Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
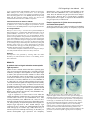
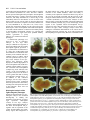
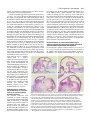
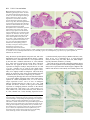
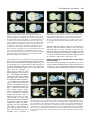
959 Development 124, 959-969 (1997) Printed in Great Britain © The Company of Biologists Limited 1997 DEV9525 Evidence that FGF8 signalling from the midbrain-hindbrain junction regulates growth and polarity in the developing midbrain Scott M. K. Lee1, Paul S. Danielian1, Bernd Fritzsch2 and Andrew P. McMahon1 1Department 2Department of Molecular and Cellular Biology, The Biolabs, Harvard University, 16 Divinity Avenue, Cambridge, MA 02138, USA of Biomedical Sciences, Creighton University, Omaha, Nebraska 68178, USA SUMMARY The developing vertebrate mesencephalon shows a rostrocaudal gradient in the expression of a number of molecular markers and in the cytoarchitectonic differentiation of the tectum, where cells cease proliferating and differentiate in a rostral to caudal progression. Tissue grafting experiments have implicated cell signalling by the mesencephalicmetencephalic (mid-hindbrain) junction (or isthmus) in orchestrating these events. We have explored the role of Wnt-1 and FGF8 signalling in the regulation of mesencephalic polarity. Wnt-1 is expressed in the caudal mesencephalon and Fgf8 in the most rostral metencephalon. Wnt1 regulates Fgf8 expression in the adjacent metencephalon, most likely via a secondary mesencephalic signal. Ectopic expression of Fgf8 in the mesencephalon is sufficient to activate expression of Engrailed-2 (En-2) and ELF-1, two genes normally expressed in a decreasing caudal to rostral gradient in the posterior mesencephalon. Ectopic expression of Engrailed-1 (En-1), a functionally equivalent homologue of En-2 is sufficient to activate ELF-1 expression by itself. These results indicate the existence of a molecular hierarchy in which FGF8 signalling establishes the graded expression of En-2 within the tectum. This in turn may act to specify other aspects of A-P polarity such as graded ELF-1 expression. Our studies also reveal that FGF8 is a potent mitogen within the mesencephalon: when ectopically expressed, neural precursors continue to proliferate and neurogenesis is prevented. Taken together our results suggest that FGF8 signalling from the isthmus has a key role in coordinately regulating growth and polarity in the developing mesencephalon. INTRODUCTION The vertebrate mesencephalon exhibits many aspects of rostrocaudal polarity. Anatomically there is a pronounced rostrocaudal developmental gradient of cytoarchitectonic differentiation in the chick tectum (dorsal mesencephalon) (LaVail and Cowan, 1971) which appears to be present in the dorsal mesencephalon (colliculi) (Altman and Bayer, 1981a,b) of mammals as well (for simplicity we will use the general term ‘tectum’ in reference to the chick tectum and the mammalian colliculi). As a consequence, neural precursors cease cell division and differentiate first in the most rostral tectum, while precursors in more caudal regions, close to the isthmus, are still dividing and expanding. Additionally, the rostral and caudal regions of the tectum acquire different sets of afferent inputs from the retina: axons extending from ganglion cells in the temporal retina innervate the rostral tectum, while the caudal tectum receives axons from the nasal retina (DeLong and Coulombre, 1965; Crossland et al., 1974). Clues as to the molecular regulation of mesencephalic polarity are provided by the observations that many molecules are expressed in gradients across the tectum. First En-2, a transcription factor, is expressed in a rostrocaudally increasing gradient across the tectum. (Davis and Joyner, 1988; Davis et al., 1988; Gardner et al., 1988; Patel et al., 1989; Gardner and Barald, 1992; Millet and Alvarado-Mallart, 1995). Addition- While vertical and planar signals which occur at gastrulation impart an initial anterior-posterior (A-P) pattern onto the central nervous system (CNS) subsequent interactions between adjacent cell populations within the neural plate are required to refine this pattern (Bally-Cuif and Wassef, 1995; Kelly and Melton, 1995). The role of cellular interactions between adjacent regions in the establishment of axial pattern has been intensively studied in the developing brain. For example, interspecies grafting experiments between quail and chick embryos have revealed that the mesencephalic-metencephalic junction (hereafter refered to as the isthmus) acts as an organizing center (Bally-Cuif and Wassef, 1995; Joyner, 1996). Grafts of isthmic tissue into the caudal diencephalon induce the formation of ectopic mesencephalic structures in the host (Gardner and Barald, 1991; Itasaki et al., 1991; Martinez et al., 1991; BallyCuif et al., 1992; Marin and Puelles, 1994), whereas grafts into the myelencephalon induce ectopic cerebellar structures (Martinez et al., 1995). These results suggest that signals from the isthmus may normally be involved in the induction of mesencephalic cell fates rostrally and cerebellar fates caudally. Cell interactions somewhat later in development may also regulate rostrocaudal polarity within a specific brain region. Key words: mesencephalon, polarity, Fgf8, Engrailed, ELF-1, Wnt-1, cell proliferation, pattern formation, CNS 960 S. M. K. Lee and others ally, ELF-1 and RAGS, two ligands for EPH-like receptor tyrosine kinases are expressed in rostrocaudally increasing gradients across the caudal tectum and may function to inhibit temporal axon ingrowth and/or to attract nasal axons (Cheng and Flanagan, 1994; Cheng et al., 1995; Drescher et al., 1995; Nakamoto et al., 1996). Ectopic high level expression of En genes in the rostral tectum leads to a perturbation of retinal axon guidance such that temporal axons rarely enter the tectum and nasal axons often arborize ectopically (Friedman and O’Leary, 1996; Itasaki and Nakamura, 1996; Logan et al., 1996). A molecular basis for this result was provided by the observations that ELF-1 and RAGS are ectopically expressed in these experiments (Logan et al., 1996) and that ectopic expression of ELF-1 alone is sufficient to inhibit ingrowth of temporal axons into the tectum (Nakamoto et al., 1996). Ectopic high level rostral expression of En also abolishes the rostrocaudal gradient of cytoarchitectonic development (Logan et al., 1996). These results support a model in which the graded expression of En genes across the rostrocaudal axis of the tectum is involved in conferring positional specification to these cells. A role for graded En expression in determining tectal polarity is further supported by grafting experiments in the chick. If the rostral mesencephalon is inverted, En-2 expression is reprogrammed in the grafted tissue to form a gradient consistent with its new environment (Martinez and AlvaradoMallart, 1990; Itasaki et al., 1991). Subsequently, a normal mesencephalon develops from the grafted tissue (Matsuno et al., 1991), demonstrating that mesencephalic rostrocaudal polarity is plastic at the time of the experiment (2 days of incubation) and implicating signals from adjacent tissue in mesencephalic rostrocaudal patterning and En-2 regulation. Additionally, ectopic mesencephalic structures resulting from isthmic grafts into the diencephalon display both an En-2 gradient and anatomical polarity which are the mirror image of those in the endogenous mesencephalon (Alvarado-Mallart et al., 1990; Itasaki and Nakamura, 1992; Itasaki et al., 1991; Martinez et al., 1991; Marin and Puelles, 1994). Together, these data implicate isthmic derived signals in the establishment of mesencephalic rostrocaudal polarity and imply that such signals may act via the regulation of En expression. Recent experiments have demonstrated the importance of FGF8 in mesencephalic development. FGF8 is expressed in cardiac mesoderm underlying the presumptive mes/met region (Heikinheimo et al., 1994; Crossley and Martin, 1995; Mahmood et al., 1995). Application of beads soaked in FGF8 results in ectopic mesencephalic development suggesting that FGF8 may normally play a role in mesencephalic induction (Crossley et al., 1996a) (see Discussion). Fgf8 is also expressed in the neural plate itself, in the rostral metencephalon, from 3somites (Mahmood et al., 1995), with expression refining to a tight band of cells immediately caudal to the isthmus by 16somites (Heikinheimo et al., 1994; Crossley and Martin, 1995; Mahmood et al., 1995; note that the region of isthmus used in the above mentioned grafting experiments includes Fgf8expressing cells). Thus FGF8 may also play a role in regulating the rostrocaudal polarity of the mesencephalon. We have explored the role of cell signalling at the isthmus in establishing rostrocaudal polarity in the mesencephalon. By analyzing Wnt-1 mutants we show that a mesencephalic derived signal(s) is required to maintain Fgf8 expression at the isthmus. To address the possibility that FGF8 regulates mesencephalic polarity we have used the Wnt-1 enhancer to generate a new domain of Fgf8 expression at the dorsal midline of the mesencephalon. Our results are consistent with FGF8 signalling playing a central role in regulating growth and polarity in the developing mesencephalon. MATERIALS AND METHODS Production and genotyping of Wnt-1 mutant embryos Wnt-1 mutant embryos were obtained by intercrossing mice heterozygous for a Wnt-1 null mutation (McMahon and Bradley, 1990). Yolk sac DNA was used for genotyping as described by McMahon et al. (1992). Fgf8 misexpression construct To ectopically express Fgf8 under the control of the Wnt-1 enhancer we utilized a variant of the previously described pWEXPZ vector (Danielian and McMahon, 1996), here designated pWEXPZ2. Each contains a Wnt-1 minigene upstream of the 5.5 kb 3-prime located Wnt-1 enhancer sequences. In pWEXPZ2 an approximately 800 base pair region in the Wnt-1 3-prime UTR has been deleted. This has no effect on the previously reported expression pattern regulated by the Wnt-1 enhancer (Echelard et al., 1993, 1994). The WEXPZ2-Fgf8 construct was obtained by cutting pWEXPZ2 with EcoRV and inserting an end filled EcoRI-XhoI cut piece of the Fgf8 cDNA (Mahmood et al., 1995). This cDNA corresponds to Fgf8 variant 4 of Crossley and Martin (1995). Production of Fgf8 transgenic embryos Transgenic mice were generated essentially as described by Echelard et al. (1993). Linearized DNA was purified by agarose gel electrophoresis followed by electroelution into 1× TAE and concentrated using the Wizard DNA Cleanup System (Promega). C57BL/6J×CBA/J F1 mice were used as embryo donors. Injected embryos were cultured overnight in M-16 medium following microinjection, and transferred at the 2-cell stage to pseudopregnant recipient SWB females. Noon of the day of the transfer was considered to be 0.5 d.p.c. For some experiments, pregnant females were injected intraperitoneally with 50 µg/g body weight of 5-bromo-2′ deoxyuridine (BrdU; Sigma) 1.5 hours prior to killing. Staged embryos were collected at 10.5 d.p.c., 12.5 d.p.c., 13.5 d.p.c., 14.5 d.p.c., and 18.5 d.p.c. into PBS and photographed using an Olympus SZH stereo photomicroscope on Kodak EPY 64T color slide film. The genotype of embryos was determined by PCR analysis of genomic DNA extracted from embryonic yolk sacs with care taken to avoid cross-contamination. The primers used were directed against the lacZ sequences present in the transgene construct. 5′ primer: 5′-TTTAACGCCGTGCGCTGTTCG-3′ 3′ primer: 5′-GATCCAGCGATACAGCGCGTC-3′ The PCR program used was as follows: cycle 1: 94°C, 3 minutes; cycle 2: 94°C, 45 seconds; cycle 3: 55°C, 45 seconds; cycle 4: 72°C, 1 minute; cycle 5: 35 times to cycle 2; cycle 6: 72°C, 5 minutes. PCR products were visualized on ethidium bromide stained agarose gels. Of a total of 366 embryos obtained between 10.5 d.p.c. and 18.5 d.p.c., 126 (34.4%) were transgenic. Of these 68 (54.0%) showed the reported brain phenotype. Histological analysis Embryos were fixed in 4% paraformaldehyde in PBS for 18-24 hours at 4°C while 18.5 d.p.c. brains were fixed for 18-24 hours in Bouin’s fixative at room temperature. Paraformaldehyde-fixed tissue was transferred into 70% ethanol and stored at 4°C. Bouin’s-fixed tissue was washed extensively in 70% ethanol for 2-4 days with subsequent storage at 4°C. For embedding, samples were dehydrated through a FGF8 signalling in the midbrain series of graded ethanols and embedded in paraffin wax (Fibrowax, BDH). Embryos were serially sectioned at 6-7 µm in the sagittal plane. Representative sections were stained with haemotoxylin and eosin and photographed on a Leitz DMRD photomicroscope using Kodak 64T film. Selected remaining sections were used for the immunohistochemical detection of BrdU incorporation. Immunohistochemical detection of BrdU Immunohistochemical detection of BrdU was performed as described by Dickinson et al. (1994). Photographs of representative sections were taken on a Leitz DMRD photomicroscope using Kodak 64T film. In situ hybridization Embryos at 10.5-13.5 d.p.c. were fixed in 4% paraformaldehyde in PBS overnight at 4°C, washed in PBS, dehydrated through a series of graded methanols, and stored at −20°C. For 12.5 and 13.5 d.p.c. embryo brains were dissected prior to in situ hybridization. Wholemount in situ hybridization was performed as described by Parr et al. (1993) and modified according to Knecht et al. (1995). Digoxigenin probes were synthesized using the Digoxigenin RNA Labeling Kit (Boehringer Mannheim). ELF-1 expression was detected by the RAP in situ technique using a Mek4-AP probe as described by Cheng and Flanagan (1994); Cheng et al. (1995). Stained embryos and brains were transferred to 80% glycerol and photographed using a Leica Wild M10 photomicroscope using Kodak 64T film. For all markers analyzed a minimum of three transgenic embryos were analyzed and identical results where always observed. 961 abolished (Fig. 1E,F), even though the metencephalon is still intact at this time (McMahon et al., 1992; Serbedzija et al., 1996). Thus, the initiation of Fgf8 expression occurs independently of Wnt-1. However, Wnt-1, or more likely a secondary mesencephalic derived signal, is required for the maintenance of Fgf8 expression (see Discussion). Ectopic expression of Fgf8 in the mesencephalon and caudal diencephalon To directly explore the role of FGF8 signalling in establishing mesencephalic rostrocaudal polarity we ectopically expressed Graphics All composites were generated by scanning slides into Adobe Photoshop v3.0, and transferred to Canvas v3.5 for labeling, all on a Power Macintosh. RESULTS A midbrain derived signal maintains metencephalic Fgf8 expression Studies of Wnt-1 mutants indicate that Wnt-1 signalling plays a key role in mes/met development. Wnt-1 is broadly expressed within the presumptive mesencephalon at neural plate stages (1-somite), but expression becomes refined to a narrow band of cells immediately anterior to the isthmus, as well as much of the dorsal central nervous system (CNS) following neural tube closure (16-somites; Wilkinson et al., 1987; McMahon et al., 1992; Parr et al., 1993). In embryos homozygous for a Wnt1 null mutation mesencephalic development proceeds normally up to the 5-somite stage, but by 14 somites most of the mesencephalon is absent. The metencephalon is lost subsequently, most likely reflecting a requirement of mesencephalic derived signals for metencephalic development (McMahon and Bradley, 1990; Thomas and Capecchi, 1990; McMahon et al., 1992; Serbedzija et al., 1996). Fgf8 is expressed in a narrow band of cells within the isthmus immediately caudal to the Wnt-1-expressing cells from 3-somites until at least 14.5 d.p.c.(Heikinheimo et al., 1994; Crossley and Martin, 1995; Mahmood et al., 1995). To investigate the possibility that Fgf8 expression is dependent on Wnt1 or, more generally, on any mesencephalic derived signals, we analyzed Fgf8 expression in Wnt-1 mutant embryos. Fgf8 expression in Wnt-1 mutants was indistinguishable from that in the wild type up to 6-somites (Fig. 1A,B). However, by 9somites expression is markedly reduced (Fig. 1C,D) and by 14somites metencephalic Fgf8 expression was completely Fig. 1. Fgf8 expression in Wnt-1 mutant embryos. (A,B) Fgf8 expression appears identical in the mes-met region of wild-type (A) and mutant (B) embryos at the 6-somite stage, indicating that Wnt-1 is not required for activation of Fgf8 expression. (C,D) Comparison of wild-type (C) and mutant (D) 9-somite embryos reveals that by this time mesencephalon development is reduced in Wnt-1 mutants. Fgf8 expression in the metencephalon (arrow) is also markedly reduced. (E) By 15 somites Fgf8 expression in wild-type embryos is restricted to a narrow band of cells immediately posterior to the isthmus. (F) In a 14-somite mutant embryo the metencephalon is still present (arrow) but metencephalic Fgf8 expression is completely abolished. 962 S. M. K. Lee and others Fgf8 in the rostral mesencephalon of mouse embryos using the Wnt-1 enhancer (Echelard et al., 1994). The Wnt-1 enhancer directs expression throughout the presumptive mesencephalon at neural plate stages (1-somite). Expression rapidly becomes refined to, and stably maintained in, a narrow ring of cells immediately anterior to the isthmus as well as much of the dorsal central nervous system (CNS) by the time that neural tube closure in the brain is complete (16-somites; Wilkinson et al., 1987; McMahon et al., 1992; Parr et al., 1993). Consequently, this expands the domain of Fgf8 expression into the dorsal aspect of the mesencephalon and caudal diencephalon coincident with the initiation of a distinct rostrocaudal polarity in mesencephalic development. Because we expected that ectopic expression of Fgf8 would cause postnatal lethality, founder (generation 0 [G0]) embryos were examined for aberrant phenotypes. A characteristic phenotype was observed in 54% of transgenic embryos. At 10.5 d.p.c. these embryos exhibited an overgrowth of the mesencephalon and most caudal diencephalic segment, prosomere 1 (p1) (nomenclature of Rubenstein et al., 1994), (Fig. 2A,B) together with the formation of a constriction at the boundary of the midbrain and diencephalon (arrows in 2B,D), which was more pronounced by 12.5 d.p.c. (Fig. 2C,D). Analysis at 18.5 d.p.c. showed that transgenic brains displayed a dramatic hyperplasia of the mesencephalon and caudal diencephalon which partially overgrew the cerebellum and telencephalon (compare Fig. 2E,G with F,H). The volume of the mesencephalon of transgenic embryos was as much as 3 times greater than that of sibling non-transgenic controls (Figs 2, 4). In addition to this consistent phenotype, four 10.5 d.p.c. embryos appeared to have more widespread and non-specific defects most likely resulting from widespread activation of the transgene. These were not studied further. Histological analysis of the Fgf8 transgenic phenotype Sagittal sections through 12.5 d.p.c. transgenic embryos were examined histologically to identify the anatomical consequences of ectopic Fgf8 expression. Mid-sagittal sections of 12.5 d.p.c. embryos revealed a rostrocaudal expansion of the dorsal mesencephalon and p1, together with an expansion of the ventral mesencephalon and mammillary region, and enlargement of the third ventricle (Fig. 3A,B). We also observed an apparent reduction in the medial thalamus, which most likely is secondary to the expansion of the third ventricle (Fig. 3B). In all cases the overgrowth correlated with the presence of the transgene and the characterized expression pattern of the Wnt1 enhancer (Echelard et al., 1993, 1994). Analysis of more lateral regions revealed the remainder of the diencephalon had formed essentially normally (Fig. 3C,D). Interestingly, lateral regions of the expanded rostral mesencephalon and p1 consisted of only a thin ventricular zone and very few differentiated cell types, unlike wild-type embryos which have undergone considerable differentiation in the rostral mesencephalon by this time (compare Fig. 3C,D). This suggests that the morphological expansion may therefore result from a main- Fig. 2. Views of WEXPZ2-Fgf8 transgenic embryos and brains revealing an overgrowth of the mesencephalon and caudal diencephalon. (A-D) Lateral views of wild-type (A,C) and transgenic (B,D) embryos at 10.5 d.p.c. (A,B) and 12.5 d.p.c. (C,D). The mesencephalon and most caudal diencephalic segment, p1, are expanded in the transgenics, leading to a large increase in the flexion of the brain by 12.5 d. p. c. (compare C and D). Additionally, an ectopic constriction is present between the rostral mesencephalon and p1 (arrows in B and D). (E-H) Lateral (E,F) and dorsal (G,H) views of 18.5 d.p.c. wild-type (E,G) and transgenic (F,H) brains in which the mesencephalon-p1 region is overgrown. The expansion in the mediolateral dimension is greater rostrally than caudally, probably due the fact that the Wnt-1 enhancer drives expression of Fgf8 to a broad dorsal region of p1 and only to a very narrow dorsally restricted band of cells in the mesencephalon. cb, cerebellum; ch, cerebral cortex; i, isthmus; ms, mesencephalon; mt, metencephalon; my, myelencephalon; p1, prosomere 1 (pretectum); t, telencephalon. FGF8 signalling in the midbrain tenance of proliferative ventricular zone cells and a concomitant reduction in neuronal differentiation. Consistent with this suggestion, expression of Fgf8 leads, by 18.5 d.p.c., to a severe reduction of differentiated cell types in the dorsal mesencephalon and caudal diencephalon and an even greater expansion of the third ventricle than that observed at 12.5 d.p.c. (Fig. 4A,B). As FGF8-soaked beads have been shown to induce the formation of an ectopic mesencephalon in the caudal diencephalon of chick embryos (Crossley et al., 1996a), we examined the diencephalon for evidence of mesencephalic transformation. In such experiments the ectopic mesencephalon forms at the expense of the dorsal thalamus and pretectum of the host, and duplicated cranial nerves III and IV are formed ventrally. In our transgenics the pretectum is significantly overgrown but both the dorsal and ventral parts of the thalamus are present and the habenulopeduncular tract has formed close to its normal location (Fig. 4C,D). Additionally, a detailed histological analysis of four 18.5 d.p.c. trangenic brains revealed no evidence for duplications of mesencephalic cranial nerves III or IV (Fig. 4 and data not shown). This was further confirmed by placing DiI into the orbit of 14.5 d.p.c. transgenic embryos. While we were consistently able to label the oculomotor, trochlear and abducens projections to their normal locations we never observed any ectopic projections to more rostral positions (data not shown). Based on these observations and molecular analysis (see below) we conclude that the continued ectopic expression of Fgf8 using the Wnt-1 regulatory element leads to a massive expansion of neural precursor cells at the expense of differentiated cell types in both mesencephalon and caudal diencephalon, but is not sufficient to induce the formation of ectopic mesencephalic structures. The differences between our results and the bead experiments may reflect variability in the experimental paradigm, species differences in the competence to respond to FGF8 at different developmental stages, or alternatively may represent differences in the activities of FGF8 isoforms (see Materials and Methods, Crossley et al., 1996a, and MacArthur et al. 1995a,b). Fgf8 expression enhances the proliferation of precursor cells in the mesencephalon and diencephalon The observed ventricular zone expansion in regions expressing Fgf8 suggests that FGF8 signalling leads to enhanced or prolonged mitotic activity in these regions. To investigate this issue further we directly assayed cell proliferation by monitoring the incorporation of bromodeoxyuridine (BrdU) at 14.5 d.p.c. In the mesencephalon of wild 963 type embryos only the most caudal regions of the tectum, close to the isthmus, retain a thick ventricular zone with a large number of dividing precursor cells (Fig. 5A,C). Note that in this region almost the entire neuroepithelium consists of mitotically active ventricular zone cells and few differentiated cell types are present (Fig. 5C). More rostrally the converse is true, there are very few mitotically active precursor cells and a thick layer of differentiated cell types has formed (Fig. 5E). In contrast, transgenic embryos have a thick layer of mitotically active precursor cells along the entire rostrocaudal extent of the expanded midbrain-p1 ventricle (Fig. 5B,D,F). A corresponding reduction in the number of differentiated cell types is also observed (compare Fig. 5E with F). The expanded mammillary recess of the third ventricle present in transgenic brains also contains a layer of mitotically active cells (data not shown). In summary, we observed a strict correlation between ectopic Fgf8 expression, increased neural precursor cell proliferation, and a reduction of neural differentiation. Characterization of rostrocaudal patterning in embryos ectopically expressing Fgf8 in the rostral mesencephalon and caudal diencephalon In all transgenic embryos analyzed between 10.5 and 13.5 d.p.c. Fgf8 is ectopically expressed in a ring of cells rostral to the isthmus, along the dorsal and ventral midlines of the mesencephalon, in a dorsally restricted transverse stripe in p1, Fig. 3. Histological analysis of 12.5 d.p.c. WEXPZ2-Fgf8 transgenic embryos. Sections were stained with haematoxylin and eosin. A and C are sections through the wild-type embryo shown in Fig. 2C, A is mid-sagittal and C more lateral. B and D are sections through the transgenic embryo shown in Fig. 2D, B is mid-sagittal and D is more lateral. Mid-sagittal sections reveal a rostrocaudal expansion of the dorsal mesencephalon and most caudal diencephalic segment, prosomere 1 (p1), together with an expansion of the ventral mesencephalon and mammillary region, and enlargement of the third ventricle (compare A and B). An apparent reduction in the medial thalamus is most likely secondary to the expansion of the third ventricle. In all cases, the overgrowth correlates with expression of the transgene. Analysis of more lateral regions reveals that the remainder of the diencephalon formed essentially normally (compare C and D), consistent with the fact that transgene expression is largely limited to medial regions. cb, cerebellum; mr, mammilary region; ms, mesencephalon; p, pituitary gland; my, myelencephalon; p1, prosomere 1; t, telencephalon; th, thalamus; III, third ventricle. 964 S. M. K. Lee and others Fig. 4. Histological analysis of 18.5 d.p.c. WEXPZ2-Fgf8 transgenic brains. Sections were stained with haematoxylin and eosin. A and C are sections through the wild-type embryo shown in Fig. 2E,G, A is midsagittal and C more lateral. B and D are sections through the transgenic embryo shown in Fig. 2F,G, B is mid-sagittal and D is more lateral. Transgenic brains display a reduction in differentiated cell types in the dorsal mesencephalon and caudal diencephalon and a concomitant expansion in the number of precursor cells, which leads to a large expansion of the effected regions tangential to the ventricular surface (compare A,C with B,D). In some regions, which are likely expressing Fgf8, the roof of the mesencephalon is only a few cell layers thick (* in B). Additionally, the third ventricle is expanded as observed at 12.5 d.p.c. (B). Note that while the thalamus exhibits some dismorphology, likely a secondary consequence of the expanded third ventricle, it appears to be anatomically rather normal and the habenulopeduncular tract is present (compare C and D). Transgenic brains also display a consistent reduction in the cerebellum. cb, cerebellum; ch, cerebral cortex; ht, habendopenduncular tract; mr, mammilary region; ms, mesencephalon; pt, pretectum; my, myelencephalon; th, thalamus; III, third ventricle. along the dorsal myelencephalon and spinal cord, and in the mammillary region (Fig. 6A,B and data not shown), a pattern identical to Wnt-1 itself and to that of a lacZ reporter driven by the same Wnt-1 enhancer (Echelard et al., 1993; Echelard et al., 1994). We never observed an ectopic Wnt-1 expression domain nor any apparent increase in the levels of Wnt-1 expression in transgenic embryos analyzed between 10.5 and 13.5 d.p.c. (compare Fig. 6C with D). The dorsal mesencephalic stripe of Wnt-1 expression however appears consistently to be slightly broader in transgenics. This most likely reflects an expansion of the entire mesencephalon along the mediolateral axis by this time (compare Fig. 6C with D). The transgenic phenotype does not result from the formation of an ectopic isthmus organizing center in the rostral mesencephalon or caudal diencephalon. First, we never observed ectopic expression of En-1, Pax-2, or Pax-5 in transgenic embryos examined between 10.5 and 13.5 d.p.c. (Fig. 7A,B and data not shown). All three of these genes are normally expressed in overlapping domains centered on the isthmus (Davis and Joyner, 1988; Davis et al., 1988; Nornes et al., 1990; Adams et al., 1992; Asano and Gruss, 1992; Puschel et al., 1992; Urbanek et al., 1994). Secondly, expression of PaxFig. 5. BrdU incorporation in transgenic brains at 14.5 d.p.c. A is a section through a mid-sagittal region of a wild-type control and B is a section through a corresponding mid-sagittal region of a transgenic embryo. C,D,E,F are higher magnification views of the regions indicated in A and B. Cells that have incorporated BrdU appear black and they are the only stained cells in these photomicrographs. In the wild-type mesencephalon (A) high mitotic activity is limited to the most caudal region (C) and few mitotic cells are observed rostrally (E). In contrast, the expanded dorsal mesencephalon and p1 of transgenic brains contains a thick ventricular zone of mitotically active cells along its entire rostrocaudal extent (B,D,F). 6, which is normally expressed in p1 (Walther and Gruss, 1991; arrow in Fig. 7C) is maintained in p1 of Fgf8 transgenic embryos (arrow in Fig. 7D). Thus, these cells, while morphologically abnormal, maintain a p1 identity. Our results are more consistent with FGF8 signalling playing a later role in the regulation of rostrocaudal polarity. In wild type embryos En-2 expression in the mes/met region is highest at the isthmus and decreases both rostrally and caudally (Davis and FGF8 signalling in the midbrain Fig. 6. Fgf8 and Wnt-1 expression in the mesencephalon of transgenic brains. Whole-mount in situ hybridization analysis of Fgf8 (A,B) or Wnt-1 (C,D) expression in isolated 13.5 d.p.c. brains. (A,B) The WEXPZ2-Fgf8 transgene drives expression of Fgf8 along the dorsal midline of the mesencephalon, in a broad dorsal domain of p1, and into the ventral mesencephalon (which is not visible in this view; B) whereas endogenous Fgf8 expression is only detected at the mes-met junction at this time (A). Comparison of Wnt-1 expression in wild-type (C) and transgenic (D) brains reveals that Wnt-1 is not ectopically activated by Fgf8. The dorsal mesencephalic stripe of Wnt-1 expression in transgenics (D) may be broader than in the wildtype (C), but this is likely due to a mediolateral expansion of the entire mesencephalon, and not ectopic Wnt-1 activation. Accordingly, the WEXPZ2-Fgf8 expression domain is also consistently broader (B) than the wild-type Wnt-1 domain (C). 965 Fig. 7. Analysis of En-1 and Pax-6 expression in transgenic brains. Isolated brains of (A,B) 12.5 d.p.c. (En-1) or (C,D) 13.5 d.p.c. (Pax6) were stained as whole mounts. En-1 expression is limited to the caudal midbrain in both wild-type (A) and transgenic (B) brains. Pax-6 is expressed in the dorsal part of p1 in wild-type brains (arrow in C) and this expression domain is maintained in transgenic brains (arrow in D). contained within the domain of ectopic En-2 expression (Fig. 8B,C,E,F). Note that scattered clumps of En-2 and ELF-1 expressing cells are always observed in dorsal p1 (Fig. 8B,C,E,F). These may be mislocated mesencephalic cells which crossed the disrupted mesencephalon-p1 boundary. However, we can not rule out the possibility that FGF8 signalling induced En-2 and ELF-1 expression in a subset of p1 cells. Ectopic En expression activates ELF-1 in the rostral Joyner, 1988; Davis et al., 1988), however En-2 expression was mesencephalon observed in a very broad dorsal domain all along the rostrocauIn the chick tectum ectopic high level expression of En-1 or Endal axis of the mesencephalon of transgenic embryos (Fig. 8A2 in the rostral mesencephalon leads to ectopic activation of ELFC). Interestingly, En-2 is not activated by ectopic expression of 1 (Logan et al., 1996). To test whether FGF8 activation of ELFFgf8 in the dorsal diencephalon (see below), but is activated in 1 in our transgenics is mediated by Engrailed genes we examined the dorsal myelencephalon (white arrows in Fig. 8B,C). This argues that the competence to express En-2 in response to dorsal FGF8 signalling has a rostral limit at the mesencephalon-diencephalon boundary, and that FGF8 signalling can regulate En-2 expression in the mesencephalon and metencephalon. To investigate the possibility that Fgf8 is a more general regulator of posterior mesencephalon development we examined the expression of ELF-1, a ligand for the Mek-4 and Sek-1 receptor tyrosine kinases which is implicated in the regulation of the retinal-tectal connection (Cheng and Flanagan, 1994). In transgenic brains, ELF-1 is ectopically activated in a broad stripe along the dorsal mesencephalon and Fig. 8. En-2 and ELF-1 are ectopically expressed in the rostral mesencephalon of transgenics. Isolated 12.5 d.p.c. brains were stained as whole mounts using either in situ hybridization to detect in a transverse stripe at the mesen- En-2 or the RAP in situ technique to detect ELF-1. In wild-type brains mesencephalic expression of cephalon-p1 boundary (Fig. 8D-F), En-2 (A) and ELF-1 (D) is limited to caudal regions. In contrast, En-2 is expressed throughout the suggesting that all mesencephalic alar region of the entire mesencephalon in transgenic brains (B,C). Similarly, ELF-1 is ectopically cells within a certain distance of expressed in a very broad dorsal domain along the entire rostrocaudal extent of the mesencephalon FGF8 signal are induced to express and in a transverse stripe (limited to alar regions) in the rostral mesencephalon of transgenic brains ELF-1. The domain of ectopic mes- (E,F). Note that clumps of En-2- and ELF-1- (black arrows in B,D) expressing cells are also present encephalic ELF-1 expression is in dorsal p1. 966 S. M. K. Lee and others Fig. 9. Ectopic expression of En-1 activates ectopic expression of ELF-1. In a transgenic mouse line which expressed En-1 under the control of the Wnt-1 enhancer En-1 is ectopically expressed in the dorsal mesencephalon and p1 at 12.5 d.p.c. (A, and compare with Fig. 7A). ELF-1 expression is ectopically activated along the dorsal midline of the entire mesencephalon of transgenic brains from this line but not in dorsal p1 (B). The arrow in B indicates the rostral limit of high level dorsal ELF-1 expression in wild-type brains (compare with Fig. 8D). ELF-1 expression in transgenic mice expressing En-1 (En-1 and En-2 are functionally interchangeable; Hanks et al., 1995) under the control of the Wnt-1 enhancer. Transgenic embryos from this line express En-1 in the typical Wnt-1 expression pattern (Danielian and McMahon, 1996; Fig. 9A). In these transgenic embryos ELF-1 expression was ectopically activated in a narrow strip of cells along the dorsal midline of the anterior mesencephalon (Fig. 9B), most likely in only those cells that also express En-1 (compare Fig. 9A with B). Interestingly, En-1 expression in the dorsal diencephalon was not sufficient to activate ELF-1 expression in this region. Since Fgf8 expression is normal in this line of mice (Danielian and McMahon, 1996, data not shown) ectopic En expression is sufficient to activate expression of ELF-1 independently of FGF8 signalling. DISCUSSION We have provided evidence that an interaction between mesencephalic and metencephalic cells is required for the localized expression of Fgf8 at the isthmus. Continuous ectopic expression of FGF8 along the dorsal mesencephalon results in a posteriorization of much of the tectum. These results imply that the localized production of FGF8 signal at the isthmus serves to regulate many aspects of rostrocaudal polarity across the tectum. FGF8 signalling regulates rostrocaudal polarity in the mesencephalon The normal expression of En genes in a rostrocaudally increasing gradient across the mesencephalon (Davis and Joyner, 1988; Davis et al., 1988; Gardner et al., 1988; Patel et al., 1989; Gardner and Barald, 1992; Millet and Alvarado-Mallart, 1995) and the ability of ectopic high level En expression to caudalize the rostral tectum (Friedman and O’Leary, 1996; Itasaki and Nakamura, 1996; Logan et al., 1996) implies that En expression level acts to confer positional specification on cells along the rostrocaudal axis of the tectum. In our experiments the ectopic expression of Fgf8 along the dorsal midline of the mesencephalon leads to ectopic activation of En-2 expression in a broad dorsal domain. This implies that the localized production of FGF8 at the isthmus is normally involved in establishing the graded pattern of En-2 expression across the mesencephalon and thus in regulating the rostrocaudal specific expression of En-2 target genes. Indeed, in normal development Fgf8 expression is initiated just prior to the onset of En-2 expression (Davis and Joyner, 1988; Davis et al., 1988; Heikinheimo et al., 1994; Crossley and Martin, 1995; Mahmood et al., 1995) consistent with FGF8 activating En-2 expression. A putative En target, ELF-1, is indeed activated in the rostral mesencephalon of Fgf8 transgenic embryos. The domain of ectopic ELF-1 expression is always contained within the domain of ectopic En-2 expression and the ectopic expression of En-1 (En-1 and En-2 are functionally interchangeable) is sufficient to activate ectopic ELF-1 expression in the mesencephalon. The simplest interpretation of these results is that FGF8 signalling indirectly regulates ELF-1 expression via regulation of En-2. Together with the recently published observation that ectopic expression of En-1 in the rostral tectum of the chick is sufficient to activate ectopic expression of ELF-1 (Logan et al., 1996), these results imply that Engrailed proteins act additively to generate the graded expression of ELF-1 in the tectum. Normal En-2 expression in the metencephalon is the mirror image of mesencephalic En-2 expression. Expression is highest rostrally and forms a caudally decreasing gradient (Davis and Joyner, 1988; Davis et al., 1988; Gardner et al., 1988; Patel et al., 1989; Gardner and Barald, 1992; Millet and AlvaradoMallart, 1995). Our results indicate that expression of Fgf8 in the dorsal myelencephalon is sufficient to activate expression of En-2 in this brain region as well. This result is reminiscent of the ability of isthmus (Fgf8 expressing tissue) grafted into the myelencephalon to induce En-2 expression in adjacent host tissue (Martinez et al., 1995). This implies that FGF8 signalling from the isthmus may act on both rostrally and caudally adjacent cells of the mes/met region to establish a broad domain of graded En-2 expression. It should be pointed out that while our gain-of-function experiments strongly implicate En-2 in the regulation of mesencephalic rostrocaudal polarity, En-2 mutant mice seem to have a normal mesencephalon and display only minor defects in cerebellar foliation (Millen et al., 1994; Millen et al., 1995). A likely explanation is provided by the fact that En-1 and En2 are expressed in overlapping domains centered on the isthmus such that the domain of En-2 expression is only slightly broader than the domain of En-1 expression (Davis and Joyner, 1988; Davis et al., 1988; McMahon et al., 1992). Additionally, En-1 expression initiates at 1-somite, prior to initial expression of En-2 at 5-somites. Therefore, En-1 and En-2 are likely to serve redundant functions, an idea supported by rescue of the En-1 mutant phenotype by expression of En-2 under the control of the En-1 promoter (Hanks et al., 1995). Thus it is likely to be the additive concentration of En protein a tectal cell expresses which specifies its rostrocaudal position. Interestingly, we have never observed the ectopic expression of En-1 in Fgf8 transgenic embryos, suggesting the existence of a distinct mechanism which is responsible for establishing the rostrocaudal domian of En-1 expression. Consistent with this explanation, normal activation of En-1 (Davis and Joyner, 1988; McMahon et al., 1992) precedes the onset of Fgf8 expression in the brain (Mahmood et al., 1995). FGF8 mediated regulation of neural proliferation and neurogenisis An important aspect of tectal polarity is the rostrocaudal FGF8 signalling in the midbrain gradient of cytodifferentiation (LaVail and Cowan, 1971; Altman and Bayer, 1981a,b). Rostral regions differentiate first, have a greater overall thickness, and manifest a more advanced laminar structure than caudal regions, which retain mitotic activity in neural precursors late in development. Our studies show that ectopic expression of Fgf8 in transgenic mice leads to a dramatic overproliferation in all areas of the dorsal mesencephalon and diencephalon expressing the transgene. The continued expression of Fgf8 leads to a large expansion of the ventricular zone and subsequently to the perdurance of a large population of dividing ventricular zone cells. This phenotype is complemented by an apparent reduction in the number of differentiated cell types formed. These data are consistent with a model in which FGF8 is a potent mitogen for neural precursor cells and suggest that FGF8 signalling also has a role in regulating the rate of neural differentiation from dividing precursor cells. In this model, as the mesencephalon increases in size, the most rostral cells may come to lie beyond the effective range of FGF8 signal produced by the isthmus and as a result these cells cease dividing and differentiate, leading to the normal rostrocaudal cytoarchitectonic developmental gradient. It is interesting to note that widespread ectopic expression of En in the tectum disrupts the gradient of differentiation as well (Logan et al., 1996). Thus, the widespread activation of En-2 in our transgenics may be responsible for a retardation of cytoararchitectonic differentiation. However, as the widespread expression of En does not lead to overgrowth (Logan et al., 1996) we conclude that the mitogenic role of FGF8 we observe is independent of En. Several lines of evidence indicate that FGFs may regulate neural proliferation. For example bFGF (FGF2) has been shown to act as a potent mitogen for precursor cells of many CNS regions in vitro (Gensburger et al., 1987; Temple and Qian, 1995). In these experiments bFGF acts primarily to increase the number of precursor cells in the culture without promoting the production of differentiated cell types. Interestingly, as FGF2 and FGF8 behave similarly in various assays, such as their ability to induce ectopic limb development in the flank of chick embryos (Cohn et al., 1995; Crossley et al., 1996b; Vogel et al., 1996), these two factors may have similar activities in the developing brain. We propose that the local production of FGF8 at the isthmus may be responsible for the known gradient of growth and differentiation along the rostrocaudal axis of the tectum. As development proceeds and the tectal neuroepithelium expands along the rostrocaudal axis, cells progressively become positioned further away from the isthmic source of FGF8 and thus beyond its mitogenic influence, leading to a cessation of mitosis and subsequent differentiation. Interestingly, despite the fact that Fgf8 is expressed in the myelencephalon and spinal cord of transgenic embryos we observe no apparent growth phenoytpe in these regions. This suggests a polarity in the responsiveness of cells. Caudal to the isthmus neural cells (including those normally expressing Fgf8) are not competent to respond mitogenically to FGF8 signalling, although they do clearly activate En-2 transcription in response to ectopic FGF8. Our experiments have identified two overlapping yet distinct domains of competence for two unique responses to FGF8 signalling. The ability of FGF8 to induce En-2 expression in our experiments extends at least as far caudally as the myelencephalon and has a rostral limit at the mesen- 967 cephalon-p1 boundary. In contrast, the mitogenic response to FGF8 signalling extends from p1 (the most rostral domain tested) to a caudal limit at the isthmus. The mesencephalon is competent for both responses. Mes/met pattern formation and the midbrain organizer Our results have only addressed the role of isthmic-derived FGF8 signalling in regulating mesencephalic polarity. However, it is interesting to consider these experiments in the light of other work that has implicated FGF8 in mesencephalon induction. The ability of FGF8 protein coated beads to induce ectopic mesencephalon formation in chick embryos suggests the involvement of FGF8 in normal mesencephalon induction (Crossley et al., 1996a). If FGF8 signalling normally has a role in mesencephalon induction it seems unlikely that the normal source of FGF8 is from the brain, as expression is not initiated in the mes/met region until 3-somites (Mahmood et al., 1995). This is after the regional activation of several genes in the presumptive mes/met region including Pax-2, Wnt-1 and En-1 (Davis and Joyner, 1988; McMahon et al., 1992; Rowitch and McMahon, 1995). Fgf8 is also expressed in the cardiogenic mesoderm (Heikinheimo et al., 1994; Crossley and Martin, 1995; Mahmood et al., 1995) and it has been suggested that vertical FGF8 signalling may induce mesencephalon formation in the overlying neural plate (Crossley et al., 1996a). However, Pax-2 expression is initiated at 7.75 d.p.c. in the presumptive mes/met domain (Rowitch and McMahon, 1995), before Fgf8expressing cardiogenic mesodermal cells underlie the presumptive mesencephalon. This suggests that mesencephalon development initiates prior to FGF8 signalling. Our data argue strongly that following mesencephalon induction, FGF8 is involved in a signalling loop which regulates the rostrocaudal polarity of the mesencephalon. It is possible that one of the targets of this pathway (e.g. En) may play an earlier role in mesencephalic specification. Consequently, its activation at earlier stages in responsive tissue may trigger mesencephalon development. Thus, the issue of whether FGF8 plays a direct role in mesencephalon induction remains open. Analysis of Wnt-1 mutants indicates that the maintenance of Fgf8 expression in the rostral metencephalon is dependent on mesencephalic derived factors. Because mesencephalon development is initiated normally in these mutants we can not rule out the possibility that mesencephalic derived factors might also be required to initiate Fgf8 expression. Recent studies in which Fgf8 expression is maintained in Wnt-1 mutants argue that it is not Wnt-1 directly which maintains metencephalic Fgf8 expression, but rather a secondary mesencephalic derived signal(s) (Danielian and McMahon, 1996). Our results suggest that FGF8 signalling in turn activates En-2 expression in the caudal mesencephalon and rostral metencephalon. The fact that Fgf8 expression initiates at 3-somites (Mahmood et al., 1995) and En-2 expression shortly thereafter, at 5-somites (Davis and Joyner, 1988; Davis et al., 1988; McMahon et al., 1992), suggests that isthmic derived FGF8 signalling may actually be involved in En-2 activation. Consensus sites for Pax-2,5, or 8 have been shown to be required for proper activation of an En2 minimal enhancer and are therefore implicated in the regulation of En-2 (Song et al., 1996). However, we have observed continued expression of En-2 in the absence of ectopic Pax-2 968 S. M. K. Lee and others or Pax-5 expression. Thus, the relative importance of Pax-2/5 and FGF8 signalling in regulating En-2 expression during normal development remains unclear. Once established, the En-2 expression gradient then acts to regulate midbrain polarity through the regulation of genes such as ELF-1. While our data strongly implicate FGF8 signalling in the regulation of growth and polarity in the mesencephalon, lossof-function experiments are required to directly assess the requirement for Fgf8 in regulating these processes. Additionally, it will be interesting to determine the range of FGF8 signalling from the isthmus and whether FGF8 acts directly to regulate mesencephalic development or whether secondary, FGF8 induced signals are involved. We thank Bianca Klumpar for sectioning; Clive Dickson for the Fgf8 cDNA; John Flanagan for Mek4-AP supernatant; Alex Joyner and Peter Gruss for in situ probes; Andreas Kispert and Terry Yamaguchi for helpful comments on an earlier version of the manuscript; and especially Elisa Marti for many valuable discussions. These studies were supported by a grant from the NIH to A. P. M., and by a postdoctoral fellowship from the Human Frontiers Science Program to P. S. D. REFERENCES Adams, B., Dorfler, P., Aguzzi, A., Kozmik, Z., Urbanek, P., Maurer-Fogy, I. and Busslinger, M. (1992). Pax-5 encodes the transcription factor BSAP and is expressed in B lymphocytes, the developing CNS, and adult testis. Genes Dev. 6, 1589-1607. Altman, J. and Bayer, S. A. (1981a). Time of origin of neurons of the rat inferior colliculus and the relations between cytogenesis and tonotopic order in the auditory pathway. Exp. Brain Res. 42, 411-423. Altman, J. and Bayer, S. A. (1981b). Time of origin of neurons of the rat superior colliculus in relation to other components of the visual and visuomotor pathways. Exp. Brain Res. 42, 424-434. Alvarado-Mallart, R.-M., Martinez, S. and Lance-Jones, C. C. (1990). Pluripotentiality of the 2-day-old avian germinative neuroepithelium. Dev. Biol. 139, 75-88. Asano, M. and Gruss, P. (1992). Pax-5 is expressed at the midbrain-hindbrain boundary during mouse development. Mech. Dev. 39, 29-39. Bally-Cuif, L., Alvarado-Mallart, R.-M., Darnell, D. K. and Wassef, M. (1992). Relationship between Wnt-1 and En-2 expression domains during early development of normal and ectopic met-mesencephalon. Development 115, 999-1009. Bally-Cuif, L. and Wassef, M. (1994). Ectopic induction and reorganization of Wnt-1 expression in quail/chick chimeras. Development 120, 3379-3394. Bally-Cuif, L. and Wassef, M. (1995). Determination events in the nervous system of the vertebrate embryo. Curr. Opin. Genet. Dev. 5, 450-458. Cheng, H. and Flanagan, J. G. (1994). Identification and cloning of ELF-1, a developmentally expressed ligand for the Mek4 and Sek receptor tyrosine kinases. Cell 79, 157-168. Cheng, H., Nakamoto, M., Bergemann, A. D. and Flanagan, J. G. (1995). Complementary gradients in expression and binding of ELF-1 and Mek-4 in development of the topographic retinotectal projection map. Cell 82, 371381. Cohn, M. J., Izpisua-Belmonte, J.-C., Abud, H., Heath, J. K. and Tickle, C. (1995). Fibroblast growth factors induce additional limb development from the flank of chick embryos. Cell 80, 739-746. Crossland, W. J., Cowan, W. M., Rogers, L. A. and Kelly, J. P. (1974). Specification of the retino-tectal projection in the chick. J. Comp. Neurol. 155, 127-164. Crossley, P. H. and Martin, G. R. (1995). The mouse Fgf8 gene encodes a family of polypeptides and is expressed in regions that direct outgrowth and patterning in the developing embryo. Development 121, 439-451. Crossley, P. H., Martinez, S. and Martin, G. R. (1996a). Midbrain development induced by FGF8 in the chick embryo. Nature 380, 66-68. Crossley, P. H., Minowada, G., MacArthur, C. A. and Martin, G. R. (1996b). Roles for FGF8 in the induction, initiation, and maintenance of chick limb development. Cell 84, 127-136. Danielian, P. S. and McMahon, A. P. (1996). Engrailed-1 as a target of the Wnt-1 signalling pathway in vertebrate midbrain development. Nature 383, 332-334. Davis, C., A. and Joyner, A. L. (1988). Expression patterns of the homeoboxcontaining genes En-1 and En-2 and the proto-oncogene int-1 diverge during mouse development. Genes Dev. 2, 1736-1744. Davis, C. A., Noble-Topham, S. E., Rossant, J. and Joyner, A. L. (1988). Expression patterns of the homeobox containing gene En-2 delineates a specific region in the developing mouse brain. Genes Dev. 2, 361-371. DeLong, G. R. and Coulombre, A. J. (1965). Development of the retinotectal projection in the chick embyro. Exp. Neurol. 13, 351-363. Dickinson, M. E., Krumlauf, R. and McMahon, A. P. (1994). Evidence for a mitogenic effect of Wnt-1 in the developing mammalian central nervous system. Development 120, 1453-1471. Drescher, U., Kremoser, C., Handwerker, C., Loschinger, J., Noda, M. and Bonhoeferr, F. (1995). In vitro guidance of retinal ganglion cell axons by RAGS, a 25 kDa tectal protein related to ligands for eph receptor tyrosine kinases. Cell 82, 359-370. Echelard, Y., Epstein, D. J., St-Jacques, B., Shen, L., Mohler, J., McMahon, J. A. and McMahon, A. P. (1993). Sonic hedgehog, a member of a family of putative signalling molecules, is implicated in the regulation of CNS polarity. Cell 75, 1417-1430. Echelard, Y., Vassileva, G. and McMahon, A. P. (1994). Cis-acting regulatory sequences governing Wnt-1 expression in the developing mouse CNS. Development 120, 2213-2224. Friedman, G. C. and O’Leary, D. D. M. (1996). Retroviral misexpression of engrailed genes in the chick optic tectum perturbs the topographic targeting of retinal axons. J. Neurosci. 16, 5498-5509. Gardner, C. A. and Barald, K. (1992). Expression patterns of Engrailed-like proteins in the chick embryo. Dev. Dyn. 193, 370-388. Gardner, C. A. and Barald, K. F. (1991). The cellular environment controls the expression of engrailed-like protein in the cranial neuroepithelium of quail-chick chimeric embryos. Development 113, 1037-1048. Gardner, C. A., Darnell, D. K., Poole, S. J., Ordahl, C. P. and Barald, K. F. (1988). Expression of an engrailed-like gene during development of the early embryonic chick nervous system. J. Neurosci. Res. 21, 426-437. Gensburger, C., Labourdette, G. and Sensenbrenner, M. (1987). Brain basic fibroblast growth factor stimulates the proliferation of rat neuronal precursor cells in vitro. FEBS Lett. 217, 1-5. Hanks, M., Wurst, W., Anson-Cartwright, L., Auerbach, A. B. and Joyner, A. L. (1995). Rescue of the En-1 mutant phenotype by replacement of En-1 with En-2. Science 269, 679-682. Heikinheimo, M., Lawshe, A., Shackleford, G. M., Wilson, D. B. and MacArthur, C. A. (1994). Fgf-8 expression in the post-gastrulation mouse suggests roles in the development of the face, limbs, and central nervous system. Mech. Dev. 48, 129-138. Itasaki, N., Ichijo, H., Hama, C., Matsuno, T. and Nakamura, H. (1991). Establishment of rostrocaudal polarity in tectal primordium: engrailed expression and subsequent tectal polarity. Development 113, 1133-1144. Itasaki, N. and Nakamura, H. (1992). Rostrocaudal polarity of the tectum in birds: correlation of en gradient and topographic order in retinotectal projection. Neuron 8, 787-798. Itasaki, N. and Nakamura, H. (1996). A role for gradient en expression in positional specification on the optic tectum. Neuron 16, 55-62. Joyner, A. L. (1996). Engrailed, Wnt and Pax genes regulate midbrainhindbrain development. Trends Genet. 12, 15-20. Kelly, O. G. and Melton, D. A. (1995). Induction and patterning of the vertebrate nervous system. Trends Genet. 11, 273-278. Knecht, A. K., Good, P. J., Dawid, I. B. and Harland, R. M. (1995). Dorsalventral patterning and differentiation of noggin-induced neural tissue in the absence of mesoderm. Development 121, 1927-1936. LaVail, J. H. and Cowan, W., M. (1971). The development of the chick optic tectum. I. Normal morphology and cytoarchitectonic development. Brain Res. 28, 391-419. Logan, C., Wizenmann, A., Drescher, U., Monschau, B., Bonhoeffer, F. and Lumsden, A. (1996). Rostral optic tectum acquires caudal characteristics following ectopic Engrailed expression. Curr. Biol. 6, 1006-1014. MacArthur, C. A., Lawshe, A., Shanker, D. B., Heikenheimo, M., Shackleford, G. M. (1995a). FGF-8 isoforms differ in NIH3T3 cell transforming potential. Cell Growth Diff. 6, 817-825. MacArthur, C. A., Lawshe, A., Xu, J., Santos-Ocampo, S., Heikinheimo, M., Chellaiah, A. T., and Ornitz, D. M. (1995b). FGF-8 isoforms activate receptor forms that are expressed in mesenchyme regions of mouse development. Development 121, 3603-3613. FGF8 signalling in the midbrain Mahmood, R., Bresnick, J., Hornbruch, A., Mahony, C., Morton, N., Colquhoun, K., Martin, P., Lumsden, A., Dickson, C. and Mason, I. (1995). A role for FGF-8 in the initiation and maintenance of vertebrate limb bud outgrowth. Curr. Biol.5, 797-806. Marin, F. and Puelles, L. (1994). Patterning of the embryonic avian midbrain after experimental inversion: a polarizing activity form the isthmus. Dev. Biol. 163, 19-37. Martinez, S. and Alvarado-Mallart, R.-M. (1990). Expression of the homeobox chick-en gene in chick/quail chimeras with inverted mesmetencephalic grafts. Dev. Biol. 139, 432-436. Martinez, S., Marin, F., Nieto, M. A. and Puelles, L. (1995). Induction of ectopic engrailed expression and fate change in avian rhombomeres: intersegmental boundaries as barriers. Mech. Dev. 51, 289-303. Martinez, S., Wassef, M. and Alvarado-Mallart, R.-M. (1991). Induction of a mesencephalic phenotype in the 2-day-old chick prosencephalon in preceded by the early expression of the homeobox gene en. Neuron 6, 971981. Matsuno, T., Ichijo, H. and Nakamura, H. (1991). Regulation of the rostrocaudal axis of the optic tectum: histological study after rostrocaudal rotation in quail-chick chimeras. Dev. Brain Res. 58, 265-270. McMahon, A. P. and Bradley, A. (1990). The Wnt-1 (int-1) proto-oncogene is required for development of a large region of the mouse brain. Cell 62, 10731085. McMahon, A. P., Joyner, A. L., Bradley, A. and McMahon, J. A. (1992). The midbrain-hindbrain phenotype of Wnt-1−/Wnt-1− mice results from stepwise deletion of engrailed-expressing cells by 9.5 days postcoitum. Cell 69, 581595. Millen, K. J., Wurst, W., Herrup, K. and Joyner, A. L. (1994). Abnormal embryonic cerebellar development and patterning of postnatal foliation in two mouse Engrailed-2 mutants. Development 120, 695-706. Millen, K. J., Hui, C.-C. and Joyner, A.L. (1995). A role for En-2 and other murine homologues of Drosophila segment polarity genes in regulating positional information in the developing cerebellum. Development 121, 3935-3945. Millet, S. and Alvarado-Mallart, R.-M. (1995). Expression of the homeoboxcontaining gene En-2 during the development of the chick central nervous system. Eur. J. Neurosci. 7, 777-791. Nakamoto, M., Cheng, H.-J., Friedman, G. C., McLaughlin, T., Hansen, M. J., Yoon, C. H., O’Leary, D. D. M. and Flanagan, J. G. (1996). Topographically specific effects of ELF-1 on retinal axon guidance in vitro and retinal axon mapping in vivo. Cell 86, 755-766. 969 Nornes, H. O., Dressler, G. R., Knapik, E. W., Deutsch, U. and Gruss, P. (1990). Spatially and temporally restricted expression of Pax-2 during murine neurogenesis. Development 109, 797-809. Parr, B. P., Shea, M. J., Vassileva, G. and McMahon, A. P. (1993). Mouse Wnt genes exhibit discrete domains of expression in the early embryonic CNS and limb buds. Development 119, 247-261. Patel, N. H., Martin-Blanceo, E., Coleman, K. G. P., S.J.Ellis, M.C., Kornberg, T. B. and Goodman, C. S. (1989). Expression of engrailed proteins in arthropods, annelids, and chordates. Cell 58, 955-968. Puschel, A. W., Westerfield, M. and Dressler, G. R. (1992). Comparative analysis of Pax-2 protein distributions during neurulation in mice and zebrafish. Mech. Dev. 38, 197-208. Rowitch, D. H. and McMahon, A. P. (1995). Pax-2 expression in the murine neural plate precedes and encompasses the expression domains of Wnt-1 and En-1. Mech. Dev. 52, 3-8. Rubenstein, J. L. R., Martinez, S., Shimamura, K. and Puelles, L. (1994). The embryonic vertebrate forebrain: the prosomeric model. Science 266, 578-580. Serbedzija, G. N., Dickinson, M. E. and McMahon, A. P. (1996). Cell death in the CNS of the Wnt-1 mutant mouse. J. Neurobiol. In Press. Song, D., Chalepakis, G., Gruss, P. and Joyner, A. L. (1996). Two Paxbinding sites are required for early embryonic brain expression of an Engrailed-2 transgene. Development 122, 627-635. Temple, S. and Qian, X. (1995). bFGF, neurotrophins, and the control of cortical neurogenesis. Neuron 15, 249-252. Thomas, K. R. and Capecchi, M. R. (1990). Targeted disruption of the murine int-1 proto-oncogene resulting in severe abnormalities in midbrain and cerebellar development. Nature 346, 847-850. Urbanek, P., Wang, Z., Fetla, I., Wagner, E. F. and Busslinger, M. (1994). Complete block of early B cell differentialtion and altered patterning of the posterior midbrain in mice lacking Pax5/BSAP. Cell 79, 901-912. Vogel, A., Rodriguez, C. and Izpisua-Belmonte, J.-C. (1996). Involvement of FGF-8 in initiation, outgrowth and patterning of the vertebrate limb. Development 122, 1737-1750. Walther, C. and Gruss, P. (1991). Pax-6, a murine paired box gene, is expressed in the developing CNS. Development 113, 1435-1449. Wilkinson, D. G., Bailes, J. A. and McMahon, A. P. (1987). Expression of the proto-oncogene int-1 is restricted to specific neural cells in the developing mouse embryo. Cell 50, 79-88. (Accepted 23 December 1996)