Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Immune system wikipedia , lookup
12-Hydroxyeicosatetraenoic acid wikipedia , lookup
Molecular mimicry wikipedia , lookup
Adaptive immune system wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
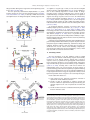
Pharmacology & Therapeutics 125 (2010) 128–137 Contents lists available at ScienceDirect Pharmacology & Therapeutics j o u r n a l h o m e p a g e : w w w. e l s ev i e r. c o m / l o c a t e / p h a r m t h e r a Associate editor: P. Foster Inhibitory receptors on myeloid cells: New targets for therapy? Ariel Munitz ⁎ Department of Microbiology and Clinical Immunology, The Sackler Faculty of Medicine, Tel-Aviv University, Tel-Aviv, Israel a r t i c l e i n f o Keywords: Inflammation Myeloid cells Inhibitory receptor Immunoreceptor tyrosine-based inhibitory motif (ITIM) a b s t r a c t Immune regulation of cellular activation is a tightly regulated process dictated by a balance of activation and inhibitory signals. Although initially described and characterized on natural killer cells, it has become increasingly apparent that inhibitory receptors are expressed and functional on myeloid cells. These receptors can override signals elicited by activation pathways including cytokine and chemokine receptors, growth factor signaling and more recently innate immune receptor signaling. Inhibitory receptors have key roles in various cellular and pathological processes and are thus potential targets for future therapeutics. In this review, the structure and function of inhibitory receptors will be discussed. Furthermore, utilization of these receptors as pharmacological targets and recent examples of strategies targeting inhibitory receptors will be analyzed. © 2009 Elsevier Inc. All rights reserved. Contents 1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2. Structure of inhibitory receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3. Inhibitory signaling: mechanism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4. Ligands for inhibitory receptors. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5. Inhibitory receptors: function. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6. Negative regulation of myeloid cell differentiation, proliferation and survival by inhibitory receptors . 7. Inhibition of cell migration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8. Inhibitory receptors in innate immune inflammatory responses. . . . . . . . . . . . . . . . . . 9. Activating ITIMs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10. Inhbitory receptors as future therapeutic target: lessons from allergy . . . . . . . . . . . . . . . 11. Concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Abbreviations: NK, Natural killer; ITIM, Immunoreceptor tyrosine-based inhibitory motif; IgSR, Immunoglubulin (Ig) super family receptors; LIR, Leukocyte immunoglobulinlike receptor; ILT, Immunoglobulin like transcript; LAIR, Leukocyte-associated immunoglobulin-like receptor; IRp60, Inhibitory receptor protein 60; Siglec, Sialic acid binding Ig-like lectins; SHP, Src homology 2 (SH2)-containing phosphates; SHIP, SH2-containing inositol phosphatase; PTB, Phosphotyrosine-binding domain; Dok, Downstream of kinase; Ras, Rat sarcoma; ERK, Extracellular signal-regulated kinase; Csk, c-src tyrosine kinase; Sirpα, Signal-regulatory protein α; RasGAP, Ras GTPase activating protein; ITAM, Immunoreceptor tyrosine-based activation motif; DAP, DNAX activation protein; MHC, Major histocompatibility complex; PIR-B, Paired immunoglobulin-like receptor B; GM-CSF, Granulocyte-macrophage colony stimulating factor; CML, Chronic myelogenous leukemia; AML, Acute myelogenous leukemia; ROS, Reactive oxygen species; IL, Interleukin; JAK, Janus kinase; GPCR, G-protein coupled receptor; LTB4, Leukotriene B4; LPS, Lipopolysaccharide. ⁎ Room 703a, Department of Clinical Immunology and Microbiology, The Sackler School of Medicine, Tel-Aviv University, Ramat Aviv, Tel-Aviv, 69978, Israel. Tel.: +972 3 6405846; fax: +972 3 6409160. E-mail addresses: [email protected], [email protected]. 0163-7258/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.pharmthera.2009.10.007 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 128 129 129 130 131 131 132 133 134 134 135 136 136 1. Introduction An intricate network of positive and negative signals mediates regulation of immune responses (Ravetch & Lanier, 2000). These signals are the basis for immune cell homeostasis in health and disease and are often achieved at the cellular level through a combination of signals from cell surface receptors, intracellular signaling intermediates and gene transcription (Daeron et al., 2008). Historically, studies on the immune system have been largely concentrated on pathways that activate immune cell function in response to antigens and pathogens. However, over the last two decades it has become increasingly apparent that in addition to pathways that stimulate immune cell activation, a counter regulatory system exists that can restrain their action and thereby promote cellular homeostasis (Cooper, 2008). These suppressive signals can regulate A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 various checkpoints in immune cell function including differentiation, maturation, adhesion, chemotaxis and mediator release. In 1998, Lanier first set the term “inhibitory receptor superfamily” in his description of receptors that suppress activation of natural killer (NK) cells (Lanier, 1998). The “classical” view of inhibitory receptors is that these receptors inhibit cell activation in trans, contain immunoreceptor tyrosine-based inhibitory motifs (ITIMs) and bind phosphatases for their inhibitory function (Lanier, 1998). In addition, they are capable of inhibiting multiple activation pathways albeit require interaction with an activating receptor (Fig. 1). Much of the data on inhibitory receptor function has emerged from studies on NK cell function or B and T cell receptor signaling (Moretta et al., 1996; Lanier, 1998; Ravetch & Lanier, 2000; Campbell & Colonna, 2001; Moretta et al., 2002). However it has become noticeable that myeloid cells including macrophages, dendritic cells, neutrophils, eosinophils and mast cells express functional inhibitory receptors that are likely to have key roles in the function of these cells, particularly in settings where these cells are recruited and activated (Munitz & Levi-Schaffer, 2004; Kanazawa et al., 2004; Katz, 2007; Munitz & Levi-Schaffer, 2007). The focus of this review will be on the expression and function of inhibitory receptors on myeloid cells. Basic structure and mechanism of action will be discussed and lessons from in vitro and in vivo studies will be illustrated. Finally, utilizing inhibitory receptors as pharmacological targets for immune suppression will be presented and discussed. 2. Structure of inhibitory receptors Inhibitory receptors can be broadly divided into two groups, belonging either to the immunoglobulin (Ig) superfamily receptors (IgSR) or the C-type lectin inhibitory receptors (Lanier, 1998; Ravetch & Lanier, 2000; Robinson et al., 2006). The IgSR is characterized by a single V-type Ig-like domain in the extracellular portion and may contain various C-type domains. Myeloid cells express various IgSR including leukocyte immunoglobulin-like receptors/immunoglobulinlike transcript (LIRs/ILTs) (Cella et al., 1997), Leukocyte-associated immunoglobulin-like receptor (LAIR) (Meyaard et al., 1997), gp49B1 (Katz, 2007), inhibitory receptor protein 60 (IRp60)/CD300a (Clark et al., 2009) and sialic acid binding Ig-like lectins (Siglecs) (Crocker 129 et al., 2007) (see Table 1). The term “C-type lectin” indicates a calciumdependent carbohydrate-binding protein motif, yet many C-type lectin inhibitory receptors posses no obvious calcium binding or carbohydrate specificity (Kanazawa et al., 2004; Robinson et al., 2006). Thus, this term is a structural rather than functional term. The C-type lectin inhibitory receptors also include proteins with a structural domain that was originally identified as a protein fold in the carbohydrate-recognition domain of mannose-binding lectin (Robinson et al., 2006). The prototypical C-type lectin inhibitory receptor is the NK receptor NKG2/CD94 or Ly49 (Held et al., 1996; Brooks et al., 1997). However, myeloid cells express various C-type lectin inhibitory receptors such as mast cell function-associated antigen (MAFA), dendritic-cell-associated C-type lectin 2 (DCAL-2) (Chen et al., 2006) and dendritic cell inhibitory receptor (DCIR) (Kanazawa et al., 2004) (Table 1). 3. Inhibitory signaling: mechanism The prototype immune inhibitory receptor (either an IgSR or C-type lectin) can be identified by a consensus amino acid sequence, the ITIM, which is present in the cytoplasmic domain of these receptors. The ITIM sequence is composed of 6 amino acids (Ile/Val/Leu/Ser)-XTyr-X-X-(Leu/Val), where X represents any amino acid. Inhibitory receptors can express either one or several ITIM domains. Upon engagement with their ligands, the ITIM motifs undergo tyrosine phosphorylation (often by a Src-family kinase), which provides a docking site for the recruitment of cytoplasmic phosphatases having a Src homology 2 (SH2) domain such as SH2-containing phosphates 1 (SHP-1), -2 and SH2-containing inositol phosphatase (SHIP-1) (Long, 1999, 2008) (Fig. 1). These phosphatases are perceived to dephosphorylate (and therefore elicit inhibition) tyrosine residues that provide docking sites for signaling kinases that are recruited by activation receptors. In general it is perceived that optimal inhibition will be achieved with maximal ITIM phosphorylation. Notably, the majority of inhibitory receptors recruit the tyrosine phosphatases SHP-1 or -2 but not the lipid phosphatase SHIP with the exception of FcγRIIB, which primarily recruits SHIP but not SHPs (Fong et al., 1996). In addition, and unlike the majority of immune inhibitory receptors, the inhibitory receptor CD200 lacks an ITIM but contains an NPXY motif in its cytoplasmic domain and can bind proteins with phosphotyrosine- Fig. 1. The mechanism of inhibitory receptor function. Upon binding to their ligand, inhibitory receptors recruit a Src-family kinase (e.g. Lyn, Fgr, Hck) that rapidly phosphorylates their intracellular ITIM domain. Consequently, SH2-containing phosphatases such as SHP-1, -2 and SHIP bind the receptor and are capable to suppress proximal activation signals mediated by signaling kinases that are recruited to activation receptors (either via ITAMs or independent of ITAMs). Alternatively, several inhibitory receptors can recruit Csk that will suppress immune cell activation independent of SHP binding. In addition, inhibitory receptors can recruit and activate the phosphotyrosine-binding adaptor molecules such as Dok. These can either bind SHIP and mediate inhibition through SHIP activation or independently bind GAP to inhibit the RAS:ERK pathway, Ptp-phosphatase. 130 A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 Table 1 Expression pattern of inhibitory receptors on myeloid cells. Inhibitory receptor Alternative name Ig-superfamily receptors FcγRIIB CD32B ILTs/LIR family ILT-2/LIR-1 CD85j ILT-3/LIR-5 CD85k ILT-4/LIR-2 CD85d ILT-5/LIR-3 CD85a Gp49B1 LILRB4 PIR-B LILRB3 LAIR-1 CD305 Sirp-1α PILRα CD200 CD300 family CD300a CD300f Inhibitory siglecs Human CD33 Siglec-5 Siglec-6 Siglec-7 Siglec-8 Siglec-9 Siglec-10 Siglec-11 Mouse CD33 Siglec-E Siglec-F Ligand Myeloid cell expressiona IgG MC, Mono, Mac, E, N, DC Mono, Mac, DC Mono, Mac, DC Mono, Mac, DC, N Mono, Mac, DC, N, E, B MC, Mono, Mac, DC, N, E MC, Mono, Mac, DC, N, E Mono, Mac, DC Collagen CD47, SP-A, SP-D Herpes Simplex Mono Virus 1 MC, Mono, Mac LMIR-1, CLM-1 CLM-8 Siglec-3 CD170 CD327 CD328, p75/AIRM CD329 C-type lectin receptors DCIR CLEC4A DCAL-2 MAFA MC, Mono, Mac, DC, N, E MC, Mono, Mac, DC, N Sialic acid " " " " Mono, Myop N, Mono, B B Mono, E " " " " MC, E Mono, Mac, N Mono, E Mono, Mac " " " Mono, Mac, N, E N, Mono, Mac, DC E DC, Mac, N DC MC This table represents a partial list only of receptors mentioned in this review. MC—mast cell, Mono—monocyte, Mac—macrophage, DC—dendritic cell, N—neurophil, E—eosinophil, B—basophi, MyoP—myeloid progenitor celll. a The expression data has not been reported for all myeloid cell types. binding domains (PTBs) such as downstream of kinase (Dok) family members (Barclay et al., 2002; Zhang & Phillips, 2006). Interestingly, Dok family members have been shown to interact with SHP and SHIP molecules and inhibit rat sarcoma (Ras)-Extracellular signal-regulated kinase (Erk) mediated activation (Berg et al., 1999; Robson et al., 2004; Zhang & Phillips, 2006) (Fig. 1). It is important to note that despite the consistent findings of inhibitory receptors binding SH2 containing phosphatases (i.e. SHP-1, SHP-2, SHIP-1), the precise inhibitory mechanism is not fully understood. Two of the main questions regarding the inhibitor mechanism are: 1) Is there a differential role for SHP-1 and SHP-2 mediated inhibition? 2) Is the inhibitory pathway dependent of phosphatase activity and if so to what extent? Rationalizing these queries are a few examples. For instance, SHP-1 rapidly associates with the inhibitory receptor gp49B1 after co-ligation of gp49B1 with FcεRI in mast cells. Yet, SHP-1-deficient mast cells exhibited only a partial loss of gp49B1-mediated inhibition of exocytosis, revealing a central, but not exclusive, SHP-1 requirement in the counter-regulatory pathway, which may be SHP-2-dependent (Lu-Kuo et al., 1999). Furthermore, recent studies utilizing a unique structural approach revealed that differential binding of SHP-1 might have a distinctive inhibitory outcome than recruitment and binding of SHP-2. This analysis uncovered that SHP-1 is most active when actually bound to the ITIM receptor whereas SHP-2, which possesses a longer spacing between its two C-terminal tyrosines, can be active even when unbound to the ITIM (Lu et al., 2001; Zhang et al., 2003). Thus, specific recruitment and consequent binding of phosphatase to the intracellular ITIM may govern the extent and character of inhibition that is elicited by inhibitory receptors. In addition to the aforementioned studies, recent data highlight a possibility for ITIM-bearing receptors to inhibit even in the absence of SHPs. For example, upon phosphorylation, LAIR-1 can bind the c-Src tyrosine kinase (Csk), which is a negative regulator of Src family kinases and inhibit B cell receptor induced activation even in the absence of SHP-1 and -2 (Verbrugge et al., 2006). The binding of inhibitory receptors to Csk and consequent SHP-independent inhibition is likely a shared phenomenon between inhibitory receptors as the ITIM of immunoglobulin-like transcript (ILT) 2 and signalregulatory protein α (SIRP-1α) can also bind Csk (Fig. 1) (Sayos et al., 2004). In addition, negative adaptor molecules such as Dok family members may also mediate inhibition that is either mediated via SHIP/SHPs or independently through direct binding of Dok to Ras GTPase activating protein (RasGAP) (Berg et al., 1999; Robson et al., 2004; Zhang & Phillips, 2006). The simplistic and traditional view of inhibitory receptor function described the ITIM as an opposing signal to immune receptor tyrosinebased activation motif (ITAM) phosphorylation (Long, 2008). These motifs are present on various adaptor molecules including DNAX activation protein 10 (DAP10) and DAP12 as well as on the Fcγ chain (Wu et al., 2000). Nevertheless, recent data demonstrating negative regulation of inhibitory receptors towards receptors that do not contain ITAMs indicate a broader inhibitory capacity for these receptors (Sinclair, 1999; Dietrich et al., 2000; Brown, 2005). In fact, inhibitory receptors are relatively outnumbered by activation pathways (including cytokine/chemokine receptors, pattern recognition receptors, adhesion molecules, Fc receptors, complement receptors, co-stimulatory molecules, etc.) and a single inhibitory receptor can suppress multiple pathways. For example, CD300a was found to inhibit both G-protein coupled receptor signaling and cytokine receptor signaling (Munitz et al., 2006a). PIR-B can inhibit chemokine receptor activation, integrin signaling as well as innate immune receptor mediated-responses (Pereira et al., 2004; Zhang et al., 2005; Nakayama et al., 2007; Munitz et al., 2008; Torii et al., 2008). Thus, suggesting a relative dominance of inhibitory signaling over activation. Furthermore, this also suggests redundancy as one ITIM-bearing receptor can regulate multiple activation pathways. 4. Ligands for inhibitory receptors Assuming such an important role for inhibitory receptors in cellular homeostasis, one would predict that a) the ligands for inhibitory signaling would be widely expressed and b) each cell type can express various inhibitory receptors with redundant functions. Early studies in the NK and cytotoxic T cell field have illustrated the “missing self” hypothesis in which NK inhibitory receptors recognize a repertoire of major histocompatibility complex (MHC) class I molecules that prevent cellular activation upon recognition with a “self” cell (Colonna, 1996; Held et al., 1996; Moretta et al., 1996; Lanier, 1998; Ravetch & Lanier, 2000). However, upon viral infection or acquiring a malignant phenotype, MHC molecules are significantly downregulated and therefore the cytotoxic cell “loses” its inhibition thus becoming activated by the activation receptors that are present on its surface. While this paradigm is well understood for cells that are specialized in cell-mediated cytotoxicity, the biological rationale for MHC recognition by inhibitory receptors on myeloid cells is not quite clear (Sinclair, 1999). As characterization of inhibitory receptors on myeloid cells expanded it became evident that another class of inhibitory recognition that is independent of MHC binding is present on myeloid (and also NK) cells (Lebbink & Meyaard, 2007). For example, the A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 ligand for gp49B1 has been reported to be the αVβ3 integrin and collagens have been shown to serve as functional, high affinity ligands for the inhibitory immune receptor LAIR-1 (Castells et al., 2001; Lebbink et al., 2006). Interestingly, even paired immunoglobulin-like receptor B (PIR-B), which was originally identified as a myeloidspecific inhibitory receptor that binds MHC class I molecules, has been recently shown to recognize and bind various bacteria (Takai, 2005; Masuda et al., 2007; Nakayama et al., 2007). Substantiating a role for non-MHC-restricted inhibitory signaling are also recent reports demonstrating that the inhibitory receptor paired immunoglobulinlike type 2 receptor alpha (PILRα) recognizes CD99, which is widely expressed on all leukocytes (Shiratori et al., 2004). Furthermore, ligand(s) for other inhibitory receptors such as IRp60/CD300a are yet to be defined. In fact, CD300a was considered at one point to be a potential NK cell inhibitory molecule, but it failed to bind to HLA-class I molecules (Cantoni et al., 1999; Clark et al., 2009), making it unlikely that MHC class I molecules are its natural ligand. Future studies defining the ligands of myeloid-associated inhibitory receptors are likely to contribute to our understanding of the physiological role for such receptors in immune regulation. 5. Inhibitory receptors: function All of the abovementioned evidence suggests a broad role for inhibitory receptors in suppressing myeloid cell function. Cells from the myeloid lineage including macrophages, neutrophils and eosinophils respond to different cytokines, chemokines and express differential receptors. Nevertheless, the main pathway of myeloid cell recruitment into the inflamed tissue is similar and may be divided into a few checkpoints (Fig. 2). In general, myeloid cells originate in the bone marrow where they undergo initial differentiation and proliferation and thereafter enter the blood stream. During an inflammatory process, chemokines are secreted which induce the recruitment of these cells into the inflamed area where they are further stimulated, can secrete their specific mediators and/or interact with other cells (Rothenberg, 1998; Gordon & Taylor, 2005). This 131 paradigm highlights multiple checkpoints, which can be potentially regulated by inhibitory receptors and will be discussed below. 6. Negative regulation of myeloid cell differentiation, proliferation and survival by inhibitory receptors Hematopoeisis is a multi-step process in which the development of each cell along the different cell lineages occurs as a consequence of a sequential effect of growth factors and transcription factors. In this process, the cells can acquire or lose expression of cell surface molecules including lineage markers and inhibitory receptors. For example, the expression of CD33, an inhibitory receptor belonging to the siglec family of receptors, corresponds with the cell commitment towards the myelomonocytic precursors (Dexter & Spooncer, 1987). In fact, CD33 expression is maintained in the myeloid and monocytic lineage, whereas it is lost in mature neutrophils (Vitale et al., 1999, 2001). CD33 is homologous to siglec-7, which is also highly expressed by cells of the myeloid lineage. Flow cytometeric analysis of the expression of CD33 and siglec-7 on cord blood derived CD34+ cells that have been cultured in the presence of stem cell factor (SCF) and granulocyte-macrophage colony stimulating factor (GM-CSF) revealed that all CD34+ cells steadily expressed CD33 as early as day 4 in culture. On the other hand and under similar conditions, siglec-7 expression was gradually acquired and only 50% of the cultured cells expressed siglec-7 by day 10 under the same conditions (Vitale et al., 1999, 2001). Despite differential kinetics in expression, antibody cross-linking of siglec-7 or CD33 were capable to suppress in vitro proliferation of myeloid cell precursors. In fact, cross-linking of both receptors inhibited the proliferation of leukemic cells obtained from chemotherapy-free patients with chronic myeloid leukemia (Vitale et al., 1999, 2001). While this may suggest a broad non-specific capability of inhibitory receptors to suppress proliferation, this does not appear to be the case since CD300a/IRp60, which is expressed in a similar fashion to CD33, does not inhibit CD34+ myeloid cell proliferation or chronic myelogenous leukemia (CML) proliferation (Mingari et al., 2001). Interestingly, examination of the effects of all Fig. 2. Cellular checkpoints regulated by inhibitory receptors. Myeloid cells including polymorphonuclear cells (PMN) such as neutrophils and eosinophils and monocytes originate in the bone marrow and enter the blood. Upon inflammatory stimulus or tissue injury these cells transmigrate to the tissue through a process of adhesion and chemotaxis. In the tissue, and under the influence of local cytokines monocytes undergo final maturation to macrophages or dendritic cells (MΦ and DC). All of the aforementioned cellular processes can be regulated by inhibitory receptors. 132 A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 three receptors (i.e. CD33, siglec-7 and CD300a/IRp60) on the proliferation of acute myeloid leukemic cells revealed that siglec-7 acts to suppress acute myelogenous leukemia (AML) cell proliferation whereas CD33 induces their apoptosis and cross-linking of CD300a/ IRp60 has no effect (Vitale et al., 2001; Mingari et al., 2001). Another line of evidence supporting a role for inhibitory receptors in myeloid cell proliferation and apoptosis arise from studies conducted on LAIR-1. Similar to the previous inhibitory receptors discussed above, engagement of LAIR-1 prevents GM-CSF-induced proliferation of AML cell blasts. LAIR-1 activation blocked the cells in the G0/G1 phase of cell cycle and the cells underwent apoptosis within 4 days of LAIR-1 activation (Zocchi et al., 2001). Mechanistically, this latter phenomenon was associated with inhibition of GM-CSFinduced Ca2+ mobilization and activation of the AKT1/protein kinase B α (PKBα) pathway (Zocchi et al., 2001). It is important to note that LAIR-1 was capable of inducing its inhibitory effect even on AML blasts that do not express CD33. Thus the inhibitory activity of LAIR-1 is CD33-independent. Although cells obtained from myeloid leukemia provides a good experimental system to dissect the role of inhibitory receptors on cell proliferation, apoptosis and survival, an important question is whether they can influence these parameters in non-proliferating cells such as mature eosinophils or neutrophils. Recently it has been shown that activation of siglec-8 on human eosinophils by means of antibody cross linking suppressed their survival by initiating mitochondrial injury, reactive oxygen species (ROS) generation, rapid cleavage of caspase-3, -8 and -9 and inducing apoptosis (Nutku et al., 2003, 2005). This effect was even further enhanced in the presence of interleukin (IL)-5, IL-3 and GM-CSF, the hallmark eosinophil “survival cytokines” which seem to prime siglec-8 activity and diminish the need for secondary antibody cross-linking (Nutku et al., 2003; Nutku-Bilir et al., 2008). In contrast to the observations with siglec-8, which actively induced eosinophil apoptosis, cross-linking of CD300a/IRp60 on human eosinophils suppressed their survival in response to the same “survival cytokines” (Munitz et al., 2006a). As such, cross-linking of CD300a/IRp60 inhibited IL-5 and GM-CSF signaling and thus suppressed survival signals rather than actively induce apoptosis. Supporting this notion was the finding that the inhibitory effect of CD300a/IRp60 was only seen in the presence of IL-5 or GM-CSF and quite interestingly the ability of CD300a/IRp60 to suppress eosinophil activation was proportional to cytokine concentration. Thus, increased concentration of IL-5/GM-CSF enhances the inhibitory effect (Munitz et al., 2003). In the case of CD300a/IRp60 the inhibitory process is likely due to a crosstalk between CD300a/IRp60 and the IL5/GM-CSF/IL-3 common β chain and involves the recruitment of intracellular phosphatases and consequent inhibition of Janus kinas (JAK) 2 phosphorylation (Munitz et al., 2006a). Unlike CD300a, CD300LF (an additional CD300-inhibitory receptor belonging to the CD300 family of IgS receptors) has been recently shown to actively induce cell death in macrophages in an ITIM-independent caspaseindependent fashion (Can et al., 2008). Thus, the molecular basis for the negative regulation of survival by inhibitory receptors may not be dependent on ITIM-activity. Similar to siglec-8 on eosinophils, siglec-9 ligation can induce neutrophil apoptosis. Interestingly, neutrophils obtained from patients with acute septic shock or rheumatoid arthritis demonstrated increased siglec-9 expression and activity. Mechanistically, siglec-9 was capable to induce an apoptotic (ROS- and caspase-dependent) and non-apoptotic (ROS-dependent) death pathways (von Gunten et al., 2005). The findings that cytokines and/or disease state can increase the inhibitory capability of such receptors require further investigation. Inhibitory receptors can also negatively regulate myeloid cell differentiation. The potent activity of these receptors in this process may be illustrated by recent observations demonstrating a key role for PIR-B and CD300LF in osteoclast formation and development (Chung et al., 2003; Mori et al., 2008). Both PIR-B and CD300LF were downregulated in receptor activator for nuclear factor κ B ligand (RANKL)- and macrophage colony-stimulating factor (M-CSF)induced osteoclast formation. In fact, deletion of PIR-B lead to accelerated osteoclastogenesis and forced expression of CD300LF prevents osteoclastogenesis induced by RANKL and transforming growth factor (TGF)-β1 (Chung et al., 2003; Mori et al., 2008). Of note, the aforementioned LAIR-1 was implicated in myeloid cell differentiation as well. In vitro studies have shown the LAIR-1 was capable to inhibit GM-CSF-induced monocyte differentiation into dendritic cells. Interestingly, this effect was specific to LAIR-1 since stimulation of FDF03, a highly similar inhibitory receptor, had no effect (Fournier et al., 2000). Collectively, these data suggest nonredundant roles and mechanisms for inhibitory receptor function in the regulation of myeloid cell proliferation, survival and differentiation. Nevertheless, there is currently limited amount of data regarding the functions of these receptors in bone marrow-derived cells and addressing their function in vivo. 7. Inhibition of cell migration Being the sentinels of host defense, myeloid leukocytes respond to perturbations in immune homeostasis by rapid mobilization into the site of injury or infection. The directed migration of these cells is often coordinated in response to chemokines. These small (∼10–20 kDa) secreted proteins induce the migratory effect through interactions with heterotrimeric GPCRs on the responding cell. In response to chemokine stimulation, GPCRs induce a signaling cascade that is accompanied by Ca2+ mobilization and activation of Ras:ERKdependent pathways (Cotton & Claing, 2009). These events are often associated with interactions of the GPCR with Src-family kinases (Rozengurt, 2007). Recent data indicate a direct cross talk between GPCR signaling and inhibitory receptor signaling. Zhang et al., have shown that the kinases feline sarcoma viral (Fgr) and hemopoietic cell kinase (Hck), which are responsible for the baseline ITIM phosphorylation of the inhibitory receptor PIR-B act as negative regulators of neutrophil and dendritic cell migration (Zhang et al., 2005). While the exact mechanism is not fully understood, it appears that under baseline conditions, the ITIMs in PIR-B are phosphorylated by Hck and Fgr leading to association of PIR-B with SHP-1 and -2 and subsequent dephosphorylation of yet unknown targets leading to cell arrest. Upon binding of chemokines to GPCRs, PIR-B is dephosphorylated, the inhibitory effect of SHP-1 dismantled and therefore inhibition of migratory responses is decreased. Indeed, neutrophils and dendritic cells that lack PIR-B or SHP-1 display enhanced chemokine signaling and functional responses, as do Fgr and Hck deficient cells (Brown, 2005; Zhang et al., 2005). We have recently shown that PIR-B may actually have a dual role in the regulation of eosinophil and neutrophil migration. In response to chemokine signaling PIR-B serves as a negative regulator of GPCR signaling via interactions with SHP-1 and -2. However, upon stimulation with leukotriene B4 (LTB4), an arachidonic acid metabolite that induces cell migration (also via GPCR signaling) PIR-B serves as a co-activator of cell migration (Munitz et al., 2008). In attempt to define the downstream molecular events that may cause this duality, biochemical analysis revealed that PIR-B differentially interacts with Src-family kinases or kinase adaptor molecules to either suppress or co-activate migratory responses (Munitz et al., 2008) (Fig. 3). In human cells, we have recently shown that antibody crosslinking of IRp60/CD300a on eosinophils suppresses their response to eotaxin-induced migration and inhibits eotaxin-induced Ca2+ mobilization and ERK and p38 activation (Munitz et al., 2006a). Similarly, it has been reported that activation of dendritic cells with a ligand to the inhibitory receptor LIR-1/ILT-2 (a human PIR-B orthologue) inhibits their response to the chemokine C–C ligand 5 (CCL5) (Wagner et al., 2008). A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 133 Fig. 3. The molecular basis for the dual role of PIR-B. PIR-B has been recently shown to possess a dual function in respect to eosinophil (and neutrophil) chemotaxis. Upon stimulation of eosinophils with eotaxin that binds the CCR3 receptor, PIR-B undergoes rapid tyrosine phosphorylation and recruits the phosphatases SHP-1 and -2. This leads to inhibition of GPCR-induced Erk activation and therefore PIR-B has an inhibitory role. In contrast, upon stimulation of eosinophils with the arachidonic acid metabolite LTB4 (via the BLT1 receptor), PIR-B is capable to recruit various kinases and adaptor molecules inclusing Btk, JAK1/2, Shc and Crk. Consequently leading to activation of ERK signaling and thus PIR-B serves as an co-activator for LTB4-induced signaling. It is important to note that inhibitory receptors have been also shown to regulate myeloid cell adhesion and integrin signaling (Pereira et al., 2004). Thus, negative regulation of adhesive properties of myeloid cells by inhibitory receptors may also add to the overall suppressive activity that has been observed and reported from in vivo chemotaxis studies. However, in vitro findings clearly point to a direct effect of inhibitory receptors in suppression of chemokine receptor signaling that is independent of integrin signaling. Supporting this, increased Ca2+ mobilization (a process that is negatively regulated by inhibitory receptors) in response to chemokine stimulation is independent of integrin signaling and results from direct stimulation of phospholipase C (PLC) β by the βγ subunit of stimulated Gi heterotrimeric G proteins (Brown, 2005). Taken together these results clearly demonstrate a key role for inhibitory receptors in regulation of myeloid cell migration. 8. Inhibitory receptors in innate immune inflammatory responses Following recruitment, myeloid cells are important in coordinating inflammatory responses. Not only do they generate proinflammatory agents in the course of the phagocytic activities or release toxic mediators, but they also produce cytokines that modulate the inflammatory response including IL-6, TNF-α, IL-1β, IL-8, GM-CSF and others (Taylor & Gordon, 2003). Innate immune signaling components have key roles in multiple diseases including autoimmune, metabolic and allergic diseases (Ravetch & Lanier, 2000). Experimentation in the innate immune field has been able to delineate a complex network of receptors (e.g. Toll-like receptors, nucleotide-binding oligomerization domain (NOD) family receptors) and intracellular adaptor molecules and kinases that are readily available for the immediate response of the host cell (Akira et al., 2006). Given the role of myeloid cells in innate immunity, one would assume that their innate functions could be counter regulated by inhibitory receptors. Indeed, accumulating data indicate a role of inhibitory receptors in the regulation of innate immune inflammatory responses. A common phenomenon to various inhibitory receptors is that proinflammatory mediators can regulate the expression of inhibitory receptors. Thus, proinflammatory mediators can influence the “threshold” of immune cell inhibition either by upregulating or downregulating inhibitory receptor expression. Stimulation of human neutrophils with lipopolysaccharide (LPS) and GM-CSF increases the cell surface expression of IRp60/CD300a as a result of the rapid translocation of an intracellular pool of the receptor to the cell surface (Alvarez et al., 2008). Intradermal injection of LPS increases the expression of gp49B1 on mouse bone marrow and blood neutrophils (Zhou et al., 2003). Furthermore, the levels of PIR-B on osteoclasts and macrophages are also upregulated by TNF-α and LPS (Ochi et al., 2007 and Munitz et al., unpublished data). In contrast, stimulation of neutrophils and macrophages with LPS downregulated the expression of the inhibitory receptors DCIR and SIRP-1α (Richard et al., 2003; Kong et al., 2007). The expression patterns of these ITIM-bearing receptors in response to inflammatory stimuli suggest an important role for these receptors in the control of inflammation. In support of this concept, various reports have demonstrated an in-vivo role for inhibitory receptors in innate immune responses. It has been recently shown that gp49B1 is a negative regulator of LPS induced activation of neutrophils (Zhou et al., 2003; Katz, 2007). Intradermal LPS injection in mice deficient in gp49B1 resulted in significantly more intravascular thrombi, occluding venules in comparison with wild type mice. In addition, neutrophil accumulation was increased in the vascular thrombi and in the ear of the gp49B1 knockout mice. It is interesting to note that this effect is due to a specific inhibition of neutrophils responses to LPS by gp49B1 but not mast cell responses. In fact, the increased pathology that was observed in the absence of gp49B1 was prevented following neutrophil depletion (Zhou et al., 2003; Katz, 2007). Another example for such a role comes from SIRP-1α. Mice that have been transferred with 134 A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 SIRP-1α-deficient macrophages were highly susceptible to endotoxic shock, developing multiple organ failure and exhibited a remarkable increase in mortality. Of note the inhibitory activity of SIRP-1α was mainly due to the recruitment of SHP-2 and prevention of LPSinduced SHP-2:IKK complex formation (Kong et al., 2007). Similar to SIRP-1α, PIR-B has been recently shown to regulate macrophage function in response to bacterial activation. Upon binding and recognition of bacterial by various receptors, macrophages are activated and secrete large amounts of pro-inflammatory cytokines. Interestingly, PIR-B deficient mice display increased susceptibility to Salmonella infection (Torii et al., 2008). Furthermore, PIR-B has been also shown to bind gram-positive and gram-negative bacteria such as S. aureus and E. coli and to suppress macrophage activation in response to S. aureus activation in vitro. This result is particularly interesting as it indicates that inhibitory receptors may serve as pattern recognition receptors to bacterial products (Nakayama et al., 2007). Although it is assumed that inhibitory receptors operate mainly via recruitment of phosphatases and subsequent dephosphorylation of signaling targets an additional mechanism for suppression of inflammatory responses by inhibitory receptors has been proposed. Recently, it has been shown that siglec-9 and -5 can enhance (rather than inhibit) IL-10 production in response to multiple TLR ligands in a process that is dependent on tyrosine phosphorylation of intracellular ITIMs (Ando et al., 2008). The ability to enhance IL-10 production is not specific for siglecs as PIR-B was also described to increase IL-10 production in response to S. aureus stimulation (Nakayama et al., 2007). Thus, inhibitory receptors may regulate immune activation and inflammation via an-IL-10 dependent pathway. In fact, and in addition to the aforementioned inhibitory receptors, FcγRIIB, a well-established and characterized inhibitory receptor (Daeron et al., 2008) can suppress innate immune responses via binding to IgG. In fact, passively immunized mice with Abs to LPS to reversed proinflammatory cytokine production, and the transfer of macrophages whose FcγR had been cross-linked (and thereby activated and display increased IL-10 production and decreased IL-12) could rescue mice from lethal endotoxemia (Gerber & Mosser, 2001; Grazia Cappiello et al., 2001; Kane & Mosser, 2001). Thus, in-vivo activation of the macrophage FcγR, can be exploited to prevent inappropriate inflammatory cytokine responses. Indeed, elegant studies in human patients with active visceral leishmaniasis indicate that IgG (the ligand of FcγR) can cause a novel form of immune enhancement due to its ability to induce IL-10 production from macrophages (Miles et al., 2005). Although the majority of the studies on inhibitory receptors in innate immunity examine their role in TLR-dependent activation, inhibitory receptors can also suppress non TLR-dependent pathways. Recently CD24-deficient mice have been shown to exhibit increased susceptibility to danger- but not pathogen-associated molecular patterns (Chen et al., 2009). CD24 can associate with high mobility group box 1 (HMGB1), heat shock protein 70 (Hsp70), and heat shock protein 90 (Hsp90) and negatively regulates their pro-inflammatory activity resulting in inhibition of nuclear factor kappa B (NFκB) activation. The inhibitory effect of CD24 is mediated at least in part via association with Siglec-10 or Siglec-G in human and mice, respectively (Chen et al., 2009). Importantly, inhibitory receptors belonging to the c-type lectin family of receptors have been also shown to possess innate immune roles by suppression of dendritic cell function via receptors such as DC-SIGN (Relloso et al., 2002; Gagliardi et al., 2005; den Dunnen et al., 2009). However the role of such receptors has been recently reviewed elsewhere and will thus not be included in this chapter (Geijtenbeek & Gringhuis, 2009). Taken together, these studies indicate that inhibitory receptors utilize distinct mechanisms that are capable to suppress innate immune responses in a cell-specific manner. Future studies are likely to shed light on the exact mechanism by which these receptors operate and how they regulate specific innate components. 9. Activating ITIMs The involvement of inhibitory receptors in the negative regulation of myeloid cell function highlights them as potential targets for therapy. However, when taking into account their inhibitory role, one should also consider another side for these receptors. Recent data has suggested that ITIM-bearing receptors can actually mediate activation and ITAM-bearing receptors could induce inhibition (Barrow & Trowsdale, 2006; Pinheiro da Silva et al., 2008). Although the subject of activation through ITIMs is not fully understood, there are now various reports on reciprocal activity for such receptors. As mentioned previously, we have recently shown that in response to LTB4 stimulation, PIR-B can serve as a docking site for various adaptor kinases that can promote cellular activation. In fact, our data support a model in which following LTB4 activation PIR-B can interact with several activating kinases, such as JAK1, JAK2, Shc, and Crk (Fig. 4) (Munitz et al., 2008). Similar to PIR-B, the inhibitory receptor SIRP-1α, was recently reported to stimulate nitric oxide production in macrophages via the Jak/STAT and PI3K/Rac1 pathways, thereby suggesting an activating role for SIRP-1α in a similar molecular mechanism to that of PIR-B (Alblas et al., 2005). Furthermore, the inhibitory receptors TREM-like transcript-1 (TLT-1) and cholecystokinin (CCK) receptor have been also shown to enhance cellular activation (Barrow & Trowsdale, 2006). For example TLT-1, amplifies FcεRI-mediated calcium signaling and CCK (via recruitment of SHP-2 to its ITIM) activates the AKT pathway (Vatinel et al., 2006). Thus, although it appears that the primary role of inhibitory receptors is to indeed suppress cellular activation, under specific circumstances they can also induce cellular activation. 10. Inhbitory receptors as future therapeutic target: lessons from allergy As a therapeutic approach to suppress inflammation, one can target activation pathways and inhibit them. Numerous approaches have used this strategy and several biological agents targeting cytokines, angiogenic factors, adhesion molecules and kinase inhibitors are either in development, clinical trials or in actual use. However, the findings that inhibitory receptors display key roles in dampening immune cell activation highlights them as a potential target for therapeutic intervention as well. In this set-up, the anticipated drug will act to enhance inhibition rather than to suppress activation. One of the best examples for usage of inhibitory receptors as therapeutic targets comes from studies in the allergy field. Several groups including ours have utilized inhibitory receptors for anti-allergic treatment in an overall similar approach of bispecific-antibodies or -fusion proteins (Zhu et al., 2002; Tam et al., 2004; Zhu et al., 2005; Bachelet et al., 2006; Munitz et al., 2006b; Saxon et al., 2008). Notably, the use of similar strategy by three independent groups reveals the necessity to consider designing molecular targets that will display cell-specific regulation. Allergic responses are initiated by mast cell degranulation in response to allergen cross-linking of FcεRI-bound IgE (Fig. 4A). Thus, specific targeting of mast cells (but not general suppression of other myeloid cells) may yield a potent anti-allergic therapeutic approach. Saxon et al., have undertaken two approaches targeting inhibitory pathways in mast cells and basophils (Fig. 4B) (Zhu et al., 2002, 2005). One platform uses negative signaling to drive nonallergen-specific suppression of allergic reactivity. The other platform uses a similar negative pathway but is actually allergen-specific (Fig. 4B). These two drug modalities can be used either to chronically treat allergic patients independent of their allergenicity or specifically “tailored” for the patient to distinctively target an antigen. In general, both platforms (termed GE2 and GFD respectively) are based on a fusion protein that inhibits FcεRI mediated responses by cross-linking it to FcγRIIB (GE2) or cross-links FcγRIIB with Feld1 a well known cat A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 allergen. Studies utilizing these reagents have shown promising results in-vivo (Zhu et al., 2002, 2005). We have previously described that IRp60/CD300a is a potent negative regulator of mast cell and eosinophil responses (Bachelet et al., 2005; Munitz et al., 2006a). In attempt to inhibit mast cell and eosinophil functions we designed bispecific antibody fragments that 135 are capable to recognize IgE or CCR3 (as mast cell and eosinophil specific molecules) and CD300a/IRp60 (Fig. 4C). In vivo administration of these antibody fragments in murine models of allergic peritonitis, passive cutaneous anaphylaxis and a chronic model of established allergic eosinophilic airway inflammation was capable to suppress the allergic response and even reverse the inflammatory process and associated remodeling (Bachelet et al., 2006; Munitz et al., 2006b). Similar to this approach, Tam et al. have designed a bispecific antibody against IgE and FcγRIIB that inhibits antigeninduced histamine release by human mast cells and basophils in-vitro (Tam et al., 2004). As mentioned, inhibitory receptors can interact with various inhibitory adaptor proteins such as Dok-1 (Fig. 1) (Berg et al., 1999; Saxon et al., 2004; Robson et al., 2004; Zhang & Phillips, 2006). Interestingly, activation of mast cells with nanomolar concentrations of dexamethasone, upregulates the expression and association of Dok-1 with RasGAP (Hiragun et al., 2005). Thus, it is possible that lowdose glucocorticosteroid treatment will potentiate inhibitory receptorbased therapeutic approaches that are aimed to cross-link (and thereby activate) inhibitory receptors. Collectively, these independent and timely reports highlight the vast potential and possibilities that inhibitory receptors may have as future therapeutic targets including inflammatory, autoimmune, allergic and malignant disorders. However, when targeting these receptors one should take into account various factors including cell-specificity, general immune-suppression and the inflammatory context, which may cause ITIMs to co-activate rather than inhibit. 11. Concluding remarks Since the description of the first inhibitory motif in FcγRIIB (Daeron, 1995), the number of ITIM-bearing receptors has noticeably increased. A recent protein sequence analysis that is based on sequence context (i.e. predictions of signal peptide, transmembrane helices and protein domains) it is now estimated that 109 proteins express ITIMs and may represent novel inhibitory receptor candidates (Staub et al., 2004). Currently, there is still need to enhance our understanding on the role of such receptors in various aspects of myeloid cell biology and to define these pathways capable of inhibiting their functions. Identification of the in-vivo ligands and definition of receptor:ligand interactions using gene targeted mice is likely to shed light on the potential of these receptors to serve as therapeutic targets. Future studies will enable us to: A) Gain insight into the molecular mechanisms involved in myeloid cell cellular inhibition/activation. B) Provide us with an opportunity to suppress myeloid cell function in different experimental settings and thereby gain valuable insight into the role of myeloid cells in health and disease. C) Provide us with novel tools to combat detrimental myeloid cell functions in disease states. Fig. 4. (A) The allergic/inflammatory response is triggered by cross-linking of Fcε by allergen-bound IgE. In the absence of inhibitory signals, the FcεRI receptor chain ITAMs are phosphorylated and mediate mast cell activation and initiation of inflammation. (B) In attempt to utilize the ability of FcγRIIB to inhibit IgE-mediated activation of mast cells and basophils, a platform of two fusion proteins has been designed. A bispecific fusion protein consisting of the Fc portion of IgG (Fcγ) and the Fc portion of IgE (Fcε) were linked (GE2). This fusion protein cross-links FcγRIIB to FcεRI and activates the inhibitory mechanism of FcγRIIB. Similarly, another fusion protein linking a major cat allergen (FelD1) to Fcγ was designed. This fusion protein mimics allergen binding to FcεRI portion and induces allergen-specific cellular inhibition. (C) An additional approach that targets inhibitory receptors as anti-allergic drugs utilizes chemically conjugated bispecific antibodies that cross-link FcεRI-bound IgE to IRp60/Cd300a (LE1) or FcγRIIB (AT10XE10) and therefore activate the inhibitory mechanism of IRp60/ CD300a or FcγRIIB. 136 A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 As the rates of diseases involving expansion, recruitment and activation of myeloid cells is currently on the rise, studies that are focused on pathways that inhibit rather than activate are likely to prove key in the regulation of allergic, autoimmune, inflammatory and metabolic diseases. These may represent novel pathways for future therapy. Acknowledgments The authors wish to thank Drs Marc Rothenberg, Nives Zimmerman and Joe Sherill for critically reviewing the manuscript, Dr. Simon Hogan for the helpful discussions and insights and to Dr. Ido Bachelet for assistance with generating the figures. Grant support: This work was supported by a Research Fellowship Award from the American Heart Association and internal Tel-Aviv University Funds. References Akira, S., Uematsu, S., & Takeuchi, O. (2006). Pathogen recognition and innate immunity. Cell 124, 783−801. Alblas, J., Honing, H., de Lavalette, C. R., Brown, M. H., Dijkstra, C. D., & van den Berg, T. K. (2005). Signal regulatory protein alpha ligation induces macrophage nitric oxide production through JAK/STAT- and phosphatidylinositol 3-kinase/Rac1/NAPDH oxidase/H2O2-dependent pathways. Mol Cell Biol 25, 7181−7192. Alvarez, Y., Tang, X., Coligan, J. E., & Borrego, F. (2008). The CD300a (IRp60) inhibitory receptor is rapidly up-regulated on human neutrophils in response to inflammatory stimuli and modulates CD32a (FcgammaRIIa) mediated signaling. Mol Immunol 45, 253−258. Ando, M., Tu, W., Nishijima, K., & Iijima, S. (2008). Siglec-9 enhances IL-10 production in macrophages via tyrosine-based motifs. Biochem Biophys Res Commun 369, 878−883. Bachelet, I., Munitz, A., & Levi-Schaffer, F. (2006). Abrogation of allergic reactions by a bispecific antibody fragment linking IgE to CD300a. J Allergy Clin Immunol 117, 1314−1320. Bachelet, I., Munitz, A., Moretta, A., Moretta, L., & Levi-Schaffer, F. (2005). The inhibitory receptor IRp60 (CD300a) is expressed and functional on human mast cells. J Immunol 175, 7989−7995. Barclay, A. N., Wright, G. J., Brooke, G., & Brown, M. H. (2002). CD200 and membrane protein interactions in the control of myeloid cells. Trends Immunol 23, 285−290. Barrow, A. D., & Trowsdale, J. (2006). You say ITAM and I say ITIM, let's call the whole thing off: the ambiguity of immunoreceptor signalling. Eur J Immunol 36, 1646−1653. Berg, K. L., Siminovitch, K. A., & Stanley, E. R. (1999). SHP-1 regulation of p62(DOK) tyrosine phosphorylation in macrophages. J Biol Chem 274, 35855−35865. Brooks, A. G., Posch, P. E., Scorzelli, C. J., Borrego, F., & Coligan, J. E. (1997). NKG2A complexed with CD94 defines a novel inhibitory natural killer cell receptor. J Exp Med 185, 795−800. Brown, E. J. (2005). Leukocyte migration: dismantling inhibition. Trends Cell Biol 15, 393−395. Campbell, K. S., & Colonna, M. (2001). Human natural killer cell receptors and signal transduction. Int Rev Immunol 20, 333−370. Can, I., Tahara-Hanaoka, S., Hitomi, K., Nakano, T., Nakahashi-Oda, C., Kurita, N., et al. (2008). Caspase-independent cell death by CD300LF (MAIR-V), an inhibitory immunoglobulin-like receptor on myeloid cells. J Immunol 180, 207−213. Cantoni, C., Bottino, C., Augugliaro, R., Morelli, L., Marcenaro, E., Castriconi, R., et al. (1999). Molecular and functional characterization of IRp60, a member of the immunoglobulin superfamily that functions as an inhibitory receptor in human NK cells. Eur J Immunol 29, 3148−3159. Castells, M. C., Klickstein, L. B., Hassani, K., Cumplido, J. A., Lacouture, M. E., Austen, K. F., et al. (2001). gp49B1-alpha(v)beta3 interaction inhibits antigen-induced mast cell activation. Nat Immunol 2, 436−442. Cella, M., Dohring, C., Samaridis, J., Dessing, M., Brockhaus, M., Lanzavecchia, A., et al. (1997). A novel inhibitory receptor (ILT3) expressed on monocytes, macrophages, and dendritic cells involved in antigen processing. J Exp Med 185, 1743−1751. Chen, C. H., Floyd, H., Olson, N. E., Magaletti, D., Li, C., Draves, K., et al. (2006). Dendriticcell-associated C-type lectin 2 (DCAL-2) alters dendritic-cell maturation and cytokine production. Blood 107, 1459−1467. Chen, G. Y., Tang, J., Zheng, P., & Liu, Y. (2009). CD24 and Siglec-10 selectively repress tissue damage-induced immune responses. Science 323, 1722−1725. Chung, D. H., Humphrey, M. B., Nakamura, M. C., Ginzinger, D. G., Seaman, W. E., & Daws, M. R. (2003). CMRF-35-like molecule-1, a novel mouse myeloid receptor, can inhibit osteoclast formation. J Immunol 171, 6541−6548. Clark, G. J., Ju, X., Tate, C., & Hart, D. N. (2009). The CD300 family of molecules are evolutionarily significant regulators of leukocyte functions. Trends Immunol 30, 209−217. Colonna, M. (1996). Natural killer cell receptors specific for MHC class I molecules. Curr Opin Immunol 8, 101−107. Cooper, M. D. (2008). Inhibition of immune cell function. Immunol Rev 224, 7−10. Cotton, M., & Claing, A. (2009). G protein-coupled receptors stimulation and the control of cell migration. Cell Signal 21, 1045−1053. Crocker, P. R., Paulson, J. C., & Varki, A. (2007). Siglecs and their roles in the immune system. Nat Rev Immunol 7, 255−266. Daeron, M. (1995). Intracytoplasmic sequences involved in the biological properties of low-affinity receptors for IgG expressed by murine macrophages. Braz J Med Biol Res 28, 263−274. Daeron, M., Jaeger, S., Du Pasquier, L., & Vivier, E. (2008). Immunoreceptor tyrosinebased inhibition motifs: a quest in the past and future. Immunol Rev 224, 11−43. den Dunnen, J., Gringhuis, S. I., & Geijtenbeek, T. B. (2009). Innate signaling by the C-type lectin DC-SIGN dictates immune responses. Cancer Immunol Immunother 58, 1149−1157. Dexter, T. M., & Spooncer, E. (1987). Growth and differentiation in the hemopoietic system. Annu Rev Cell Biol 3, 423−441. Dietrich, J., Nakajima, H., & Colonna, M. (2000). Human inhibitory and activating Ig-like receptors which modulate the function of myeloid cells. Microbes Infect 2, 323−329. Fong, D. C., Malbec, O., Arock, M., Cambier, J. C., Fridman, W. H., & Daeron, M. (1996). Selective in vivo recruitment of the phosphatidylinositol phosphatase SHIP by phosphorylated Fc gammaRIIB during negative regulation of IgE-dependent mouse mast cell activation. Immunol Lett 54, 83−91. Fournier, N., Chalus, L., Durand, I., Garcia, E., Pin, J. J., Churakova, T., et al. (2000). FDF03, a novel inhibitory receptor of the immunoglobulin superfamily, is expressed by human dendritic and myeloid cells. J Immunol 165, 1197−1209. Gagliardi, M. C., Teloni, R., Giannoni, F., Pardini, M., Sargentini, V., Brunori, L., et al. (2005). Mycobacterium bovis Bacillus Calmette–Guerin infects DC-SIGN — dendritic cell and causes the inhibition of IL-12 and the enhancement of IL-10 production. J Leukoc Biol 78, 106−113. Geijtenbeek, T. B., & Gringhuis, S. I. (2009). Signalling through C-type lectin receptors: shaping immune responses. Nat Rev Immunol 9, 465−479. Gerber, J. S., & Mosser, D. M. (2001). Stimulatory and inhibitory signals originating from the macrophage Fcgamma receptors. Microbes Infect 3, 131−139. Gordon, S., & Taylor, P. R. (2005). Monocyte and macrophage heterogeneity. Nat Rev Immunol 5, 953−964. Grazia Cappiello, M., Sutterwala, F. S., Trinchieri, G., Mosser, D. M., & Ma, X. (2001). Suppression of Il-12 transcription in macrophages following Fc gamma receptor ligation. J Immunol 166, 4498−4506. Held, W., Cado, D., & Raulet, D. H. (1996). Transgenic expression of the Ly49A natural killer cell receptor confers class I major histocompatibility complex (MHC)-specific inhibition and prevents bone marrow allograft rejection. J Exp Med 184, 2037−2041. Hiragun, T., Peng, Z., & Beaven, M. A. (2005). Dexamethasone up-regulates the inhibitory adaptor protein Dok-1 and suppresses downstream activation of the mitogen-activated protein kinase pathway in antigen-stimulated RBL-2H3 mast cells. Mol Pharmacol 67, 598−603. Kanazawa, N., Tashiro, K., & Miyachi, Y. (2004). Signaling and immune regulatory role of the dendritic cell immunoreceptor (DCIR) family lectins: DCIR, DCAR, dectin-2 and BDCA-2. Immunobiology 209, 179−190. Kane, M. M., & Mosser, D. M. (2001). The role of IL-10 in promoting disease progression in leishmaniasis. J Immunol 166, 1141−1147. Katz, H. R. (2007). Inhibition of pathologic inflammation by leukocyte Ig-like receptor B4 and related inhibitory receptors. Immunol Rev 217, 222−230. Kong, X. N., Yan, H. X., Chen, L., Dong, L. W., Yang, W., Liu, Q., et al. (2007). LPS-induced down-regulation of signal regulatory protein alpha contributes to innate immune activation in macrophages. J Exp Med 204, 2719−2731. Lanier, L. L. (1998). NK cell receptors. Annu Rev Immunol 16, 359−393. Lebbink, R. J., & Meyaard, L. (2007). Non-MHC ligands for inhibitory immune receptors: novel insights and implications for immune regulation. Mol Immunol 44, 2153−2164. Lebbink, R. J., de Ruiter, T., Adelmeijer, J., Brenkman, A. B., van Helvoort, J. M., Koch, M., et al. (2006). Collagens are functional, high affinity ligands for the inhibitory immune receptor LAIR-1. J Exp Med 203, 1419−1425. Long, E. O. (1999). Regulation of immune responses through inhibitory receptors. Annu Rev Immunol 17, 875−904. Long, E. O. (2008). Negative signaling by inhibitory receptors: the NK cell paradigm. Immunol Rev 224, 70−84. Lu, W., Gong, D., Bar-Sagi, D., & Cole, P. A. (2001). Site-specific incorporation of a phosphotyrosine mimetic reveals a role for tyrosine phosphorylation of SHP-2 in cell signaling. Mol Cell 8, 759−769. Lu-Kuo, J. M., Joyal, D. M., Austen, K. F., & Katz, H. R. (1999). gp49B1 inhibits IgE-initiated mast cell activation through both immunoreceptor tyrosine-based inhibitory motifs, recruitment of src homology 2 domain-containing phosphatase-1, and suppression of early and late calcium mobilization. J Biol Chem 274, 5791−5796. Masuda, A., Nakamura, A., Maeda, T., Sakamoto, Y., & Takai, T. (2007). Cis binding between inhibitory receptors and MHC class I can regulate mast cell activation. J Exp Med 204, 907−920. Meyaard, L., Adema, G. J., Chang, C., Woollatt, E., Sutherland, G. R., Lanier, L. L., et al. (1997). LAIR-1, a novel inhibitory receptor expressed on human mononuclear leukocytes. Immunity 7, 283−290. Miles, S. A., Conrad, S. M., Alves, R. G., Jeronimo, S. M., & Mosser, D. M. (2005). A role for IgG immune complexes during infection with the intracellular pathogen Leishmania. J Exp Med 201, 747−754. Mingari, M. C., Vitale, C., Romagnani, C., Falco, M., & Moretta, L. (2001). p75/AIRM1 and CD33, two sialoadhesin receptors that regulate the proliferation or the survival of normal and leukemic myeloid cells. Immunol Rev 181, 260−268. Moretta, A., Bottino, C., Mingari, M. C., Biassoni, R., & Moretta, L. (2002). What is a natural killer cell? Nat Immunol 3, 6−8. Moretta, A., Bottino, C., Vitale, M., Pende, D., Biassoni, R., Mingari, M. C., et al. (1996). Receptors for HLA class-I molecules in human natural killer cells. Annu Rev Immunol 14, 619−648. A. Munitz / Pharmacology & Therapeutics 125 (2010) 128–137 Mori, Y., Tsuji, S., Inui, M., Sakamoto, Y., Endo, S., Ito, Y., et al. (2008). Inhibitory immunoglobulin-like receptors LILRB and PIR-B negatively regulate osteoclast development. J Immunol 181, 4742−4751. Munitz, A., & Levi-Schaffer, F. (2004). Eosinophils: ‘new’ roles for ‘old’ cells. Allergy 59, 268−275. Munitz, A., & Levi-Schaffer, F. (2007). Inhibitory receptors on eosinophils: a direct hit to a possible Achilles heel? J Allergy Clin Immunol 119, 1382−1387. Munitz, A., Bachelet, I., Eliashar, R., Moretta, A., Moretta, L., & Levi-Schaffer, F. (2006a). The inhibitory receptor IRp60 (CD300a) suppresses the effects of IL-5, GM-CSF, and eotaxin on human peripheral blood eosinophils. Blood 107, 1996−2003. Munitz, A., Bachelet, I., & Levi-Schaffer, F. (2006b). Reversal of airway inflammation and remodeling in asthma by a bispecific antibody fragment linking CCR3 to CD300a. J Allergy Clin Immunol 118, 1082−1089. Munitz, A., McBride, M. L., Bernstein, J. S., & Rothenberg, M. E. (2008). A dual activation and inhibition role for the paired immunoglobulin-like receptor B in eosinophils. Blood 111, 5694−5703. Munitz, A., Piliponsky, A. M., & Levi-Schaffer, F. (2003). IgE-independent activation of human mast cells indicates their role in the late phase reaction of allergic inflammation. Cell Tissue Bank 4, 25−28. Nakayama, M., Underhill, D. M., Petersen, T. W., Li, B., Kitamura, T., Takai, T., et al. (2007). Paired Ig-like receptors bind to bacteria and shape TLR-mediated cytokine production. J Immunol 178, 4250−4259. Nutku, E., Aizawa, H., Hudson, S. A., & Bochner, B. S. (2003). Ligation of Siglec-8: a selective mechanism for induction of human eosinophil apoptosis. Blood 101, 5014−5020. Nutku, E., Hudson, S. A., & Bochner, B. S. (2005). Mechanism of Siglec-8-induced human eosinophil apoptosis: role of caspases and mitochondrial injury. Biochem Biophys Res Commun 336, 918−924. Nutku-Bilir, E., Hudson, S. A., & Bochner, B. S. (2008). Interleukin-5 priming of human eosinophils alters siglec-8 mediated apoptosis pathways. Am J Respir Cell Mol Biol 38, 121−124. Ochi, S., Shinohara, M., Sato, K., Gober, H. J., Koga, T., Kodama, T., et al. (2007). Pathological role of osteoclast costimulation in arthritis-induced bone loss. Proc Natl Acad Sci U S A 104, 11394−11399. Pereira, S., Zhang, H., Takai, T., & Lowell, C. A. (2004). The inhibitory receptor PIR-B negatively regulates neutrophil and macrophage integrin signaling. J Immunol 173, 5757−5765. Pinheiro da Silva, F., Aloulou, M., Benhamou, M., & Monteiro, R. C. (2008). Inhibitory ITAMs: a matter of life and death. Trends Immunol 29, 366−373. Ravetch, J. V., & Lanier, L. L. (2000). Immune inhibitory receptors. Science 290, 84−89. Relloso, M., Puig-Kroger, A., Pello, O. M., Rodriguez-Fernandez, J. L., de la Rosa, G., Longo, N., et al. (2002). DC-SIGN (CD209) expression is IL-4 dependent and is negatively regulated by IFN, TGF-beta, and anti-inflammatory agents. J Immunol 168, 2634−2643. Richard, M., Thibault, N., Veilleux, P., Breton, R., & Beaulieu, A. D. (2003). The ITIMbearing CLECSF6 (DCIR) is down-modulated in neutrophils by neutrophil activating agents. Biochem Biophys Res Commun 310, 767−773. Robinson, M. J., Sancho, D., Slack, E. C., LeibundGut-Landmann, S., & Reis, C. (2006). e Sousa: Myeloid C-type lectins in innate immunity. Nat Immunol 7, 1258−1265. Robson, J. D., Davidson, D., & Veillette, A. (2004). Inhibition of the Jun N-terminal protein kinase pathway by SHIP-1, a lipid phosphatase that interacts with the adaptor molecule Dok-3. Mol Cell Biol 24, 2332−2343. Rothenberg, M. E. (1998). Eosinophilia. N Engl J Med 338, 1592−1600. Rozengurt, E. (2007). Mitogenic signaling pathways induced by G protein-coupled receptors. J Cell Physiol 213, 589−602. Saxon, A., Kepley, C., & Zhang, K. (2008). “Accentuate the negative, eliminate the positive”: engineering allergy therapeutics to block allergic reactivity through negative signaling. J Allergy Clin Immunol 121, 320−325. Saxon, A., Zhu, D., Zhang, K., Allen, L. C., & Kepley, C. L. (2004). Genetically engineered negative signaling molecules in the immunomodulation of allergic diseases. Curr Opin Allergy Clin Immunol 4, 563−568. Sayos, J., Martinez-Barriocanal, A., Kitzig, F., Bellon, T., & Lopez-Botet, M. (2004). Recruitment of C-terminal Src kinase by the leukocyte inhibitory receptor CD85j. Biochem Biophys Res Commun 324, 640−647. 137 Shiratori, I., Ogasawara, K., Saito, T., Lanier, L. L., & Arase, H. (2004). Activation of natural killer cells and dendritic cells upon recognition of a novel CD99-like ligand by paired immunoglobulin-like type 2 receptor. J Exp Med 199, 525−533. Sinclair, N. R. (1999). Why so many coinhibitory receptors? Scand J Immunol 50, 10−13. Staub, E., Rosenthal, A., & Hinzmann, B. (2004). Systematic identification of immunoreceptor tyrosine-based inhibitory motifs in the human proteome. Cell Signal 16, 435−456. Takai, T. (2005). Paired immunoglobulin-like receptors and their MHC class I recognition. Immunology 115, 433−440. Tam, S. W., Demissie, S., Thomas, D., & Daeron, M. (2004). A bispecific antibody against human IgE and human FcgammaRII that inhibits antigen-induced histamine release by human mast cells and basophils. Allergy 59, 772−780. Taylor, P. R., & Gordon, S. (2003). Monocyte heterogeneity and innate immunity. Immunity 19, 2−4. Torii, I., Oka, S., Hotomi, M., Benjamin, W. H., Jr., Takai, T., Kearney, J. F., et al. (2008). PIR-Bdeficient mice are susceptible to Salmonella infection. J Immunol 181, 4229−4239. Vatinel, S., Ferrand, A., Lopez, F., Kowalski-Chauvel, A., Esteve, J. P., Fourmy, D., et al. (2006). An ITIM-like motif within the CCK2 receptor sequence required for interaction with SHP-2 and the activation of the AKT pathway. Biochim Biophys Acta 1763, 1098−1107. Verbrugge, A., Rijkers, E. S., de Ruiter, T., & Meyaard, L. (2006). Leukocyte-associated Iglike receptor-1 has SH2 domain-containing phosphatase-independent function and recruits C-terminal Src kinase. Eur J Immunol 36, 190−198. Vitale, C., Romagnani, C., Falco, M., Ponte, M., Vitale, M., Moretta, A., et al. (1999). Engagement of p75/AIRM1 or CD33 inhibits the proliferation of normal or leukemic myeloid cells. Proc Natl Acad Sci U S A 96, 15091−15096. Vitale, C., Romagnani, C., Puccetti, A., Olive, D., Costello, R., Chiossone, L., et al. (2001). Surface expression and function of p75/AIRM-1 or CD33 in acute myeloid leukemias: engagement of CD33 induces apoptosis of leukemic cells. Proc Natl Acad Sci U S A 98, 5764−5769. von Gunten, S., Yousefi, S., Seitz, M., Jakob, S. M., Schaffner, T., Seger, R., et al. (2005). Siglec-9 transduces apoptotic and nonapoptotic death signals into neutrophils depending on the proinflammatory cytokine environment. Blood 106, 1423−1431. Wagner, C. S., Walther-Jallow, L., Buentke, E., Ljunggren, H. G., Achour, A., & Chambers, B. J. (2008). Human cytomegalovirus-derived protein UL18 alters the phenotype and function of monocyte-derived dendritic cells. J Leukoc Biol 83, 56−63. Wu, J., Cherwinski, H., Spies, T., Phillips, J. H., & Lanier, L. L. (2000). DAP10 and DAP12 form distinct, but functionally cooperative, receptor complexes in natural killer cells. J Exp Med 192, 1059−1068. Zhang, S., & Phillips, J. H. (2006). Identification of tyrosine residues crucial for CD200Rmediated inhibition of mast cell activation. J Leukoc Biol 79, 363−368. Zhang, H., Meng, F., Chu, C. L., Takai, T., & Lowell, C. A. (2005). The Src family kinases Hck and Fgr negatively regulate neutrophil and dendritic cell chemokine signaling via PIR-B. Immunity 22, 235−246. Zhang, Z., Shen, K., Lu, W., & Cole, P. A. (2003). The role of C-terminal tyrosine phosphorylation in the regulation of SHP-1 explored via expressed protein ligation. J Biol Chem 278, 4668−4674. Zhou, J. S., Friend, D. S., Feldweg, A. M., Daheshia, M., Li, L., Austen, K. F., et al. (2003). Prevention of lipopolysaccharide-induced microangiopathy by gp49B1: evidence for an important role for gp49B1 expression on neutrophils. J Exp Med 198, 1243−1251. Zhu, D., Kepley, C. L., Zhang, K., Terada, T., Yamada, T., & Saxon, A. (2005). A chimeric human-cat fusion protein blocks cat-induced allergy. Nat Med 11, 446−449. Zhu, D., Kepley, C. L., Zhang, M., Zhang, K., & Saxon, A. (2002). A novel human immunoglobulin Fc gamma Fc epsilon bifunctional fusion protein inhibits Fc epsilon RI-mediated degranulation. Nat Med 8, 518−521. Zocchi, M. R., Pellegatta, F., Pierri, I., Gobbi, M., & Poggi, A. (2001). Leukocyte-associated Ig-like receptor-1 prevents granulocyte-monocyte colony stimulating factordependent proliferation and Akt1/PKB alpha activation in primary acute myeloid leukemia cells. Eur J Immunol 31, 3667−3675.