Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Mitochondrion wikipedia , lookup
Gaseous signaling molecules wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Beta-Hydroxy beta-methylbutyric acid wikipedia , lookup
Adenosine triphosphate wikipedia , lookup
Basal metabolic rate wikipedia , lookup
Lactate dehydrogenase wikipedia , lookup
Biochemistry wikipedia , lookup
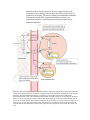
Muscle Energetics and Fatigue Joseph Feher, Ph.D. LECTURE OUTLINE: I. II. III. IV. Rate of ATP consumption by active muscle depends on the load. A. The free energy of ATP hydrolysis is the chemical source of mechanical work. B. ATP hydrolysis is linked to mechanical work through the actomyosin ATPAse. C. The rate of cross-bridge cycling and ATPase activity depend on the load. D. ATP hydrolysis is also needed for control reactions. Rate and amount of ATP consumption varies with the intensity and duration of exercise Muscle consumption of ATP is always fast A. Normal activation of muscle is by trains of impulses B. In vigorous exercise, frequency of muscle activation increases C. Each activation of the muscle requires fast ATP hydrolysis D. Intensity of exercise changes the rest period for metabolism to recover energy reserves Metabolism regenerates ATP in different time scales and capacities A. B. Direct phosphorylation regenerates ATP fastest. 1. Creatine phosphate provides the first buffer for ATP in muscle, but it increases Pi. 2. Myokinase can regenerate ATP from ADP. Glycolysis provides a rapid but low capacity supply of ATP for fast twitch fibers. C. Oxidative phosphorylation provides a slower but high capacity supply of ATP. V. The fuel used by muscle varies with intensity and duration of exercise VI. At high intensity, glucose and glycogen is the preferred fuel for muscle VII. VIII. A. Carbohydrates are stored as glycogen and mobilized by glycogenolysis B. Glycolysis uses muscle glycogen and blood glucose C. Glycolysis produces pyruvate, ATP and NADH without requiring oxygen D. Generation of lactic acid regenerates cytoplasmic NAD+ Muscle and muscle fibers can be classified by metabolic properties A. Muscles can be classified as SO, FOG and FG B. Whole muscles are mixtures of muscle fiber types C. Muscle fiber types differ in the isomers of many different proteins D. Muscle fiber types differ in the relative amount of organelles E. Muscle fiber types are more continuously distributed than suggested by the classification schemes Mismatch of lactic acid production and oxidation determines the rate of lactic acid release into the blood A. Blood lactate levels rise progressively with increases in exercise intensity B. Mitochondria can metabolize lactic acid C. Lactic acid forms a carrier system for NADH oxidation D. Lactic acid is produced by fully oxygenated tissue E. Three lactic acid shuttles remove lactate from its site of production 1. the intracellular shuttle removes lactate to the mitochondria 2. The cell-cell shuttle removes lactate to adjacent oxidative fibers 3. F. IX. X. The Cori cycle shuttles lactate to the liver for gluconeogenesis The “anaerobic threshold” really has to do with mismatch of lactic acid production and oxidation 1. The rest periods between contractions become shorter 2. The fast glycolytic fibers are increasingly recruited over oxidative fibers 3. Sympathetic stimulation increases the rate of glyocogenolysis Exercise increases glucose transporters in the muscle sarcolemma A. Glucose uptake by muscle depends on the number and activity of GLUT4 transporters B. Insulin increases the number of GLUT4 transporters by recruiting latent transporters C. Exercise increases the number of active GLUT4 transporters independent of insulin D. Mechanism of GLUT4 recruitment is probably through AMPK and CAMK The site of Fatigue depends on the muscle and intensity of exercise. A. Fatigue is a reduction in developed force. B. Metabolic fatigue occurs with repetitive stimulation C. Sites of fatigue could be in any process from brain to contractile element D. Fatigue during maximum sustained contractions is not in the brain. E. Pi and H+ in muscle interferes with force development by actomyosin ATPase. F. Fatigue at submaximal work rates depends on glycogen stores. 1. Fatigue accompanies glycogen depletion 2. XI. The importance of glycogen in exercise is evident from muscle glyogenoses XII. Training regimens are designed to increase strength of endurance A. The most rapid training involves training the brain B. Training for strength induces muscle hypertrophy C. XIII. XIV. Fatigue can be delayed by glycogen supercompensation 1. Muscle strength depends on muscle size 2. Training sends a signal for muscle growth a. Myostatin inhibits muscle growth b. Insulin-like growth factor-1 and muscle growth factor also influence muscle size Endurance training uses repetitive movements Our ability to switch muscle fiber types is limited A. Muscle capabilities are determined by the mixture of muscle fiber types B. Single muscle fibers can be hybrids C. It is unlikely that muscle types interconvert in human training regimens Practice Questions OBJECTIVES: 1. Describe how intensity of exercise differs in heavy resistance vs. endurance exercise 2. Explain why ATP hydrolysis in active muscle is always fast 3. List the sources of ATP generation in order of their rapidity 4. List the sources of ATP generation in order of their capacity 5. Describe how fuel use varies with intensity and duration of exercise 6. Explain how lactic acid production allows for more rapid glycolysis 7. List the types of muscle fibers based on metabolic properties 8. List the metabolic fates of lactic acid 9. Explain why exercise reduces diabetic’s need for insulin 10. Identify the likely site of fatigue in high-intensity, short-duration exercise 11. Identify the likely site of fatigue in moderate-intensity, long-duration exercise 12. Describe the overall changes that occur during hypertrophy 13. Describe the function of myostatin and the consequence of its lack 14. Explain how muscle hybrid types can arise 15. Describe our limited ability to switch muscle types Suggested Reading: Berne and Levy, pp. 237-241 I. MUSCULAR ACTIVITY CONSUMES ATP AT RATES DEPENDENT ON LOAD A. The free energy of ATP hydrolysis is the chemical source of mechanical work. In adenosine triphosphate ( ATP), energy is required to attach each of the phosphates on the ribose moiety. These bonds are less stable than their starting materials: The γ bond makes ATP less stable than ADP and Pi ; the β Pi bond makes ADP less stable than AMP and Pi. When these bonds are hydrolyzed, chemical energy becomes available for work with the remainder dissipated as heat. B. ATP hydrolysis is linked to mechanical work through the myosin ATPase. The cross-bridge cycle links shortening or force production to the hydrolysis of ATP through the kinetics of the actin and myosin interactions. In an intact muscle, macroscopic force results from thousands of these cross-bridges cycling at furious rates. Figure 1. Structures of ATP and ADP and the ATP hydrolysis reaction. Under the conditions of the myoplasm [ATP] = 5 x 10-3M, [ADP] = 0.04 x 10-3 M, [Pi] = 5 x 10-3 M and pH = 7.0, hydrolysis of ATP liberates approximately 57 kjoules mol-1 C. The rate of cross-bridge cycling and ATPase activity depend on the load. Unlike smooth muscle, which can generate force without large amounts of ATP hydrolysis, skeletal muscle hydrolyzes ATP whether or not it shortens. Isometric contractions consume ATP because the cross bridges continue to cycle even when shortening does not occur. Eccentric contractions also require ATP hydrolysis to resist stretch and thereby impart rigidity to the joints. But fewer muscle fibers are recruited in eccentric contractions, because more force results from the eccentric contractions. Thus, the rate of ATP consumption for whole muscles varies with the speed and type of contraction. D. ATP hydrolysis is also needed for control reactions. Muscles also use ATP to re-accumulate activator Ca2+ into the SR, for active ion pumping by the Na+-K+-ATPase , and for maintenance chores. During activity, the actomyosin ATPase is the main cause of ATP hydrolysis. II. RATE AND AMOUNT OF ATP CONSUMPTION VARIES WITH THE INTENSITY AND DURATION OF THE EXERCISE The rate of ATP utilization by the aggregate muscles of the body depends on the intensity of the exercise. The total amount of ATP used during a bout of exercise is its rate of utilization times the duration of the event. High intensity exercise can be sustained only for short periods, whereas moderate intensity exercise can be endured for long times. The relationship between intensity and sustainable effort is not linear. Table 1 below lists the approximate rates and amounts of ATP needed for different track events. Rate of ATP consumption Amount of ATP needed (mol/min) (mol) Rest 0.07 ---100 m sprint 2.6 0.4 800 m run 2.0 3.4 1500 m run 1.7 6 42200 m marathon 1.0 150 Table 1. Rate and amount of ATP needed for different track events. Event Adapted from Hultman, E. and Sjoholm, H. Biochemical causes of fatigue, in “Human Muscle Power” (1986) Human Kinetics Publishers, Inc, Champaign Illinois. III. MUSCLE CONSUMPTION OF ATP IS ALWAYS FAST WHEN THE MUSCLE IS ‘ON’ A. Normal activation of muscle is by trains of impulses The motor neuron carries a single code for activating its muscle fibers: the temporal pattern of its action potentials. For particular movements, motor units in groups of muscles are activated in a particular sequence to coordinate the movement. Figure 2 shows an electromyogram (EMG) of rat leg muscles obtained at a slow walking speed. The EMG records electrical activity of the muscle, not force. Each muscle is activated at appropriate times for a definite length of time, and this activity alternates with periods of rest. B. In vigorous exercise, frequency of muscle activation increases The EMGs in Fig. 2 illustrate muscle activation in slow walking. In vigorous walking, the frequency of activation increases and the duration of each activation decreases. The duty cycle, the fraction of time that the muscle is activated, increases with increased intensity. Many animals engage different sets of muscle coordination, called gaits, to vary their speed of locomotion. In humans, walking and running form distinctive gaits, but running fast and running slowly differ mainly in the speed and recruitment rather than the sequence of activation. Weight lifting differs from running in that it activates a selected few muscles. Increasing the intensity of weight lifting means increasing the weight, and this is achieved by increasing recruitment of the muscle fibers until 100% recruitment is reached at the maximum weight. The maximum weight can be lifted only slowly, so the muscles must be activated by a train of impulses that last as long as it takes to lift the weight. This differs from the activation pattern of repetitive exercise such as walking or running in that in weight lifting there is no rest phase for the muscle fibers until the exercise is over. This is the origin of the different kind of fatigue for heavy resistance exercise versus endurance exercise. Figure 2. Electromyogram of rat leg muscles at a slow walking speed, 1 mph, on a treadmill. Modified from G.A. Brooks, T.D. Fahey, T.P. White and K.M. Baldwin, Exercise Physiology, Third Edition, McGraw Hill, 1999. C. Each activation of the muscle requires fast ATP hydrolysis Larger EMG amplitudes (recorded in volts) indicate greater number of muscle fibers that are firing action potentials. Each muscle fiber, when activated, is activated completely. The control of force for the entire muscle is achieved by the temporal recruitment of the fibers: which fibers are being activated and with what frequency and in what sequence. Each activation of a muscle fiber entails fast rates of ATP hydrolysis because all of the actomyosin cross-bridges are activated by the Ca2+ transient with each action potential on the motor neuron. D. Intensity of exercise changes the rest period for metabolism to recover energy reserves Increasing intensity of exercise increases the overall frequency at which individual muscle fibers are activated. Each activation entails furious rates of ATP hydrolysis. Increasing the frequency of activation necessarily decreases the time that the muscle is not activated, and this is the time the muscle has for metabolism to oxidize substrates produced during the contractions. Thus, every muscle contraction requires rapid ATP consumption and regeneration, but at low intensity exercise there is more time for metabolism to recover the resting state. IV. METABOLISM REGENERATES ATP IN DIFFERENT TIME SCALES AND CAPACITIES (See Figure 3 and Table 2) A. Direct phosphorylation re-generates ATP fastest. 1. Creatine phosphate provides the first buffer for ATP in muscle, but it increases Pi. Muscles contain about 15-20 mM creatine phosphate. Creatine phosphokinase (CPK) catalyzes the phosphorylation of ADP from creatine phosphate to form ATP. This extremely rapid reaction helps to “buffer” ATP concentrations near the normal 5 mM in the muscle fiber myoplasm. When creatine phosphate is used to regenerate ATP, myoplasmic [Pi] increases. Creatine phosphokinase is located nearby ATP-utilizing reactions such as the myosin ATPase and SR Ca-ATPase and may directly transfer ATP to these enzymes. This is called substrate channeling. These ATPase enzymes split ATP faster in the presence of CPK. All contractions of muscle require creatine phosphate regeneration of ATP for maximum force. 2. Myokinase can re-generate ATP from ADP. Another enzyme, myokinase, converts two molecules of ADP into ATP and AMP. AMP levels may be what is sensed by the “fuel gauge” of muscle fibers. When it goes up, you are running out of fuel. B. Glycolysis provides a rapid but low capacity supply of ATP for fast twitch fibers. Glycolysis begins with glucose that may arise from glycogen or from the blood, and ends with two pyruvate molecules. Glycolysis requires 2ATP molecules and generates 4ATP, for a net gain of only 2 ATP molecules per molecule of glucose. Additional ATP (4 or 6 per molecule of glucose) can be generated from the NADH produced by the oxidation of glyceraldehyde-3- phosphate during glycolysis. C. Oxidative phosphorylation provides a slower but high capacity supply of ATP. Cytosolic pyruvate formed by glycolysis enters the mitochondria to be converted to acetyl CoA by pyruvate dehydrogenase. In this process, 1 CO2 is released and reducing equivalents as NADH are produced. The acetyl CoA is converted to 2 more CO2 molecules through the TCA cycle, which also produces NADH, FADH2 and GTP. In the presence of oxygen, the NADH and FADH2 is oxidized through the Electron Transport Chain (ETC). The ETC pumps H+ ions out of the mitochondrial matrix, establishing a [H+] gradient and an electrical potential across the inner mitochondrial membrane. This electrochemical gradient for H+ is used by the mitochondrial ATP synthase to synthesize ATP from ADP and Pi. Net ATP production from the complete oxidation of pyruvate is 30 ATP molecules per molecule of glucose. Oxygen is needed as the final electron acceptor from the ETC. Without oxygen, the ETC remains reduced and everything backs up. The TCA stops for lack of NAD+, and beta oxidation of fats stops for the same reason. However, lack of oxygen is pathological rather than physiological. In normal physiology, the issue is how fast oxidative phosphorylation is going with respect to ATP consumption. V. THE FUEL USED BY MUSCLE VARIES WITH INTENSITY AND DURATION OF EXERCISE Muscles can use fats, carbohydrates and proteins as fuels. Which is used at what rates depends on the type, intensity and duration of exercise. At rest, muscles use mainly free fatty acids. At moderate exercise (< 50% maximum O2 consumption (VO2)), muscle use blood glucose and free fatty acids. At higher intensities of exercise (>50% VO2) the proportion contributed by glycogen becomes increasingly important so that at 70-80% VO2 aerobic metabolism of glycogen is predominant. Figure 3. Overall energy metabolism driving contraction in skeletal muscle. ATP is consumed in a variety of reactions including the actomyosin cross-bridges and the SR Ca-ATPase pump. ATP is provided by a variety of routes including glycolysis and complete oxidation of carbohydrates through the TCA cycle and electron transport chain (ETC) in the mitochondria. The source of glucose for glycolysis can be muscle glycogen or plasma glucose. The glucose is imported into the muscle by a glucose transporter, GluT4. Plasma glucose originates from liver and extrahepatic tissues either through glycolysis (liver) or gluconeogenesis (liver, kidneys, intestine). Fatty acids form acetylCoA through beta oxidation and the acetyl CoA is then completely oxidized, in the presence of adequate oxygen, in the mitochondria. These fatty acids may derive from muscle or from adipose stores. When glycolytic flux is rapid and myoplasmic NADH accumulates, glycolysis continues by the regeneration of NAD+ by converting pyruvate to lactic acid by lactate dehydrogenase (LDH). Production of lactic acid thereby allows glycolysis to continue. Lactic acid produced in this way is transported into the blood and from there to the liver where it can be converted to glucose again. This cycle of muscle glucose to lactate to liver lactate to glucose is the Cori cycle. Rate of ATP production Amount of ATP available (mol/min) (mol) ATP and creatine phosphate 4.4 0.7 Glycogen to lactate 2.4 1.6 Muscle glycogen to CO2 1.0 84 Liver glycogen to CO2 0.4 19 Fatty acids to CO2 0.4 4000 Table 2. Rate and amount of ATP available for contraction from various fuel sources Source of energy Table 2 shows the rates of ATP production and amounts of ATP available from various sources. It is important to remember that every muscle contraction utilizes creatine phosphate and glycogen to lactate, but at low frequency the oxidation of lactate or blood glucose during the rest period pays to resynthesize glycogen. VI. AT HIGH INTENSITY, GLUCOSE AND GLYCOGEN IS THE PREFERRED FUEL FOR MUSCLE A. Carbohydrates are stored as glycogen and mobilized by glycogenolysis. Muscle cells burn glucose, but the amount of free glucose in the blood is limited and cannot fuel muscle activity alone. Muscles and liver store carbohydrates as glycogen. Glycogen is mobilized through glycogenolysis to provide glucose for muscle activity. Glycogenolysis is controlled by sympathetic nervous activity and circulating epinephrine. It is mediated by phosphorylase, and is controlled by a Gs protein linked to adenylyl cyclase, the production of cAMP and the activation of protein kinase A. The rapid utilization of ATP in normal contractions appears to require glycogenolysis. Glycogen is re-generated during the resting phase of the muscle between trains of impulses. B. Glycolysis uses muscle glycogen and blood glucose. Glycogen stored in muscle is dedicated to glycolysis because muscle lacks glucose 6-phosphatase that converts G-6P to glucose. Only glucose can cross the cell membrane. Ionically charged G-6P cannot. Because it lacks the enzyme to make free glucose, muscle cannot export significant glucose. Liver and other tissues produce glucose that can travel to muscle through the blood. Muscle tissues take up glucose by a transporter, Glut4, that is sensitive to exercise. The Glut4 transporter is recruited to the cell membrane by insulin, but exercise also recruits these transporters in the absence of insulin. This is why diabetics cut back on their insulin when they exercise. C. Glycolysis produces pyruvate, ATP and NADH without requiring oxygen. Glycolysis produces a net gain of 2 molecules of ATP per molecule of glucose. It also produces NADH from NAD+ as an obligatory cofactor for the reaction of glyceraldehyde-3-phosphate to 1,3-diphosphoglycerate. If the cell runs out of cytoplasmic NAD+, glycolysis will stop. Generating ATP without requiring oxygen is called anaerobic metabolism. D. Generation of lactic acid regenerates cytoplasmic NAD+ As noted above, glycolysis requires NAD+ in order to proceed. Cytoplasmic NADH produced by glycolysis can be oxidized back to NAD+ by the mitochondria through shuttle systems (the malate shuttle and the glycerolphosphate shuttle) that transfer reducing equivalents (NADH) into the mitochondrial matrix. Conversion of NADH to NAD+ requires an oxidized electron transport chain. During rapid bursts of glycolysis, the mitochondria cannot keep up with the NADH generated by glycolysis. In these rapid bursts, both NADH and pyruvate concentrations momentarily spike. Lactic dehydrogenase converts pyruvic acid to lactic acid, simultaneously converting NADH to NAD+. This NAD+ can then be used to allow glycolysis to proceed at the glyceraldehyde 3-phosphate step. Thus, lactic acid production allows glycolysis to proceed during rapid glycolytic bursts of ATP production, but the reaction goes faster in exercise because the concentrations of its substrate, pyruvate, increases. VII. MUSCLES AND MUSCLE FIBERS CAN BE CLASSIFIED BY THEIR METABOLIC PROPERTIES A. Muscles can be classified as slow oxidative (SO), fast glycolytic (FG) and fast oxidative-glycolytic (FOG). Muscles can be classified by their mechanical properties (see Skeletal Muscle Mechanics) and by their myosin staining (see Contractile Mechanisms). Muscle fibers can also be classified on the basis of their metabolic capabilities. Peter and coworkers described three types of fibers: slow oxidative (SO); fast glycolytic (FG) and fast oxidative-glycolytic (FOG). The three main classification schemes are shown in Table 3. Muscle Property Used to Classify Types Burke mechanical properties Brooke myosin ATPase staining Peter metabolic capacity Table 3. Muscle Fiber Type Classification Schemes. Classification Scheme B. Fiber Types S, FR, FI, FF I, IIA, IIB, IIC SO, FOG, FG Whole muscles are mixtures of muscle fiber types. Whole muscles consist of thousands of muscle fibers and these muscle fibers are distributed among the various muscle fiber types. Specific muscles may be predominately one type or another, and the distribution of fiber types in a given muscle varies between individuals. C. Muscle fiber types differ in the isoforms of many different proteins. Brooke’s classification scheme is based on the expression of different myosin isoforms in skeletal muscle. Many other proteins also can be expressed as one of several different isoforms. The SERCA Ca-ATPase has a fast-twitch (SERCA1a) and slow-twitch isoform (SERCA2a); calsequestrin within the SR lumen has at least two different isoforms, a “fast” type and a “cardiac” type. The RyR has different isoforms (RyR1 in skeletal muscle and RyR2 in cardiac muscle). TnC is expressed in different forms in fast skeletal (TnC2) and slow twitch skeletal muscle and cardiac muscle (TnC1). Why muscles have some of these isoforms is not yet clear. Table 4 compares some of the different proteins expressed in different muscle types and the relative abundance of selected organelles. D. Muscle fiber types also differ in the relative amount of organelles. Oxidative fibers contain a lot of mitochondria compared to glycolytic fibers. The relative amounts of the SR also vary depending on the speed of contraction. Slow twitch fibers generally have about one-half as many SR Ca2+ pumps as fast twitch fibers. The myoplasmic parvalbumin content correlates well with the speed of the fibers, whereas the myoglobin content correlates well with its oxidative capacity. Type I Muscle Type IIa Muscle Twitch Slow Fast Fatigue Resistant Resistant Metabolism Oxidative Oxidative Mitochondria +++ ++++ SR volume ++ +++ Glycogen + +++ Myosin Heavy Chain MHC-I MHC-IIa Myosin Light Chain MLC-1aS, -1bS MLC-1f, -3f SR Ca-ATPase SERCA2a SERCA1a Phospholamban ++ Calsequestrin fast and cardiac fast RyR RyR1 RyR1 Troponin C TnC1 TnC2 Myglobin +++ +++ Paravalbumin + Table 4. Comparison of different muscle types. Type IIb Muscle Fast Fatigable Glycolytic + ++++ ++++ MHC-IIb, -IIx MLC-1f, -3f SERCA1a fast RyR1 TnC2 ++ Cardiac Muscle Resistant Oxidative ++++ + ++ MHC-α, MHC-β MLC-1v, -1a SERCA2a + cardiac RyR2 TnC1 +++ - VIII. MISMATCH OF LACTIC ACID PRODUCTION AND LACTIC ACID OXIDATION DETERMINES THE RATE OF LACTIC ACID RELEASE A. Blood lactate levels rise progressively with increases in exercise intensity Progressive increases in exercise intensity causes a progressive rise in blood lactic acid, as shown in Fig. 4. The increased circulating lactic acid is caused by release of lactic acid from the working muscles. Originally lactic acid was thought to be produced during anaerobic metabolism, so that the increased levels in the blood was thought to represent increased reliance on anaerobic metabolism. For this reason, the knee in the curve was called the “anaerobic threshold”. Newer views suggest that the tissue is not anaerobic, even though lactic acid production is increased. Figure 4. Blood lactate concentration as a function of relative work load. Lactate levels in blood increase only gradually until about 60% of VO2 max is reached, and then lactate concentration increases markedly with further increases in exercise intensity. B. Mitochondria can metabolize lactic acid Mitochondria possess a monocarboxylic acid transporter (MCT1) that transports lactic acid into the mitochondria. Mitochondria also possess lactic dehydrogenase, LDH, that converts lactic acid to pyruvate. Lactate enters the mitochondria over MCT1 (Monocarboxylic acid transporter) and is converted back to pyruvate by mitochondrial LDH. The pyruvate is then oxidized by the TCA cycle. C. Lactic acid forms a carrier system for NADH oxidation Because lactic acid can enter the mitochondria and be converted back to pyruvate, it carries cytoplasmic reducing equivalents, as NADH, into the mitochondria (Fig. 5). NADH is converted to NAD+ by LDH in the cytoplasm, lactic acid crosses over to the mitochondria and converts NAD+ to NADH in the matrix. The lactic acid, converted to pyruvate, is then consumed by the mitochondria. Figure 5. Lactic acid carries reducing equivalents into the mitochondria. Cytosolic NAD+ is an obligatory requirement for glycolysis. Conversion of pyruvate to lactic acid in the cytoplasm regenerates NAD+ so that glycolysis can continue. The lactic acid enters the mitochondria over the MCT1 carrier (which also transports pyruvate) and is converted back to pyruvate in the mitochondria by mitochondrial lactate dehydrogenase (LDH). The NADH generated in the mitochondria can be oxidized back to NAD+ by the electron transport chain. D. Lactic acid is produced by fully oxygenated tissue The classical view is that lactate is produced only under anaerobic conditions, when muscle PO2 falls below levels that fully energize mitochondria. This view is now thought to be wrong. The main fact that doesn’t fit is that lactate is produced by exercising muscles that are fully oxygenated. E. Three lactate shuttles remove lactate from the site of production 1. The intracellular shuttle removes lactate to the mitochondria in the same cell Lactate produced in the cytosol moves into the mitochondria of the cell where the lactate is converted to pyruvate and oxidized by the TCA cycle coupled to oxidative phosphorylation. 2. The cell-cell shuttle removes lactate to adjacent oxidative cells The fastest fibers produce lactate at the highest rate. They first become unable to oxidize all of the lactic acid themselves, and the lactate enters the blood. Neighboring oxidative fibers, which are generally smaller than the large glycolytic fibers, take up some of this lactate and oxidize it. This constitutes the cell-cell shuttle (Fig. 6.) 3. The Cori cycle shuttles lactate to the liver for gluconeogenesis The liver takes up lactate that is released into the blood by the active muscles. The liver either metabolizes the lactate for energy or uses it to make new glucose through gluconeogenesis, and exports the glucose into the blood. Muscles can then take up this glucose and use it again for energy. This cycle of blood glucose to muscle lactate to blood lactate to liver lactate and back to blood glucose is called the Cori cycle. F. The “anaerobic threshold” really has to do with mismatch of lactic acid production and oxidation The increases in blood lactate with intensity of exercise is caused by the release of more lactic acid by the exercising muscles than can be metabolized by the aggregate tissues of the body. Release outstrips oxidation plus gluconeogenesis. This has the appearance of an increase of anaerobic metabolism, and in one sense it is. Every muscle contraction involves a period of “anaerobic” generation of ATP. In less intense exercise, there is sufficient rest time for the lactic acid produced during this period to be oxidized. When exercise intensity increases, three different things happen: 1. The rest period between contractions becomes shorter 2. The fast glycolytic fibers are increasingly recruited over the oxidative 3. Sympathetic nervous system increases the rate of glycogenolysis, further increasing the supply of pyruvate, and, by mass action, the production of lactate Because of these, lactate release by the active muscles soars with increased exercise intensity and outstrips the ability of the tissues to metabolize the lactate. This occurs without gross anaerobic conditions. The muscle are still fully oxygenated. In this sense there is no anaerbiosis yet there is increased periods of lactate production by anaerobic pathways. Figure 6. The lactate shuttles. Lactate is produced in the cytoplasm by LDH acting on pyruvate and NADH. Lactate can be shuttled from the cytoplasmic compartment to the mitochondrial compartment by importing the lactate into the mitochondria and linking it to synthesis of pyruvate and generation of NADH in the mitochondrial matrix. This is the intracellular lactate shuttle. Secondly, lactate can be exported into the blood where it is taken up by adjacent oxidative muscle fibers and completely oxidized by its mitochondria. This is the cell-to-cell lactate shuttle. Third, lactate released into the blood when lactic acid production is high can be taken up by liver cells (also called hepatocytes). The hepatocytes resynthesize glucose from the lactate and export it back into the blood where it can be taken up by the exercising muscle, for example. This is the Cori cycle. IX. EXERCISE INCREASES GLUCOSE TRANSPORTERS IN THE MUSCLE SARCOLEMMA A. Glucose uptake by muscles depends on the number and activity of GLUT4 transporters One of the important fuels for muscle is blood glucose, which originates mainly from the liver, intestine and kidney through glycogenolysis or gluconeogenesis. Blood glucose enters the muscle fibers through specific GLUT4 transporters in the muscle fiber membrane. The rate of uptake depends on the number of these transporters in the membrane and their activity. B. Insulin increases the number of GLUT4 transporters by recruiting latent transporters One of the most important effects of insulin is to increase the uptake of glucose by the peripheral tissues, especially muscle. Insulin increases glucose uptake by recruiting GLUT4 transporters from latent storage in vesicles in the muscle fiber. This mechanism is shown in Fig. 7. C. Exercise increases the number of active GLUT4 transporters independent of insulin Exercise itself exerts an insulin-like effect and increases glucose uptake by increasing the number of GLUT4 transporters, but without increases in insulin. Diabetic persons who inject insulin and then exercise could experience too much glucose removal and possibly suffer from hypoglycemia. For this reason, diabetics should reduce their injection of insulin when they anticipate they will exercise. D. Mechanism of GLUT4 recruitment is probably through AMPK and CAMK The mechanism by which exercise increases GLUT4 transporters is not yet completely worked out. However, researchers believe than AMPK, a protein kinase stimulated by AMP, and calmodulin-dependent protein kinase, CAMK, may be involved. AMPK is stimulated by AMP, which is produced from ADP by myokinase when ADP concentrations rise during contractions. The AMPK is thought to be a kind of “fuel gauge” that senses low fuel levels and then switches off ATP-consuming reactions and turns on ATP-producing reactions. AMP acts in a negative feed-back mechanism to restore ATP levels. CAMK, on the other hand, is activated by Ca2+ when it rises to activate the myofilaments. If it simultaneously activates GLUT4 transporters, it acts as a feed-forward mechanism to begin increasing ATP supply in anticipation of its need to support contraction. Figure 7. Insulin, AMPK and CAMK increase GLUT4 incorporation into the sarcolemma of muscle fibers. All three increase the number of sarcolemma GLUT4 from a population of latent transporters located in vesicles in the cell. The increased numbers of GLUT4 transporters increase glucose uptake and generation of ATP through glycolysis and oxidation of pyruvate or lactate. X. THE SITE OF FATIGUE DEPENDS ON THE MUSCLE AND INTENSITY OF EXERCISE A. Fatigue is a reduction in developed force. Fatigue is a transient loss of work capacity resulting from preceding work. Reduction in the maximal force from a muscle, compared to its rested force, is viewed as fatigue. By this criteria, fatigue sets in very soon after maximum effort. Usually a person can bench press a maximal weight only once. B. Metabolic fatigue occurs with repetitive stimulation. A second kind of fatigue follows exercise at submaximal force for many repetitions. After some time, we tire and eventually become unable to develop even this submaximal force. C. Sites of fatigue could be in any process in the chain from brain to contractile element. In principle, impairment of any one of the chain of events starting with the central nervous system and ending with the contractile elements of muscle could reduce developed force. These events include excitatory drive to the higher motor centers (motivation or effort), balance between excitatory and inhibitory pathways in spinal motor neurons; conduction of action potential on the motor neuron to the neuromuscular junction; transmission across the neuromuscular junction; propagation of the muscle action potential over the SL and into the T-tubules; excitation-contraction coupling; generation of force at the actin and myosin filaments. D. Fatigue during maximum sustained contractions in humans is not in the brain. In classic experiments, Merton tested the adequacy of central nervous system activation of fatiguing muscles by directly stimulating the nerve leading to a muscle during maximal voluntary contractions. If the voluntary contraction was maximal, the electrical stimulation would not increase the force. He found no increase in force upon external stimulation and concluded that the CNS is not the cause of fatigue. Direct stimulation of the muscle also did not increase force of fatigued muscle, suggesting that the neuromuscular junction failure also does not cause fatigue. E. Pi and H+ in muscle interferes with force development by actomyosin ATPase. In fast twitch fibers activated for short bursts, creatine phosphate regenerates ATP from ADP. The terminal phosphate comes from the creatine phosphate, so that regeneration of ATP increases Pi in the myoplasm. At the same time, activation of anaerobic glycolysis produces ATP with a build up of lactic acid and H+ ions. During exercise the pH of muscle can fall from pH 7.0 to pH 6.0. Both Pi and H+ interfere with force production, by directly inhibiting the acto-myosin ATPase or by making the myofilaments less sensitive to activator Ca2+. The resulting reduction in force is perceived as fatigue. F. Fatigue at submaximal work rates depends on glycogen stores. 1. Fatigue accompanies glycogen depletion The performance times at high but submaximal work loads depends on the size of the glycogen stores before exercise. Fatigue appears when glycogen levels fall but before they are zero. This has led to the hypothesis of the “glycogen shunt” in which glycogenolysis is necessary to maintain ATP during contraction, and is resynthesized during the rest period between contractions. When glycogen becomes low, it can no longer sustain ATP levels during contraction and force falls, even though glycogen is not completely used up. 2. Fatigue can be postponed by glycogen supercompensation Glycogen stores can be increased by a combination of exercise and carbohydrate consumption. This is referred to as carbohydrate loading. It is usually accomplished by exhaustive exercise followed within 2 hours by a high-carbohydrate meal. Under these conditions the glycogen stores supercompensate and store larger than normal amounts of glycogen. For serious athletic contests such as marathons the race preparation is more complicated, taking place over the two weeks prior to the race. XI. THE IMPORTANCE OF GLYCOGEN IN EXERCISE IS EVIDENT FROM MUSCLE GLYCOGENOSIS (Figure 8) There are 11 well known hereditary disorders of glycogen or carbohydrate metabolism that affect muscle alone of together with other tissues. These disorders cause two major clinical syndromes: A. acute, recurrent, reversible muscle dysfunction manifesting as exercise intolerance, myalgia upon exercise, and cramps, often culminating in muscle breakdown and myoglobinuria B. fixed, often progressive weakness, sometimes simulating dystrophic, inflammatory or even neurogenic processes Fig. 8 illustrates the points in metabolism characterized by these glycogenoses. Figure 8. Muscle Glycogenoses. XII. TRAINING REGIMENS ARE DESIGNED TO INCREASE STRENGTH OR ENDURANCE A. The most rapid training involves training the brain. During initial training (the first few weeks) the maximal voluntary contraction increases whereas the maximal evoked contraction (produced by direct and maximal stimulation of the motor nerve) does not. This suggests that trainees learn to activate their muscles more fully or they improve coordination of the voluntary contraction. Increases in maximal evoked contraction require a longer training period. B. Training for strength induces muscle hypertrophy. 1. Muscle strength depends on muscle size. The maximum force that can be exerted by a muscle depends on its cross-sectional area and architecture such as pinnation. Strength training employs contractions against large resistances with few repetitions. It is called resistance training. 2. Training sends a signal for muscle fiber growth. Beginning training is associated with delayed onset muscle soreness, or DOMS. Part of the soreness could be due to microscopic tears in muscle fiber membranes or connective tissue or to the stretching of sensory neurons from edema or mechanical stretch. The exercise signals muscle hypertrophy: the diameter of the fibers increases but the number of fibers stays the same. Although the dogma states that training increases the size of muscle fibers but not their number, some electron micrographs of muscles appear to show muscle fibers splitting length wise. Some limited amount of hyperplasia (increase in cell number) may also occur. Both type I and type II fibers hypertrophy in response to resistance training. Hypertrophy occurs two ways: muscle fibers make more myofibrils and satellite cells within the muscle are recruited to fuse with existing muscle fibers to help control the extra cytoplasm. During development, satellite cells are recruited to form myotubes that further differentiate to become muscle fibers. a. Myostatin inhibits muscle growth Stretch, hypoxia, and intracellular [Ca2+] signal muscle cell nuclei to produce a host of transcription factors that increase synthesis of myofibrillar proteins. One product that inhibits muscle differentiation and growth is myostatin. Myostatin is an autocrine and paracrine hormone produced by muscle cells that is a negative regulator of muscle mass. Watch the newspapers for athletic abuse of myostatin inhibitors. People with nonfunctioning myostatin mutations have gross muscle hypertrophy (N Engl J Med 350:2682-2688, 2004). b. Insulin-like growth factor-1 and muscle growth factor (MGF) also influence muscle size Other muscle growth factors include insulin-like growth factor-1 (IGF-1) and muscle growth factor (MGF). Muscles have receptors for IGF-1 that activate the cascade leading from phosphatidyl inositol 3-kinase (PI3K) to activation of protein kinase B (PCB-ACK) and mammalian target of rapamycin (mTOR). This is inhibited by cAMP-dependent protein kinase when AMP levels rise during hypoxia, for example. Figure 9 illustrates the likely signaling pathways for muscle hypertrophy. 3. Strength training decreases relative mitochondrial volume. Heavy resistance training results in a reduction in the mitochondrial volume density and the ratio of mitochondrial to myofilament volume. Strength training appears to have no effect on the muscle capillarity. C. Endurance training uses repetitive movements. Endurance training increases the capillarity of muscles and tunes the muscles’ metabolic capabilities. Concentrations of myoglobin and TCA cycle enzymes are increased as well as both the size and number of mitochondria. Muscles of endurance trained subjects use fats as the primary fuel for moderate exercise, thereby sparing glycogen for bursts of high intensity activity. Figure 9. The signaling events in muscle hypertrophy. Muscle grows in response to stretch, increases in the integrated cytoplasmic [Ca2+], androgens and glucocorticoids and other cytokines. Calcium activates calcineurin, a protein phosphatase that dephosphorylates NFAT (nuclear factor of activated T cells) and activates it. Myostatin produced by muscle inhibits satellite cell division and differentiation. Muscle cells also respond to IGF-1 (insulin-like growth factor-1) and MGF (muscle growth factor). XIII. OUR ABILITY TO SWITCH MUSCLE FIBERS TYPES IS LIMITED A. Muscle capabilities are determined by the mixture of muscle fiber types Earlier we saw that muscles are heterogeneous mosaics of different muscle fiber types, and that the overall mechanical and metabolic performance of the muscles is a consequence of the composition of that mosaic. One basis of this heterogeneity is the expression of specific myosin isoforms. There are at least 20 structurally distinct classes of myosin heavy chains. Eleven of these are expressed in adult mammalian muscles, but some are specific to one muscle. The most common isoforms are MHCIb, MHCIIa, MHCIIb and MHC IId. B. Single muscle fibers can be hybrids Immunohistochemistry reveals that some muscle fibers are “pure” types that express only a single myosin heavy chain (MHC) isoform. However, some muscle fibers contain two or more isoforms and are “hybrids”. How can this be? Recall that each muscle fiber contains many nuclei, typically located at the periphery of the cell near the sarcolemma, as shown in Fig. 10. Each nucleus controls a volume of cytoplasm or surface of the fiber called its “nuclear domain”. A separate population of nuclei congregate near the neuromuscular junction. During transitions between fiber types, it is possible that some of these nuclei receive different signals than others, and therefore transcribe different genes for the expression of myosin. In this way, hybrid muscle fiber types arise. The existence of hybrid muscle fibers allows a more continuous gradation between muscle types, as shown in Table 5. Figure 10. Nuclear domains in muscle fibers. Muscle fiber nuclei are located in the periphery of the fiber, nearly aligned in rows with more or less regular spacing, with some 35-80 nuclei per mm of fiber. Each nucleus controls protein expression in a volume or surface element called its domain. Muscle Fiber Type Type I Pure Fiber Myosin Heavy Chain Expression MHCI MHCI>MHCIIa Hybrid MHCIIa>MHCI Type IIa Pure Fiber MHCIIa MHCIIa>MHCIIx Hybrid MHCIIx>MHCIIa Muscle Fiber Description Slow Fast Fatiguable Type IIb Pure Fiber MHCIIx Fast Fatiguable Table 5. The muscle fiber type continuum. Hybrid muscle fibers allow transitional forms intermediate between Types I, IIa, and IIb. Humans do not make MHCIIb as in experimental animals, so the human form in fast fatiguable muscles is named MHCIIx. C. It is unlikely that muscle types interconvert in human training regimens. Is it possible for humans to convert a type I slow fiber into a type II fast fiber, or vice-versa? In cross-innervation experiments in animals, a fasttwitch muscle is removed from its bed and transplanted to a slow-twitch muscle bed, and a slow-twitch muscle is transplanted to a fast-twitch bed. The muscles in these cases are converted part way from slow twitch to fast twitch, and vice-versa. This demonstrates that it is the pattern of neural stimulation that determines muscle type. Chronic low frequency stimulation of fast twitch fibers increases the expression of proteins normally expressed only by slow-twitch fibers. Denervation or muscle unloading increase the levels of proteins normally expressed by fast-twitch fibers. The evidence for human transformation of muscle types is inconclusive. It appears that the stimulation of muscle necessary to transform the fiber types is so severe that no human can train that hard. The scientific consensus is that the transformation of muscle types is limited in part by the original position of the muscle on the muscle fiber type continuum. The transformation by exercise is always towards a slower type of muscle, but frank conversion of Type IIB fiber to a Type I fiber does not occur. D. The mechanism of muscle type switching appears to involve calcineurin Calcineurin is a calcium-dependent protein phosphatase that is located in the Z disks of skeletal muscle. With continued activity, the integrated [Ca2+] in muscle cells increases, which may signal calcineurin activation. It cleaves a phosphate off NFAT (nuclear factor of activated T cells) that migrates to the nucleus and begins switching off transcription of DNA coding for MHCIIa mRNA, and switches on the gene for MHCI. The result is a switching of muscle fiber types. (See Fig. 11). Figure 11. Mechanism of fiber type switching. XIV. PRACTICE QUESTIONS 1. 2. The main source of ATPase activity in exercising muscle is A. The SR Ca-ATPase B. The Na-K-ATPase C. Acto-myosin ATPase D. Myokinase E. Creatine kinase Because of lack of glucose 6 phosphatase A. Muscles can import glucose from the blood B. Liver cells can contribute to blood glucose homeostasis 3. 4. 5. C. The Cori cycle can occur D. Muscles cannot contribute substantially to blood glucose homeostasis E. Muscles cannot participate in glycolysis Fatigue in high-intensity, brief exercise is thought to be caused by A. A build up of Pi and H+ in the myoplasm B. Lactic acid C. Failure of T-tubule transmission D. Depletion of glycogen E. Failure of neuromuscular transmission When people with insulin-dependent diabetes exercise, they reduce their insulin shots because A. Exercise increases glucose uptake by muscle B. Exercise increases insulin secretion C. Exercise increases glycogenolysis D. Exercise increases lactic acid production E. They do not do this, tricky question person Lactic acid levels in blood A. Increases dramatically at 60-70% VO2 max because muscles become anaerobic B. Increases only when blood supply to muscle is compromised C. Is normally close to zero D. Explains most of short-term, high intensity fatigue E. Increases at 60-70% VO2 max because glycolysis increases dramatically and lactic oxidation cannot keep up 6. Persons with low levels of myostatin should A. Have predominately Type I muscle fibers B. Show exercise intolerance C. Have predominately Type II muscle fibers D. Have larger muscles than normal E. Have smaller muscle than normal Answers: 1C; 2D; 3A; 4A; 5E; 6D