Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
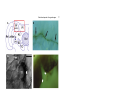
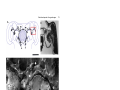
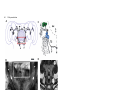
Development 121, 75-86 (1995) Printed in Great Britain © The Company of Biologists Limited 1995 75 Axogenesis in the embryonic brain of the grasshopper Schistocerca gregaria: an identified cell analysis of early brain development George Boyan1, Stavros Therianos1, J. Leslie D. Williams2 and Heinrich Reichert1 1Laboratory of Neurobiology, Department of Zoology, University of Basel, CH-4051 Basel, Switzerland 2Max-Planck-Institut für Verhaltensphysiologie, Arbeitsgruppe Kaissling, D-82319 Seewiesen, Germany SUMMARY Axogenesis in the embryonic brain was studied at the single cell level in the grasshopper Schistocerca gregaria. A small set of individually identifiable pioneer neurons establishes a primary axon scaffold during early embryogenesis. At the beginning of scaffold formation, pioneering axons navigate along and between glial borders that surround clusters of proliferating neuroblasts. In each brain hemisphere, an axonal outgrowth cascade involving a series of pioneer neurons establishes a pathway from the optic ganglia to the brain midline. At the midline the primary preoral commissural interconnection in the embryonic brain is pioneered by a pair of midline-derived pioneer neurons. A second preoral commissural connection is pioneered by two pairs of pars intercerebralis pioneer neurons. Descending tracts are pioneered by the progeny of identified neuroblasts in the pars intercerebralis, deutocerebrum and trito- cerebrum; the postoral tritocerebral commissure is pioneered by a pair of tritocerebral neurons. All of the pioneering brain neurons express the cell adhesion molecule fasciclin I during initial axon outgrowth and fasciculation. Once established, the primary axon scaffold of the brain is used for fasciculation by subsequently differentiating neurons and, by the 40% stage of embryogenesis, axonal projections that characterize the mature brain become evident. The single cell analysis of grasshopper brain development presented here sets the stage for manipulative cell biological experiments and provides the basis for comparative molecular genetic studies of embryonic brain development in Drosophila. INTRODUCTION In this report, we analyse the cellular processes that give rise to the initial set of axonal projections in the embryonic brain of the grasshopper. We use immunocytochemical, intracellular dye injection and electron microscopical techniques to identify as individuals the neurons that pioneer the initial projections in the embryonic brain and to determine the lineage of these neurons from identified brain neuroblasts. During early neurogenesis, the axons from these neurons initially navigate along glial-bound aggregates of proliferating neuroblasts, termed proliferative clusters, that are established before axogenesis begins. Through axonal outgrowth and fasciculation, the identified pioneering neurons rapidly establish a primary axon scaffold that interconnects all of the proliferative clusters in the embryonic brain. This scaffold is used for fasciculation by many of the subsequently differentiating neurons and, by the 40% stage of embryogenesis tracts, commissures and connectives of the mature brain become apparent. In the past decade, the embryonic central nervous system (CNS) of insects has become an important model system for investigations of neuronal development. Many of the cellular mechanisms of neurogenesis and axonal pathfinding have been analyzed in detail (i.e. Goodman et al., 1984; Bastiani et al., 1985; Doe et al., 1985). Insight into the molecular mechanisms that control these developmental processes has also been gained, both through the use of hybridoma technology (Bastiani et al., 1987; Harrelson and Goodman, 1988; Snow et al., 1988) and through molecular genetic analyses of neuronal development in Drosophila (for reviews see Grenningloh et al., 1990; Campos-Ortega and Knust, 1990; Goodman and Doe, 1994). Most of the investigations of embryonic development in the insect CNS have been carried out on the accessible and easily identifiable cells in the segmental ganglia. The brain, by contrast, is an enormously complex structure which in large insects can consist of over a million neurons (Farrel and Kuhlenbeck, 1964). While an initial analysis of neurogenesis in the grasshopper brain has described neuroblasts from which the brain develops (Zacharias et al., 1993), virtually nothing is known about the mechanisms by which the progeny of these neuroblasts produce the complex axonal projections of the adult brain (Boyan et al., 1993). Key words: axogenesis, brain, fasciculation, glia, embryo, grasshopper, central nervous system. MATERIALS AND METHODS Animals Schistocerca gregaria eggs were kept in moist aerated containers at 30°C. Embryo staging was according to Bentley et al. (1979). This method stages embryos at time intervals equal to percentage of embryogenesis completed. 76 G. Boyan and others Whole-mount histology and histological sections Whole-mount preparations (dorsal side up) were dissected to remove the stomodium and reveal the interior of brain structures. For serial sections of osmium-ethyl gallate stained brains, embryos were staged, dissected and treated according to Zacharias et al. (1993). Immunocytochemistry Immunocytochemistry was carried out according to Meier et al. (1993). Primary antibodies were diluted in preincubation solution as follows: anti-horseradish peroxidase (HRP), 1:1 (see Jan and Jan, 1982; Snow et al., 1987); anti-fasciclin I, 1:1 (see Bastiani et al., 1987; Zinn et al., 1988); anti-REGA-1, 1:200 (see Carpenter and Bastiani, 1991); anti-engrailed, 1:1 (see Patel et al., 1989); anti-annulin, 1:1 (see Bastiani et al., 1992). Intracellular dye filling Identified cells were impaled with glass microelectrodes, filled electrophoretically with lucifer yellow dye and processed for light microscopy according to Raper et al. (1983). Alternatively, injected dye was photoconverted and processed for electron microscopy according to Sandell and Masland (1988). Microscopy For light microscopy, histological material or whole-mount embryos were viewed in a Zeiss Axioskop microscope equipped for epifluorescence and DIC (differential interference contrast). For documentation, microscopic images were either photographed or, alternatively, recorded by a CCD camera, digitized and subjected to image processing using the GenIIsys (MTI) and Image 1 systems (Universal Imaging). For immunoelectron microscopy, embryos were processed as described by Meier et al. (1993). The primary antibody dilutions were as above. Cell lineage analysis Brain neuroblast identification was according to Zacharias et al. (1993). Three methods were used to determine the cell lineage of pioneer neurons. First, an initial identification of the lineage of pioneer neurons was possible by determining the position of the neurons in the glia-ensheathed columns relative to their neuroblast of origin. Second, to confirm this initial analysis, lucifer yellow dye was injected into the neuroblast of origin of the pioneer neuron. Due to dye coupling among the neuronal precursors and their early progeny, it was possible to assign a given pioneer neuron to an identified neuroblast. Third, to analyse the generation of neurons from their neuroblasts of origin in vivo and in situ, 5-bromodeoxyuridine (BrdU) incorporation into proliferating neuroblasts was performed according to Zacharias et al. (1993). For each pioneer neuron investigated, this and all of the other experimental analyses reported here were carried out on one or both contralateral homologs of a minimum of 5 different preparations. RESULTS Formation of glial borders around proliferating neuroblasts precedes axogenesis In the embryonic brain of the grasshopper, axogenesis begins at approximately the 29% stage of embryogenesis. At this stage, considerable development of the brain has already occurred, giving rise to a highly structured set of large bilaterally symmetrical neuroblasts (Zacharias et al., 1993), as well as midline progenitors (see below), developing neurons and non-neural cells. Since initial axogenesis relies heavily on the cells in the brain that are in place before the first axon Fig. 1. Formation of glial-bound proliferative clusters in the preaxogenesis brain. REGA-1 immunoreactivity. (A) Schematic of proliferative clusters in one hemisphere of the embryonic brain. Neuroblast 12 of lateral protocerebrum where the first REGA-1 immunoreactive glial processes appear is highlighted. (B) REGA-1 immunoreactive glial processes (white arrow) surround neuroblast 12 of lateral protocerebrum. No other neuroblasts or neurons in the brain hemispheres have REGA-1 immunoreactive glial processes at this stage (26%). (C) At 30% stage REGA-1 immunoreactive glial processes (black arrows) completely surround individual PI neuroblast. (D) Glial processes delimit future axonal pathway inside developing PI proliferative cluster. Dorsal layer of PI neuroblasts has been removed revealing numerous neuronal cell bodies among which the labelled glial process (black arrows) is found. 29% stage. (E) REGA-1-immunoreactive glial processes (white arrows) delimit borders of PI (upper right) and PI(ALDL) (lower left) proliferative clusters. Overlying dorsal sheath removed. 30% stage. (F) Preparation in E at higher magnification. REGA-1 immunoreactive glial processes delimit proliferative cluster border (white arrows). Glia also extend processes into proliferative clusters and surround columnar arrays of neuroblasts and progeny (black arrows). The following abbreviations apply to all figures: GMC, ganglion mother cell; NB, neuroblast; PI, pars intercerebralis proper; PI(ALDL), anterior lateral dorsal lobe of pars intercerebralis; PC, protocerebrum proper; LPC, lateral protocerebrum; DC, deutocerebrum; TC, tritocerebrum; La, lamina; Me, medulla; Re, retina; M, midline. In this and subsequent figures, a rectangular box in the summary diagram indicates the brain region being analysed. Embryos are viewed dorsally unless otherwise stated. Anterior, A, is to the top. Scale bar: B, F, 10 µm; C, 6 µm; D, 14 µm; E, 20 µm. outgrowth is initiated, it is important to characterize the features of the preaxogenesis brain. At early embryonic stages (before 25%), the bilaterally symmetrical neuroblasts in the neurogenic region of the future brain hemispheres are arranged in a sheet-like array, which is largely homogeneous in appearance. Between 26% and 28%, this structural homogeneity disappears and the neuroblasts together with their progeny become arranged into distinct clustered aggregates. The boundaries of these clusters are delineated by glial support cells and the differentiation of these glial borders continues throughout embryonic development. We refer to the glial-bound aggregates of proliferating neuroblasts, ganglion mother cells and neurons in each of the embryonic brain hemispheres as proliferative clusters (Fig. 1A). The proliferative clusters prove to be important for axogenesis within the brain hemispheres because the earliest outgrowing axons navigate in the space between these clusters as well as along their glial borders. Aspects of glial development can be characterized by the expression pattern of REGA-1, an antigen present in nonmidline glia in the embryo (Carpenter and Bastiani, 1991; Seaver et al., 1993). Earliest REGA-1 expression in the brain hemispheres is seen at the 26% stage and is highly restricted; immunoreactive glial processes begin to surround a single brain neuroblast (Fig. 1B). This type of focal initial expression of an antigen by developing glial support cells is also observed for annulin (see Bastiani et al. 1992; and below). Subsequently the spatial expression of the REGA-1 glial marker rapidly expands to surround the surface of other brain neuroblasts (Fig. 1C) as well as the columnar arrays of their progeny (Fig. 1F). In addition, the REGA-1 antigen also appears on elongated glial processes, which extend inside a given proliferative Brain development in the grasshopper D A B C cluster and demarcate future pathways along which the neurites will grow as they project from their neurons of origin outwards toward the surface of the proliferative cluster (Fig. 1D). Although REGA-1 immunoreactive glial processes of this type are observed repeatedly, it has not yet been possible to relate these processes to identified glial cells or determine the identity of the neurons whose axons project along the processes. By the 30% stage, all aggregates of proliferating neuroblasts and their progeny have become bordered by REGA-1 immunoreactive 77 E F glial processes (Fig. 1E,F). These proliferative clusters subdivide the embryonic brain into the different neurogenic regions that will give rise to the pars intercerebralis proper, the anterior lateral dorsal lobe of the pars intercerebralis, the protocerebrum proper, the lateral protocerebrum, the deutocerebrum and the tritocerebrum. In summary, before axogenesis begins, the bilaterally symmetrical brain neuroblasts are subdivided into the proliferative clusters, which give rise to the major brain regions. This sub- 78 G. Boyan and others division is accompanied by the formation of glial borders and by the glial demarcation of future axon pathways. Subsequently both of these glial structures are used as substrata for axon outgrowth. Brain pioneer neurons construct an embryonic axon scaffold Initial axogenesis in the embryonic brain is carried out by a small subset of neurons, which are located at discretely spaced sites in the developing brain. These neurons are found near the periphery of the proliferative clusters and, in many cases, near the abutment of two different clusters. Through axonal outgrowth and fasciculation, this small subset of neurons establishes a primary axon scaffold that interconnects all of the proliferative clusters in the embryonic brain. Given that they are the first to initiate axogenesis in the insect brain, we refer to the neurons that establish the initial interconnections among all of the proliferative clusters in the brain as pioneer neurons in accordance with the terminology used in the peripheral nervous system and in the segmental ganglia (Bate, 1976; Bastiani et al., 1985; Caudy and Bentley, 1986). We studied axonal pathfinding of the brain pioneer neurons by direct observation of developing axonal processes using DIC optics, by intracellular injection of lucifer yellow dye and by immunocytochemistry with an anti-HRP antibody that labels the axons and cell bodies of all outgrowing neurons in the developing insect nervous system (Jan and Jan, 1982; Snow et al., 1987). A semi-schematic diagram of the brain pioneer neurons based on a representative anti-HRP stained preparation is shown in Fig. 2A. Many of the brain pioneer neurons can be identified on the basis of their position, the trajectory of their axonal projections and their molecular expression patterns. For several of the brain pioneers, it has also been possible to identify their neuroblast of origin and determine their lineage; in all of these cases, the brain pioneer neurons belong to the initial set of progeny that a given neuroblast produces. How do the pioneer neurons establish the initial axonal pathways in the embryonic brain? To address this question at the single cell level, we first investigate the formation of the pioneering axonal pathways that link the peripheral part of the brain to more medial structures in each hemisphere. We then analyse the cellular processes through which the commissural connections between the two hemispheres are established. Finally we characterize the formation of descending and tritocerebral connections. In each case, we limit our analysis to initial pioneering neurons and pathfinding processes. A wave of axonal outgrowth interconnects brain and optic lobes At the onset of axogenesis in the brain, a wave of differentiation occurs in which the axons from pioneer neurons located in lateral and more peripheral parts of the brain extend towards the axons of more centrally located pioneers, which then project towards the midline. Throughout this process, the growth cones of all of the pioneer neurons extend along and between proliferative clusters and their filopodia make transient contact with border cells of the proliferative clusters. The first pioneer neurons that initiate axogenesis in the brain are found in the developing optic lobes at the interface between lamina and medulla (Fig. 2A,B). (While the exact mechanisms are still unclear, at least the distal part of the medulla appears to form from cells differentiating from the most proximal regions of the lamina.) There, at the 29% stage, one or a pair of neurons begin to send their axons towards the protocerebrum. These neurons pioneer a pathway, whose location suggests that it may be used later by visual neurons of the medulla (Fig. 2C). The axonal growth cones of these pioneering neurons first grow anteriorly along the border between lamina and medulla. Then near the anterior edge of the border they turn medially. At the turn, their growth cones pass filopodia from a pioneer neuron located at the anterior medullalamina border; subsequently their growth cones encounter filopodia from a pioneer neuron located in the pars intercerebralis (Fig. 2B). Following reciprocal filopodial contact at the interface between the developing optic lobes and the lateral edge of the pars intercerebralis proliferative cluster, the growth cones of all three sets of pioneer neurons fasciculate with one another. The axons from the optic lobes then project along the pars intercerebralis axon into a specific axonal junction zone in the brain hemisphere. During the formation of the pioneering connections between optic lobes and pars intercerebralis, an axonal junction zone is established in the brain at the interface between the pars intercerebralis, anterior lateral dorsal lobe of the pars intercerebralis and protocerebrum (Fig. 3A). There, the axon of a pioneer neuron from the lateral protocerebrum grows out of its proliferative cluster of origin and extends into the intercluster space. At the interface between the three proliferative clusters, its growth cone contacts and fasciculates with the growth cone of the pars intercerebralis neuron which pioneers the pathway extending peripherally towards the optic lobes (Fig. 3B,C). (Note that this is the pathway used by the axons from the optic lobes to grow into the brain hemisphere, see above.) This pioneer neuron is one of the first two neuronal progeny of pars intercerebralis neuroblast 14; its putative sibling neuron pioneers a pathway projecting centrally between proliferative clusters towards the brain midline (Fig. 3D). In this case, the lineage analysis of neurons is based on the position of the neurons in the glial-ensheathed columns relative to their neuroblast of origin; in other cases, intracellular dye-injection experiments and BrdU incorporation were also used (see methods). After fasciculation, the growth cone of the protocerebral pioneer neuron first grows centrally along the axon of the peripherally projecting neuron of the pars intercerebralis. Subsequently (not shown) the axon of the protocerebral pioneer switches its growth cone to the axon of the other sibling pars intercerebralis neuron, fasciculates with this cell’s axon and then continues to grow along the axon of this centrally projecting neuron until it reaches the medial edge of the pars intercerebralis. In this way, a pathway is established from the pars intercerebralis-protocerebral junction zone to the medial edge of the brain hemisphere. Thus, in each hemisphere, the growth cones of brain pioneer neurons extend along the borders of the proliferative clusters of the brain and carry out a series of axonal fasciculation processes at the interfaces between proliferative clusters. Through these fasciculation processes, initial axon pathways are established from the optic lobes into the pars intercerebralis as well as from the lateral protocerebrum via the pars intercerebralis towards the brain midline. With the linkage of these axon pathways at the pars intercerebralis-protocerebral Brain development in the grasshopper A 79 C B Fig. 2. Pioneer neurons establish an axonal pathway from the optic lobes into the brain. (A) Semi-schematic diagram of neurons that establish a primary axon scaffolding; summary based on anti-HRP immunocytochemistry of 29-34% stages. (B) Digitized montage of anti-fasciclin I immunocytochemistry at the 29% stage. The axon (black arrow) of a visual pioneer neuron (white asterisk) from the lamina-medulla interface is growing along the border of a proliferative cluster into the brain. En route the growth cone (open arrowhead) of the visual pioneer neuron grows past filopodia (white short arrow) growing into the brain from a neuron (partly visible, white star) located at the edge of the anterior medulla-lamina border and subsequently encounters filopodia (black arrowheads) directed peripherally into the optic region from another neuron at the PC/PI(ALDL) interface. All growth cones extend between proliferative clusters; their filopodia (long white arrows) make contact with border cells within the proliferative cluster. (C) The axon of the visual pioneer neuron establishes a pathway that may later be utilized by cells of medulla (black arrow). 45% stage. 16 µm horizontal section. Scale bar: B, 15 µm; C, 60 µm. junction zone, a set of interconnecting neuronal pathways are established, which extend from the developing optic lobes towards the midline of the brain. As soon as these initial pathways in the brain are pioneered, they are used by numerous other neurons which project their axons along them. During the construction of the axonal pathways from the optic lobes to the midline, the growth cones and axons of all of these reciprocally fasciculating brain pioneer neurons express the homophilic cell adhesion molecule fasciclin I (Bastiani et al., 1987; Snow et al., 1988; Zinn et al., 1988). Fasciclin I expression is dynamic; expression of fasciclin I by the growth cones and axons of the pioneer neurons is strong during the formation of the primary axon scaffold early in embryogenesis; expression of fasciclin I is no longer evident in these neurons at 40% of embryonic development. Interestingly, the filopodia of the pioneering growth cones often contact cells located at the borders of the proliferative clusters that they grow along and some of these contacted cells also express fasciclin I (see Figs 2B,3B). Formation of the embryonic brain commissure By the time the wave of outgrowing axons in each brain hemisphere reaches the medial border of the hemisphere (31% stage), an initial brain commissural connection has already been established. How is this embryonic brain commissure formed? A central role in pioneering the brain commissure is played by neurons that are generated by a small set of midline progenitor cells. These midline progenitors do not stain with G. Boyan and others 80 C A B D Fig. 3. Contact and fasciculation of pioneer neurons at PI/PI(ALDL)/PC interface. (A) Semi-schematic summary diagram. (B) Digitized montage, anti-fasciclin I immunocytochemistry of 30% stage. Fasciculation of two outgrowing axons, one from the LPC proliferative cluster (white asterisk), the other from the PI proliferative cluster (partly visible, white star). Axonal growth cones (black arrows) meet in the space between the two clusters. (C) Digitized montage showing fasciculating growth cones from PI neuron (white star) and LPC neuron (white asterisk) in B at higher magnification. Filopodia (black arrows) extend from growth cones to proliferative cluster borders. (D) Schematic summary data describing lineage relationships of the neuron indicated by white star in B. One of the first born neurons (white star) of neuroblast 14 projects its axon laterally along a proliferative cluster border to fasciculate with the growth cone of a neuron (white asterisk) from LPC. The other neuron pioneers a pathway projecting centrally towards brain midline along posterior proliferative cluster border of PI. Neuroblast 14 and its first born progeny are in solid blue; other progeny outlined in blue, GMC outlined in black. Unlabelled neuroblasts are protocerebral. Scale bar: B, 20 µm; C, 10 µm. the anti-HRP antibody. Moreover, they do not express the engrailed protein. This contrasts with the segmental ganglia, where large engrailed-positive midline neuroblasts and progeny are seen (data not shown). However, as is the case for neuroblasts in the segmental ganglia, the midline progenitors in the brain do become surrounded by glial sheath cells which express annulin in correlation with the mitotic activity of the cells they surround (Bastiani et al., 1992; Singer et al., 1992). Thus, before the midline progenitors differentiate and become active, no annulin staining is seen in this region of the brain Brain development in the grasshopper A B C D (Fig. 4A). When the midline progenitors generate their progeny, their surrounding glial sheath cells become annulinimmunoreactive and immunoreactive processes surround the proliferating precursors as well as their progeny (Fig. 4B,C). We have identified a set of three midline progenitors based on their position at the midline, on the annulin immunoreactivity of their glial sheath cells and on the incorporation of BrdU into the precursors and their progeny (Fig. 4D). Additional, as yet unidentified midline progenitors may exist. The first pair of neurons that are generated by the lateralmost progenitors on each side of the midline are fasciclin I expressing neurons and these pioneer the first commissural connection in the embryonic brain (Fig. 5A,B). We call these neurons the primary commissural pioneers (PCP). We carried out a detailed analysis of the axonal outgrowth process of these pioneer neurons by intracellular dye filling. At the onset of axogenesis, a single small process extends from each pioneer neuron towards the midline (Fig. 5B). Shortly thereafter a large growth cone with numerous filopodia separated by lamellipodial structures is generated (not shown). This growth cone extends towards the midline and, in doing so, becomes smaller in size and gives rise to a trailing axonal process (Fig. 5B). The growth cones of these pioneer neurons extend over the midline just posterior to the soma of the medial midline precursor. They then contact and fasciculate with the axons of their contralateral homologs and grow along these axons into the contralateral hemisphere, thus establishing an initial axonal bridge across the brain midline (Fig. 5A,C). Once the PCP neurons have established the first axonal pathway across the midline, another pair of neurons at the medioposterior edge of the developing pars intercerebralis in each brain hemisphere, the secondary commissural neurons (SCN), extend their growth cones medially onto this axon pathway and fasciculate with it (Fig. 5D,F). At the midline, the 81 Fig. 4. Midline progenitors in the brain. (A-C) Annulin immunoreactivity; (D) BrdU incorporation. (A) At 26% stage annulin expression is seen in developing PI proliferative clusters (asterisks) but not in preoral brain midline. Annulin expression is also seen in optic lobes, deutocerebrum, tritocerebrum and ventral nerve cord. (B) At 30% stage annulin expression is seen in preoral midline progenitor cells (arrow) and in the PI proliferative clusters (asterisks). (C) Higher magnification showing annulin expression surrounding preoral midline progenitor cells between the PI proliferative clusters (asterisks). Note prominent annulin immunoreactive border (arrowheads) delimiting the cluster of large progenitor cells from surrounding epithelial cells. (D) BrdU labelling of three midline progenitor cells (arrows). Note progeny (star) of the central midline progenitor. BrdU incorporation is also seen in cells of PI proliferative clusters (upper left and right). Scale bar: A, 166 µm; B, 120 µm; C, 14 µm; D, 25µm. growth cone of each SCN neuron contacts and fasciculates with its contralateral homolog (Fig. 5F) just like the PCP neurons did previously (Fig. 5C). Each SCN growth cone then extends along the axon of its contralateral homolog and grows into the contralateral brain hemisphere. Soon after these two initial commissural pathways in the brain are established (within 1-2% of embryonic development), numerous other neurons project axons onto them, some extending growth cones in different directions. The lateral pioneer (LP), for example, has a cell body at the medial edge of the developing pars intercerebralis (Fig. 5D). The LP neuron extends its axon in the opposite direction to the PCP neuron along the initial midline pathway and establishes a lateral pathway that extends from the midline commissure peripherally into the brain hemisphere (Fig. 5D,E). Both the secondary commissural neurons and the lateral pioneer express fasciclin I during axonal outgrowth. The results of an ultrastructural analysis of such a nascent commissural fascicle are shown in Fig. 6. In this experiment, the lateral pioneer and the paired (contralateral) primary commissural pioneers have been filled with dye in a 33-34% stage embryo (Fig. 6A). Closer to the brain midline, the axons of the primary commissure pioneers are tightly apposed to one another and still at some distance from the axon of the lateral pioneer (Fig. 6B). Closely associated with these pioneer axons are numerous other unstained axonal profiles. These unstained axons have projected onto the initial pioneering pathway and together now form the developing commissural fascicle. Interestingly, a large putative midline progenitor cell is in close association with the fascicle, suggesting that this precursor cell might play a role in the commissural pathfinding processes. Further away from the midline, the primary commissure pioneers have fasciculated with the lateral pioneer as they run laterally along the posterior margin of the pars intercerebralis proliferative cluster (Fig. 6C,D). G. Boyan and others 82 A B C D F Once again, numerous unstained axonal profiles are closely associated with the commissural pioneer axons. Together these axons represent the developing commissural fascicle just before it enters the brain hemisphere. A large glial cell, which can be identified by the typical involuted membrane structure surrounding its nucleus, is in close contact with the fascicle. This glia cell extends processes that envelop all of the axons in the fascicle. Numerous glial cells of this type are found E G wherever axonal outgrowth occurs in the brain. They are seen at the midline and in the spaces between the proliferative aggregates of the brain hemispheres; they can be seen extending processes along pathways that are subsequently used by pioneer neurons for axonal outgrowth. These glia are likely to play important roles in early axogenesis, however, a more detailed understanding of their function awaits further investigation. Brain development in the grasshopper Fig. 5. Formation of the primary brain commissure and of other protocerebral pathways. (A) A pair of neurons that originate from a lateral progenitor cell on each side of the midline pioneer the primary preoral commissure. 30-31% stage. Top: camera lucida drawing of these primary commissure pioneer (PCP) neurons (arrowheads, cell bodies). Bottom: anti-HRP immunocytochemistry showing PCP neurons (white arrows) and axons extending across midline (open arrows). (B) Details of the primary commissural pioneers following intracellular lucifer yellow labelling. A PCP neuron initiating process outgrowth at 30% stage (top). Digitized montage (bottom) shows axon extension across midline towards contralateral hemisphere at 31% stage of a different preparation. (C) Intracellular lucifer yellow labelling of a PCP neuron from each brain hemisphere at 32-33% stage. Arrowhead indicates brain midline. Digitized montage. (D) Details of the secondary commissural neurons (SCN), digitized montage of fasciclin I immunoreactivity. The cell body of one SCN neuron in left PI is shown (asterisk). At midline (white short arrow) the fascicle bends around midline cells. The lateral pioneer (LP) (star) located at the medial border of the PI proliferative cluster directs a process laterally along the axon fascicle. Growth cones (white arrow) extending into left PI express fasciclin I as they contact surrounding neurons and proliferative cluster borders. 31% stage. (E) Intracellular lucifer yellow labelling of lateral pioneer neuron (star) described in D; different preparation. Arrow indicates growth cone. (F) Intracellular labelling of SCN neuron (asterisk) from each brain hemisphere. 31-32% stage. Arrowhead indicates preoral brain midline. Digitized montage. (G) Summary diagram showing lineage relationships of neurons involved in early pathway formation in the brain. 32% stage. Progeny of PI neuroblast 3 contribute to commissural and descending pathways; progeny of neuroblast 1 contribute to laterally projecting pathway. Progeny from other unidentified PI neuroblasts contribute to commissural and descending pathways. Arrowheads indicate direction of initial axon outgrowth; lineage-related progeny are coloured the same. Scale bar: A,B, 8 µm; C, 14 µm; D, 10 µm; E, 11 µm; F, 17 µm. A C 83 Formation of embryonic descending pathways and tritocerebral commissure With the establishment of the primary preoral commissure, the axon pathways in the two brain hemispheres become interconnected. Concurrently, descending pathways are pioneered that connect the brain hemispheres with the ganglia of the ventral nerve cord (Figs 5G,7A). This is a multistage process. In the protocerebrum, a descending ipsilateral pathway is pioneered at 31-32% of embryonic development by a pair of cells derived from neuroblast 3 of the pars intercerebralis (Fig. 5G). Within 1% of embryonic development, the axons of neurons derived from other pars intercerebralis neuroblasts project onto this pioneer pathway and through the protocerebrum proper towards the deutocerebrum. Other pioneer neurons derived from neuroblast 1 of the pars intercerebralis project axons laterally towards the optic lobes (Fig. 5G). In the deutocerebrum, neurons derived from neuroblast 5 pioneer a descending ipsilateral pathway along the medial edges of the deutocerebrum to the tritocerebrum and from there through the circumesophageal connectives into the ventral nerve cord (Fig. 7B,C). The axons of all of these descending pioneers express fasciclin I. In the tritocerebrum, two very different pathways are pioneered by the early progeny of tritocerebral neuroblast 12: a descending ipsilateral pathway and the postoral tritocerebral commissure (Fig. 7B,C). The neurons responsible for pioneering both pathways are putative sibling cells. Their axons express fasciclin I and are arranged symmetrically on either side of the brain. Initially the axons of both siblings follow an identical route through the tritocerebrum but they soon part and one sibling directs its axon posteriorly towards the circumesophageal connective. The other directs its growth cone towards the midline where it meets the growth cone of its D Fig. 6. Ultrastructural analysis of commissural development. (A) Schematic summary. Two primary commissural pioneer neurons (PCPs) and the contralateral lateral pioneer (LP) were stained in the same preparation (34% stage) by B intracellular injection and photoconversion of lucifer yellow. Dashed lines indicate plane of sections in B, C. (B) Section shows the PCP axons (open arrowhead) separated from the LP axon (filled arrowhead). All profiles are closely associated with the developing commissure (arrow) and with a large midline precursor (asterisk). (C) Section more lateral than (B). Axon of the PCP neurons and the LP neuron are now fasciculated (arrowhead). The labelled axon profiles and other unlabelled axons of the developing commissure are surrounded by processes of a large glial cell (asterisk). (D) Enlargement of labelled axon profiles (star) seen in C. Scale bar: B,C, 3 µm; D, 1 µm. G. Boyan and others 84 A C B D Fig. 7. Formation of descending pathways and of the tritocerebral commissure. (A) Summary diagram. (B) Formation of the tritocerebral commissure. Digitized montage based on fasciclin I immunoreactivity. 33-34% stage. Two bilaterally symmetrical tritocerebral neurons (stars), the tritocerebral commissure pioneers (TCCP), project axons (white arrows) towards postoral midline (open arrow). Axons (arrowheads) from two pairs of deutocerebral neurons form descending pathways through the tritocerebrum. Inset (white rectangle) is a magnification of midline region and shows that axons from both TCCP neurons have made contact and fasciculated at the midline (black arrow). (C) Summary schematic showing lineage relationships of the neurons described in B. TCCP neuron and sibling (white asterisks) derive from tritocerebral neuroblast 12. TCCP projects across midline; its sibling cell forms a descending axonal projection (black asterisk). Descending pathway from deutocerebrum into ventral nerve cord (more lateral axons in B) is pioneered by progeny (white triangle) of DC neuroblast 5 (33% stage). Only one of the sibling progeny contributing axons is shown. Other lineage-related cells are colored alike. (D) Horizontal section (16µm) through brain and part of subesophageal ganglion showing orthogonal axonal scaffold at 45% stage. Osmium-ethyl gallate staining. P, protocerebrum; T, tritocerebrum; S1, first subesophageal segment. Scale bar: B, 17 µm; inset, 11 µm; D, 108 µm. homolog from the other brain hemisphere; these two axons then fasciculate and grow along their homologs into the contralateral tritocerebrum (Fig. 7B). This initial fascicle of the tritocerebral commissure is pioneered at the 33-34% stage. Aspects of the early axon scaffold remain evident in later embryonic stages Once the axonal scaffolding in the embryonic brain is established, the axonal projections rapidly grow in size. Within 3% of the initial pioneering of the primary preoral brain commissure, over 100 axons have crossed the midline. Indeed, by the 37% stage, some fascicles of the adult brain, such as the Ushaped commissure PC20 (Boyan et al., 1993), can already be recognized. By the 40% stage, a massive amount of axonal outgrowth and fasciculation has occurred in the brain. Tracts, commissures and descending pathways have increased enormously in size and complexity. Moreover, nerves that extend from the peripheral nervous system into the brain have formed. Nevertheless, some aspects of the early orthogonal scaffold of brain axons are still visible at this stage (Fig. 7D). Lateral axon tracts extend from the pars intercerebralis and protocerebrum regions into the optic lobes; descending axon tracts extend from the pars intercerebralis towards the deutocerebrum and tritocerebrum and from there through the connectives into the ventral nerve cord; preoral and postoral brain commissures formed around the gut link the brain hemispheres. Interest- Brain development in the grasshopper ingly, the orthogonal scaffold described above is formed in the absence of all higher brain structures such as the mushroom bodies and the central body. These structures only take shape after 40% of embryonic development. DISCUSSION Investigations on insect axogenesis have become increasingly important for analyzing the cellular and molecular mechanisms involved in guided outgrowth and pathway formation in the nervous system (for recent reviews see Reichert, 1993; Goodman and Doe, 1994). Virtually all of this work has, however, been carried out on the simple segmental ganglia or the peripheral nervous system of insects. In consequence, very little is known about the development of more complex structures such as the insect brain. In this study, we investigate axogenesis in the early embryonic brain of the grasshopper Schistocerca gregaria. Our results show that, despite the enormous complexity of the adult brain, a set of simple developmental processes involving individually identifiable cells and axons underlies early axogenesis in the brain. Axogenesis in the brain commences in a restricted group of cells and leads to a simple orthogonal scaffolding of axonal pathways. The events that occur in the formation of this scaffolding resemble those that are seen in the segmental nervous system in several ways. First, identifiable pioneering neurons establish a simple axonal scaffold (Bate, 1976; Bentley and Keshishian, 1982; Ho and Goodman, 1982). In the brain, pioneers are involved in the construction both of the commissures that link the two hemispheres and of the tracts and connectives that interconnect the different parts of the brain in each hemisphere. Second, in the brain and in the segmental ganglia, all of the initially outgrowing axons derive from the central nervous system itself (Goodman et al., 1984). Axons that grow into the brain from structures on the head such as the antennae, head hairs or the ocelli grow into the axonal scaffolding later and appear to depend on the existence of the central nervous scaffolding for proper navigation. Third, the growth cones of neurons pioneering the preoral and postoral brain commissures meet and then fasciculate with their contralateral homologs in the midline as described for commissure formation in the segmental ganglia (Myers and Bastiani, 1993). Fourth, glia play an important role in axonal pathfinding (Bastiani and Goodman, 1986; Meyer et al., 1987; Jacobs and Goodman, 1989; Klämbt and Goodman, 1991; Carpenter and Bastiani, 1991). The pioneering axons of the brain follow glial borders during their outgrowth and glia are also important in establishing connections across the midline. Interestingly, glial bound neuronal aggregates are likely to play important roles in the postembryonic development of the insect brain (Tolbert and Oland, 1989) as well as in embryonic brain development in vertebrates (see Steindler, 1993). Fifth, some cell bodies in the developing brain may have a function equivalent to that of ‘guidepost cells’ in the peripheral nervous system (Bentley and Keshishian, 1982; Ho and Goodman, 1982). Such cell bodies are observed near the borders of proliferative clusters. These cells express the cell adhesion molecule fasciclin I and are contacted by outgrowing axons that also express fasciclin I. These cells, therefore, might play an important role in directing axon outgrowth. 85 The primary scaffold of axons in the embryonic brain is established in the absence of higher brain structures such as the central body, the protocerebral bridge or the mushroom bodies. These structures are established as part of a second stage of development that leads to the brain acquiring the anatomical organization characteristic of the adult. In hemimetabolous insects like the grasshopper, this second stage of development occurs in the embryo once the first stage has been achieved (Boyan et al., unpublished data). Thus, the early embryonic brain is not constructed as a miniature model of the adult brain. It initially acquires features that are reminiscent of the developing ventral ganglia. Only later does the embryonic brain assume the features that are specific for the mature supraesophageal ganglion. In consequence, for a complete understanding of axogenesis in the insect brain, the formation of the nerve tracts and connections in these later developing, higher brain structures must also be investigated. The single cell analysis of early brain axogenesis described here lays the foundation for further cell biological studies in which the many experimental advantages of the grasshopper embryo, such as large size of embryonic neurons, easy accessibility for intracellular labelling and ablation, existence of a robust embryo culture system and established antibody-block protocols (Raper et al., 1984; Bastiani et al., 1985; Kolodkin et al., 1993; Xie et al., 1994) can be exploited. Moreover, this analysis sets the stage for following molecular genetic investigations of embryonic brain development in Drosophila.(Therianos et al., unpublished data). The merger of work on the grasshopper with its highly accessible identified embryonic neurons with work on Drosophila with its powerful genetics and molecular biology has led to significant advances in our understanding of neuronal development in general (Thomas et al., 1984; Goodman et al., 1984; Zinn et al., 1988; Grenningloh et al., 1990). Indeed, the expectation that mechanisms and molecules, first discovered in insects, have equivalents in the developing vertebrate nervous system has already been fulfilled in several cases (Thomas and Capecchi, 1990; McGinnis and Krumlauf, 1992; Goodman and Shatz, 1993; Kolodkin et al., 1993). Anti-fasciclin I, anti-annulin and anti-engrailed antibodies were generous gifts of C. S. Goodman and N. Patel; anti-REGA-1 antibodies were generous gifts of M. J. Bastiani and E. C. Seaver. We thank T. Meier, F. Xie, D. Sanchez, L. Ganfornina and E. Ball for technical advice. Supported by the Swiss NSF, the FAG Basel and the Max-Planck Gesellschaft, Seewiesen. REFERENCES Bastiani, M. J., de Couet, H. G., Quinn, J. M. A., Karlstrom, R. O., Kotrla, K, Goodman, C. S. and Ball, E. E. (1992). Position-specific expression of the annulin protein during grasshopper embryogenesis. Dev. Biol. 154, 129142. Bastiani, M. J., Doe, C. Q., Helfand, S. L. and Goodman, C. S. (1985). Neuronal specificity and growth cone guidance in grasshopper and Drosophila embryos. Trends in Neurosci. 8, 257-266. Bastiani, M. J. and Goodman, C. S. (1986). Guidance of neuronal growth cones in the grasshopper embryo. III. Recognition of specific glial pathways. J. Neurosci. 6, 3542-3551. Bastiani, M. J., Harrelson, A. L., Snow, P. M. and Goodman, C. S. (1987). Expression of fasciclin I and II glycoproteins on subsets of axon pathways during neuronal development in the grasshopper. Cell 48, 745-755. Bate, M. (1976). Pioneer neurons in an insect embryo. Nature 260, 54-56. 86 G. Boyan and others Bentley, D. H. and Keshishian, H. (1982). Pathfinding by peripheral pioneer neurons in grasshoppers. Science 218, 1082-1088. Bentley, D., Keshishian, H., Shankland, M. and Toroian-Raymond, A. (1979). Quantitative staging of embryonic development of the grasshopper, Schistocerca nitens. J. Embryol. Exp. Morph. 54, 47-74. Boyan, G., Williams, L. and Meier, T. (1993). Organization of the commissural fibers in the adult and early embryonic brain of the locust. J. Comp. Neurol. 332, 358-377. Campos-Ortega, J. A. and Knust, E. (1990). Genetic mechanisms in early neurogenesis of Drosophila melanogaster. Ann. Rev. Genet. 24, 387-407. Carpenter, E. M. and Bastiani, M. J. (1991). Developmental expression of REGA-1, a regionally expressed glial antigen in the central nervous system of grasshopper embryos. J. Neurosci. 11, 277-286. Caudy, M. and Bentley, D. (1986). Pioneer growth cone steering along a series of neuronal and non-neuronal cues of different affinities. J. Neurosci. 6, 1781-1795. Doe, C. Q., Kuwada, J. Y. and Goodman, C. S. (1985). From epithelium to neuroblasts to neurones: the role of cell interactions and cell lineage during insect neurogenesis. Phil. Trans. R. Soc. Lond. B 312, 67-81. Farrel, S. and Kuhlenbeck, H. (1964). Preliminary computation of the number of cellular elements in some insect brains. Anat. Rec. 148, 369-370. Goodman, C. S., Bastiani, M. J., Doe, C. Q., du Lac, S., Helfand, S. L., Kuwada, J. Y. and Thomas, J. B. (1984). Cell recognition during development. Science 225, 1271-1279. Goodman, C. S. and Doe, C. Q. (1994). Embryonic development of the Drosophila central nervous system. In The Development of Drosophila.vol. 1 (ed. M. Bate and A. Martinez-Arias), pp. 1131-1206. New York: Cold Spring Harbor. Goodman, C. S. and Shatz, C. J. (1993) Developmental mechanisms that generate precise patterns of neuronal connectivity. Neuron 10, 77-98. Grenningloh, G., Bieber, A., Rehm, E. J., Snow, P., Traquina, Z., Hortsch, M., Patel, N. and Goodman, C. S. (1990). Molecular genetics of neuronal recognition in Drosophila: Evolution and function of immunoglobulin superfamily cell adhesion molecules. Cold Spring Harbor Symp. Quant. Biol. 55, 327-340. Harrelson, A. L. and Goodman, C. S. (1988). Growth cone guidance in insects: fasciclin II is a member of the immunoglobulin superfamily. Science 242, 700-708. Ho, R. K. and Goodman, C. S. (1982). Peripheral pathways are pioneered by an array of central and peripheral neurons in grasshopper embryos. Nature 297, 404-406. Jacobs, J. R. and Goodman, C. S. (1989). Embryonic development of axon pathways in the Drosophila CNS. I. A glial scaffold appears before the first growth cones. J. Neurosci. 9, 2402-2411. Jan, L. Y. and Jan, Y. N. (1982). Antibodies to horseradish peroxidase as specific neuronal markers in Drosophila and grasshopper embryos. Proc. Natl. Acad. Sci. USA 79, 2700-2704. Klämbt, C. and Goodman, C. S. (1991). The diversity and pattern of glia during axon pathway formation in the Drosophila embryo. Glia 4, 205-213. Kolodkin, A. L., Matthes, D. J., O’Connor, T. P., Patel, N. H., Admon, A., Bentley, D. and Goodman, C. S. (1992). Fasciclin IV: Sequence, expression, and function during growth cone guidance in the grasshopper embryo. Neuron 9, 831-845. McGinnis, W. and Krumlauf, R. (1992) Homeobox genes and axial patterning. Cell 68, 283-302. Meier, T., Therianos, S., Zacharias, D. and Reichert, H. (1993). Developmental expression of the TERM-1 glycoprotein on growth cones and terminal arbors of individual identified neurons in the grasshopper. J. Neurosci. 13, 1498-1510. Meyer, M. R., Reddy, G. R. and Edwards, J. S. (1987). Immunological probes reveal spatial and developmental diversity in insect neuroglia. J. Neurosci. 7, 512-521. Myers, P. Z. and Bastiani, M. J. (1993). Growth cone dynamics during the migration of an identified commissural growth cone. J. Neurosci. 13, 127143. Patel, N. H., Kornberg, T. B. and Goodman, C. S. (1989). Expression of engrailed during segmentation in grasshopper and crayfish. Development 107, 201-212. Raper, J. A., Bastiani, M. J. and Goodman, C. S. (1983). Pathfinding by neuronal growth cones in grasshopper embryos. I. Divergent choices made by the growth cones of sibling neurons. J. Neurosci. 3, 20-30. Raper, J. A., Bastiani, M. J. and Goodman, C. S. (1984). Pathfinding by neuronal growth cones in grasshopper embryos. IV. The effects of ablating the A and P axons upon the behavior of the G growth cone. J. Neurosci. 4, 2329-2345. Reichert, H. (1993). Molecular correlates of neuronal specificity in the developing insect nervous system. Mol. Neurobiol. 7, 349-362. Sandell, J. H. and Masland, R. H. (1988). Photoconverson of some fluorescent markers to a diaminobenzidine product. J. Histochem. Cytochem. 36, 555-559. Seaver, E. C., Carpenter, E. M. and Bastiani, M. J. (1993). Glial cells in the development of the nervous system: developmental expression and molecular characterization of REGA-1 in grasshoppers. Neurosci. Abstr. 19, 266. Singer, M. A., Hortsch, M., Goodman, C. S. and Bentley, D. (1992). Annulin, a protein expressed at limb segment boundaries in the grasshopper embryo, is homologous to protein cross-linking transglutaminases. Dev. Biol. 154, 143-159. Snow, P. M., Patel, N. H., Harrelson, A. L. and Goodman, C. S. (1987). Neural specific carbohydrate moiety shared by many surface glycoproteins in Drosophila and grasshopper embryos. J. Neurosci. 7, 4137-4144. Snow, P. M., Zinn, K., Harrelson, A. L., McAllister, L., Schilling, J., Bastiani, J. J., Makk, G. and Goodman, C. S. (1988). Characterization and cloning of fasciclin I and fasciclin II glycoproteins in the grasshopper. Proc. Natl. Acad. Sci. USA 85, 5291-5295. Steindler, D. A. (1993). Glial boundaries in the developing nervous system. Annu. Rev. Neurosci. 16, 445-470. Thomas, J. B., Bastiani, M. J., Bate, M. and Goodman, C. S. (1984). From grasshopper to Drosophila : a common plan for neuronal development. Nature 310, 203-207. Thomas, K. R. and Capecchi, M. R. (1990) Targeted disruption of the murine int-1 proto-oncogene resulting in severe abnormalities in midbrain and cerebellar development. Nature 346, 847-850. Tolbert, L. P and Oland, L. A. (1989). A role for glia in the development of organized neuropilar structures. Trends Neurosci. 12, 70-75. Xie, F., Meier, T. and Reichert, H. (1994). Coexpression and function of TERM-1 and fasciclin II during axonal outgrowth of identified brain interneurons in the grasshopper. Proc. R. Soc. London B 257, 199-204. Zacharias, D., Williams, J. L. D., Meier, T. and Reichert, H. (1993). Neurogenesis in the insect brain: Cellular identification and molecular characterization of brain neuroblasts in the grasshopper embryo. Development 118, 941-955. Zinn, K., McAllister, L. and Goodman, C. S. (1988). Sequence and expression of fasciclin I in grasshopper and Drosophila. Cell 53, 577-587. (Accepted 20 September 1994)