Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Permeability of blood-ocular barriers of
neonatal and adult cats to
fluorescein-labeled dextrans of selected
molecular sizes
Roy W. Bellhorn
The permeability of the ocular blood vessels and neuroepithelial layers (the blood-ocular barriers) to fluorescein-labeled dextrans of selected molecular sizes was evaluated in neonatal and
adult cats by fluorescence microscopy. The iris and ciliary process vessels were permeable to
molecules as large as 85A effective diffusion radius (EDR). In the kitten, the choriocapillaris
was permeable to molecules as large as 58A EDR, and in the adult to molecules only as large as
32A EDR. The retinal vessels and the retinal pigment epithelium were impermeable to all
markers. The role of the cat's iris vessel in aqueous humor formation appears dissimilar to that
of other species in that molecules larger than serum proteins traverse the walls of the iris
capillaries.
Key words: neonatal kitten, cat, permeability, blood-ocular barriers, fluorescein
isothiocyanate labeled dextrans, ocular development, vessels, aqueous humor,
fluorescence microscopy, tracer molecules
c
certain morphologic and physiologic characteristics of the ocular blood vessels and
neuroepithelial layers constitute what are
known as the blood-ocular barriers. 1 The
basic function of those barriers is to control
the movement of molecules from the vascular
lumen to the paravascular tissues and/or to
the ocular fluids. Size is one determinant
whereby a molecule may or may not traverse
a vascular endothelial lining, and dextran
molecules of selected sizes have proved use-
From the Department of Ophthalmology, Montefiore
Hospital and Medical Center, the Albert Einstein College of Medicine, Bronx, N.Y.
Supported by U.S. Public Health Service grant 1 R01
EY 02038 and by Research to Prevent Blindness, Inc.
Submitted for publication Jan. 29, 1980.
Reprint requests: Dr. Roy W. Bellhorn, Department of
Ophthalmology, Montefiore Hospital and Medical
Center, 111 East 210th St., Bronx, N.Y. 10467.
282
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
ful in the study of that permeability determinant. 2 ' 3 Fluorescein isothiocyante (FITC)labeled dextrans of selected molecular sizes
provide the added potential of in vivo as well
as in vitro localization of the molecules by
fluorescent techniques. We have previously
demonstrated the usefulness of FITCdextrans in studies of normal and abnormal
rat retinal vessels,4' 5 the avian pecten, 4 and
the rat anterior segment. 6 Fluorescent microscopic localization of FITC-dextrans has
been utilized in studies of aqueous humor
outflow pathways7' 8 and in blood-ocular barrier permeability determinations. 9
Fluorescent microscopic evaluations of
freeze-dried tissues wherein sodium fluorescein (NaFl) was the permeability marker
have been performed for both aqueous humor pathway and blood-ocular barrier investigations.10"14 In a study of the blood-ocular
barriers of neonatal and adult cat, the iris
0146-0404/81/080282+09$00.90/0 © 1981 Assoc. for Res. in Vis. and Ophthal., Inc.
Volume 21
Number 2
Feline blood-ocular barrier permeability
283
Table I. Penetrance of selected molecular sized FITC-dextrans into ocular
tissues of kittens
Molecule (EDR)
FITC/dex 3 (12A)
FITC-dex 20 (32A)
FITC-dex 40 (45A)
FITC-dex 70 (58A)
FITC-dex 150 (85A)
Circulation
time (min)
Iris
stroma
Ciliary process Ciliary process
epithelium
stroma
Choroidal
stroma
Retina via
Retina via
RPE
retinal vessels
1
4
2
8
3
12
5
20
5
30
Relative degree of fluorescence within tissue as indication of permeability of associated barrier vessels or epithelium: + + + = marked;
+ + = moderate; + = minimal; — = negative.
capillaries were found to be markedly permeable to NaFl,14 a finding decidedly dissimilar
to that reported for other species.11' 12j 15 It
again brought to mind the question concerning the role of those vessels in aqueous formation.16 Because NaFl is partially bound to
serum proteins, it is not known in such
studies whether it is the free NaFl (376 mol.
wt; 5A effective diffusion radius (EDR)) or
the protein-bound NaFl (35A EDR) that does
or does not pass the barriers.
To begin assessing that question, FITCdextrans of selected molecular sizes were
utilized to evaluate the permeability of the
blood-ocular barriers in the neonatal and
adult cat. The results form the basis of this
report.
Materials and methods
A total of 16 kittens, ages 5, 13, and 21 days,
and 13 adult cats were anesthetized with sodium
pentobarbital (35 mg/kg I.P.) and injected intravenously with 0.1 cc of 33% FITC-dextran/
0.1M sodium phosphate buffer solution per 200
gin body weight. The compounds utilized were
FITC-dex 3 (mol.wt. 3000; EDR 12A), FITC-dex
20 (mol.wt. 20,000; EDR 32A), FITC-dex 40
(mol.wt. 40,000; EDR45A), FITC-dex 70 (mol.wt.
70,000; EDR 58A), and FITC-dex 150 (mol.wt.
150,000; EDR 85A). Details of the properties of
these compounds are given in a previous publication.4 After circulation times from 30 sec to 30 min,
depending on the molecular size of the injected
FITC-dextran (Table I), the eyes were removed
and quick-frozen in isopentane cooled to —105° C
by a liquid nitrogen bath. The eyes were freeze-
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
dried for 12 to 18 days in a molecular sieve apparatus in a —35° C environment. After drying, the
eyes were grossly dissected, and portions were
embedded under vacuum in wax. Sections were
cut at 10 /Ltm and examined by epi-illumination
fluorescence microscopy with a Zeiss Photomicroscope II equipped with a BG-12 excitor and a Zeiss
50 barrier filter. Photographs were taken with 400
ASA Ektachrome color film.
Results
Neonatal cat. The penetrance of the different FITC-dextran markers into the ocular tissues of the kitten is shown in Table I. Those
molecules with an EDR of 58A or less readily
passed the iris capillaries (Figs. IB, 2A, and
2B) whereas the 85A molecule did so only
minimally. There was evidence of fluorescence in greater concentration in the posterior stroma adjacent to the iris pigment epithelium (Figs. IB and 2A), and this was interpreted as evidence that the FITC-dextrans
entered the iris stroma from the iris capillaries in that region rather than by movement
from the anterior chamber aqueous. This
gradient was more pronounced in the shorter
circulation time specimens (Figs. 2A and 2B).
All FITC-dextran molecules up to and including the 85A molecule were markedly
evident in the ciliary process stroma. However, fluorescence was evident in the intercellular spaces of the nonpigmented epithelium of the ciliary processes only with the
FITC-dex 3 (Fig. 3) and possibly with the
FITC-dex 20 markers. Topographic evalua-
284
Invest. Opkthalmol. Vis. Sci.
August 1981
Bellhorn
AC
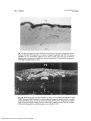
Fig. 1A. Photomicrograph of 5-day-old kitten freeze-dried iris tissue for orientation purposes.
Posterior chamber (PC) adjacent to two layers of pigment epithelial cells (PE). Iris stromal
capillaries (C) are immediately adjacent to PE layers. Large vessel area (L) lies between
capillary area and anterior iris border and anterior chamber (AC). (Freeze-dried preparation;
white-light transillumination; X86.)
Fig. IB. Photomicrograph of 13-day-old kitten iris after 1 min circulation of FITC-dex 3 (12A
EDR). Presence of dye in posterior iris stroma adjacent to pigment epithelium (PE) is marked.
Diminished fluorescence in anterior .stroma and marked fluorescence along posterior walls of
large vessels (arrowheads) suggest movement of dye from capillaries toward anterior chamber.
(Freeze-dried preparation; epi-il!umination; wax-embedded; X25.)
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
Volume 21
Number 2
Feline blood-ocular barrier permeability
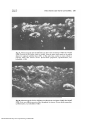
Fig. 2A. Photomicrograph of 5-day-old kitten iris after 5 min circulation of FITC-dex 70 (58A
EDR). Presence of dye within vessels is marked. Note the many small vessels in iris stroma
adjacent to pigment epithelial layers (PE). At this early time, marker is more evident in
posterior rather than anterior stroma. (Freeze-dried preparation; epi-illumination; waxembedded; X125.)
Fig. 2B. Photomicrograph of 5-day-old kitten iris after 20 min circulation of FITC-dex 70 (58A
EDR). Note more diffuse presence of dye throughout iris stroma. (Freeze-dried preparation;
epi-illumination; wax-embedded; X320.)
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
285
286
Bellhorn
Invest. Ophthalmol. Vis, Set.
August 1981
Fig. 3. Photomicrograph of ciliaiy process of 13-day-old kitten after 1 min circulation of
FITC-dex 3 (12A EDR). Note marked presence of dye in stronia and presence of dye between
nonpigmented epithelial cells (arrows) in this tangential section. PC, Posterior chamber.
(Freeze-dried preparation; epi-illumination; wax-embedded; X800.)
tions of nonembedded pieces of iris and
ciliary body also showed an overlying fluorescence on the surface of the ciliary processes
in just those specimens receiving FITC-dex 3
and 20.
The choriocapillaris appeared permeable
to those markers with an EDR of 58A or less
(Fig. 4), whereas the 85A marker remained
within the vessels (Fig. 5). The retinal blood
vessels and pigment epithelium were impermeable to all the FITC-dextrans, even in
the very immature (5-day-old) kitten (Fig. 6).
No age-related differences were found in
the permeability characteristics of the bloodocular barriers in the kittens; similarly, the
status of the pupil at the time of enucleation
had no effect on the results. Depending on
the depth of anesthesia, the pupil was either
constricted or only partially dilated; in some
instances pupil dilation occurred during
enucleation.
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
Adult cat. The permeability of the adult
cat's iridial and especially the choroidal capillaries appeared less than that of the kitten. In
the iris, the 85A marker was observed in the
stroma adjacent to the pigment epithelial layers, but its presence was markedly less evident than in the kitten. The autofluorescence
of the adult cat's iridial chromatophores could
mask to some degree the fluorescein markers, but this did not appear solely responsible
for the diminished observance of those markers. Similar lessened fluorescence was also
observed in the adult iris with the 45A and
the 58A markers. The degree of fluorescence
in the iris concerning the 32A and 12A markers was similar in both neonatal and adult
cats.
In the choroid, the 12A marker was observed to a moderate degree in the adult, and
the 32A marker to a minimal degree. The
45A, 58A, and 85A markers did not appear to
Volume 21
Number 2
Feline blood-ocular barrier permeability 287
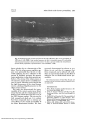
Fig. 4. Photomicrograph of retina and choroid of 5-day-old kitten after 20 inin circulation of
FITC-dex 70 (58A EDR). Note presence of dye within tapetal tissue (T) and retinal vessels
(arrows). No dye is present within retinal tissue. (Freeze-dried preparation; epi-illumination;
wax-embedded; X320.)
be permeable in the adult cat's choriocapillaris.
The ciliary processes of the adult cat
fluoresced to a similar degree for all markers
as in the kitten.
Discussion
A previous study demonstrating the marked
permeability of the feline iris capillaries to
NaFl raised the question of whether free or
protein-bound NaFl was passing the vessel
endothelium.14 It is now apparent that both
forms may do so, inasmuch as this present
study demonstrated passage of molecules as
large as 85A EDR across the endothelium of
the iris capillaries in kittens and adult cats.
These data substantiate the finding of CunhaVaz et al.17 that serum protein-bound trypan
blue also permeated the iris vessels of kittens. This species, then, is dissimilar to
man,12 monkey and rabbit,11 and rat15 in that
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
iris capillaries of those species are impermeable to both the free and protein-bound
forms of NaFl. The degree of permeability
to larger molecules was less for the adult
cat's iris capillaries than for those of the kitten; this suggests that the maturation of the
iris tissues and/or blood-aqueous barriers has
caused an alteration in the permeability of
the mature iris vessels. This phenomenon of
the alteration of vessel permeability by concurrent altered tissue requirements is evidenced in other tissues and under other circumstances.18
The role of the iris vessels in aqueous formation in the monkey has been demonstrated
to be negligible1; however, the role of the
feline iris vessels may be significant. A study
concerning totally iridectomized cats found a
lowered intraocular pressure in those cats.16
Those authors suggested that the cat's iris
vessels may be necessary to optimal aqueous
288
Bellhorn
Invest. Ophthalmol. Vis. Sci.
August 1981
Fig. 5. Photomicrograph of 21-day-old kitten choroid after 20 min circulation of FITC-dex 150
(85A EDR). Note absence of dye from tapetal tissue but marked presence within the choroidal
blood vessels and choriocapillaris (large arrow). Faint blocks of lightness (small arrow) are
autofluorescent quality of developing tapetal cells. (Freeze-dried preparation; epi-illumination; wax-embedded; X320.)
humor formation. It is interesting, though,
that they found no significant differences in
aqueous humor protein content between iridectomized and control cats. If serum proteins passing the iris capillaries are a significant part of the aqueous humor values, a
marked lowering of that value could be expected in the iridectomized cats. It is possible, though, that there was a faculative response on the part of the ciliary body epithelium in the iridectomized cats.
In this study, fluorescence was observed
on the surface of the ciliary processes in kittens and adult cats that had received FITCdex 3 and, to a lesser extent, FITC-dex 20. A
molecular seive effect for serum proteins has
been demonstrated for the blood-aqueous
barrier in man. 19 In that study, serum protein
analysis of aqueous humor samples from
normal patients showed an inverse relationship between molecular size and aqueous
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
humor concentration. We have demonstrated
a similar relationship for the dextran molecules in our in vivo study in rats6 and in this
present study in cats.
The choriocapillaris in the kitten, like the
iris capillaries, was more readily permeable
to the larger dextrans than in the adult cat.
Again it would seem that maturation of the
tissues had affected the permeability of those
vessels. In the adult cat, molecules with an
EDR as large as 32A passed from the
choriocapillaris into the tissue stroma, and
this is compatible with a study of the adult rat
choriocapillaris wherein hemeprotein markers of a similar size were permeable whereas
larger molecules were not.20
It is interesting that there was no apparent
effect on permeability of the ciliary process
capillaries based on molecular size and/or
age. Because studies of the this nature have
not been performed in other species, it is not
Volume 21
Number 2
Feline blood-ocular barrier permeability 289
Fig. 6. Photomicrograph of retina and choroid of 5-day-old kitten after 4 min circulation of
FITC-dex 3 (12A EDR). Note marked presence of dye in choroidal stroma (C) and within
wide-luniened immature retinal vessels (arrow). No dye is present within the retinal tissue.
(Freeze-dried preparation; epi-illumination; wax-embedded; xl25.)
known whether this is a characteristic of the
feline. That the ciliary process capillaries appear more permeable than the iris or choroidal capillaries may be a reflection on the
amount of bulk-flow necessary for aqueous
humor production via the ciliary process epithelium. The stroma of the ciliary process is
not extensive; in fact, there is a marked presence of vessels. Therefore it may also be that
the high fluorescence of the vessel lumens
conveys a feeling of high fluorescence in the
ciliary process stroma.
This study also demonstrated the impermeability of the retinal vessels (inner bloodretinal barrier) and the retinal pigment epithelium {outer blood-retinal barrier) to any of
the FITC-dextran markers. This was true for
both neonatal and adult cats. It is evident
that those compounds have no intrinsic adverse effects on the normal permeability of
the feline blood-retinal barriers. We have
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
previously demonstrated no adverse in vivo
effects in rats, and thus it is apparent that
these compounds can be utilized with confidence in experimental studies on the effect of
molecular size on blood-retinal barrier permeability.
The technical assistance of Mary Ellen Murphy and
Noel Roa, and the secretarial assistance of Marion Lutrin
are appreciated.
REFERENCES
1. Bill A: Blood circulation and fluid dynamics in the
eye. Physiol Rev 55:383, 1975.
2. Grotte G: Passage of dextran molecules across the
blood-lymph barrier. Acta Chir Scand [Suppl] 211:
1, 1956.
3. Caulfield JP and Farquhar MG: The permeability of
glomerular capillaries to graded dextrans. Identification of the basement membrane as the primary
filtration barrier. J Cell Biol 63:883, 1974.
4. Bellhom MB, Bellhom RW, and Poll DS: Permeability of fluorescein-labeled dextrans in fundus
290
Bellhorn
fluorescein angiography of rats and birds. Exp Eye
Res 24:595, 1977.
5. Rabkin MD, Bellhorn MB, and Bellhorn RW:
Selected molecular weight dextrans for in vivo permeability studies of rat retinal vascular disease. Exp
Eye Res 24:607, 1977.
6. Burns-Bellhorn MB, Bellhorn RW, and Benjamin
JV: Anterior segment permeability to fluoresceinlabeled dextrans in the rat. INVEST OPHTHALMOL VIS
7.
8.
9.
10.
11.
12.
SCI 17:856, 1978.
Cole DF and Munro PAG: The use of fluoresceinlabeled dextrans in investigation of aqueous humor
outflow in the rabbit. Exp Eye Res 23:571, 1976.
Tripathi RC: Uveoscleral drainage of aqueous
humor. Exp Eye Res 25(Suppl):305, 1977.
Salminen L, Chioralia G, and Schreiber S: Effect of
acute ocular hypotony on the blood-ocular barrier.
Trans Ophthalmol Soc UK 97:621, 1977.
Grayson MC and Laties AM: Ocular localization of
sodium fluorescein. Arch Ophthalmol 85:600, 1971.
Grayson MC, Tsukahara S, and Laties AM: Tissue
localization in rabbit and monkey eye of intravenously administered fluorescein. In Fluorescein
Angiography, Shimizu K, editor. Tokyo, 1974,
Kgaku Shoin Ltd. pp. 235.
McMahon RT, Tso MOM, and McClean IW: His-
Downloaded From: http://iovs.arvojournals.org/ on 08/01/2017
Invest. Ophthalmol. Vis. Sci.
August 1981
tologic localization of sodium fluorescein in human
ocular tissues. Am J Ophthalmol 80:1058, 1975.
13. Sherman SH, Green K, and Laties AM: The fate of
anterior chamber fluorescein in the monkey eye. I.
The anterior chamber outflow pathways. Exp Eye
Res 27:159, 1978.
14. Bellhorn RW: Permeability of blood-ocular barrier
of neonatal and adult cat to sodium fluorescein. INVEST OPHTHALMOL VIS SCI 19:870, 1980.
15. Szalay J, Nunziata B, and Henkind P: Permeability
of iridial blood vessels. Exp Eye Res 21:531, 1975.
16. Scheie HG, Moore E, and Adler FH: Physiology of
aqueous in completely iridectomized eyes. Arch
Ophthalmol 30:70, 1943.
17. Cunha-Vaz JG, Shakib M, and Ashton N: Studies on
the permeability of the blood-retinal barrier. I. On
the existence, development, and site of a bloodretinal barrier. Br J Ophthalmol 50:441, 1966.
18. Bellhorn RW: Control of blood vessel development.
Trans Ophthalmol Soc UK (in press).
19. Dernouchamps JP and Heremans JF: Molecular
sieve effect of the blood-aqueous barrier. Exp Eye
Res 21:289, 1975.
20. Pino RM and Essner E: Permeability of rat choriocapillaris endothelium. INVEST OPHTHALMOL VIS SCI
18(ARVO Suppl.):16, 1979.