during coculture. Neither the coculture of enriched lymphocyte ( S O % ) nor neutrophil (>95%) populations induced the production of either IL-8 or MCP-I, indicating leukocyte specificity for the response. Furthermore, in experiments using transwell plates to separate the two cell populations, no increased expression of chemokine was observed (data not shown), indicating a requirement for cell-to-cell contact. The expression of IL-8 and MCP-I mRNA was analyzed by Northern blot analysis. Little constitutive expression of IL-8 or MCP-I mRNA was detected in unstimulated HUVEC cultures. Addition of monocytes to the HUVEC monolayers significantly increased the expression of bothIL-8 (Fig 1A) and MCP-l (Fig IB) mRNA. As with the results of the protein data, prestimulation of the HUVEC cultures with IFN-.)I did not increase chemokine mRNA expression over unstimulated HUVEC cultures. In fact, the cultures that were not prestimulated with IFN-y appeared to express a higher level of the chemokine mRNA. Inability of Ab to either adhesion molecules or inJlammatory cytokines to inhibit IL-8 or MCP-I production. Our laboratory has demonstrated that the ability to induce MIPla, a C-C chemokine, during HUVEC:monocyte coculture was dependent on ICAM-l interactions.’3 To determine whether IL-8 and MCP-I were also induced by adhesion molecules, unstimulated and IFN-y-stimulated HUVEC were layered with enriched monocytes in the presence or absence of anti-ICAM-I MoAb. The results showed that no inhibition of IL-8 (Fig 2) or MCP-I (Fig 3) production was observed. In addition, Northern blot analysis of IL-8 and MCP- 1 mRNA also demonstrated no decrease in the mRNA T 50 A A I I . E .- 0 -30 V U 10 0 P> 3 X Unstimulated C m 0 Y E > 3 X IFN-g Pre-stimulated Fig 2. Blockage of adhesion molecule, ICAM-1, interaction does not inhibitIL-8 production during monocyte:endothelial cell interactions. Enriched monocytes (5 x 105/mL)were plated onto unstimulated and IFN-y-stimulated (1.000 UlmL) HUVEC monolayer cultures. MoAbs (5 pg/mL) directed against CAM-l were added to the cocultures and24-hoursupernatantswereharvestedandassayedby ELlSA for IL-8. From www.bloodjournal.org by guest on August 3, 2017. For personal use only. 2770 LUKACS ET AL ‘I .I t m T T T E v ti 0 I 1 + 8 > C m 3 I unStirnulated 1 production was inhibited only by the addition of soluble collagen (P < .05), butnot significantly withfibronectin (Fig 5 ) , possibly suggesting differences inthe activation pathway for these two chemokines. The examination of the mRNA expression during the coculture also demonstrated similar findings.When the monocytes were preincubated withmatrix proteins, the expression of IL-8 andMCP-I mRNAwas decreased (Fig 6). Similar to the results for protein levels of IL-8 and MCP-1, IL-8 mRNA levels were diminished when monocytes were incubated with either soluble collagen or fibronectin, whereas MCP-1 demonstrated that only collagen was effective in decreasing the mRNA expression. There appeared to be some specificity for the matrix protein used, as laminin did not inhibit either IL-8 or MCP-1 (data not shown). Altogether, the latter data suggest that IL-8 and MCP-I production during cell-to-cell contact may be induced, in part, via matrix protein binding events. IFNg Pre-Stimulated Fig 3. Blockage of adhesionmolecule, CAM-1, interaction does not inhibit MCP-1 productionduring monocyte:endothelial cell interactions. Enrichedmonocytes(5 x 106/mL)were plated onto unstimulated and IFN-y-stimulated (1.000 UlmL) HUVEC monolayer cultures. MoAbs (5 pg/mLl directed against ICAM-1 were added to thecocultures and 24-hour supernatants were harvested andassayed by ELISA for MCP-1. expression by treatment of cocultures with anti-ICAM-11 VCAM-I molecule MoAbs (data not shown). In additional experiments, we were also interested in whether IL-1 and TNF played a role in the production of IL-8 and MCP- 1. These two potent inflammatory cytokines have previously been shown to induce chemokines in most cell types. In coculture supernatants from different time points ( 1 , 4, 8, 12, and 24 hours) little IL-I (< 100 pg/ mL) and no TNF was detected. Subsequently, addition of neutralizing antibodies to the coculture assays directed against IL- 1 and TNF had no effect on either IL-8 or MCPl production (data not shown). Altogether, these studies show that neither adhesion molecule nor inflammatory cytokine cascades were responsible for the upregulation of IL8 and MCP-1. Inhibition of chemokine production by blocking matrix protein interactions. To determine whether additional cellto-cell interactions were influencing the production of IL-8 and MCP-I, we examined the role of matrix protein interactions. Previous studies have identified matrixprotein interactions as inducing IL-8 production.” To identify therole matrix proteins played in the HUVEC:monocyte interactions, the mononuclear cells were preincubated with either soluble fibronectin or type I collagen (30 pg/mL) for 15 minutes and then layered onto unstimulated HUVEC monolayers. These studies demonstrated a significant inhibition in chemokine production during the cell-to-cell interactions, (Figs 4 and 5). IL-8 production during cell-to-cell interactions was inhibited by the addition of either soluble fibronectin or collagen to the culture media (Fig 4) ( P <c .05). MCP- DISCUSSION The extravasation of cells from the lumen of a vessel into inflamed tissue depends on leukocyte interaction with activated endothelium.’,’ The initial adhesion relies on interactions between P-integrins on the surface of leukocytes and 30- 20- 10 - P 0 S + U W 2 I o + W > + + 3 I I: Fig 4. Inhibition of IL-8 productionby solublematrix proteinsduring monocyte:endothelialcell interactions. H U M C and monocyte populations were preincubated with soluble collagen or fibronectin for 30 minutes before coculture. Enriched monocytes 15 x 1O5/mL1 were plated onto unstimulated HUVEC monolayer cultures. Cell-free culture supernatants were harvested after 24 hours and analyzedfor IL-8 production. Data represent the mean of three repeat experiments. From www.bloodjournal.org by guest on August 3, 2017. For personal use only. PRODUCTION OF CHEMOKINES 277 1 T Fig 5. Inhibition of MCP-1 production by soluble matrix proteins during monocyte:endothelial cell interactions. HUVEC and monocyte populations were preincubated with soluble collagen or fibronectin for 30 minutes before coculture. Enriched monocytes l5 x 105/mL) were plated onto unstimulated HUVEC monolayer cultures. Cell-free culture supernatants were harvested after 24 hours and analyzedfor MCP-1 production. Data represent the mean of three repeat experiments. adhesion molecules on the activated endothelium. Additional interactions between the endothelium and adhered leukocytes, which may include binding via matrix proteins as well as other specific receptor-ligand interactions, also appear to be ~perative.'"'~ In the present study, our results demonstrate activation of IL-8 and MCP-I expression during monocyte:HUVEC interactions. The upregulation of the two chemokines was dependent on cell-to-cell contact andnot soluble mediators, as the addition of enriched monocytes to HUVEC cultures in transwell separated culture dishes did not induce the expression of the chemokines. Because monocyte adherence and stretching over the endothelium is a reversible and dynamic event,*').*'the interaction of monocytes with the HUVEC must be transient. Interestingly, the increased production of the two chemokines was notdependent on activation of the endothelium, as IFN-y pretreatment of HUVEC monolayers had little effect on IL-8 or MCP-I production. In addition, the interaction between cells leading to the production of IL-8 and MCP-l wasnot dependent on ICAM- 1 or VCAM- 1 interactions, as MoAbs to these molecules also had no effect on the chemokine production. These findings are in contrast to our previous study that examined MIP-la production during monocyte:HUVEC interaction^.'^ In the previous study, we showed that MIP-la production was dependent on ICAM-I expression and interactions of monocytes with activated endothelial cells. The present study also investigated production of potentinflammatory cytokines, IL-I, and TNF, which could potentially induce the increased IL-8 and MCP-I production through cytokine cascades. These results demonstrated little detectable IL-I and TNF production at any time throughout the coculture assay. In addition, antibodies directed to IL-I and TNF did not inhibit the production of either IL-8 or MCP-I. These data suggest that the increased production of the two chemokines is independent of traditional inflammatory cytokine ACTIN IL-8 MCP-1 A Fig 6. Inhibition of K-8 and MCP-1 mRNA expression during monocyte:endothelial cell interactions by soluble matrix proteins. RNA was analyzed from HUVEC alone (A) or HUVEC and monocyte populations preincubated with BSA (B), solublefibronectin IC), or soluble collagentype I ID) for 30 minutes be(5 x 105/mL) fore coculture.Enrichedmonocytes were plated onto unstimulated HUMC monolayer cultures. Total cellular RNA was isolatedfrom 8-hour cultures (time of peak mRNA expression). Northern blot analysis was performed using IL-8 and MCP-1 specific P3*-labeled oligo-probes. Repeat experiments showed similar results. B C c A B C D D 5- A B C D From www.bloodjournal.org by guest on August 3, 2017. For personal use only. 2772 cascades. However, the production of IL-8 and MCP-1 was dependent on the interaction of monocytes with matrix proteins, possibly via p-integrins." This latter mechanism was verified using soluble matrix proteins to inhibit chemokine production. The fact that both soluble fibronectin and collagen decreased IL-8, while only soluble collagen diminished MCP-1 production, suggests differential regulation during the cell-to-cell interactions. Interestingly, IL-8 was produced by both cell types, whereas MCP-1 was produced by only the HUVEC monolayer. This discrepancy in chemokine production may explain the differential regulation by the soluble matrix proteins. Because the preincubation of cells before coculture only partially decreased the production of chemokines, it is likely that other mechanisms are operative during the binding events that facilitate the upregulation of the chemokine production other than matrix binding. Altogether, these results may be indicative of regulational events that occur during cellular interactions, adhesion events, and chemokine production. The induction of cytokines during monocyte adhesion has previously been shown in other studies."." In addition, the upregulation of L - 8 protein in monocytes during fibronectin adherence events has demonstrated a relationship of cellular adhesion to chemokine production." The mechanism of these interactions during leukocyte extravasation under flow conditions would be dependent on the initial binding of monocytes to endothelial expressed selectin and adhesion molecules, thus allowing secondary adhesive events to conAs previously demonstrated, the specific binding to ICA"1 was required to upregulate the production of MIPla from monocyte^.'^ The binding of monocytes under nonflow conditions to other ligands on endothelium,'"20as demonstrated in the present study, can upregulate the expression of IL-8 and MCP-1. However, in vivo, the secondary interactions under flow conditions would always be dependent on the initial requirement of selectin and ICAM-1 expression on the endothelial cell layer','and subsequent monocyte binding to additional ligands, such as matrix proteins. Therefore, the limiting step for upregulation of IL-8 and MCP-1 under the shear forces of flow conditions would always be inflammation-induced expression of adhesion molecules on the endothelium. This would give the system the fidelity needed to control the induction and release of the chemokines during homeostatic and inflammatory events. These interactions may be a contributing step in exacerbation of vascular diseases, such as atherosclerosis, in which there is prolonged and intimate interaction of monocytes and endothelial cells correlated with chemokine expression within the Iesion.22-25 The significance of the increased production of chemokines during extravasational events may be multifunctional. The activation of leukocytes as they extravasate into inflamed tissue may allow deposition of chemokines into the surrounding tissue, intensifying the chemotactic gradient, and increasing the accumulation and activational status of the infiltrate. Additionally, these chemokines may also function to activate not only the infiltrating leukocytes, but also the surrounding structural cells, therefore, maintaining the inflammatory milieu. Finally, the increased production of IL-8 and MCP-l may aid in roles other than recruitment of LUKACS ET AL leukocytes. For example, IL-8 has recently been described as a potent angiogenic whereas MCP-1 (JE) wasfirst described as a competence factor for fibroblast activation and proliferation." Both of these understudied areas of chemokine functions could also contribute to the maintenance of an inflammatory response. The production of cytokines during cell-to-cell interactions may be an important mechanism for maintenance and regulation of an inflammatory response. Increased IL-8 and MCP-1 production during monocyte:endothelial cell interaction was not dependent on inflammatory cytokine (IL-1 or TNF) cascades or expression of adhesion molecules (ICAM1 or VCAM-l), but, in part, on interactions with matrix proteins. These studies suggest that interactions of leukocytes with endothelial cells can induce XL-8 and MCP-I production, possibly influencing the intensity of the inflammatory infiltrate and maintenance of the response in vivo. REFERENCES 1. Springer TA: Adhesion receptors of the immune system. Nature 346:425, 1990 2. Zimmerman CA, Prescott SM, McIntyre TM: Endothelial cell interactions with granulocytes: Tethering and signaling molecules. Immunol Today 13:93, 1992 3. Luscinskas F W , Cybuisky MI, Kiely JM, Peckins CS, Davis VM, Gimbrone MA Jr: Cytokine-activated human endothelial monolayers support enhanced neutrophil transmigration via a mechanism involving both endothelial-leukocyte adhesion molecule- 1 and intracellular adhesion molecule 1. J Immunol 146:1617, 1991 4. Bevilacqua MP, Pobex JS, Wheeler ME, Cotran RS, Gimbrone MA Jr: Interleukin 1 acts on cultured human vascular endothelium to increase the adhesion of polymorphonuclear leukocytes, monocytes, and related leukocyte cell lines. J Clin Invest 76:2003, 1985 5 . Dustin ML, Rothlein R, BhanAK, Dinarello CA, Springer TA: Induction by IL-I and interferon-gamma: tissue distribution, biochemistry, and function of a natural adherence molecule (ICAMl). J Immunol 137:245, 1986 6. Hakkert BC, Kuijpers T W , Leeuwenberg JF, van Mourik JA, Roos D: Neutrophil and monocyte adherence to and migration across monolayers of cytokine-activated endothelial cell: The contribution of CD18, ELAM-l, and VLA-4. Blood 78:2721, 1991 7. Carlos T, Kovach N, Schwartz B, Rosa M, Newman B, Wayner E, Benjamin C, Osborn L, Lobb R, Harlan J: Human monocytes bind to two cytokine-induced adhesive ligands on cultured human endothelial cells: Endothelial-leukocyte adhesion molecule-l and vascular cell adhesion molecule-l. Blood 77:2266, 1991 8. Oppenheim JJ, Zachariae COC, Mukaida N, Matsushima K: Properties of the novel proinflammatory "intercrine" cytokine family. Annu Rev Immunol 9:617, 1991 9. Kunkel SL, Lukacs N W , Chensue SW, Strieter RM: Adhesion molecules and chemokines dictate leukocyte recruitment, in Faist E (ed): Third International Congress on the Immune Consequences of Trauma, Shock, and Sepsis. Munich, Germany, Biomedical Press, 1994 10. Kasahara K, Strieter RM, Chensue SW, Standiford TJ, Kunkel SL: Mononuclear cell adherence induces neutrophil chemotactic factor/interleukin-8 gene expression. J Leukoc Biol 50:287, 1991 11. Haskill S, Johnson C, Eierman D, Becker S, Warren K: Adherence induces selective mRNA expression of monocyte mediators and proto-oncogenes. J Immunol 1401690, 1988 12. Eierman DF, Johnson CE, Haskill JS: Human monocyte inflammatory mediator gene expression is selectively regulated by adherence substrates. J Immunol 142:1970, 1989 13. Kasahara K, Strieter RM, Standiford TJ, Kunkel SL: Adher- From www.bloodjournal.org by guest on August 3, 2017. For personal use only. PRODUCTION OF CHEMOKINES ence in combination with lipopolysaccharide, tumor necrosis factor or interleukin-lb potentiates the induction of monocyte-derived interleukin-8. Pathobiology 61:57, 1993 14. Lukacs N W , Strieter RM, Elner VM, Evanoff HL, Burdick MD, Kunkel SL: Intracellular adhesion molecule-l mediates the expression of monocyte-derived MIP-la during monocyte-endothelial cell interactions. Blood 83:1174, 1994 15. Chirgwin JM, Przbyca AE, MacDonald RI, Rutter WJ: Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5294, 1979 16. Jonas E, Sargent TD, David IB: Epidermal keratin gene expressed in embryos of Xenopus laevis. Proc Natl Acad Sci USA 825135, 1985 17. Evanoff H, Burdick MD, Moore SA, Kunkel SL, Strieter RM: A sensitive ELISA for the detection of human monocyte chemoattractant protein-l (MCP-I). Immunol Invest 21:39, 1992 18. de Sousa M, Tilney N L , Kupiec-Weglinski J W : Recognition of self within self: Specific lymphocyte positioning and the extracellular matrix. Immunol Today 12:248, 1991 19. Levesque JP, Hatzfeld A, Hatzfeld J: ECM-immune-system interactions: Mitogenic properties of major extracellular proteins. Immunol Today 12:258, 1991 20. Albelda SM, Buck CA: Integrins and other cell adhesion molecules. FASEB J 4:2868, 1990 21. Gruber MF, Webb DSA, Gerrard TL: Stimulation of human 2773 monocytes via CD45, CD44,and LFA-3 triggers macrophage-colony-stimulating factor production: Synergism with lipopolysaccharide and L-lb. J Immunol 148:1113, 1992 22. Takeya M, Yoshimura T, Leonard EJ, Takahashi K: Detection of monocyte chemoattractant protein-l in human atherosclerotic lesions by an anti-monocyte chemoattractant protein-l monoclonal antibody. Hum Path01 24:534, 1993 23. Valente AJ, Rozek MM, Sprague EA, Schwartz CJ: Mechanisms in intimal monocyte-macrophage recruitment. A special role for monocyte chemotactic protein- 1. Circulation 86: 11 120, 1992 24. Neiken NA, Coughlin SR, Gordon D, Wilcox JN: Monocyte chemoattractant protein-l in human atheromatous plaques. J Clin Invest 88:1121, 1991 25. Strieter RM, Koch AE, Antony VB, Fick RB Jr, Standiford TJ, Kunkel SL: The immunopathology of chemotactic cytokines: The role of interleukin-8 and monocyte chemoattractant protein-l. J Lab Clin Med 123:183, 1994 26. Koch M , Ploverini PJ, Kunkel SL, Harlow LA, DiPietro LA, Elner VM, Elner SG, Strieter RM: Interleukin-8 as a macrophagederived mediator of angiogenesis. Science 258:1798, 1992 27. Rollins BJ, Momso ED, Stiles CD: Cloning and expression of JE, a gene inducible by platelet-derived growth factor and whose product has cytokine-like properties. Proc Natl Acad Sci USA 85:3738, 1988 From www.bloodjournal.org by guest on August 3, 2017. For personal use only. 1995 86: 2767-2773 Production of chemokines, interleukin-8 and monocyte chemoattractant protein-1, during monocyte: endothelial cell interactions NW Lukacs, RM Strieter, V Elner, HL Evanoff, MD Burdick and SL Kunkel Updated information and services can be found at: http://www.bloodjournal.org/content/86/7/2767.full.html Articles on similar topics can be found in the following Blood collections Information about reproducing this article in parts or in its entirety may be found online at: http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requests Information about ordering reprints may be found online at: http://www.bloodjournal.org/site/misc/rights.xhtml#reprints Information about subscriptions and ASH membership may be found online at: http://www.bloodjournal.org/site/subscriptions/index.xhtml Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036. Copyright 2011 by The American Society of Hematology; all rights reserved.