Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Protein moonlighting wikipedia , lookup
Purinergic signalling wikipedia , lookup
Hedgehog signaling pathway wikipedia , lookup
NMDA receptor wikipedia , lookup
List of types of proteins wikipedia , lookup
Protein phosphorylation wikipedia , lookup
Tyrosine kinase wikipedia , lookup
VLDL receptor wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
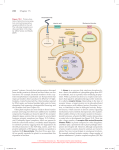
Review TRENDS in Plant Science Vol.8 No.5 May 2003 231 Making inroads into plant receptor kinase signalling pathways Gabrielle Tichtinsky1, Vincent Vanoosthuyse2, J. Mark Cock3 and Thierry Gaude1 1 Reproduction et Développement des Plantes, UMR 5667 CNRS-INRA-ENSL-UCBL, École Normale Supérieure de Lyon, 46 allée d’Italie, F-69364 Lyon Cedex 07, France 2 Swann 508, ICMB, University of Edinburgh, King’s buildings, Mayfield Road, Edinburgh, UK EH9 3JR 3 UMR 1931 CNRS-Goëmar, Station Biologique, Place Georges Teissier, BP74, 29682 Roscoff Cedex, France Cell-membrane-located receptor kinases play important roles in many plant signal-transduction pathways. Exciting progress has been made in recent years with the characterization of four ligand–receptor systems involved in physiological processes as diverse as selfpollen rejection, stem-cell maintenance and differentiation at the shoot meristem, the response to the brassinosteroid hormones and the innate response to bacterial pathogens. These new findings emphasize the remarkably high diversity of these signalling pathways, although some downstream components are shared. This observation supports the idea that the wide diversification of plant receptors is associated with a high degree of specialization, one receptor potentially regulating a single developmental process. However, the possibility that one receptor might have a dual recognition function cannot be ruled out. The plant receptor kinase (PRK, see Glossary), formally, receptor-like kinase [1], gene family constitutes a large, monophyletic group that includes .400 genes in Arabidopsis. These genes encode typical single-pass, membrane-anchored receptor kinases, with an extracellular N-terminal domain and an intracellular C-terminal kinase domain [2]. Although PRKs are less well characterized than their animal counterparts, there appear to be many features common to receptor signalling pathways in the two kingdoms [1]. The leucine-rich repeat (LRR) subfamily of receptors (Table 1) represents a large proportion of the PRKs characterized so far. This family includes CLAVATA1 (CLV1), which controls stem-cell maintenance in apical meristems [3], the brassinosteroid-insensitive 1 (BRI1) receptor and its co-receptor BRI1-associated receptor kinase 1 (BAK1 ), which mediate the response to brassinosteroid (BR ) hormones [4 – 6], and the flagellin-sensing 2 (FLS2) receptor, which mediates the innate response to bacterial pathogens [7]. The S-locus receptor kinase (SRK) is the female determinant of self-incompatibility in Brassicaceae [8]. It belongs to the S-domain subfamily of PRKs (Table 1). Exciting progress has been made in recent years with the identification of specific ligands and characterization of components of the downstream signalling pathways of these four regulators. In this article, we compare and contrast these four signalling systems. Nature of the cognate ligands Most of the identified ligands for PRKs are peptides (Table 1). S-locus cysteine-rich protein (SCR), for example, is the male determinant of self-incompatibility in the Brassicaceae. It encodes the SRK ligand, a highly polymorphic secreted peptide [9,10]. Interestingly, a search of the Arabidopsis genome revealed the presence of at least 114 genes that are predicted to encode peptides with a similar primary structure to SCR [11]. It is tempting to postulate that some of these genes might encode ligands for other members of the S-domain PRK subfamily [11]. CLAVATA3 (CLV3), the CLV1 ligand, is also a secreted peptide but it shares no sequence similarity with SCR [12,13]. As for plant defences, flagellin, a major protein constituent of bacterial flagella, elicits plant responses efficiently. Its most conserved peptide, flg22 , is sufficient to elicit these responses [7,14]. Several other peptides, such as AtGRP-3, systemin, phytosulfokine and Lat52, have also been identified as putative PRK ligands [15 – 18]. Nevertheless, not all putative PRK ligands are peptides: BRI1 and BAK1 mediate the response to plant steroid Glossary ARC1: Armadillo-repeat-containing 1 BAK1: BRI1-associated receptor kinase 1 BES2: BRI-EMS-suppressor 2 BIN2: BR-insensitive 2 kinase BL: brassinolide BR: brassinosteroid BRI1: brassinosteroid-insensitive 1 receptor BZR1: brassinazole-resistant 1 EGF: epidermal growth factor EGFR: EGF receptor eSRK: extracellular domain of SRK flg22: flagellin 22 FLS2: flagellin-sensing 2 KAPP: kinase-associated protein phosphatase LRR: leucine-rich repeat MAPK: mitogen-activated protein kinase PRK: plant receptor kinase Rop: Rho-GTPase-related protein SCR: S-locus cysteine rich SLG: S-locus glycoprotein SRK: S-locus receptor kinase THL: thioredoxin-h-like Corresponding author: Thierry Gaude ([email protected]). http://plants.trends.com 1360-1385/03/$ - see front matter q 2003 Elsevier Science Ltd. All rights reserved. doi:10.1016/S1360-1385(03)00062-1 232 Review TRENDS in Plant Science Vol.8 No.5 May 2003 Table 1. Characteristics of the four ligand –receptor systems analysed in this article Physiological process Receptor Cognate ligand Refs Name Subfamilya Name Nature Molecular massb Stem-cell maintenance in apical meristems CLV1 LRR-PRK CLV3 Secreted peptide Response to the brassinosteroid hormones Innate response to bacterial pathogens Self-incompatibility BRI1, BAK1 FLS2 SRK LRR-PRK LRR-PRK S-domain receptor kinases BR, BL Flagellin SCR Steroid hormones Bacterial protein Secreted peptide, basic and cysteine-rich Predicted: 8.7 kDa; observed: 25.0 kDa 480 Da 33.0 kDa Predicted: 5.7 –8.0 kDa; observed: 16.0 kDa [3,12,13] [4,6,19] [7,30] [8,9,62] Abbreviations: BAK1, BRI1-associated receptor kinase 1; BL, brassinolide; BR, brassinosteroids; BRI1, brassinosteroid-insensitive 1; CLV1, CLAVATA 1; CLV3, CLAVATA 3; FLS2, flagellin-sensing 2; LRR-PRK, leucine-rich repeat; SCR, S-locus cysteine rich; SRK, S-locus receptor kinase. a The subfamilies of plant receptor kinases differ in their N-terminal extracellular domains. The LRR-PRK subfamily has an extracellular domain consisting of tandemly repeated LRRs, flanked in most cases by two conservatively spaced cysteine pairs. The S-domain subfamily is characterized by an extracellular domain similar to the S (selfincompatibility) locus glycoprotein [36]. b Predicted molecular mass is based on the deduced amino acid sequence. The range of molecular masses indicated for SCR refers to the many allelic forms of this protein. Observed molecular masses might correspond to dimers or complexes of ligands. hormones (brassinosteroids), particularly brassinolide (BL), and the bacterial Nod factors, which are lipochitin oligosaccharides essential for symbiotic nodule development in leguminous plants, might be sensed by a LRR PRK, the nodulation receptor kinase NORK [5,6,19,20]. Ligand –receptor binding The analysis of the ligand-binding properties at the cellular level provides information about the functional sites in vivo. However, the identification of the biochemical nature of these functional sites (i.e. whether they are composed of a single PRK or of homo- or heterodimers, or of bigger protein complexes) remains in most cases a major challenge. Functional binding sites for flg22, BL and SCR have been characterized at the cellular level (Table 2). Binding of 125I-labelled flg22 to suspension-cultured cells of tomato or Arabidopsis is rapid (maximal binding reached within 15 – 20 min), stable (over at least 90 min) and irreversible under physiological conditions [21,22]. The high stability of the interaction might be due to either a low rate of dissociation or stabilization of the complex. Investigation of the cellular fate of ligand– receptor complexes after ligand perception should provide some insight into this putative stabilization process. In contrast to flg22 and BL binding (Table 2), 125I-labelled SCR binding to stigma microsomal membranes revealed two types of binding site: an abundant low-affinity site and a less-abundant highaffinity site [10]. Interestingly, a similar feature has been described when the animal epidermal growth factor (EGF) binds to its receptor (EGFR), ErbB1 [23,24]. Direct protein interaction between SCR and SRK has been demonstrated in vitro using SCR produced in the periplasmic space of Escherichia coli and the extracellular domain of SRK (eSRK) produced in Nicotiana benthamiana leaves [25]. This physical interaction was confirmed using an in vitro binding and immunoprecipitation assay in which synthetic radioactive-iodinated SCR was mixed with stigma microsomal membranes containing SRK [10]. Interaction between CLV3 and CLV1, and BL and BRI1 was detected in vivo by co-immunoprecipitation from plant extracts [26,27]. These observations do not rule out the possibility that the interaction between the ligand and its receptor might require additional partners. For example, a dominant mutation, brs1, was shown to suppress weak alleles of bri1 in genetic backgrounds in which normal levels of brassinosteroids are produced [28]. Because BRS1 encodes a secreted carboxypeptidase, it was suggested that BRS1 processes either the receptor itself or, more probably, a proligand, such as a steroid-binding protein [28]. Recently, genetic data indicated that BRI1 cooperates with BAK1 to transduce the BL signal. Both proteins also interact in vitro and in vivo [5,6]. In previous work, when a chimeric receptor composed of the extracellular and transmembrane domains of BRI1 fused to the intracellular kinase domain of Xa21 (an LRR PRK involved in the defence response) was produced in rice cells, treatment with BL triggered cellular defence responses. This indicates that the extracellular and transmembrane domains of BRI1 can recognize BL and transduce this signal to a heterologous kinase domain [29]. Whether BAK1 is required for BL binding is unclear. It could simply be necessary to initiate downstream signalling leading to the BL response. However, an endogenous rice homologue of BAK1 might be present in the cells, allowing BL binding. Further work is required to distinguish between these alternatives. Binding of flagellin or of flg22 to purified FLS2 has not yet been demonstrated. Nevertheless, binding of flg22 to crude plant extracts is much reduced in fls2 mutants and binding is restored following complementation with a wildtype copy of FLS2 [30]. These data indicate that FLS2 is a Table 2. Ligand-binding properties at the cellular level Ligand Binding system Binding capacitya Dissociation constant (Kd) Refs 125 Suspension-cultured cells Microsomal membranes Microsomal membranes 2.00 pmol mg21 0.23 pmol mg21 1.00 pmol mg21 0.25 pmol mg21 1.3 –1.6 nM 7.4 –10.8 nM 32.0 nM (low-affinity site) 1.2 nM (high-affinity site) [22] [27] [10] I-flg22 3 H-BL 125 I-SCR a Expressed as pmol of bound ligand per mg of microsomal proteins. http://plants.trends.com Review TRENDS in Plant Science crucial component of the membrane-located flg22 binding site. This assumption is supported by an analysis of the Arabidopsis ecotype Ws-0, which is insensitive to flg22. Specific binding of flg22 to leaf microsomal membranes is greatly reduced in Ws-0 compared with ecotype Landsberg erecta [21]. The locus responsible for this difference between ecotypes, originally referred to as FLS1, has recently been shown to be identical to FLS2 [31]. In support of these genetic data, affinity cross-linking of flg22 to microsomal membrane fractions allowed the identification of a single protein of ,115 kDa, which is the predicted size of FLS2 [22]. Receptor complexes When analysed under native conditions, PRKs are usually associated with other proteins. Thus, two protein complexes including CLV1 were detected in cauliflower extracts [26,32]. The smaller one (185 kDa) includes a second protein linked by one or more disulfide bonds. CLV2, a gene acting in the CLV1– CLV3 pathway, encodes an LRR-receptor-like protein resembling CLV1 but with only a short cytoplasmic tail and no intracellular kinase domain [33,34]. In clv2 mutants, the stability of CLV1 is reduced [3], whereas the 185 kDa complex is partially disrupted in a clv1 mutant in which the LRR structure is altered [32]. Therefore, CLV1 and CLV2 probably interact through their LRR domains to form the 185 kDa complex (Fig. 1). CLV3 is not detected in the 185 kDa complex but is present in a bigger, 450 kDa complex that also contains kinase-associated protein phosphatase (KAPP) and Rho-GTPase-related proteins (Rop) [26,32]. Analysis of CLV1 complexes in mutant backgrounds revealed that a functional CLV3 allele and the kinase activity of CLV1 are required for assembly of the 450 kDa complex [26]. Thus, the 450 kDa complex is probably the active ligand– receptor complex (Fig. 1). Like CLV1, SRK was found to form protein complexes in vivo. In the basal state (i.e. in unpollinated stigmas), cross-linking experiments indicated that SRK interacts with other cellular components, in complexes of , 161 kDa and , 233 kDa [35]. The second complex of , 233 kDa has the expected size for an SRK dimer. Interestingly, soluble proteins encoded by S-locus genes are secreted by stigmatic papillar cells. They include a soluble truncated form of SRK (eSRK), which corresponds to the SRK extracellular domain alone, and a highly similar protein, SLG (S-locus glycoprotein) [36]. Using co-immunoprecipitation and sucrose-gradient-based approaches, neither eSRK nor SLG were detected in complexes including SRK in unpollinated stigmas [35] (D. Cabrillac et al., unpublished). However, cross-linking experiments performed on stigma proteins after binding of labelled SCR to stigma microsomal fractions identified two radiolabelled proteins of 120 kDa and 65 kDa, which correspond to SRK and either eSRK or SLG, respectively [10]. The 65 kDa protein was also co-immunoprecipitated with SRK from stigma extracts. These results suggest that SCR can bind to both SRK and eSRK or SLG, and that SCR binding initiates association of SRK with the soluble stigma proteins to form heterodimers (Fig. 1). Binding affinity data suggest that the complex of SRK plus eSRK or SLG, might constitute http://plants.trends.com Vol.8 No.5 May 2003 233 the high-affinity binding site for SCR on the papillar cell membrane [10]. Similarly to the self-incompatibility recognition system, EGF binding by EGFR implies two states of different affinities [23,37,38]. The proportion of EGFR in the high-affinity state is modulated by its heteromerization with ErbB2, another member of the ErbB family [23,38]. Whether SRK behaves in a similar way through its association with soluble proteins remains to be confirmed. Activation of the receptor All the PRKs discussed in this article share a homologous intracellular kinase domain and autophosphorylate in vitro on serine and threonine residues when this domain is produced in E. coli [5,30,36,39]. The physiological importance of this kinase activity is stressed by the many PRK mutants with lesions affecting the kinase domain [7,40,41]. In vivo, there is evidence that the binding of ligands to PRKs induces autophosphorylation and activation. SRK, for example, is phosphorylated in vivo within an hour of an incompatible pollination [42]. This autophosphorylation can also be triggered in vitro by treating plasma-membrane fractions with SCR peptides [10]. Transphosphorylation of a kinase-inactive form by an active one was shown to occur when both forms are co-expressed in a membranous environment [35]. BL treatment also induces BRI1 phosphorylation in plants previously treated with inhibitors of brassinolide production [27]. However, the ability of the complete BRI1 protein expressed in either bacterial or yeast cells to autophosphorylate remains controversial [6,43]. Experiments in which wild-type and kinase-negative forms of BRI1 and BAK1 were expressed in yeast suggest that activation of the kinase activity of these receptors requires heterodimerization and transphosphorylation [6]. There are no data available for the integral CLV1 receptor but a kinase-active form of the cytosolic domain of CLV1 does transphosphorylate a kinase-inactive form of the same domain in vitro [44]. Taken together, these observations indicate that activation of receptor kinases (in either a homo- or a hetero-oligomeric configuration) by transphosphorylation might be a common feature of PRK signalling pathways. Regulation of kinase activity Analysis of animal receptor kinases has highlighted the importance of regulating kinase activity as a means to prevent inappropriate activation of downstream signal transduction pathways. The plant phosphatase KAPP interacts in vitro with the kinase domain of many PRKs including those of SRK [45], CLV1 [44 – 46] and FLS2 [30]. In the case of CLV1 and SRK, interaction does not occur with mutant forms altered in their kinase activity [44,46] (V. Vanoosthuyse et al., unpublished), suggesting that the interaction depends on the phosphorylation status or activity of the receptor. KAPP is also phosphorylated by CLV1 and SRK, and can dephosphorylate both receptors [44] (V. Vanoosthuyse et al., unpublished). The role of KAPP in PRK regulation was investigated by producing transgenic plants with up- or downregulated KAPP Review 234 TRENDS in Plant Science Vol.8 No.5 May 2003 (a) (b) Inactive complex Active complex Down-regulated complex Active Sterol binding complex peptide? CLV2 CLV1 CLV3 BRI1 BAK1 BR X SS SS SS SS ? KAPP POL SS ROP ? MAPK cascade? BIN2 Stem cell identity genes BES2 and BZR1 BR response genes WUS (c) Active complex FLS2 Downregulated complex flg22 (d) Inactive complex eSRK or SLG SRK Active complex Down-regulated complex SCR THL KAPP ARC1 MAPK cascade MEKK1 MKK4/5 MPK3/6 Defence response genes Self-incompatibility response TRENDS in Plant Science Fig. 1. Models of signal transduction through four plant receptor kinases (PRKs). (a) CLV3–CLV1 signalling. The inactive CLAVATA complex is composed of disulfide-bound CLAVATA 1 (CLV1) and CLAVATA 2 (CLV2) proteins. Binding of the ligand CLAVATA 3 (CLV3), perhaps in association with an unknown X factor, induces receptor oligomerization and transphosphorylation followed by phosphorylation of kinase-associated protein phosphatase (KAPP). Rho-GTPase-related protein (Rop), (which might interact directly or via an unknown intermediate with the CLV1– CLV2 complex), might be responsible for signal transduction downstream of CLV1. Repression of WUSCHEL (WUS) expression enables progression of meristem stem cells toward differentiation. Genetic evidence indicates that dephosphorylation by KAPP downregulates the activated receptor complex. The POLTERGEIST (POL) protein functions as a negative regulator of CLV1 signalling. (b) BR– BRI1– BAK1 signalling. Binding of brassinosteroids (BR), perhaps in association with a sterol binding protein, activates heterodimers of brassinosteroid-insensitive 1 receptor (BRI1) and BRI1-associated receptor kinase 1 (BAK1), leading to mutual transphosphorylation. Signal transduction from the activated receptor complex leads to inactivation of the BR-insensitive 2 (BIN2) kinase probably via one or more unknown intermediates. The reduced BIN2 activity in turn leads to an increase in unphosphorylated BRI-EMS-suppressor 2 (BES2) and brassinazole-resistant 1 (BZR1), which move into the nucleus and trigger at least some of the cell responses to BR. (c) flg22– FLS2 signalling. Binding of flagellin 22 (flg22) to flagellin sensing 2 (FLS2) triggers kinase activation. It is not known whether this involves FLS2 oligomerization. FLS2 activation, in turn, triggers a mitogen-activated protein (MAP) kinase cascade, putatively through signal-transduction intermediates. KAPP also interacts with FLS2, probably to downregulate its activity. (d) SCR –SRK signalling. In the basal (unpollinated) state, S-locus receptor kinase (SRK) is present as a monomer and as a homodimer and is maintained in an inactive state by interacting with thioredoxin-hlike (THL). After self-pollination, the ligand, S-locus cysteine-rich (SCR), binds as a dimer either to an SRK homodimer or to a hetetodimer of SRK with extracellular SRK (eSRK) or S-locus glycoprotein (SLG). SCR binding is thought to induce receptor oligomerization and transphosphorylation, followed by phosphorylation of armadillorepeat-containing 1 (ARC1) and KAPP. ARC1 is a positive effector of the self-incompatibility response. KAPP probably dephosphorylates SRK and consequently downregulates the activated SRK complex. Red stars indicate protein phosphorylation. expression. A floral phenotype reminiscent of clv1 loss of function alleles was observed in plants overexpressing KAPP [44,46]. Moreover, reducing the level of KAPP expression suppressed the clavata phenotype of the weak clv1 – 1 mutant allele [46]. These results suggest that KAPP probably acts as a negative regulator of http://plants.trends.com CLAVATA signalling, probably by dephosphorylating CLV1. KAPP-overexpressing plants also mimic fls2 mutants in their response to treatment with flg22, and have a much lower flg22 binding capacity [30]. Taken together, these data allow us to suggest a general model in which KAPP binds to activated, phosphorylated PRKs, is Review TRENDS in Plant Science in turn phosphorylated by PRKs and then downregulates them by dephosphorylating the kinase domain (Fig. 1). A possible mechanism for this downregulation has been suggested by recent work on another PRK, AtSERK1 [47]. AtSERK1 interacts with KAPP in a phosphorylationdependent manner and in vivo protein –protein interaction experiments indicate that this interaction occurs in intracellular vesicles. This raises the possibility that KAPP mediates PRK downregulation by modulating receptor endocytosis. Recently, we discovered two novel SRK-interacting proteins using a yeast two-hybrid screen (V. Vanoosthuyse et al., unpublished). These interactors (calmodulin and a sorting-nexin-like protein) bind to the kinase domain of SRK and of several other PRKs in vitro. Remarkably, calmodulin and sorting nexin have both been shown to interact with the kinase domain of EGFR [48,49]; sorting nexin is involved in EGFR internalization [49]. Although the physiological relevance of the interactions of calmodulin or sorting-nexin-like protein with PRK remains to be elucidated, we can presume a conserved regulatory function of these proteins in plant cell signalling. Another protein, thioredoxin-h-like (THL) has been shown to act as a negative regulator of SRK. In vitro, THL inhibits the kinase activity of recombinant SRK from microsomal fractions of insect cells [42]. This inhibition is released following the addition of self-pollen proteins, presumably as a result of specific SCR binding to SRK [42]. We suppose that thioredoxins maintain SRK in an inactive state in vivo in the absence of its ligand (Fig. 1). Downstream signalling events In general, signal transduction downstream of receptor kinases involves cell effectors that interact with and are phosphorylated by the cytoplasmic domain of the receptor. Armadillo (ARM)-repeat-containing protein 1 (ARC1) is an SRK-interacting partner that satisfies both criteria [50]. Transgenic plants with reduced levels of ARC1 transcripts showed a partial breakdown of self-incompatibility, indicating that ARC1 is a positive effector of self-incompatibility signalling [51]. Interestingly, ARC1 belongs to a subclass of U-box- and ARM-repeat-containing proteins that are encoded by at least 18 genes in Arabidopsis [52]. Although the U-box and ARM repeats are present in some yeast and animal proteins, no clear homologues of the ARC1 subclass of genes are detected in these organisms, suggesting that ARC1 belongs to a plant-specific class of signalling proteins. The signalling pathway downstream of flg22 and FLS2 has largely been unravelled using an in vivo system consisting of mesophyll protoplasts in which defence reactions were induced by exposure to flg22 [53]. A specific mitogen-activated protein kinase (MAPK) cascade is involved, including AtMEKK1, AtMAPKK4/AtMAPKK5 and AtMAPK3/AtMAPK6. This MAPK cascade stimulates the expression of early (within 30 min of flg22 induction) transcribed genes. It is not known whether MEKK1 is a direct substrate for FLS2 or whether additional signalling intermediates are involved. A possible intermediate is the plant-specific protein AtPhos43, which is specifically http://plants.trends.com Vol.8 No.5 May 2003 235 phosphorylated in cultured cells within 4 min of activation of the flg22/FLS2 pathway [54]. Several components that function downstream of the BRI1 receptor have recently been discovered by genetic screens. BR-insensitive 2 (bin2) is a semidominant mutation conferring brassinolide insensitivity, suggesting that BIN2 acts as a negative regulator of BRI1 signalling [55]. BIN2 encodes a SHAGGY/GSK-3 protein kinase, which in its dominant mutant form (bin2-D) has an increased kinase activity [55]. The two mutations BRI-EMS-suppressor 1 (bes1-D) and brassinazole-resistant 1 – 1D (bzr1–1D) suppress phenotypes of BR-signalling mutants [56,57]. This suggests a function of BZR1 and BES1 as positive regulators of the signalling cascade. BZR1 and BES1 (which is identical to BZR2) encode proteins that belong to a small plant-specific protein family [56,57]. They both accumulate in nuclei in response to BL treatment, as early as 30 min after addition of BL. In the nucleus, BES1 accumulates predominantly as an unphosphorylated form. Interestingly, in a bin2-D/BIN2 background (i.e. in plants with enhanced BIN2 kinase activity), the accumulation of the dephosphorylated form of BES1 in response to BL is greatly reduced [57]. Taken together these data suggest a model [57] in which, in the absence of a BL signal, BES1 is phosphorylated by BIN2 and remains in the cytoplasm, where it is probably degraded. In the presence of brassinosteroids, BIN2 is inhibited and the unphosphorylated form of BES1 accumulates in the nucleus, where it modifies the expression of BL target genes. Like BES1, BZR1 has conserved phosphorylation sites that match the consensus site for SHAGGY/GSK-3 protein kinase phosphorylation and might function in a similar way [56]. In addition to the BRI1 –BAK1 pathway described above, there is also evidence for alternative pathways involving a Hþ-ATPase, MAPKs and calcium-dependent protein kinases downstream of BRI1 [58,59]. Recent data suggest that the tomato homologue of BRI1 is a dual ligand receptor for systemin and brassinosteroids [60]. If this dual recognition function is confirmed, it would be particularly interesting to investigate how the same receptor can trigger two different downstream signalling pathways. In contrast to the other three signalling systems described above, the CLAVATA signal-transduction pathway remains largely unknown. Genetic data indicate that stem-cell maintenance is regulated through a regulation loop involving WUSCHEL (a homeobox gene) and the locus POLTERGEIST, which encodes a protein phosphatase 2C [3,61]. Moreover, it has been suggested that Rop, which is a component of the activated CLV1 receptor complex, might act in a manner analogous to the related Ras protein in animals and activate a downstream MAPK pathway [32]. Conclusion Recently, significant advances have been made in the molecular characterization of signal-transduction pathways mediated by PRKs by studying a limited number of model systems. Several basic features of cell signalling, such as dimerization of receptors following ligand binding, receptor transphosphorylation and the existence of receptor complexes, appear to be common to both plants and 236 Review TRENDS in Plant Science animals. However, there are marked differences between the two kingdoms. The large number of PRKs supports the idea of a high degree of specialization, even though recent data suggest that some PRKs might have a dual recognition function. PRK signalling pathways also seem to be remarkably diverse, with few downstream components being shared by more than one receptor. However, this impression might be partly because of the fragmentary nature of current data and it is likely that common features (such as MAPK cascades) will emerge as each pathway is analysed in more detail and as additional pathways are characterized. The elucidation of many mechanistic aspects of PRK signalling, such as the identification of novel cytosolic signalling components, the mechanism of turnover of activated receptor complexes and the nature of cross-talk between signalling pathways, remain challenges for the coming years. Vol.8 No.5 May 2003 20 21 22 23 24 25 26 27 References 1 Cock, J.M. et al. (2002) Receptor kinase signalling in plants and animals: distinct molecular systems with mechanistic similarities. Curr. Opin. Cell Biol. 14, 230– 236 2 Shiu, S.H. and Bleecker, A.B. (2001) Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. U. S. A. 98, 10763 – 10768 3 Clark, S.E. (2001) Cell signalling at the shoot meristem. Nat. Rev. Mol. Cell Biol. 2, 276 – 284 4 Clouse, S.D. et al. (1996) A brassinosteroid-insensitive mutant in Arabidopsis thaliana exhibits multiple defects in growth and development. Plant Physiol. 111, 671 – 678 5 Li, J. et al. (2002) BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 110, 213– 222 6 Nam, K.H. and Li, J. (2002) BRI1/BAK1, a receptor kinase pair mediating brassinosteroid signaling. Cell 110, 203– 212 7 Gomez-Gomez, L. and Boller, T. (2000) FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 5, 1003 – 1011 8 Takasaki, T. et al. (2000) The S receptor kinase determines selfincompatibility in Brassica stigma. Nature 403, 913– 916 9 Schopfer, C.R. et al. (1999) The male determinant of self-incompatibility in Brassica. Science 286, 1697– 1700 10 Takayama, S. et al. (2001) Direct ligand – receptor complex interaction controls Brassica self-incompatibility. Nature 413, 534– 538 11 Vanoosthuyse, V. et al. (2001) Two large Arabidopsis thaliana gene families are homologous to the Brassica gene superfamily that encodes pollen coat proteins and the male component of the self-incompatibility response. Plant Mol. Biol. 46, 17– 34 12 Fletcher, J.C. et al. (1999) Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 283, 1911 – 1914 13 Rojo, E. et al. (2002) CLV3 is localized to the extracellular space, where it activates the Arabidopsis CLAVATA stem cell signaling pathway. Plant Cell 14, 969– 977 14 Felix, G. et al. (1999) Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 18, 265– 276 15 Park, A.R. et al. (2001) Interaction of the Arabidopsis receptor protein kinase Wak1 with a glycine-rich protein, AtGRP-3. J. Biol. Chem. 276, 26688 – 26693 16 Scheer, J.M. and Ryan, C.A. Jr (2002) The systemin receptor SR160 from Lycopersicon peruvianum is a member of the LRR receptor kinase family. Proc. Natl. Acad. Sci. U. S. A. 99, 9585– 9590 17 Matsubayashi, Y. et al. (2002) An LRR receptor kinase involved in perception of a peptide plant hormone, phytosulfokine. Science 296, 1470 – 1472 18 Tang, W. et al. (2002) A cysteine-rich extracellular protein, LAT52, interacts with the extracellular domain of the pollen receptor kinase LePRK2. Plant Cell 14, 2277– 2287 19 Li, J. and Chory, J. (1997) A putative leucine-rich repeat receptor http://plants.trends.com 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 kinase involved in brassinosteroid signal transduction. Cell 90, 929– 938 Endre, G. et al. (2002) A receptor kinase gene regulating symbiotic nodule development. Nature 417, 962– 966 Bauer, Z. et al. (2001) Sensitivity of different ecotypes and mutants of Arabidopsis thaliana toward the bacterial elicitor flagellin correlates with the presence of receptor-binding sites. J. Biol. Chem. 276, 45669 – 45676 Meindl, T. et al. (2000) The bacterial elicitor flagellin activates its receptor in tomato cells according to the address – message concept. Plant Cell 12, 1783 – 1794 Schlessinger, J. (2002) Ligand-induced, receptor-mediated dimerization and activation of EGF receptor. Cell 110, 669– 672 Stein, R.A. et al. (2001) An analytical approach to the measurement of equilibrium binding constants: application to EGF binding to EGF receptors in intact cells measured by flow cytometry. Biochemistry 40, 6142– 6154 Kachroo, A. et al. (2001) Allele-specific receptor– ligand interactions in Brassica self-incompatibility. Science 293, 1824– 1826 Trotochaud, A.E. et al. (2000) CLAVATA3, a multimeric ligand for the CLAVATA1 receptor-kinase. Science 289, 613– 617 Wang, Z.Y. et al. (2001) BRI1 is a critical component of a plasmamembrane receptor for plant steroids. Nature 410, 380 – 383 Li, J. et al. (2001) BRS1, a serine carboxypeptidase, regulates BRI1 signaling in Arabidopsis thaliana. Proc. Natl. Acad. Sci. U. S. A. 98, 5916– 5921 He, Z. et al. (2000) Perception of brassinosteroids by the extracellular domain of the receptor kinase BRI1. Science 288, 2360 – 2363 Gomez-Gomez, L. et al. (2001) Both the extracellular leucine-rich repeat domain and the kinase activity of FSL2 are required for flagellin binding and signaling in Arabidopsis. Plant Cell 13, 1155 – 1163 Gomez-Gomez, L. and Boller, T. (2002) Flagellin perception: a paradigm for innate immunity. Trends Plant Sci. 7, 251 – 256 Trotochaud, A.E. et al. (1999) The CLAVATA1 receptor-like kinase requires CLAVATA3 for its assembly into a signaling complex that includes KAPP and a Rho-related protein. Plant Cell 11, 393– 406 Kayes, J.M. and Clark, S.E. (1998) CLAVATA2, a regulator of meristem and organ development in Arabidopsis. Development 125, 3843– 3851 Jeong, S. et al. (1999) The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell 11, 1925 – 1934 Giranton, J.L. et al. (2000) The integral membrane S-locus receptor kinase of Brassica has serine/threonine kinase activity in a membranous environment and spontaneously forms oligomers in planta. Proc. Natl. Acad. Sci. U. S. A. 97, 3759 – 3764 Gaude, T. and McCormick, S. (1999) Signaling in pollen – pistil interactions. Semin. Cell Dev. Biol. 10, 139 – 147 Johannessen, L.E. et al. (2001) Heterodimerization of the epidermalgrowth-factor (EGF) receptor and ErbB2 and the affinity of EGF binding are regulated by different mechanisms. Biochem. J. 356, 87 – 96 Wilkinson, J.C. and Staros, J.V. (2002) Effect of ErbB2 coexpression on the kinetic interactions of epidermal growth factor with its receptor in intact cells. Biochemistry 41, 8 – 14 Oh, M.H. et al. (2000) Recombinant brassinosteroid insensitive 1 receptor-like kinase autophosphorylates on serine and threonine residues and phosphorylates a conserved peptide motif in vitro. Plant Physiol. 124, 751 – 766 Clark, S.E. et al. (1997) The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell 89, 575– 585 Clouse, S.D. (2002) Brassinosteroid signal transduction: clarifying the pathway from ligand perception to gene expression. Mol. Cell 10, 973– 982 Cabrillac, D. et al. (2001) The S-locus receptor kinase is inhibited by thioredoxins and activated by pollen coat proteins. Nature 410, 220– 223 Friedrichsen, D.M. et al. (2000) Brassinosteroid-insensitive-1 is a ubiquitously expressed leucine-rich repeat receptor serine/threonine kinase. Plant Physiol. 123, 1247– 1256 Williams, R.W. et al. (1997) A possible role for kinase-associated protein phosphatase in the Arabidopsis CLAVATA1 signaling pathway. Proc. Natl. Acad. Sci. U. S. A. 94, 10467 – 10472 Review TRENDS in Plant Science 45 Braun, D.M. et al. (1997) Interaction of the maize and Arabidopsis kinase interaction domains with a subset of receptor-like protein kinases: implications for transmembrane signaling in plants. Plant J. 12, 83 – 95 46 Stone, J.M. et al. (1998) Control of meristem development by CLAVATA1 receptor kinase and kinase-associated protein phosphatase interactions. Plant Physiol. 117, 1217– 1225 47 Shah, K. et al. (2002) The Arabidopsis kinase-associated protein phosphatase controls internalization of the somatic embryogenesis receptor kinase 1. Genes Dev. 16, 1707– 1720 48 Aifa, S. et al. (2002) Interactions between the juxtamembrane domain of the EGFR and calmodulin measured by surface plasmon resonance. Cell. Signal. 14, 1005 49 Kurten, R.C. et al. (1996) Enhanced degradation of EGF receptors by a sorting nexin, SNX1. Science 272, 1008– 1010 50 Gu, T. et al. (1998) Binding of an arm repeat protein to the kinase domain of the S-locus receptor kinase. Proc. Natl. Acad. Sci. U. S. A. 95, 382 – 387 51 Stone, S.L. et al. (1999) A breakdown of Brassica self-incompatibility in ARC1 antisense transgenic plants. Science 286, 1729– 1731 52 Azevedo, C. et al. (2001) The U-box protein family in plants. Trends Plant Sci. 6, 354 – 358 53 Asai, T. et al. (2002) MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415, 977 – 983 54 Peck, S.C. et al. (2001) Directed proteomics identifies a plant-specific Vol.8 No.5 May 2003 55 56 57 58 59 60 61 62 237 protein rapidly phosphorylated in response to bacterial and fungal elicitors. Plant Cell 13, 1467 – 1475 Li, J. et al. (2001) BIN2, a new brassinosteroid-insensitive locus in Arabidopsis. Plant Physiol. 127, 14 – 22 Wang, Z.Y. et al. (2002) Nuclear-localized BZR1 mediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis. Dev. Cell 2, 505– 513 Yin, Y. et al. (2002) BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell 109, 181– 191 Schumacher, K. et al. (1999) The Arabidopsis det3 mutant reveals a central role for the vacuolar Hþ-ATPase in plant growth and development. Genes Dev. 13, 3259 – 3270 Sharma, A. et al. (2001) Antisense inhibition of a BRI1 receptor reveals additional protein kinase signaling components downstream to the perception of brassinosteroids in rice. FEBS Lett. 507, 346– 350 Montoya, T. et al. (2002) Cloning the tomato Curl3 gene highlights the putative dual role of the leucine-rich repeat receptor kinase tBRI1/SR160 in plant steroid hormone and peptide hormone signaling. Plant Cell 14, 3163 – 3176 Yu, L.P. et al. (2003) POLTERGEIST encodes a protein phosphatase 2C that regulates CLAVATA pathways controlling stem cell identity at Arabidopsis shoot and flower meristems. Curr. Biol. 13, 179 – 188 Takayama, S. et al. (2000) The pollen determinant of self-incompatibility in Brassica campestris. Proc. Natl. Acad. Sci. U. S. A. 97, 1920– 1925 Phloem 2003 International Meeting on Phloem Transport 2003 31 August–5 September 2003 Bayreuth, Germany TOPICS: – Source–sink relationships in nutrient allocation; phloem development and differentiation; phloem as a pathway for signal substances; phloem interactions with pathogens and herbivores. For more information contact: Professor Ewald Komor, Plant Physiology, University Bayreuth, D-95440 Bayreuth, Germany e-mail: [email protected] – http://www.phloem2003.de/ http://plants.trends.com