Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Axon guidance wikipedia , lookup
Multielectrode array wikipedia , lookup
Optogenetics wikipedia , lookup
Eyeblink conditioning wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Subventricular zone wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Synaptogenesis wikipedia , lookup
Development of the nervous system wikipedia , lookup
Node of Ranvier wikipedia , lookup
Neuroanatomy wikipedia , lookup
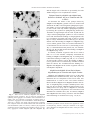
Journal of Neuropathology and Experimental Neurology Copyright q 2004 by the American Association of Neuropathologists Vol. 63, No. 5 May, 2004 pp. 461 473 Neuron-Binding Human Monoclonal Antibodies Support Central Nervous System Neurite Extension ARTHUR E. WARRINGTON, PHD, ALLAN J. BIEBER, PHD, VIRGINIA VAN KEULEN, MS, BOGOLJUB CIRIC, PHD, LARRY R. PEASE, PHD, AND MOSES RODRIGUEZ, MD Abstract. Two human IgMs (sHIgM12 and sHIgM42) were identified that supported in vitro central nervous system (CNS) neurite extension equal to the potent neurite stimulatory molecule laminin. Both IgMs bound to multiple cell types in unfixed CNS tissue and to the surface of neurons in culture. Both monoclonal antibodies (mAbs) overrode the inhibitory effect of CNS mouse myelin on granule cell neurite extension. Neither mAb bound to the surface of mature oligodendrocytes or strictly colocalized with myelin proteins. Sialidase treatment eliminated the neuronal surface binding of both mAbs, whereas blocking sphingolipid synthesis with Fumonisin B1 or removing GPI-linked proteins with PIPLC did not. When used as substrates for mixed neuron/glia aggregates, sHIgM12 and sHIgM42 supported robust neurite extension while astrocytes remained in the aggregates. In contrast, laminin supported astrocyte migration and spreading. Human mAbs that support neurite extension are novel factors that may be of use in encouraging axon repair following injury while minimizing glial cell infiltration. Both human mAbs were isolated from individuals with monoclonal gammopathy. Each individual has carried high mAb titers in circulation for years without detriment. sHIgM12 and sHIgM42 are therefore unlikely to be systemically pathogenic. Key Words: Cerebellar granule cell; Human monoclonal antibody; Laminin; Monoclonal gammopathy; Myelin; Neurite extension; Sialic acid. INTRODUCTION It is generally accepted that axons of the adult brain and spinal cord do not successfully regenerate following injury due to the synthesis of a glial scar at the lesion site and the presence of a number of inhibitory molecules exposed following the breakdown of myelin (1–6). Central nervous system (CNS) axons retain their ability to regenerate when provided with an appropriate environment, such as peripheral nerve graphs, where CNS axons will extend significant distances (7, 8). Despite such environmental hurdles, histological studies dating to Ramon y Cajal (9) revealed that the adult CNS does have a limited capacity of endogenous repair. Following injury, spinal cord tissue distal to the site of injury maintained aspects of normal histology and axon responsiveness (10, 11). Intact white matter does not inhibit axon extension to the degree that degenerated white matter does, for when CNS neurons were transplanted beyond the site of injury, axons extended within the white matter (12, 13). Therefore, a period following injury likely exists when therapeutic interventions to encourage axon growth through the area of damaged myelin and to limit glial scar formation can be beneficial. Therapeutic strategies to enhance axon regeneration fall into 2 general categories—strategies to block endogenous inhibitory factors From Departments of Neurology (AEW, AJB, MR) and Immunology (VVK, BC, LRP, MR), Mayo Clinic College of Medicine, Rochester, Minnesota. Correspondence to: Arthur E. Warrington, PhD, Department of Neurology, Guggenheim 401, Mayo Clinic College of Medicine, 200 First St. SW, Rochester, MN 55905. E-mail: [email protected] Supporting Grants: NIH grant NS24180, the National Multiple Sclerosis Society RG3172, the Multiple Sclerosis Society of Canada. and strategies to provide a factor or substrate that overrides inhibitors to axon extension. There is considerable evidence that antibodies (Abs) can modulate CNS repair. Administration of a mAb against an axon growth inhibitory fraction of myelin improved axon regeneration through the lesioned area (2). It was hypothesized that the mAb partially neutralized in vivo inhibitors to axon extension. Polyclonal Abs synthesized in situ in response to white matter immunization may mimic immune-mediated aspects of an endogenous CNS repair response. In the Theiler’s murine encephalomyelitis virus (TMEV)-mediated model of multiple sclerosis (MS), immunization with spinal cord homogenate (SCH) prior to spinal cord demyelination resulted in significantly more remyelination (14). The direct transfer of sera or purified immunoglobulin (Ig) from SCH-immunized, uninfected mice to non-immunized, TMEV-infected mice promoted the same degree of remyelination as pre-immunization (15, 16). Analogous immunization of mice with SCH prior to axonal injury resulted in more axon extension through the lesion site than that observed in non-immunized mice (17). Although it has not been directly proven that Abs are the functional agents in the axon regeneration studies, this is implied by several observations. Within individual mice an elevated titer of Abs directed against myelin correlated with the degree of axon extension, and the SCH anti-sera blocked myelinderived neurite outgrowth inhibition in vitro. Since the SCH anti-sera was thought to promote axon extension by blocking endogenous outgrowth inhibitors, it was surprising to discover that the level of Abs to known CNS inhibitors of axon regrowth (MAG, Nogo, CSPGs) were not elevated over non-immunized mice (18). 461 462 WARRINGTON ET AL Using binding to myelin and oligodendrocytes (OLs) as an initial screen, a number of mouse and human mAbs were identified that promoted remyelination in the TMEV-mediated model of demyelination (19, 20). Several of the mAbs that promoted remyelination bound to known OL plasma membrane lipids. The ability of a mAb to promote remyelination in vivo correlated with the ability to induce transient Ca21 influx in OLs in culture (21). It is reasonable to propose that mAbs that promote remyelination interact in vivo directly with the cells responsible for myelination. Similarly, if the SCH anti-sera that promoted axon regeneration acted directly upon neurons, then neuronbinding mAbs may encourage axon outgrowth in vivo. Such mAbs would offer new therapeutic strategies to influence recovery from CNS injury. To begin to test this hypothesis, a collection of human sera obtained from individuals with monoclonal gammopathy, a condition characterized by high concentrations of monoclonal serum mAb, was screened for the presence of mAbs that bind to neurons. This report details the identification of 2 human mAbs that bound to the surface of neurons, supported robust neurite extension when presented as substrates, and overrode the neurite extension inhibition of CNS myelin. Human mAbs that support neurite outgrowth are novel molecules that may be useful factors to encourage axon extension following injury. MATERIALS AND METHODS Antibodies Hybridomas A2B5, R24, and HNK-1 were obtained from ATCC (Manassas, VA). A2B5 is a mouse IgM anti-ganglioside (22, 23). R24 is a mouse IgG anti-GD3 (24). O4 is a mouse IgM anti-sulfatide (25, 26). The mouse IgG anti-MOG clone 8– 18C5 was obtained from C. Linnington. Mouse anti-Thy-1.1 (clone OX-7) was from BD PharMingen (San Jose, CA). IgG producing hybridomas were grown in serum-free, protein-free medium (Sigma, St. Louis, MO) and purified by ammonium sulfate precipitation, a protein A column and dialysis against PBS. IgM producing hybridomas were grown in 50% serumfree, protein-free medium, 50% RPMI 1640 (Sigma) with 1% FCS (Life Technologies, Grand Island, NY). IgMs were purified by PEG precipitation, water precipitation, and a superose-6 column. Purity and concentrations were determined by SDSPAGE. Fluorochrome conjugated affinity purified F(ab9)2 fragments of goat anti-mouse IgM mm chain specific, anti-mouse IgG Fcg fragment specific, anti-human IgM Fc5m fragment specific, and anti-human IgG Fcg fragment specific are from Jackson ImmunoResearch (West Grove, PA). Human Abs Human serum samples obtained from the Mayo Clinic dysproteinemia clinic were selected solely by the presence of an Ig clonal peak of greater than 20 mg/ml. Sera were from 102 patients with a variety of conditions, including Waldenström’s macroglobulinemia, multiple myeloma, lymphoma, and MUGUS. IgM-containing sera were dialyzed against water and separated on a Superose-6 column. IgM fractions were analyzed by SDS-PAGE and pooled. Human IgG-containing sera were diluted 1:20 for testing. Human peripheral B cells from normal adults, adults with rheumatoid arthritis (AKJR), adults with MS and from fetal cord blood (CB) were immortalized using Epstein-Barr virus (ATCC #CRL 1612). Ab-containing supernatants were tested directly. Cerebellar Slice Immunocytochemistry SJL mice were obtained from Jackson Labs (Bar Harbor, ME). Holtzman Sprague-Dawley rats were obtained from Harlan (Indianapolis, IN). All animal handling and procedures were done in accordance with the guidelines and approval of the Mayo Clinic and Foundation Institutional Animal Care Committee. Cerebellar slice immunocytochemistry was performed as described (27). Abs were used at 10 mg/ml in HEPES-buffered Earle’s balanced salt solution (E/H) or as undiluted supernatant with 1% BSA. A fluorescently conjugated F(ab9)2 fragment was used for visualization. Slices were mounted with Vectashield (Vector Labs, Burlingame, CA) and 10 mg/ml bisbenzimide for nuclear localization. Images were obtained with an Olympus Provis epifluorescent microscope and a SPOT digital camera (Diagnostic Instruments Inc., Sterling Heights, MI). Cell Culture Granule neurons were prepared as previously described, with some modifications (28). Cells from P6 rat cerebella were suspended in DMEM/F12 with 10% FCS, 6.0 g/l glucose, 0.1 mg/ ml penicillin/streptomycin, 2 mM glutamine, and 25 mM KCl (29) and plated onto plastic Petri dishes (BD PharMingen) to allow glial adherence. Non-adherent cells were passed through a 30-mm mesh and resuspended in Neurobasal/B27 (1:50), N2 (1:100), glutamine, pen/strep (all from Life Technologies, Carlsbad, CA), 6.0 g/l glucose, and 25 mM KCl. Glial contamination was never more than 2% as determined by immunocytochemistry. Purified OLs were isolated as in (30) and differentiated for 12 days in DMEM/F12/N2. For surface → Fig. 1. Screening for human mAbs that bound to CNS cells. Slices of unfixed cerebellum from 8 week SJL/J mice were incubated with 10 mg/ml purified human mAb or undiluted immortalized B-cell clone supernatant followed by appropriate fluorochrome-conjugated F(ab9)2 fragments. Two human mAbs bound strongly to a single cerebellar layer, AKJR4 (G), identified the internal granular layer (IGL), whereas sHIgM22 (D) identified the central white matter (WM) of the folia. Additional human mAbs bound to several cerebellar layers. sHIgM12 (A) and sHIgM42 (E) labeled the molecular layer (ML), the IGL and the WM. Double label immunocytochemistry using sHIgM12 (A) and a mAb directed against the mature myelin protein, MOG (B) J Neuropathol Exp Neurol, Vol 63, May, 2004 HUMAN mAbs SUPPORT NEURITE EXTENSION 463 demonstrated common label of WM. sHIgM14 (C) bound to the Purkinje cells and their dendritic arborizations within the ML as well as the WM. sHIgM46 (F) bound to WM, the IGL, Purkinje cell soma, and small cells within the ML. All of the reactive human mAbs were of the IgM isotype. One of the many non-reactive human IgMs was exemplified by sHIgM39 (H). The highlight of cerebellar fissures at the pial surface (*) was non-specific and observed with many mAbs. ML: molecular layer, WM: central white matter of the folia, PC: Purkinje cell layer, IGL: internal granular layer. Scale bar: 500 mm. J Neuropathol Exp Neurol, Vol 63, May, 2004 464 WARRINGTON ET AL immunocytochemistry, cells were seeded at 50,000 cells/cm2; granule cells onto poly-D-lysine-(PDL)-coated coverslips (Fisher Scientific, Chicago, IL); OLs onto poly-L-ornithine-coated coverslips. Immunolabeling was performed on ice. After blocking with 4% BSA E/H, cells were incubated with Abs at 10 mg/ ml in 1% BSA in E/H. After washing and fixation with 4% PF, fluorochrome-conjugated F(ab9)2 fragments were used for visualization. Aggregates of cerebellar cells were prepared as described (31). Ten ml containing 3.5 3 104 cerebellar cells, prior to the removal of glia, were spotted onto the lid of a 96-well dish and the lid inverted to create hanging drops. The resulting mixed aggregates were seeded onto nitrocellulose-treated laminin or mAb-coated wells. After 48 h, aggregates were fixed and stained with Coomassie blue for bright field microscopy. Parallel wells were double immunolabeled with anti-neurofilament 150 kd (Chemicon, Temecula, CA) and anti-GFAP (BD PharMingen) to localize neuronal processes and astrocytes. a Drawing Slate II and KS 400 graphics software. Statistical comparisons were done using Kruskal-Wallis 1-way ANOVA. Neurite Outgrowth Assay Human mAbs Bound to the CNS Nitrocellulose-coated 48-well tissue culture plastic (Costar) was prepared as described (32). Wells were incubated with mAbs or mouse laminin (Life Technologies) in DMEM/F12 for 16 h at 48C. Plates were washed and filled with DMEM/F12 until cell seeding. Granule cells were seeded at 20,000 cells/ cm2 in Neurobasal/B27 medium. Cultures were incubated until significant neurite extension was observed on the laminin-coated wells, normally 18 to 20 h. Cultures were fixed and neurites visualized with Coomassie blue stain. Ten to twelve 0.27 mm2 adjacent frames from at least 2 identical wells were recorded for each substrate in each experiment as bright-field digital images at a magnification of 3200. Granule cell attachment was determined using these images. Neurite length was determined using a Drawing Slate II (CalComp, Columbia, MD) and KS 400 graphics software (Kontron Elektronic, GmbH). Neurites were measured only if the end of the neurite could be visualized. Neurites were measured whether or not they contacted another soma or neurite. Five hundred to 800 neurites were measured for each group in at least 3 separate cultures. The frequency distribution of neurite length was determined and displayed using Microsoft Excel and SigmaPlot for Windows (Jandel Scientific Software, San Rafael, CA). Statistical comparisons were done using Kruskal-Wallis 1-way ANOVA on ranks pairwise in SigmaStat (Jandel). Human IgMs reactive to the surface of OLs promoted repair of spinal cord lesions in the TMEV-mediated mouse model of MS and in lysolecithin-induced demyelination (20, 34). We hypothesized that human mAbs that bind to the surface of neurons may be potential neuronal signaling molecules. Sera from 102 humans with a variety of conditions characterized by a monoclonal Ig spike and supernatants from 74 human B cell clones were screened for binding to unfixed slices of mouse and rat cerebellum. Unfixed tissue slices were used because freezing or even light fixation of tissue can compromise the reactivity of Abs that bind to lipids or carbohydrates (27, 35). Nine human mAbs (all IgMs) of 176 mAbs tested, bound to neuronal layers in slices of cerebellum (Table). The binding patterns of several human mAbs to cerebellar slices are presented in Figure 1. Several mAbs bound to multiple morphologically distinct layers (Fig. 1A, C, E, F). Two human mAbs (AKJR4 and sHIgM22) were specific for 1 cerebellar layer. The B-cell line AKJR4 produced a mAb that bound to the internal granular layer (IGL) (Fig. 1G). When viewed at high magnification the AKJR4 mAb defined individual granule cell soma (data not shown). sHIgM22 bound to the central white matter of the folia (WM) and myelinated tracts in the IGL (Fig. 1D), similar to that observed with antiMOG mAb (Fig. 1B). sHIgM14 labeled Purkinje cell soma at the slice surface, radial strands within the molecular layer (ML), and the WM (Fig. 1C). sHIgM46 bound to the WM and the Purkinje cell layer (Fig. 1F). sHIgM12 and sHIgM42 (Fig. 1A, E), bound to multiple cerebellar layers. sHIgM12 clearly labels the WM and ML in Figure 1A and at higher magnification Purkinje and granule cells (data not shown). A nearly confluent label at the cut surface of the tissue slice over the ML suggested that sHIgM12 or sHIgM42 may bind to an extracellular matrix molecule. The patterns in which human Neurite Extension on Myelin Assays were done as previously described (17). Forty-eightwell plates were coated with nitrocellulose (as above) and 25 mg/ml PDL. Two mg of SJL/J mouse spinal cord myelin, prepared according to (33), in a 20-ml drop was placed in the center of each well and dried. After washing, 100 ml of 10 mg/ml mAbs or laminin were added and the plates incubated overnight at 48C. Wells were washed 23 with DMEM/F12 and 25,000 granule cells seeded in Neurobasal/B27. After 24 h, cultures were fixed with 4% PF and labeled with anti-neurofilament 150 kd. Images were collected from 3 separate wells at 3200 using an Olympus inverted microscope equipped with a SPOT digital camera. Ten to 12 adjacent images were collected beginning 300 mm inward from the edge of the myelin spot. The mean length of at least 300 neurites per group was determined using J Neuropathol Exp Neurol, Vol 63, May, 2004 Enzyme and Inhibitor Treatment Sialidase from C. perfringens (Worthington Biochemicals Corp, Lakewood, NJ), which cleaves N-acetyl neuraminic acid from a variety of glycoproteins and glycolipids, was added to granule cells at 1 day in culture at a final concentration of 0.1 U/ml for 24 h. Fumonisin B1 (Sigma), an inhibitor of sphingolipid synthesis, was added to cultures at final concentration of 10 mM beginning at 1 day in culture and renewed every other day for 4 days. Phosphatidyl-inositol-specific phospholipase C (Sigma) was added to cultures at a final concentration of 0.05 U/ml for 1 h. Surface immunocytochemistry of unfixed cells was performed as described above. RESULTS 465 HUMAN mAbs SUPPORT NEURITE EXTENSION mAbs bound to the cerebellum were similar to those observed using anti-glycolipid (27) and anti-ganglioside mAbs (36). sHIgM39 is an example of a non-reactive human IgM (Fig. 1H). Control slices (i.e. omission of the primary mAb or of both primary and secondary Abs) were always incubated in parallel to determine nonspecific background and autofluorescence. Both control conditions presented only faint nonspecific labeling of the IGL (not shown). Treatment of cerebellar slices with chloroform:methanol (2:1) prior to immunocytochemistry reduced the intensity of label by all human mAbs in Figure 1. These results suggested that the positive human mAbs reacted primarily with lipids or molecules associated with membrane lipids. Human mAbs Supported Neurite Extension Equal to that Supported by Laminin Human mAbs that bound to neurons in cerebellar slices were tested as substrates for cerebellar granule cell attachment and neurite extension (Table). Laminin was used as a positive control throughout the studies. Neurite extension assays were performed at least 3 times, utilizing triplicate samples. At 24 h following cell seeding, only sHIgM12 and sHIgM42 supported neurite extension and cell attachment similar to that supported by laminin (Fig. 2A–C). Granule cells seeded onto BSA-coated wells distributed evenly and elaborated short neurites (not shown, similar to Fig. 1F). When seeded onto sHIgM22 (Fig. 2E), which bound to WM and OLs, granule cells settled in clumps and extended short neurites. Granule cells seeded onto AKJR4, which bound intensely to the IGL in cerebellar slices, settled in clumps and adhered weakly (data not shown). Two mouse anti-gangliosides, R24 and A2B5, were tested as substrates for neurite extension. Both supported a significant population of neurites (Fig. 2D, G). ANOVA pairwise comparison on ranks indicated no difference between the neurite population supported by 10 mg/ml of A2B5, laminin, sHIgM12, or sHIgM42 to a significance of p , 0.05, while the neurite population supported by R24 was different. R24 induced more neurites per neuron than any other Ab tested. Laminin, sHIgM12, sHIgM42, and A2B5 supported 2 long dominant neurites per cell, whereas granule cells seeded onto R24 often elaborated 5 or more processes per cell (data not shown). The population of neurites supported by all other human mAbs TABLE Monoclonal Antibodies as Substrates for Neurite Extension Substratea BSA Laminin sHIgM 4 sHIgM 5 sHIgM 8 sHIgM 10 sHIgM 12 sHIgM 14 sHIgM 22 sHIgM 39 sHIgM 41 sHIgM 42 sHIgM 46 sHIgM 47 sHIgM 50 sHIgM 51 AKJR4 AKJR8 CB2bG8 A2B5 R24 Bindingb Attachment %c Extensiond — — WM GL — WM PC GL ML WM PC GL ML WM — WM WM GL ML WM PC WM WM ML ML GL GL — PC ML GL ML GL ML GL WM 57 100 4 6 27 4 84 50 32 23 39 86 40 34 40 20 10 12 88 91 84 2 1 2 2 2 2 1 2 2 2 2 1 2 2 2 2 2 2 2 1 1 Cytochemistrye — — — — soma and neurites — — — soma and neurites — — — — — — — soma and neurites soma and neurites a Prefixes. sHIgM: serum derived human mAb, AKJR: rheumatoid arthritis human B-cell clone, CB: cord blood human B-cell clone. b Binding. mAbs that bound to neuronal layers within a slice of unfixed mouse cerebellum were tested as substrates for granule cell neurite extension. sHIgM4, 5, 10, 39 and AKJR8 did not bind to slices and were included as negative controls. Abbreviations: GL: internal granular layer, ML: molecular layer, PC: Purkinje cell layer, WM: white matter. c Attachment. The average cell density of 3 trials compared to that supported by laminin. Laminin supported 221 6 17 cells/ mm2. d Extension. Whether a mAb supported a population of neurites 250 mm or longer within 24 h. e Immunocytochemistry. mAb binding to the surface of granule cells during the first 24 h in culture. J Neuropathol Exp Neurol, Vol 63, May, 2004 466 WARRINGTON ET AL was compared to laminin, sHIgM12, and sHIgM42— none reached a significance of p , 0.05. Substantial neurite extension was supported in wells coated with sHIgM12, sHIgM42 or laminin at concentrations down to 1 mg/ml. High concentrations (250 mg/ml) of sHIgM12 or sHIgM42, used as substrates or in solution, did not induce longer neurites, alter cell attachment, or otherwise appear toxic. Although sHIgM12, sHIgM42, and laminin supported equivalent neurite extension from purified granule cells, when used as substrates for aggregates of cerebellar cells, significant differences in the support of cell migration and spreading were observed. When aggregates were seeded onto laminin, widespread GFAP1 astrocytic spreading accompanied neurofilament1 neurite outgrowth and neuron soma migration over 48 h (Fig. 3A, Coomassie blue stain shown). In contrast, when aggregates of cerebellar cells were seeded onto sHIgM12 or sHIgM42, only robust neurofilament1 neurite extension was observed. GFAP1 astrocytes remained within the cell aggregates (Fig. 3B, C). sHIgM12 and sHIgM42 did not bind to the surface of astrocytes. Human mAbs Bound to Dissociated Neurons In Vitro When human mAbs were further screened for binding to the surface of granule cells during the first 24 h in culture, only sHIgM12 and sHIgM42 bound (Fig. 4A, B). At the same time in culture R24 and A2B5 also bound to the surface of granule cells (Fig. 3C, D). AKJR4 did not bind to the surface of granule neurons in culture at any time tested (Fig. 4E). AKJR4 may bind to a surface epitope expressed once granule neurons have migrated from the external granular layer (EGL) to the IGL. Tissue dissociation selects for viable neurons from the EGL (28) ← Fig. 2. Neurite extension supported by human mAbs. Bright field photomicroscopy demonstrated that certain mAbs, when presented as substrates, supported neurite extension from cerebellar granule cells. Granule cells were fixed 24 h after seeding and stained with Coomassie blue prior to imaging and neurite measurement. Qualitative examination of granule cell J Neuropathol Exp Neurol, Vol 63, May, 2004 cultures indicated that human mAbs sHIgM12 (B) and sHIgM42 (C) induced significant neurite extension, comparable to that supported by laminin (A). The anti-ganglioside mouse mAb A2B5 (D) also supported significant neurite extension. sHIgM22 (E) and sHIgM39 (F) did not support neurite extension. Most human mAbs tested did not induce neurite extension. Some human mAbs interfered with granule cell attachment (Table). Quantifying neurite extension supported by mAbs (G). The distribution frequency of granule cell neurite lengths 24 h post-seeding when cultured on substrate mAbs. Granule cells seeded on laminin, sHIgM12, sHIgM42, and A2B5 extended a large population of neurites longer than 300 mm within 24 h. Granule cells seeded on other human mAbs or BSA did not extend a population of neurites longer than 300 mm (Table). ANOVA of the total neurite length populations indicated that the neurites extended on laminin, sHIgM12, sHIgM42 and A2B5 were not different to a significance of p , 0.05. Figure 2G represents 1 of 4 independent neurite extension assays. In all trials, sHIgM12, sHIgM42, A2B5, and laminin supported statistically similar populations of neurites. Scale bar: 100 mm. HUMAN mAbs SUPPORT NEURITE EXTENSION 467 and the fragile cells of the IGL are presumably lost. The AKJR4 antigen is not recapitulated in culture. Epitopes Bound by sHIgM12 and sHIgM42 were Sensitive to Sialidase, but not to Treatment with FB1 or PIPLC To determine the nature of the epitopes bound by sHIgM12 and sHIgM42, granule cells were treated with fumonisin B1 (FB1) to downregulate sphingolipid expression, PIPLC to cleave GPI-linked molecules, or sialidase to cleave sialic acid. FB1 inhibits sphingolipid synthesis by blocking the acylation of sphingoid long chain bases. Treatment of hippocampal neurons with 50 mM FB1 for 3 days reduced sphingolipid synthesis to 10% of normal levels and eliminated the binding of anti-ganglioside Abs (37). Treatment of granule cells with as little as 10 mM FB1 for 5 days eliminated surface GD3 (Fig. 5C, D), but did not effect the binding of sHIgM12 or sHIgM42 (Fig. 5A, B). When granule cells were treated with PIPLC at a concentration of 0.05 U/ml for 1 h, the binding of antiThy-1, a GPI-linked neuronal protein (38), was eliminated (Fig. 5G, H), but did not alter the binding of sHIgM12 or sHIgM42 (Fig. 5E, F). In contrast, treatment of granule cells for 6 h with as little as 0.1 U/ml sialidase completely eliminated the binding of sHIgM12 and sHIgM42 (Fig. 5I, J) and greatly reduced the binding of A2B5 (Fig. 4K, L). As a control for non-specific protease activity the binding of HNK-1 (39) to granule cells was unaltered by sialidase treatment (data not shown). We concluded that the binding of sHIgM12 and sHIgM42 to the surface of granule cells was carbohydrate dependent. sHIgM12 and sHIgM42 did not Bind to Mature Oligodendrocytes or Colocalize with Myelin Proteins Fig. 3. When used as substrates, sHIgM12 and sHIgM42 supported neurite extension, but not astrocyte migration or spreading from aggregates of cerebellar cells. Dissociated cerebellar cells were allowed to aggregate as hanging drops in suspension. Aggregates were then added to mAb-coated wells. After 48 h, cultures were fixed and stained with Coomassie blue. Adjacent wells were double labeled with anti-GFAP and anti-neurofilament (data not shown). Laminin (A) supported significant astrocyte migration and spreading (arrow). In contrast, aggregates seeded sHIgM12 (B) and sHIgM42 (C) sHIgM12 and sHIgM42 bound to layers of the adult cerebellum that contained neurons, OLs, and myelin. To determine the OL component of this tissue label, OLs maintained for 2 weeks in vitro were surface immunolabeled. Neither sHIgM12 nor sHIgM42 bound to the surface of morphologically complex sulfatide-positive OLs (Fig. 6A–D). In contrast, sHIgM22 bound strongly to sulfatide-positive OLs (Fig. 6E, F). sHIgM22 is a human mAb that promoted remyelination and colocalized with anti-MOG in cerebellar slices (40). In contrast, sHIgM12 and sHIgM42 did not strictly colocalize with anti-MOG immunostaining in cerebellar slices (Fig. 1A, B, E). ← extended a penumbra of short neurites and a population of 300to 400-mm-long neurites. Astrocytes remained in the aggregates. sHIgM12 and sHIgM42 did not bind to astrocytes. Scale bar: 100 mm. J Neuropathol Exp Neurol, Vol 63, May, 2004 468 WARRINGTON ET AL Fig. 4. sHIgM12 and sHIgM42 bound to the surface of granule cells. Immunocytochemistry performed on unfixed granule cells at 24 h in culture. mAbs were used to label the surface of unfixed cells; bisbenzimide was used for nuclear localization. sHIgM12 (A), sHIgM42 (B), A2B5 (C), and R24 (D), mAbs that supported neurite extension, all bound to the surface of granule cell soma and neurites. In contrast, AKJR4 (E, corresponding phase contrast, F), did not support neurite extension and did not bind to the surface of granule cells. Scale bar: 50 mm. sHIgM12 and SHIgM42 Overrode the Neurite Extension Inhibition of CNS Myelin Since laminin has been shown to override the neurite extension inhibition of CNS myelin (41), sHIgM12 and sHIgM42 were tested for the ability to allow neurite extension on myelin (Fig. 7). Two mg spots of dried mouse spinal cord myelin were incubated with mAbs at 10 mg/ ml and then seeded with granule cells. At least 300 neurofilament1 neurites elaborated on the myelin spots were measured for each group. Neurites on untreated myelin averaged 13 6 2 mm while neurites on PDL averaged 31 6 2 mm. In contrast, neurites on myelin treated with laminin, sHIgM12 or sHIgM42 were significantly longer (29 J Neuropathol Exp Neurol, Vol 63, May, 2004 6 8, 32 6 3, and 29 6 2 mm, respectively). On myelin treated with the control mAb sHIgM39, neurite length was 17 6 1 mm. Statistical comparison of neurite length on untreated myelin was different than the neurite length on myelin pre-treated with laminin, sHIgM12, or sHIgM42 (p 5 ,0.001). DISCUSSION We have identified 2 human IgMs (sHIgM12 and sHIgM42) that when presented as substrates supported neurite extension from CNS neurons equal to that induced by laminin. Screening for biologically active human mAbs was performed in an antigen independent HUMAN mAbs SUPPORT NEURITE EXTENSION Fig. 5. Biochemical characterization of the granule neuron epitopes bound by sHIgM12 and sHIgM42. Cell surface epitopes bound by sHIgM12 (A) and sHIgM42 (B) were maintained following treatment of 50 mM FB1, whereas ganglioside epitopes bound by R24 were present on untreated cells (C), but lost in the presence of FB1 (D). Treatment with FB1 was for 5 days beginning 12 h after cell seeding. Surface epitopes bound by sHIgM12 (E) and sHIgM42 (F) were also maintained following treatment with PIPLC, whereas the GPI-linked protein Thy-1, present on untreated cells (G), was lost following PIPLC cleavage. Cultures were treated with 0.05 U/ml PIPLC for 1 h prior to immunocytochemistry. In contrast, surface epitopes bound by sHIgM12 (I) and sHIgM42 (J) were lost following treatment with sialidase. Epitopes bound by A2B5 (K) were 469 manner. Any human mAb that bound to neurons in an unfixed slice of cerebellum was tested as a substrate for neurite outgrowth. The binding pattern of sHIgM12 and sHIgM42 to layers of the cerebellum and to the surface of neurons, combined with epitope sensitivity to extraction, enzymatic cleavage, and metabolic inhibitors suggested that the mAb binding sites are carbohydrates associated with lipids. In support of this conclusion, 2 mouse anti-ganglioside mAbs (A2B5 and R24), chosen because each bound to the surface of neurons, also supported neurite extension. Both sHIgM12 and sHIgM42 also reduced the inhibitory effects of mouse CNS myelin on neurite extension, a characteristic shared by laminin (41). However, an outgrowth assay using cerebellar neuron/glia aggregates demonstrated differences between the mAbs and laminin. sHIgM12 and sHIgM42 did not support astrocyte spreading or migration from the aggregates, whereas laminin did. In aggregates cells have additional choices between suitable substrates (31). Granule cells prefer to extend short neurites toward other granule cells unless provided with a more attractive substrate. The aggregate culture studies suggest that sHIgM12 and sHIgM42, if provided as substrates in vivo, may drive axon outgrowth and control astrocyte migration, minimizing glial scar formation. When assayed for binding to dried myelin by ELISA, sHIgM12 and sHIgM42 bound very weakly compared to O4, anti-MOG, and sHIgM22 (unpublished data). Further studies will determine whether sHIgM12 and sHIgM42 bind to myelin directly and cover inhibitory epitopes or act similarly to laminin, offering an adhesive substrate that effectively competes with inhibitory epitopes. sHIgM12 and sHIgM42 are fundamentally different from the IN-1 and anti-Nogo mAbs that allow axon outgrowth in several models of CNS injury, presumably by blocking inhibitory molecules (2, 42). IN-1 bound to mature OLs and strictly colocalized in tissue sections with mAbs against MOG (43). In contrast, sHIgM12 and sHIgM42 did not bind to mature OLs or colocalize with mature myelin antigens. SCH antisera that promoted remyelination (15) and axon regeneration (17) bound to myelin and myelin proteins, but not to known myelin inhibitors (18). The anti-neuronal character of SCH antisera remains largely unexplored. However, SCH antisera with demonstrated ability to promote remyelination bound strongly to neuronal layers of the cerebellum (unpublished data). Following optic nerve lesion, retinal ganglion axon growth occurred within areas permeable to Abs (18), supporting the concept that axons can utilize ← also lost following sialidase digestion (L). Granule cells at 3 days in culture were treated with sialidase for 12 h prior to immunocytochemistry. Scale bar: 100 mm. J Neuropathol Exp Neurol, Vol 63, May, 2004 470 WARRINGTON ET AL Fig. 6. sHIgM12 and sHIgM42 did not bind to the surface of morphologically mature sulfatide-positive OLs. OL progenitors were maintained in a defined differentiation promoting media for 12 days and then double labeled prior to fixation with human mAbs and the anti-sulfatide mAb, O4. sHIgM12 (A) and sHIgM42 (C) did not label the surface of O4-positive OLs (B, D). However, sHIgM22, a human mAb previously reported to promote remyelination in models of demyelination, did bind to O4positive OLs (E, F). Scale bar: 50 mm. Abs as adhesive substrates or signaling molecules in vivo. Perhaps the anti-myelin Abs of SCH antisera participate in the promotion of remyelination, whereas antineuronal Abs of SCH antisera participate in promoting axon outgrowth. At least in vitro, sHIgM12, sHIgM42, laminin, and SCH antisera all provide sufficient incentive to allow neurites to ignore inhibitory myelin substrate molecules. J Neuropathol Exp Neurol, Vol 63, May, 2004 We hypothesize that sHIgM12 and sHIgM42 bind to neuronal gangliosides. Gangliosides can serve as receptors for ligand and Ab-mediated neuronal signaling (44). Anti-GM1 mAbs suppress neurite outgrowth (45). The axon outgrowth inhibition of MAG is mediated through GD1a and GT1b. Removal of the terminal sialic acid residues from neurons eliminated the inhibitory effect of MAG (46) and Ab-mediated cross-linking of GT1b or HUMAN mAbs SUPPORT NEURITE EXTENSION 471 Fig. 7. sHIgM12 and sHIgM42 overrode the neurite extension inhibition of CNS mouse myelin. (A) Bars represent the mean and SEM of 300 neurofilament1 neurites measured on myelin spots for each test group from triplicate wells 24 h after seeding on substrates of PDL, mouse myelin, or mouse myelin treated with mAbs. On myelin treated with sHIgM12 or sHIgM42, neurite extension was significantly greater than that measured on myelin alone or myelin treated with the control human mAb, sHIgM39. Neurite extension on myelin treated with sHIgM12, sHIgM42 or laminin was not different to a significance of p , 0.05 by ANOVA. Sample fields of granule cells labeled for neurofilament seeded on PDL (B), mouse myelin (C), and mouse myelin treated with sHIgM42 (D). GD1 mimics the effect of MAG (47, 48). Abs to GD3 or the GPI-linked protein, TAG-1, activated Lyn and induced similar alterations in protein-tyrosine phosphorylation. Removal of surface carbohydrates reduced the level of membrane GD3 and eliminated Ab-mediated signaling (49, 50). Anti-ganglioside Abs, especially pentameric IgMs, likely act as ligands by clustering plasma membrane lipid rafts containing signal transduction complexes. Laminin would appear to be an ideal candidate molecule to encourage axon re-growth if delivered to the site of CNS injury. Laminin, normally absent in the adult CNS, is upregulated following injury (51) and is likely an aspect of endogenous CNS repair (52). The effectiveness of laminin for nerve regeneration has been demonstrated (53). However, laminin-1 is a large molecule, difficult to synthesize and potentially tumorigenic (54). Therefore, smaller functional domains of laminin or molecules that mimic laminin have been sought. The g1 chain of laminin or a tripeptide portion retained the ability to promote neurite extension and allowed axon extension in the presence of CNS myelin (55, 56). Both molecules supported neurite extension in vitro within a narrow concentration range and appear to be toxic to neurons at higher concentrations. The accumulation of g1 chain in the brain was associated with Alzheimer disease and Down syndrome (57). If used in vivo, controlling the local concentration of these molecules will be critical. Anti-Thy-1 Abs induced neurite extension when used as a substrate (58). However, anti-Thy-1 Abs were systemically pathogenic to the nephritic glomeruli and vascular endothelium and are routinely used to initiate a model of nephritis (59, 60). sHIgM12 or sHIgM42 have in a real sense been prescreened for in vivo toxicity in the patients that synthesize the molecules. Despite having carried high levels of these mAbs for many years neither individual presents with Ab-mediated dysfunction. Five hundred mg of J Neuropathol Exp Neurol, Vol 63, May, 2004 472 WARRINGTON ET AL sHIgM12 and sHIgM42 were injected into chronically TMEV-infected mice, where the blood brain barrier is open, with no deleterious effects. Neurons maintained in culture for weeks in the presence of 250 mg/ml of sHIgM12 or sHIgM42 were unaffected. Abs synthesized following CNS injury may participate in repair, as modeled by the SCH pre-immunization studies and equivalent repair can be obtained by the exogenous addition of mAbs. Human mAbs have a number of advantages as therapeutic molecules. They are likely to be minimally antigenic when administered systemically—Abs are normally present in the circulation—and have a limited life span in the recipient. In contrast, administering an antigen to induce an individual to synthesize their own reparative Abs may result in unpredictable immune reactions across the population. Recombinant human mAbs can be synthesized free of infectious agents and easily molecularly altered (40). When administered systemically, mAbs that promoted remyelination localized to areas of demyelination (61) and SCH antisera localized to lesioned areas of the optic nerve (18). Thus, therapeutic human mAbs administered systemically may localize to sites of injury. Human mAbs may be one component of a combinatorial reparative strategy (utilizing cell transplantation, neurotrophic factors, or degradative enzymes) delivered to the lesion site. mAbs can be incorporated into biodegradable polymer matrices that when implanted can provide a local supply (62) or used to line guidance channels of polymer implants, which may assist in promoting directional axonal growth and inhibit glial infiltration. Human mAb-based therapeutics may offer a specificity of binding and, potentially, of action not possible with other reagents. Future studies will determine whether human mAbs that promote neurite extension have utility in CNS repair. ACKNOWLEDGMENTS We thank Mr. and Mrs. Eugene Applebaum for their generous support and Eileen M. Kennedy for her editorial savvy. REFERENCES 1. Caroni P, Schwab ME. Antibody against myelin-associated inhibitor of neurite growth neutralizes nonpermissive substrate properties of CNS white matter. Neuron 1988;1:85–96 2. Schnell L, Schwab ME. Axonal regeneration in the rat spinal cord produced by an antibody against myelin-associated neurite growth inhibitors. Nature 1990;343:269–72 3. McKerracher L, David S, Jackson DL, Kottis V, Dunn RJ, Braun PE. Identification of myelin-associated glycoprotein as a major myelinderived inhibitor of neurite outgrowth. Neuron 1994;13:805–11 4. Fitch MT, Silver J. Beyond the glial scar: Cellular and molecular mechanisms by which glial cells contribute to CNS regenerative failure. In: Tuszynski MH, Kordower JH, eds. CNS regeneration: Basic sciences and clinical advances. New York: Academic Press, 1999:55–62 5. Niederost BP, Zimmermann DR, Schwab ME, Bandtlow CE. Bovine CNS myelin contains neurite growth-inhibitory activity associated with chondroitin sulfate proteoglycans. J Neurosci 1999;19: 8979–89 J Neuropathol Exp Neurol, Vol 63, May, 2004 6. Fournier AE, Strittmatter SM. Repulsive factors and axon regeneration in the CNS. Curr Opin Neurobiol 2001;11:89–94 7. Richardson PM, McGuinness UM, Aguayo AJ. Axons from CNS neurons regenerate into PNS grafts. Nature 1980;284:264–65 8. Aguayo AJ, David S, Bray M. Influences of the glial environment on the elongation of axons after injury: Transplantation studies in adult rodents. J Exp Biol 1981;95:231–40 9. Ramon y Cajal S. Degeneration and regeneration of the nervous system. 1928. Translated by May RM, New York: Oxford UP, 1991 10. Bunge RP, Puckett WR, Becerra JL, Marcillo A, Quencer RM. Observations on the pathology of human spinal cord injury. A review and classification of 22 new cases with details from a case of chronic cord compression with extensive focal demyelination. Adv Neurol 1993;59:75–89 11. Tseng GF, Prince DA. Structural and functional alterations in rat corticospinal neurons after axotomy. J Neurophysiol 1996;75:248–67 12. Davies SJ, Fitch MT, Memberg SP, Hall AK, Raisman G, Silver J. Regeneration of adult axons in white matter tracts of the central nervous system. Nature 1997;390:680–83 13. Davies SJ, Goucher DR, Doller C, Silver J. Robust regeneration of adult sensory axons in degenerating white matter of the adult rat spinal cord. J Neurosci 1999;19:5810–22 14. Lang W, Rodriguez M, Lennon VA, Lampert PW. Demyelination and remyelination in murine viral encephalomyelitis. Ann NY Acad Sci 1984;436:98–102 15. Rodriguez M, Lennon VA, Benveniste EN, Merril JE. Remyelination by oligodendrocytes stimulated by antiserum to spinal cord. J Neuropathol Exp Neurol 1987;46:84–95 16. Rodriguez M, Lennon VA. Immunoglobulins promote remyelination in the central nervous system. Ann Neurol 1990;27:12–17 17. Huang DW, McKerracher L, Braun PE, David S. A therapeutic vaccine approach to stimulate axon regeneration in the adult mammalian spinal cord. Neuron 1999;24:639–47 18. Ellezam E, Bertrand J, Dergham P, McKerracher L. Vaccination stimulates retinal ganglion cell regeneration in the adult optic nerve. Neurobio Dis 2003;12:1–10 19. Asakura K, Miller DJ, Pease LR, Rodriguez M. Targeting of IgM kappa antibodies to oligodendrocytes promotes CNS remyelination. J Neurosci 1998;18:7700–708 20. Warrington AE, Asakura K, Bieber AJ, et al. Human monoclonal antibodies reactive to oligodendrocytes promote remyelination in a model of multiple sclerosis. Proc Natl Acad Sci USA 2000;97: 6820–25 21. Paz Soldán MM, Warrington AE, Bieber AJ, et al. Remyelination promoting antibodies activate distinct Ca21 influx pathways in astrocytes and oligodendrocytes: Relationship to the mechanism of myelin repair. Mol Cell Neurosci 2003;22:14–24 22. Eisenbarth GS, Walsh FS, Nirenberg M. Monoclonal antibody to a plasma membrane antigen of neurons. Proc Natl Acad Sci USA 1979;76:4913–17 23. Fredman PJ, Magnani JL, Nirenberg M, Ginsburg V. Monoclonal antibody A2B5 reacts with many gangliosides in neuronal tissue. Arch Biochem Biophys 1984;233:661–66 24. Pukel CS, Lloyd KO, Travassos LR, Dippold WR, Oettgen HF, Old LJ. GD3, a prominent ganglioside of human melanoma. Detection and characterization by mouse monoclonal antibody. J Exp Med 1982;155:1133–47 25. Sommer I, Schachner M. Monoclonal antibodies (O1 to O4) to oligodendrocyte cell surfaces: An immunocytological study in the central nervous system. Dev Biol 1981;83:311–27 26. Bansal R, Warrington AE, Gard AL, Ranscht B, Pfeiffer SE. Multiple and novel specificities of monoclonal antibodies O1, O4, and R-mAb used in the analysis of oligodendrocyte development. J Neurosci Res 1989;24:548–57 473 HUMAN mAbs SUPPORT NEURITE EXTENSION 27. Warrington AE, Pfeiffer SE. Proliferation and differentiation of O41 oligodendrocytes in postnatal rat cerebellum: Analysis in unfixed tissue slices using anti-glycolipid antibodies. J Neurosci Res 1992;33:338–53 28. Raetzman LT, Siegel RE. Immature granule neurons from cerebella of different ages exhibit distinct developmental potentials. J Neurobiol 1999;38:559–70 29. Gault LM, Siegel RE. Expression of the GABAA receptor delta subunit is selectively modulated by depolarization in cultured rat cerebellar granule neurons. J Neurosci 1997;17:2391–99 30. Armstrong RC. Isolation and characterization of immature oligodendrocyte lineage cells. Methods 1998;16:282–92 31. Jacob J, Haspel J, Kane-Goldsmith N, Grumet M. L1 mediated homophilic binding and neurite outgrowth are modulated by alternative splicing of exon 2. J Neurobiol 2002;51:177–89 32. Lemmon V, Farr KL, Lagenaur C. L1-mediated axon outgrowth occurs via a homophilic binding mechanism. Neuron 1989;2:1597–603 33. Norton WT, Poduslo SE. Myelination in rat brain: Method of myelin isolation. J Neurochem 1973;21:749–57 34. Bieber AJ, Warrington A, Asakura K, et al. Human antibodies accelerate the rate of remyelination following lysolecithin-induced demyelination in mice. Glia 2002;37:241–49 35. Schwartz A, Futerman AH. The localization of gangliosides in neurons of the central nervous system: The use of anti-ganglioside antibodies. Biochem Biophys Acta 1996;1286:247–67 36. Molander M, Berthold C-H, Persson H, Fredman P. Imunostaining of ganglioside GD1b, GD3 and GM1 in rat cerebellum: Cellular layer and cell type specific associations. J Neurosci Res 2000;60: 531–42 37. Schwartz A, Rapapport E, Hirschberg K, Futerman AH. A regulatory role for sphingolipids in neuronal growth. J Biol Chem 1995; 270:10990–98 38. Doherty P, Singh A, Rimon G, Bolsover SR, Walsh FS. Thy-1 antibody-triggered neurite outgrowth requires an influx of calcium into neurons via N- and L-type calcium channels. J Cell Biol 1993; 122:181–89 39. Jungalwala FB. Expression and biological functions of sulfoglucuronyl glycolipids SGGLs in the nervous system—A review. Neurochem Res 1994;19:945–57 40. Mitsunaga Y, Ciric B, Van Keulen V, et al. Direct evidence that a human antibody derived from patient serum can promote myelin repair in a mouse model of chronic-progressive demyelinating disease. FASEBJ 2002;16:1325–27 41. David S, Braun PE, Jackson DL, Kottis V, McKerracher L. Laminin overrides the inhibitory effects of peripheral nervous system and central nervous system myelin-derived inhibitors of neurite growth. J Neurosci Res 1995;42:594–602 42. Bregman BS, Kunkel-Bagden E, Schnell L, Dai HN, Gao D, Schwab ME. Recovery from spinal cord injury mediated by antibodies to neurite growth inhibitors. Nature 1995;378:498–501 43. Rubin BP, Dusart I, Schwab ME. A monoclonal antibody (IN-1) which neutralizes neurite growth inhibitory proteins in the rat CNS recognizes antigens localized in CNS myelin. J Neurocytol 1994; 23:209–17 44. Ledeen RW, Wu G. Ganglioside function in calcium homeostasis and signaling. Neurochem Res 2002;27:637–47 45. Spirman N, Sela BA, Schwartz M. Antiganglioside antibodies inhibit neuritic outgrowth from regenerating goldfish retinal explants. J Neurochem 1982;39:874–77 46. Yang LJ-S, Zeller CB, Shaper NL, et al. Gangliosides are neuronal ligands for myelin-associated glycoprotein. Proc Natl Acad Sci USA 1996;93:814–18 47. Vinson M, Strijbos PJ, Rowles A, et al. Myelin-associated glycoprotein interacts with ganglioside GT1b. A mechanism for neurite outgrowth inhibition. J Biol Chem 2001;276:20280–85 48. Vyas AA, Patel HV, Fromholt SE, et al. Gangliosides are functional nerve cell ligands for myelin-associated glycoprotein (MAG), an inhibitor of nerve regeneration. Proc Natl Acad Sci USA 2002;99: 8412–17 49. Kasahara K, Watanabe Y, Yamamoto T, Sanai Y. Association of Src family tyrosine kinase with ganglioside GD3 in rat brain. J Biol Chem 1997;272:29947–53 50. Kasahara K, Watanabe K, Takeuchi K, et al. Involvement of gangliosides in glycosylphosphatidylinositol-anchored neuronal cell adhesion molecule TAG-1 signaling in lipid rafts. J Biol Chem 2000;275:34701–9 51. McLoon SC, McLoon LK, Palm SL, Furcht LT. Transient expression of laminin in the optic nerve of the developing rat. J Neurosci 1988;8:1981–90 52. Liesi P. Laminin-immunoreactive glia distinguish regenerative adult CNS systems from non-regenerative ones. EMBOJ 1985;4:2505–11 53. Manthorpe M, Engvall E, Ruoslahti E, Longo FM, Davis GE, Varon S. Laminin promotes neuritic regeneration from cultured peripheral and central neurons. J Cell Biol 1983;97:1882–90 54. Timple R, Rhode H, Robey PG, Rennaed SI, Foidart JM, Martin GR. Laminin—A glycoprotein from basement membranes. J Biol Chem 1979;254:9933–37 55. Liesi P, Risteli L. Glial cells of the mammalian brain produce a varient form of laminin. Exp Neurol 1989;105:86–92 56. Liebkind R, Laatikainen T, Liesi P. Is the soluble KDI domain of g1 laminin a regeneration factor for the mammalian central nervous system? J Neurosci Res 2003;73:637–43 57. Murtomaki S, Risteli J, Risteli L, Koivisto UM, Johansson S, Liesi P. Laminin and its neurite outgrowth-promoting domain in the brain in Alzheimer’s disease and Down’s syndrome patients. J Neurosci Res 1992;32:261–73 58. Leifer D, Lipton SA, Barnstable CJ, Masland RH. Monoclonal antibody to Thy-1 enhances regeneration of processes by rat retinal ganglion cells in culture. Science 1984;224:303–6 59. Nakayama H, Oite T, Kawachi H, et al. Comparative nephritogenicity of two monoclonal antibodies that recognize different epitopes of rat Thy-1.1 molecule. Nephron 1998;78:453–63 60. Jefferson JA, Johnson RJ. Experimental mesangial proliferative glomerulonephritis (the anti-Thy-1.1 model). J Nephrol 1999;12:297–307 61. Hunter SF, Miller DJ, Rodriguez M. Monoclonal remyelinationpromoting natural autoantibody SCH 94.03: Pharmacokinetics and in vivo targets within demyelinated spinal cord in a mouse model of multiple sclerosis. J Neurol Sci 1997;150:103–13 62. Friedman JA, Windebank AJ, Moore MJ, Spinner RJ, Currier BL, Yaszemski MJ. Biodegradable polymer grafts for surgical repair of the injured spinal cord. Neurosurg 2002;51:742–52 Received October 21, 2003 Revision received January 26, 2004 Accepted February 3, 2004 J Neuropathol Exp Neurol, Vol 63, May, 2004