Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
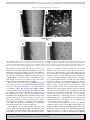
+ MODEL ARTICLE IN PRESS Research in Microbiology xx (2010) 1e8 www.elsevier.com/locate/resmic Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea Kailing Zhu a,1, Hongmiao Pan a,1, Jinhua Li b, Kui Yu-Zhang c, Sheng-Da Zhang a, Wen-Yan Zhang a, Ke Zhou a, Haidong Yue a, Yongxin Pan b,e, Tian Xiao a,e,**, Long-Fei Wud,e,* a Key Laboratory of Marine Ecology & Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China b Key Laboratory of the Earth’s Deep Interior, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing 100029, China c Laboratoire de Microscopies et d’Etude de Nanostructures, Université de Reims, 51687 Reims, France d Laboratoire de Chimie Bactérienne, Institut de Biologie Structurale et Microbiologie, UPR9043-CNRS, Marseille, France e Laboratoire International Associé de la Bio-Minéralization et Nano-Structures, CNRS-Marseille, 13009 Marseille, France Received 4 December 2009; accepted 3 February 2010 Abstract Magnetotactic bacteria (MTB) are ubiquitous in aquatic habitats. Because of their fastidious requirements for growth conditions, only very few axenic MTB cultures have been obtained worldwide. In this study, we report a novel marine magnetotactic spirillum axenic culture, designated as QH-2, isolated from the China Sea. It was able to grow in semi-solid or liquid chemically defined medium. The cells were amphitrichously flagellated and contained one single magnetosome chain with an average number of 16 magnetosomes per cell. Phosphate and lipid granules were also observed in the cells. Both rock magnetism and energy-dispersive X-ray spectroscopy characterizations indicated that the magnetosomes in QH-2 were single-domain magnetites (Fe3O4). QH-2 cells swam mostly in a straight line at a velocity of 20e50 mm/s and occasionally changed to a helical motion. Unlike other magnetotactic spirilla, QH-2 cells responded to light illumination. As a consequence of illumination, the cells changed the direction in which they swam from parallel to the magnetic field to antiparallel. This response appears to be similar to the effect of an increase in [O2]. Analysis of the QH-2 16S rRNA sequence showed that it had greater than 11% sequence divergence from freshwater magnetotactic spirilla. Thus, the marine QH-2 strain seems to be both phylogenetically and magnetotactically distinct from the freshwater Magnetospirillum spp. studied previously. Ó 2010 Elsevier Masson SAS. All rights reserved. Keywords: Magnetotactic bacteria; Response to light; Pure culture; Magnetosome; Phylogenetic analysis 1. Introduction Magnetotactic bacteria (MTB) are a heterogeneous group of aquatic microorganisms which share the ability to orient themselves along magnetic field lines. The cell reaction to the * Corresponding author. Laboratoire de Chimie Bactérienne, CNRS-Marseille, 13009 Marseille, France. Tel.: þ33 4 91164157; fax: þ33 4 91718914. ** Corresponding author. Key Laboratory of Marine Ecology & Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China. E-mail addresses: [email protected] (T. Xiao), [email protected] (L.-F. Wu). 1 These authors contributed equally to this work. magnetic field is due to the presence of magnetosomes, intracellular membrane-bound crystals of iron mineral which consist of either magnetite (Fe3O4) or greigite (Fe3S4) within the single domain (SD) size range (30e120 nm) (Bazylinski and Frankel, 2004). Because of their high abundance and their remarkable capacity for accumulating and precipitating iron minerals, MTB are assumed to have great impact on the biogeochemical cycling in natural sediments and are considered as an ideal model for understanding the mechanism of biomineralization. MTB comprise a variety of morphological types, such as coccoid, vibriod, rod-shaped, spiral-shaped, and multicellular aggregates. They are distributed worldwide and most of them are found at, or just below, the oxiceanoxic 0923-2508/$ - see front matter Ó 2010 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.resmic.2010.02.003 Please cite this article in press as: Zhu, K., et al., Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea, Research in Microbiology (2010), doi:10.1016/j.resmic.2010.02.003 ARTICLE IN PRESS 2 K. Zhu et al. / Research in Microbiology xx (2010) 1e8 transition zone (OATZ) or redoxocline in aquatic habitats (Bazylinski and Frankel, 2004). Considering their ubiquitous distribution and remarkable diversity with respect to physiology and biomineralization, axenic cultures of MTB are needed for a comprehensive understanding of the mechanisms of magnetosome biogenesis, magnetotaxis as well as of MTB ecological function. As a new type of bioresource, magnetotactic bacteria have also attracted much attention for their potential use in biotechnology, biomediation, and geobiology (Bazylinski and Frankel, 2004; Lang and Schüler, 2006). However, only very few strains are available in pure culture because of their fastidious growth requirements and strong metabolic diversity (Flies et al., 2005). Most cultivated MTB are affiliated with Alphaproteobacteria (Flies et al., 2005) except for the magnetotactic sulfatereducing bacterium Desulfovibrio magneticus (Sakaguchi et al., 2002). Axenic marine cultures include two magnetic vibrios (MV-1, Bazylinski et al., 1988; MV-2, DeLong et al., 1993), one magnetic spirillum (MMS-1, formerly known as MV-4, Meldrum et al., 1993) and three magnetic cocci (MC-1, Frankel et al., 1997; MC-2, Devouard et al., 1998; MO-1, Lefèvre et al., 2009). Here we report a novel marine magnetic spirillum axenic culture, designated strain QH-2, isolated from an intertidal zone of the China Sea. We will describe the growth features, cell structure, magnetic properties and novel motility characteristics. 2.4 ml of 0.8 M NaHCO3; 0.05 g sodium thioglycolate; and 900 ml filtered seawater collected from the pond through a 0.45 mm filter membrane. Artificial seawater and 0.1 g/L peptone were used instead of natural seawater and sediment extract in order to make the chemically defined medium, called QH-C medium. The pH value of the growth medium was adjusted to 7.6e7.8. For semi-solid medium, 0.2e0.5 g agar was added to 1 L of growth medium. After pH adjustment, the medium was autoclaved at 120 C for 20 min. Cultures were incubated at room temperature (between 22 and 26 C). 2.2. Optical and electron microscopy observations The swimming behavior of magnetotactic bacteria was analyzed by the ‘hanging drop’ method (Schüler, 2002) using a microscope (OLYMPUS BX51) connected to a ChargeCoupled Device (CCD, OLYMPUS DP71). Morphological examination of the cells was performed with fluorescence microscopy after staining with 1% acridine orange (AO). Nile red staining for lipid storage granules was performed according to Greenspan et al. (1985). Fresh cells were deposited on formvar carbon-coated copper grids, either directly or after treatment with 0.1% uranyl acetate. The grids were dried in air. TEM observations were made using a Zeiss EM9 microscope at 80 kV. The size and shape factors of magnetosomes were estimated using (length þ width)/2 and width/length, respectively. The chemical composition of magnetosomes was studied by EDXSeTEM. 2. Materials and methods 2.3. Sequence analysis of the 16S rRNA gene 2.1. Isolation and cultivation of the QH-2 strain The samples were collected from a seawater pond located at Huiquan Bay in the city of Qingdao, China. The characteristics of the pond were previously described (Pan et al., 2008). The sediments together with interface water, with a ratio of 1:2, were collected and stored in 1-L glass bottles. Magnetotactic bacteria were enriched by attaching the south pole of permanent magnets (0.37 mT) outside the bottles placed at the water/sediment interface. After 20e30 min, cells accumulating as dark spots underneath the magnets were removed with a Pasteur pipette and saved as magnetically collected samples. These samples were further magnetically purified in Pasteur pipettes according to the racetrack purification method (Wolfe et al., 1987), and inoculated into 5 ml plastic tubes that were generally filled up to 4/5th of their volume with various media and sealed with parafilm and incubated at 22e26 C in dim light. Bacteria grew and formed a sharp band or zone after several days of incubation. The optimal growth medium for QH-2 (QH medium) was modified from that used for the Magnetococcus sp. MC-1 (Frankel et al., 1997). It contained 100 ml extract solution of seawater sediments (modified from the DSMZ, Medium 12: Soil Extract Medium); 5 ml modified Wolfe’s mineral solution; 0.5 ml vitamin solution; 2.0 ml of 0.01 M ferric quinate; 1.0 g NH4Cl; 2.5 g Na2S2O3$5H2O; 1.5 ml of 0.5 M potassium phosphate buffer, pH 7.6; 1.3 g sodium lactate (50e60%); The 16S rRNA gene of the QH-2 was amplified between positions 27 and 1492 (Escherichia coli 16S rRNA gene sequence numbers), using primers 27F (50 -AGA GTY TGA TCC TGG CTC AG-30 ) and 1492R (50 -GGT TAC CTI ‘GTI’ ACG ACT T-30 ) by polymerase chain reaction (PCR) carried out with the following cycle: an initial denaturing step at 94 C for 5 min, followed by 25 cycles of 1 min at 94 C, 45 s at 50 C and 1 min at 72 C, and a final extension step of 10 min at 72 C. Then the PCR products were sequenced directly by Sinogenomax Company in Beijing. The sequences of the 16S rDNA gene were first analyzed using Advanced BLAST search program on the NCBI Website (http://www.ncbi.nlm.nih.gov/BLAST/). The related sequences were preliminarily aligned with the default setting of CLUSTALX (1.83). Phylogenetic analysis was performed with the default setting of MEGA 4 using the neighbor-joining method. Similarity was calculated using the BioEdit program. The newly determined sequence is available from GenBank under accession number EU675666. 2.4. Magnetic measurements For magnetic measurements, about 1010 QH-2 cells were collected by centrifugation from cultures. The centrifuged cells were washed once with distilled water, and then placed in a non-magnetic gelatin capsule. To avoid possible oxidization, Please cite this article in press as: Zhu, K., et al., Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea, Research in Microbiology (2010), doi:10.1016/j.resmic.2010.02.003 ARTICLE IN PRESS K. Zhu et al. / Research in Microbiology xx (2010) 1e8 3 the capsule was protected with N2 and stored at 80 C until magnetic measurements were made. Room-temperature magnetic experiments were performed on a vibrating sample magnetometer Model 3900 (Princeton Measurements Corporation, sensitivity is 5.0 1010 A m2). A hysteresis loop was measured between þ500 and 500 mT with an average time of 400 ms. Saturation magnetization (Ms), saturation remanence (Mrs) and coercivity (Bc) were determined after correction for paramagnetic phases. The saturation isothermal remanent magnetization (SIRM) was demagnetized in a backfield to obtain remanence coercivity (Bcr). First-order reversal curves (FORCs) were measured following the protocol as described by Roberts et al. (2000). A FORC diagram was calculated using FORCine version 1.05 with a smoothing factor of 3 (Harrison and Feinberg, 2008). This diagram showed a microcoercivity field along the horizontal axis (Hc) and a magnetostatic interaction field along the vertical axis (Hb) (Chen et al., 2007). Low-temperature magnetic experiments were performed on a Quantum Design Magnetic Property Measurement System (MPMS XP-5, sensitivity is 5.0 1010 A m2). Saturation remanence acquired in a 2.5-T field at 5 K (hereafter termed SIRM5K-2.5T) was demagnetized by warming from 5 to 300 K after two different pre-treatments. The first was to cool the cell sample from 300 down to 5 K in a zero field (ZFC), whereas the second was to cool the cell sample from 300 to 5 K in a 2.5-T field (FC). The Verwey transition temperature (Tv) was defined as the temperature for the maximum of the first-order derivative of dM/dT of the FC curve. The d ratio (dFC/dZFC) was calculated according to Moskowitz et al. (1993), which reflects the difference in remanence losses between the FC and ZFC when warming through the Verwey transition. 3. Results and discussion 3.1. Isolation and cultivation of the QH-2 strain Magnetotactic bacteria collected from the intertidal zone sediments displayed various cell shapes including cocci, rods, vibrios and spirilla, although the coccoid morphotype was always the dominant one. After magnetic racetrack purification, MTB were inoculated in various semi-solid media in 5 ml plastic culture tubes. More than two weeks, after the first inoculation, a sharp bacterial band or zone was formed at the oxiceanoxic interface in some of the tubes. Cells collected afterwards from the band in QH-semi-solid media (see ‘‘Materials and methods’’) were examined under an optical microscope. Unexpectedly, the dominant magnetotactic cocci in the collected samples did not grow, but magnetotactic spirilla together with non-magnetic rods and cocci were found in the first culture. To obtain a pure culture of magnetotactic spirilla, the mixed culture was repeatedly subjected to a new cycle of racetrack purification and re-inoculated in fresh semisolid media. After more than 20 subsequent cycles of racetrack purification and re-inoculation, we finally obtained a pure culture of magnetotactic spirillum, designated strain QH-2, as proven by 16S rRNA gene sequence analysis and genomic Fig. 1. Cellular characteristics of QH-2 cells. QH-2 cells were stained with acridine orange (A) or uracyl acetate (B and C) and inspected using fluorescence (A) or electron microscopy (B and C). The scale bars are 4 mm for A and 1 mm for B and C. Panel D shows the growth curve of QH-2 cultures incubated in QH-C media (see Materials and methods) at 26 C. The values are means plus standard deviation calculated from 3 sets of independent experiments. Please cite this article in press as: Zhu, K., et al., Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea, Research in Microbiology (2010), doi:10.1016/j.resmic.2010.02.003 ARTICLE IN PRESS 4 K. Zhu et al. / Research in Microbiology xx (2010) 1e8 sequencing (see below). Both the fluorescence microscope (Fig. 1A) and TEM observations (Fig. 1B) showed that the cells are vibrioid-to-helicoid in morphology, with a mean width of 0.8 0.2 mm and variable lengths ranging from 1 to 3 mm, with a mean length of 2.0 0.4 mm. They were amphitrichously flagellated with a single flagellum at each pole (Fig. 1C). Various marine culture media were analyzed for the growth of the QH-2 culture, which seemed to be heterotrophic under the conditions used, as growth of QH-2 required both the sediment extract for the QH medium and low amounts of peptone for QH-C medium. Nonetheless, we could not exclude capacity for autotrophic growth if appropriate conditions were provided. Up to now, QH and QH-C are the optimal media for QH-2 cell growth. When grown statically within the QH-C medium containing 0.05% agar at 26 C, the generation time of QH-2 was about 16 h (Fig. 1D). 3.2. Phylogenetic lineage Sequences of the 16S rRNA gene measured with about 20 independent QH-2 cultures showed 99.9% identity. Chromosomal DNA was prepared from the QH-2 culture. Genomic sequencing indicated that the QH-2 culture consists of only one species of bacteria (data not shown), confirming the pure culture of the QH-2 strain. Phylogenetic analysis showed that QH-2 was affiliated with Alphaproteobacteria and its 16S rRNA sequence was more than 11.7% divergent from all freshwater magnetotactic spirilla including Magnetospirillum magnetotacticum MS-1, Magnetospirillum magneticum AMB-1, Magnetospirillum gryphiswaldense MSR-1, and Magnetospirillum spp. MGT-1 and WM-1 (Blakemore et al., 1979; Li et al., 2007; Matsunaga et al., 1991; Okamura et al., 2003; Schüler and Koehler, 1992). Moreover, QH-2 was 88.9%, 82.4% and 84.7% identical to that of marine magnetotactic vibrio MV-1, Magnetococcus sp. MC-1 and magnetotactic ovoidal strain MO-1, respectively (Fig. 2). The closest relative was found to be the marine magnetic spirillum MMS-1 (97.2% identity) (Fig. 2), which was isolated from mud and water from School Street Marsh, Woods Hole, MA, U.S.A. (Meldrum et al., 1993). Therefore QH-2 together with MMS-1 may represent a novel phylogenetic lineage that is distinct from other axenic magnetotactic bacterial cultures. However, it is worth noting that although the 16S rRNA gene sequence of QH-2 showed high identity with that of MMS-1, they might not belong to the same species. In fact, even with 97.7% identity in their 16S rRNA sequences, the freshwater magnetospirillum strains AMB-1 (from Japan) and WM-1 (from China) actually belong to different species (Li et al., 2007). 3.3. Intracellular structures of QH-2 TEM observations and statistical analyses showed that each QH-2 cell contained 7e28 magnetosomes, with an average number of 16 5 per cell (Fig. 3A and B), which is similar to the number of magnetosomes found in MMS-1 (Meldrum et al., 1993). The magnetosome crystals of QH-2 cells were identified as iron oxide (Fe3O4) by both EDXS and electron diffraction (data not shown). The magnetosomes in a chain displayed a relatively large size distribution compared to other magnetospirilla (Fig. 3B and C). The growth conditions of QH-2 cultures might not be optimal for the formation of magnetosomes. The magnetite magnetosomes had an average width of 58 20 nm, an average length of 81 23 nm and a shape factor of 0.71 0.11. Similar to other magnetotactic bacteria, the distribution of the QH-2 crystals was asymmetric, with a cut-off toward larger size. Meanwhile, the length of magnetosomes in QH-2 cells was longer than that of MMS-1 whereas their width was roughly the same. The shape factor analysis of magnetosome crystals of QH-2 cells showed the distribution bounded by one, with a maximum around 0.7 (Fig. 3C), which is similar to those of marine magnetotactic bacteria MV-1 (shape factor was 0.65) (Devouard et al., 1998). In addition to the formation of magnetosomes (Fig. 3A, white arrow), one or two dark granules (Fig. 3A, labeled ‘P’) and a number of white globules (Fig. 3A, labeled ‘L’) were observed in QH-2 cells by TEM. EDXS analysis revealed that the dark granules were rich in phosphorus and oxygen, but the white globules could not be identified (data not shown). Similar intracellular aggregates have been identified as lipid Fig. 2. Neighbor-joining tree. The sequence determined in this study is written in bold. GenBank accession numbers of the sequences used are indicated in parentheses. Bootstrap proportions are shown. Scale bar, 0.02 substitutions per nucleotide position. Please cite this article in press as: Zhu, K., et al., Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea, Research in Microbiology (2010), doi:10.1016/j.resmic.2010.02.003 ARTICLE IN PRESS K. Zhu et al. / Research in Microbiology xx (2010) 1e8 5 Fig. 3. Magnetosome and granule characteristics of QH-2 cells. Panel A shows transmission electron microscopic micrographs of QH-2 cells and panel B shows a typical magnetosome chain inside cells. Magnetosome crystals (white arrow in panel A), phosphorouseoxygen-rich granules (‘P’ in panel A) and lipid storage globules (‘L’ in panel A) are indicated. Size (panel C1) and shape factor (panel C2) distributions were obtained by analysis of 281 crystals of QH-2 cells at stationary growth phase. Panels D1 and D2 are typical images of QH-2 cells inspected under an optical microscope as Nomarski contrast and fluorescence, respectively. D2 shows the same cells as in D1 after Nile red staining. Lipid storage granules are shown as white spots. Scale bars ¼ 1 mm for panel A, 100 nm for panel B and 4 mm for panel D. storage granules in MTB (Lefèvre et al., 2009; Silva et al., 2008). An efficient identification assay for the lipid storage granules is Nile red staining as described by Greenspan et al. (1985). Indeed, the white globules were specifically stained as red spots in QH-2 cells (Fig. 3, Panel D2, represented by white color), which confirmed their authenticity as lipid storage granules. As previously reported for the MO-1 strain, some magnetosome chains bent along the curvature of the lipid storage globules in the QH-2 cells (Fig. 3A). The fact that they contain lipid storage granules might be a common trait of MTB (Lefèvre et al., 2009; Schultheiss et al., 2005; Silva et al., 2008). The biosynthesis of lipid storage granules is supposedly promoted in response to stress imposed on the cells and during unbalanced growth. These white granules act as storage compounds for energy and carbon needed for maintenance of metabolism and synthesis of cellular metabolites during starvation, in particular if growth causes consumption to significantly increase (Waltermann and Steinbuchel, 2005). 3.4. Magnetic property of the QH-2 strain The thermal demagnetization curves of SIRM5K-2.5T are shown in Fig. 4A. Both FC and ZFC curves show sharp drops in remanence between 90 and 112 K. This confirms the magnetite magnetosome composition as revealed by the TEM study. The determined Tv was 108 K, and dFC and dZFC were 0.43 and 0.29, respectively, yielding a d ratio of 1.5. The higher dFC and lower d ratios were possibly due to the chain arrangement of magnetosomes in QH-2 cells, i.e. short axis alignment. The hysteresis loop of the QH-2 cell samples was potbellied. The values of hysteresis parameters such as Bc and Please cite this article in press as: Zhu, K., et al., Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea, Research in Microbiology (2010), doi:10.1016/j.resmic.2010.02.003 ARTICLE IN PRESS 6 K. Zhu et al. / Research in Microbiology xx (2010) 1e8 Fig. 4. Magnetic characteristics of QH-2 cells. (A) FC-SIRM5K-2.5T (solid squares) and ZFC-SIRM5K-2.5T (open squares) warming curves; (B) room temperature hysteresis loop; (C) room temperature FORC diagram and profiles of coercivity field (Hc) and interaction field (Hb) distribution through the peak of FORC distribution (r). Bcr, and ratios Bcr/Bc and Mr/Ms were deduced as 29.0 mT and 36.1 mT, and 1.24 and 0.55, respectively. These indicated that the QH-2 magnetosomes were all uniaxial SD particles (Fig. 4B). The FORC diagram of QH-2 cells had a closed concentric contour around a central peak Hc,FORC ¼ 34.4 mT, indicating a typical feature of SD magnetite magnetosomes. The characteristic interaction field, Hb1/2, which was defined as the value of the Hb field where the peak of the FORC distribution was reduced to half of its maximum value, was 2.4 mT. The narrow vertical spread of QH-2 samples in the FORC diagram indicated no or weak intercell or interchain magnetostatic interactions (Fig. 4C). 3.5. Characterization of QH-2 motility It was reported, by Spormann and Wolfe (1984) for spirilla and by Frankel et al. (1997) for cocci, that migration in the magnetic field is determined by an aerotactic sensory system. There are two kinds of magneto-aerotactic behaviors: axial or polar magnetotaxis. The axial magnetotactic Magnetospirillum spp. cells swim in both directions along the magnetic field, whereas the polar magnetotactic bacteria swim persistently toward one direction in a magnetic field (this is the case for the marine Magnetococcus sp. MC-1 strain) (Frankel et al., 1997). QH-2 cells were amphitrichously flagellated (Fig. 1C). Interestingly, QH-2 cells did not swim equally in both directions under oxic conditions in the hanging drop assay under the microscope. Most QH-2 cells were north-seeking and accumulated on the north side of the droplet (Fig. 5A). This is consistent with the behavior of polar MTB in the northern hemisphere (Qingdao: N36.1 300 , E120.3 100 ). However, the proportion of QH-2 cells at the north side compared to those at the south side was not as high as that measured with other polar MTB from the northern hemisphere (such as the case of MC-1) and the proportion varied with different culture conditions and different growth phases. Moreover, once the cells on the north side were removed and then analyzed in a fresh droplet, the population was re-segregated to the north and south sides. Polar magnetotaxis has also been observed for other freshwater magnetospirilla that are normally axial magneto-aerotactic (Frankel et al., 2006). The motility track of the QH-2 cells was recorded by using dark-field optical microscopy. They swam roughly along a straight line with a velocity ranging from 20 to 50 mm/s (Fig. 5B). Occasionally, a cell was observed to change from Please cite this article in press as: Zhu, K., et al., Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea, Research in Microbiology (2010), doi:10.1016/j.resmic.2010.02.003 ARTICLE IN PRESS K. Zhu et al. / Research in Microbiology xx (2010) 1e8 7 Fig. 5. Motility of QH-2 cells. Cells accumulated at the north and south edges of a droplet. (A) Motility tracks of QH-2 cells in geomagnetic field were recorded using dark-field optical microscopy. (B) Trajectory sections ‘1’, ‘2’ and ‘3’, separated by arrows, show cell motion in straight line, helix and straight line, respectively. Scale bar shows 30 mm, C1 shows that cells accumulated at the north end in a droplet before flashing with a blue light at 450e480 nm at the northern edge of the droplet, whereas C2 shows swimming away of cells from the droplet edge 1 min after flashing. The geomagnetic field direction is indicated. the straight line motion (Fig. 5B, trajectory section 1) to a helix (section 2) and then back to the straight line (section 3). The helical pattern in the trajectory section 2 was unlikely to result from the sticking of the flagellum onto the glass cover slip, since it corresponded to a 100 mm distance that the cell covered without an obvious change in swimming velocity. Such change of motion mode seemed to be in concordance with the change in swimming direction of QH-2. Bacteria can sense a wide range of environmental signals that steer bacterial locomotion through the extensively studied chemotaxis mechanism (Hazelbauer et al., 2008; Wadhams and Armitage, 2004). The chemoreceptors MCPs (methylaccepting chemotaxis proteins) detect the stimuli and influence cellular locomotion via histidine protein kinase CheA, the phosphorylation state of the response regulator CheY. CheY-P interacts with flagellar motor and switches the rotation direction. Rotation in one direction results in smooth swimming, whilst switching of the rotation direction may lead to backward motion, tumbling or swimming pause (Hazelbauer et al., 2008). Therefore, smooth swimming of a bacterium is periodically interrupted by a change in direction. The frequently used bacterial motility apparatus is the rotatory flagella, as is the case for QH-2 cells. When propelled by flagella, bacterial cells translate along and rotate around the long cellular axis. The overview of the swimming trajectory appears as a straight line when the translation axis overlaps with the rotation axis. However, when the two axes deviate, the trajectory displays itself as a helix. The helical trajectory was occasionally observed for QH-2 cells, as indicated by the trajectory section 2 in Fig. 5B. This point corresponds to a change in swimming direction, from parallel to perpendicular to the magnetic field. It is possible that such a helical trajectory was due to a poorly aligned magnetosome chain in the cell, as observed for the cell in Fig. 1B. Moreover, the cell might not have enough mature magnetosomes to enable constant alignment of the cell along the magnetic field lines. When the cell is well aligned along the magnetic field lines (the sections 1 and 3), the three axes (translation, rotation and magnetic dipolar moment) overlap and result in a straight trajectory. In contrast, if the cell does not align with the magnetic field lines, as observed for section 2 of which the translation direction is perpendicular to the magnetic field orientation, the three axes deviate. As a consequence, the overview of the trajectory is helical. It is well known that light is one of the major changing stimuli for many bacterial species. Blue and UV lights cause damage to almost all living systems. Blue light is particularly damaging if oxygen is present. It was reported that cells of the marine Magnetococcus sp. MC-1 strain in a capillary tube exhibited a response to a short-wavelength light (500 nm), Please cite this article in press as: Zhu, K., et al., Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea, Research in Microbiology (2010), doi:10.1016/j.resmic.2010.02.003 ARTICLE IN PRESS 8 K. Zhu et al. / Research in Microbiology xx (2010) 1e8 which caused them to swim away from the meniscus (high [O2]) and toward the petroleum jelly plug (low [O2]) persistently parallel to the magnetic field during the illumination, resulting in a reaction similar to an increase in [O2] (Frankel et al., 1997). The QH-2 cells were studied for their response to shortwavelength light in a droplet. It was observed using a light microscope that most QH-2 cells swam to the north side and accumulated at the edge under normal conditions. When illuminating the cells (Fig. 5C1) with the wavelengths ranging from 330 to 550 nm, the emitted energy triggered the QH-2 cells swimming away from the edge to the interior of the droplet (from high to low [O2]) (Fig. 5C2). Such cell behavior was similar to that observed for MC-1 cells. However, unlike the case of MC-1 in which virtually all cells left the original position, nearly half of the QH-2 cells appeared to stop motility upon illumination and become agglomerated instead of swimming away (Fig. 5C2). In contrast, magnetotactic spirillum AMB-1, one of the most extensively studied axial magneto-aerotaxis models, apparently did not respond to the illumination process in the same way (data not shown). The mechanism of QH-2 cell reaction to light requires further investigation. As the axenic culture of QH-2 was isolated from the intertidal region, the bacteria in the sediments might be regularly exposed to air and light or the changing chemical gradients with the tides. Therefore, the pure culture of QH-2 could be used as a novel strain model for the study of the mechanism of magneto-aerotaxis in combination with the cellular reaction to light and oxygen. Acknowledgments This work was supported by the NSFC (Nos. 40776094 and 40906069), the Haiwaijiechuxuezhe Fund of the Chinese Academy of Sciences (No. 2006-1-15), a CNRS scholar fellowship (to K.L. Zhu), a special fund of Creative Projects for the Postdoctors of Shandong Province (082318101N) and K.C. Wong Education Foundation, Hong Kong. We thank W. Jiang, T. Song, C. Chen, W.J. Zhang, C.L. Santini and N. Philippe for discussions and advice, and M. Jiang for assistance in electron microscopy observations. References Bazylinski, D.A., Frankel, R.B., 2004. Magnetosome formation in prokaryotes. Nat. Rev. Microbiol. 2, 217e230. Bazylinski, D.A., Frankel, R.B., Garratt-Reed, A.J., Mann, S., 1988. Anaerobic production of magnetite by a marine magnetotactic bacterium. Nature 334, 518e519. Blakemore, R.P., Maratea, D., Wolfe, R.S., 1979. Isolation and pure culture of a freshwater magnetic spirillum in chemically defined medium. J. Bacteriol. 140, 720e729. Chen, A., Egli, R., Moskowitz, B.M., 2007. First-order reversal curve (FORC) diagrams of natural and cultured biogenic magnetic particles. J. Geophys. Res. B: Solid Earth 112, B08S90. DeLong, E.F., Frankel, R.B., Bazylinski, D.A., 1993. Multiple evolutionary origins of magnetotaxis in bacteria. Science 259, 803e806. Devouard, B., Posfai, M., Hua, X., Bazylinski, D.A., Frankel, R.B., Buseck, P.R., 1998. Magnetite from magnetotactic bacteria; size distributions and twinning. Am. Mineral. 83, 1387e1398. Flies, C.B., Jonkers, H.M., de Beer, D., Bosselmann, K., Bottcher, M.E., Schüler, D., 2005. Diversity and vertical distribution of magnetotactic bacteria along chemical gradients in freshwater microcosms. FEMS Microbiol. Ecol. 52, 185e195. Frankel, R.B., Bazylinski, D.A., Johnson, M.S., Taylor, B.L., 1997. Magnetoaerotaxis in marine coccoid bacteria. Biophys. J. 73, 994e1000. Frankel, R.B., Williams, T.J., Bazylinski, D.A., 2006. Magneto-aerotaxis. In: Schüler, D. (Ed.), Magnetoreception and Magnetosomes in Bacteria. Springer-Verlag, Berlin, Heidelberg, pp. 2e24. Greenspan, P., Mayer, E.P., Fowler, S.D., 1985. Nile red: a selective fluorescent stain for intracellular lipid droplets. J. Cell. Biol. 100, 965e973. Harrison, R., Feinberg, J., 2008. FORCinel: an improved algorithm for calculating first-order reversal curve distributions using locally weighted regression smoothing. Geochem. Geophys. Geosyst. 9, Q05016. Hazelbauer, G.L., Falke, J.J., Parkinson, J.S., 2008. Bacterial chemoreceptors: high-performance signaling in networked arrays. Trends Biochem. Sci. 33, 9e19. Lang, C., Schüler, D., 2006. Biogenic nanoparticles: production, characterization, and application of bacterial magnetosomes. J. Phys. Condens. Matter 18, S2815eS2828. Lefèvre, C.T., Bernadac, A., Yu-Zhang, K., Pradel, N., Wu, L.-F., 2009. Isolation and characterization of a magnetotactic bacterial culture from the Mediterranean Sea. Environ. Microbiol. 11, 1646e1657. Li, W., Yu, L., Zhou, P., Zhu, M., 2007. Isolation of magnetotactic bacterium WM-1 from freshwater sediment and phylogenetic characterization. Arch. Microbiol. 188, 97e102. Matsunaga, T., Sakaguchi, T., Tadakoro, F., 1991. Magnetite formation by a magnetic bacterium capable of growing aerobically. Appl. Microbiol. Biotechnol. 35, 651e655. Meldrum, F.C., Heywood, B.R., Mann, S., Frankel, R.B., Bazylinski, D.A., 1993. Electron microscopy study of magnetosomes in two cultured vibroid magnetotactic bacteria. Proc. R. Soc. Lond. B 251, 237e342. Moskowitz, B., Frankel, R., Bazylinski, D., 1993. Rock magnetic criteria for the detection of biogenic magnetite. Earth Planet. Sci. Lett. 120, 283e300. Okamura, Y., Takeyama, H., Sekine, T., Sakaguchi, T., Wahyudi, A.T., Sato, R., Kamiya, S., Matsunaga, T., 2003. Design and application of a new crypticplasmid-based shuttle vector for Magnetospirillum magneticum. Appl. Environ. Microbiol. 69, 4274e4277. Pan, H., Zhu, K., Song, T., Yu-Zhang, K., Lefevre, C., Xing, S., Liu, M., Zhao, S., Xiao, T., Wu, L.F., 2008. Characterization of a homogeneous taxonomic group of marine magnetotactic cocci within a low tide zone in the China Sea. Environ. Microbiol. 10, 1158e1164. Roberts, A., Pike, C., Verosub, K., 2000. First-order reversal curve diagrams: a new tool for characterizing the magnetic properties of natural samples. J. Geophys. Res. 105, 28461e28476. Sakaguchi, T., Arakaki, A., Matsunaga, T., 2002. Desulfovibrio magneticus sp. nov., a novel sulfate-reducing bacterium that produces intracellular singledomain-sized magnetite particles. Int. J. Syst. Evol. Microbiol. 52, 215e221. Schüler, D., 2002. The biomineralization of magnetosomes in Magnetospirillum gryphiswaldense. Int. Microbiol. 5, 209e214. Schüler, D., Koehler, M., 1992. The isolation of a new magnetic spirillum. Microbiol. Res. 147, 150e151. Schultheiss, D., Handrick, R., Jendrossek, D., Hanzlik, M., Schüler, D., 2005. The presumptive magnetosome protein Mms16 is a poly (3-hydroxybutyrate) granule-bound protein (phasin) in Magnetospirillum gryphiswaldense. J. Bacteriol. 187, 2416. Silva, K.T., Abreu, F., Keim, C.N., Farina, M., Lins, U., 2008. Ultrastructure and cytochemistry of lipid granules in the many-celled magnetotactic prokaryote, ‘Candidatus Magnetoglobus multicellularis’. Micron 39, 1387e1392. Spormann, A.M., Wolfe, R.S., 1984. Chemotactic, magnetotactic and tactile behavior in a magnetic spirillum. FEMS Lett. 22, 171e177. Wadhams, G.H., Armitage, J.P., 2004. Making sense of it all: bacterial chemotaxis. Nat. Rev. Mol. Cell Biol. 5, 1024e1037. Waltermann, M., Steinbuchel, A., 2005. Neutral lipid bodies in prokaryotes: recent insights into structure, formation, and relationship to eukaryotic lipid depots. J. Bacteriol. 187, 3607e3619. Wolfe, R., Thauer, R., Pfennig, N., 1987. A ‘‘capillary racetrack’’ method for isolation of magnetotactic bacteria. FEMS Microbiol. Ecol. 45, 31e35. Please cite this article in press as: Zhu, K., et al., Isolation and characterization of a marine magnetotactic spirillum axenic culture QH-2 from an intertidal zone of the China Sea, Research in Microbiology (2010), doi:10.1016/j.resmic.2010.02.003