Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Signal transduction wikipedia , lookup
Tissue engineering wikipedia , lookup
Extracellular matrix wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cell culture wikipedia , lookup
List of types of proteins wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
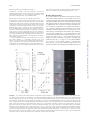
Cutting Edge: Eotaxin Elicits Rapid Vesicular Transport-Mediated Release of Preformed IL-4 from Human Eosinophils This information is current as of June 17, 2017. Christianne Bandeira-Melo, Kumiya Sugiyama, Lesley J. Woods and Peter F. Weller J Immunol 2001; 166:4813-4817; ; doi: 10.4049/jimmunol.166.8.4813 http://www.jimmunol.org/content/166/8/4813 Subscription Permissions Email Alerts This article cites 32 articles, 16 of which you can access for free at: http://www.jimmunol.org/content/166/8/4813.full#ref-list-1 Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2001 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. Downloaded from http://www.jimmunol.org/ by guest on June 17, 2017 References ● Cutting Edge: Eotaxin Elicits Rapid Vesicular Transport-Mediated Release of Preformed IL-4 from Human Eosinophils1 Christianne Bandeira-Melo, Kumiya Sugiyama, Lesley J. Woods, and Peter F. Weller2 E osinophils, prominent in Th2-driven immune responses, including asthma and allergic and parasitic diseases, may have multiple roles in these diseases. As effector cells eosinophils, based in part on their release of cationic granule proteins and lipid mediators, may contribute to the immunopathogenesis of allergic diseases (1). Additional functional roles for eosinophils are indicated by findings that eosinophils may exert immunomodulatory activities via interactions with T and B lymphocytes. For these potential interactions with lymphocytes, human eosinophils express costimulatory surface proteins, including CD40, CD28, CD86, and MHC class II Ags (2– 4), and produce over two dozen cytokines, including the prototypical Th2 cytokine IL-4 (5). Like eosinophils, IL-4 is a hallmark of allergic and parasitic disorders. IL-4 contributes to the polarization toward Th2 differentiation and promotes IgE class switching (6, 7). The actions of IL-4 are not limited to the initiation of Th2 responses, but also may stimulate other cellular responses that contribute to manifestations of allergic diseases (6). Potential cellular sources of IL-4 include CD4⫹ T cells (7), mast cells (8), basophils (9), NK1.1⫹ T cells (10), ␥␦ T cells (11), and eosinophils (12–14). Within eosinophils, unlike CD4⫹ T cells, IL-4 is stored as a preformed pool within eosinophil-specific granules (13–17). A role for eosinophil-derived IL-4 has been demonstrated in a murine system in which the i.p. instillation of Schistosoma mansoni eggs led to the enhanced generation over 12 h of IL-4 derived from peritoneal exudate eosinophils (12). Because nothing is known about how IL-4 within human eosinophils might be mobilized, we have investigated stimuli and mechanisms that lead to the extracellular release of IL-4 stored within normal donor-derived eosinophils. Materials and Methods Purification of human eosinophils Eosinophils were isolated from the blood of 18 healthy nonatopic donors by negative selection using the MACS anti-CD16 immunomagnetic bead procedure (Miltenyi Biotec, Auburn, CA) (18). Eosinophil purity and viability was ⬎99 and ⬎95%, respectively. Detection of intracellular IL-4 Department of Medicine, Harvard Thorndike Laboratories, Charles A. Dana Research Institute, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, Massachusetts 02215 Received for publication December 14, 2000. Accepted for publication February 26, 2001. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 This work was supported by National Institutes of Health Grants A20241, AI22571, AI41995, and HL56386. 2 Address correspondence and reprint requests to Dr. Peter F. Weller, Beth Israel Deaconess Medical Center, DA-617, 330 Brookline Avenue, Boston, MA 02215. E-mail address: [email protected] Copyright © 2001 by The American Association of Immunologists ● Intracellular IL-4 was analyzed by 1) immunofluorescence microscopy of cytospin preparations of paraformaldehyde-fixed/saponin-permeabilized eosinophils stained with Alexa546-labeled (Molecular Probes, Eugene, OR) anti-IL-4 mAb (clone 3010.211) or isotype control mouse IgG1 (both obtained from R&D Systems, Minneapolis, MN) and viewed with a TE300 Nikon fluorescence microscope; 2) flow cytometry (FACScan, CellQuest software; BD Biosciences, Mountain View, CA) of paraformaldehydefixed/saponin-permeabilized eosinophils stained with PE-conjugated antiIL-4 mAb (clone MP4-25D2) or isotype control rat IgG1 (both obtained from BD PharMingen, San Diego, CA); and 3) ELISA (R&D kit) of postnuclear supernatants (14,000 ⫻ g for 20 min) of eosinophil lysates (in 1 mM DTT, 1 mM EDTA, 0.1% SDS and 1% Nonidet P-40, pH 7.5, 150 mM NaCl, and 20 mM HEPES with protease inhibitors). 0022-1767/01/$02.00 Downloaded from http://www.jimmunol.org/ by guest on June 17, 2017 IL-4 release is important in promoting Th2-mediated allergic and parasitic immune responses. Although human eosinophils are potential sources of IL-4, physiologic mechanisms to elicit its release have not been established. By flow cytometry and microscopy, eosinophils from normal donors uniformly contained preformed IL-4. In contrast to cytolytic IL-4 release from calcium ionophore-activated eosinophils, eotaxin and RANTES, but not IFN-␥, elicited IL-4 release by noncytotoxic mechanisms. With a dual Ab capture and detection immunofluorescent microscopic assay, IL-4 was released at discrete cell surface sites. IL-5 enhanced eotaxin-induced IL-4 release, which was mediated by G protein-coupled CCR3 receptors, detectable as early as 5 min and maximum within 1 h. IL-4 release was not diminished by transcription or protein synthesis inhibitors, but was suppressed by brefeldin A, an inhibitor of vesicle formation. Thus, CCR3-mediated signaling can rapidly mobilize IL-4 stored preformed in human eosinophils for release by vesicular transport to contribute to immune responses. The Journal of Immunology, 2001, 166: 4813– 4817. 4814 CUTTING EDGE ELISA for detection of eosinophil-released IL-4 Eosinophils (2 ⫻ 10 cells in 1 ml) were incubated for 1 h (37°C) with chemokines (R&D Systems) or A23187 (Sigma, St. Louis, MO) in RPMI 1640 medium containing 0.1% OVA. Eosinophil supernatant IL-4 was measured by ELISA (sensitivity 10 pg/ml; R&D Systems). 6 EliCell assay for the detection of eosinophil-released IL-4 Results and Discussion IL-4, a major mediator in the induction and regulation of allergic and parasitic immune responses, is not produced exclusively by activated Th2 lymphocytes. Earlier studies established that human eosinophils synthesize and store IL-4 within their specific granules and suggested, based on immunocytochemistry, that there might be subpopulations of IL-4-positive and -negative eosinophils (13, 14). Our analyses of populations of eosinophils from normal donors extend these findings by demonstrating that all circulating eosinophils contained preformed IL-4, as evidenced by a uniformly positive, unimodal pattern of intracellular IL-4 immunoreactivity by flow cytometry (Fig. 1A, top) and by high level, granule-associated eosinophil anti-IL-4 immunostaining (94.3 ⫾ 2.5% positive cells; mean ⫾ SD, n ⫽ 4) by fluorescence microscopy (images not shown). By ELISA of cell lysates, eosinophils contained more IL-4 (32.3 ⫾ 6.2 pg/2 ⫻ 106 cells) than did PBMCs (2.3 ⫾ 0.9 pg/2 ⫻ 106 cells, means ⫾ SEM, n ⫽ 7) from the same donors (Fig. 1A, bottom). Thus, in addition to the presence of IL-4 in tissue eosinophils in bronchial biopsies of asthmatic patients and FIGURE 1. IL-5 enhances eotaxin-induced release of preformed IL-4 from eosinophils. A, Eosinophils from nonatopic donors contain preformed IL-4. Top, Flow cytometry histogram of intracellular IL-4 in saponin-permeabilized eosinophils, with Alexa-546-labeled anti-IL-4 mAb (dashed line) and nonimmune IgG1 (solid line). No anti-IL-4 staining was detected with nonpermeabilized eosinophils (data not shown). Result is representative of findings from four donors. Bottom, ELISA-assayed quantities of preformed IL-4 in lysates of eosinophils and PBMCs from seven normal donors. Bars denote the means. B, EliCell assays of IL-4 released extracellularly from eosinophils (captured with a biotinylated anti-IL-4 Ab and detected with Alexa546-labeled anti-IL-4 mAb). Dose-responses of eotaxin-induced IL-4 release at 1 h with and without concomitant 2 nM IL-5 were expressed both as the average fluorescence intensities for immunoreactive IL-4 around 50 individual eosinophils (top) and the percentages of eosinophils exhibiting extracellularly released IL-4 (bottom). Results are means ⫾ SD from five donors. ⴱ and ⴱⴱ, p ⬍ 0.05 and p ⬍ 0.01, respectively, compared with unstimulated eosinophils. C, Phase-contrast (left) and fluorescent (right) microscopic images of identical fields of eosinophils. Anti-IL-4 immunoreactive sites (red) are overlaid on phase-contrast images to facilitate their localization. Images show representative eosinophils stimulated for 1 h with 2 nM IL-5, 6 nM eotaxin, or both. Bottom image shows a representative brefeldin A (BFA; 1 g/ml)-treated IL-5/eotaxin-stimulated eosinophil. Numerical values (right) are the fluorescent intensities (in arbitrary units) of immunoreactive IL-4 released by each of the shown eosinophils. Downloaded from http://www.jimmunol.org/ by guest on June 17, 2017 The EliCell assay, a gel-phase dual Ab capture and detection assay based on microscopic observations of individual viable cells, was performed as detailed (18) to enumerate the proportion of eosinophils releasing IL-4 and to electronically quantitate the average relative amounts of IL-4 released extracellularly. A biotinylated goat polyclonal anti-IL-4 Ab (20 g/ml; R&D Systems) was used as capturing Ab and an Alexa546-labeled antiIL-4 mAb (R&D Systems) was used (400 l of 10 g/ml) to detect released IL-4. Alexa546-labeled mouse IgG1 was included as a nonimmune isotype control. An irrelevant biotinylated capture Ab was substituted in combination with the Alexa546-labeled anti-IL-4 detection Ab to ascertain that 1) cell permeabilization had not allowed detection of intracellular IL-4; and 2) retention of surface-released IL-4 was dependent on the immobilized anti-IL-4 capturing Ab. In some experiments, eosinophils were pretreated (37°C) for 30 min with specific inhibitors: 1) a neutralizing anti-CCR3 mAb (clone 61828.111) or isotype-matched control rat IgG (both 10 g/ml; R&D Systems); 2) pertussis toxin (10 or 100 ng/ml; Calbiochem, La Jolla, CA); 3) actinomycin D and cycloheximide (both at 1 and 10 M; Calbiochem); or 4) brefeldin A (0.1 and 1 g/ml; Biomol, Plymouth Meeting, PA). Statis- tical comparisons were made by ANOVA followed by Student NewmanKeuls t test with differences considered significant when p ⬍ 0.05. The Journal of Immunology 4815 (data not shown). The latter condition assured that neither intracellular nor membrane-bound IL-4 was being detected in the nonpermeabilized eosinophils, and the punctate pattern of immunoreactive IL-4 released at discrete loci proximate to the cell surface (Fig. 1C) (as confirmed by confocal microscopy; data not shown) was compatible with a vesicular transport-mediated process of IL-4 release. IL-8, which does not stimulate normal eosinophils (27), did not elicit IL-4 release (Table I). Of greater interest, IFN-␥, at a concentration that effectively elicited vesicular transport-mediated release of RANTES from eosinophils (18, 25), failed to elicit IL-4 release (Table I), suggesting that differential signaling may function to selectively mobilize at least the cytokine proteins stored preformed in eosinophil granules. IL-5 by itself did not elicit detectable IL-4 release from eosinophils (Fig. 1B), but IL-5 did enhance eotaxin-stimulated mobilization of IL-4. The simultaneous addition of IL-5 with even low concentrations of eotaxin (0.06 and 0.6 nM) evoked release of IL-4 from significant proportions of eosinophils, whereas these eotaxin concentrations alone did not stimulate detectable IL-4 release (Fig. 1B). IL-5 in concert with higher concentrations of eotaxin (6 and 60 nM) doubled the number of eosinophils releasing IL-4 and the amounts of IL-4 released (Fig. 1B). IL-5 also accelerated eotaxininduced IL-4 release from eosinophils. Even as early as 5 min, IL-5 in concert with eotaxin enhanced both the percentages of eosinophils releasing IL-4 and the quantities of IL-4 being released (Fig. 2A). We next evaluated mechanisms underlying the IL-5/eotaxinelicited release of IL-4 from eosinophils. The release of IL-4 from IL-5/eotaxin-stimulated eosinophils was mediated through the G protein-linked CCR3 chemokine receptor. Pretreatment of eosinophils with a neutralizing anti-CCR3 receptor mAb reduced the percentages of eosinophils releasing IL-4 (71 ⫾ 5% inhibition vs 0% with a control Ab) and the amounts of IL-4 released (98 ⫾ 2% inhibition vs 11% with a control Ab) (both n ⫽ 3; p ⬍ 0.05). Likewise, pertussis toxin pretreatment at 10 and 100 ng/ml inhibited eotaxin-elicited IL-4 release and reduced the percentages of eosinophils releasing IL-4 by 56 ⫾ 10 and 89 ⫾ 3%, respectively (both p ⬍ 0.05, n ⫽ 3). Neither actinomycin D, an inhibitor of transcription, nor cycloheximide, a protein synthesis inhibitor, suppressed eosinophil IL-4 release induced by IL-5/eotaxin (Fig. 2B). Therefore, the mechanisms underlying IL-5/eotaxin-elicited IL-4 release are not likely to require either the new synthesis of IL-4 or of other proteins that contribute to the secretory process. In contrast, brefeldin A, an inhibitor of vesicle formation, substantially suppressed the quantities of IL-4 released extracellularly around eosinophils without diminishing the overall number of cells releasing IL-4 (Figs. 1C and 2B). Table I. ELISA and EliCell assays of IL-4 released from human eosinophilsa ELISA EliCell Stimulus Dose n IL-4 (pg/ml) % Eosinophils releasing IL-4 IL-4 fluorescence intensity ⫻ 106/cell Medium IFN-␥ Eotaxin RANTES IL-8 A23187 500 U/ml 6.0 nM 6.4 nM 6.2 nM 0.5 M 4 4 4 3 3 4 1.6 ⫾ 0.3 ND 0.9 ⫾ 0.4 1.7 ⫾ 0.1 1.1 ⫾ 0.7 45.4 ⫾ 12.5** 1⫾1 1⫾1 44 ⫾ 11* 59 ⫾ 22* 0⫾0 89 ⫾ 5** 0.0 ⫾ 0.0 0.0 ⫾ 0.0 1.0 ⫾ 0.2* 0.5 ⫾ 0.3* 0.0 ⫾ 0.0 5.2 ⫾ 0.7** a Eosinophils were incubated with stimuli for 1 h. ELISA assessed IL-4 in 1-ml supernatants from 2 ⫻ 106 eosinophils. EliCell assessed both the percentages of eosinophils exhibiting extracellular IL-4 and the average electronically measured immunofluorescent intensities of extracellular IL-4. Results are means ⫾ SD. *, p ⬍ 0.01 and **, p ⬍ 0.001 compared with medium. n, Number of different donors. Downloaded from http://www.jimmunol.org/ by guest on June 17, 2017 in allergen-induced cutaneous late-phase reactions in atopic subjects (15, 19), human blood-derived eosinophils are a major potential source of IL-4. Because eosinophils contain preformed IL-4, the mechanisms to mobilize IL-4 from specific granule storage sites for its extracellular release need to be defined. Prior studies established the releasability of IL-4 from eosinophils activated with either serumcoated beads, IgA immune complexes, or calcium ionophore A23187 (13, 14, 20), but these nonphysiological stimuli are not selective and may be uniformly exocytotic and/or cytolytic (21, 22). Indeed with our methods, A23187 activation of eosinophils led to the cytolytic release of IL-4. Not only were the amounts of immunoreactive IL-4 released in eosinophil supernatants following A23187 stimulation (Table I) equivalent to the total preformed IL-4 content of eosinophils (Fig. 1A, bottom), but microscopy of eosinophils in EliCell assays demonstrated extensive IL-4 immunostaining of eosinophils (Table I) that exhibited morphological signs of cell damage (data not shown, but as previously illustrated for RANTES staining in A23187 stimulated eosinophils; Ref. 18). An alternative mechanism to either exocytosis or cytolysis for the release of eosinophil granule-derived proteins has been indicated by ultrastructural observations of tissue and blood eosinophils activated in vivo. At least for the major cationic protein components of eosinophil-specific granules, selective losses of the core or matrix components of the granules and other findings suggest that eosinophil-specific granule contents may be mobilized by selective incorporation into small vesicles that traffic to the cell surface and release these granule contents by a process of “piecemeal” degranulation based on vesicular transport (23, 24). Recently, Lacy and coworkers identified IFN-␥ as a physiological stimulus that in vitro induces piecemeal release of RANTES, a chemokine also stored preformed in eosinophil-specific granules (25); we have established a microscopic assay, the EliCell assay, to study the piecemeal degranulation process in eosinophils (18). We evaluated the capacity of several eosinophil agonists to elicit IL-4 release from eosinophils. The C-C chemokines, eotaxin and RANTES, did not elicit IL-4 release at levels detectable by ELISA of supernatant fluids (Table I), consistent with a recent report that these cytokines elicited ELISA-detectable “degranulation” only if eosinophils were pretreated with cytochalasin B (26). In contrast with the EliCell assay, both eotaxin and RANTES stimulated release of IL-4 detectable extracellularly (Table I). No IL-4 staining was found with unstimulated eosinophils or when the Alexa546labeled anti-IL-4 detection Ab was replaced by an Alexa546-labeled isotype IgG1 nonimmune control. Moreover, no IL-4 was detectable when the biotinylated anti-IL-4 capture Ab (which was necessary to immobilize IL-4 at its extracellular sites of release; Ref. 18) was substituted with a biotinylated irrelevant control Ab 4816 CUTTING EDGE References Our findings indicate that chemokines acting via CCR3-initiated signaling pathways can very rapidly mobilize preformed stores of IL-4 from within human eosinophils. The means of extracellular release was by noncytotoxic, vesicular transport as indicated by the microscopic patterns of focal cell surface IL-4 release, the absence of required new protein synthesis, and the inhibition by a vesicle formation inhibitor, brefeldin A. In support of this mechanism, we have localized the vesicle-associated membrane protein-2 by immunogold electron microscopy not only at vesicles within eosinophils but also at the outer membrane of eosinophil-specific granules (28), indicating the capacity of secretory vesicles to traffic from eosinophil granule membranes. Although released IL-4 concentrations in supernatant fluids were not sufficient to be detectable by conventional ELISA, local concentrations of released IL-4 may effectively stimulate responses in tissue sites of eosinophil localization. IL-4 released by eosinophils may augment effector actions of eosinophils, including enhancing the IL-4-dependent generation of airway mucous secretion (29) or even the elicitation of further eotaxin generation (30). Moreover, because airway eosinophils can traffic back to regional lymph nodes and effectively present airway-derived Ags to elicit proliferation of CD4⫹ T cells (31), local IL-4 release by eosinophils within lymph nodes, the thymus (32), or other sites may also modulate the local responses of lymphocytes. Thus, in contrast to CD4⫹ T cells in which IL-4 synthesis needs to be transcriptionally induced, human eosinophils have the capacity by means of vesicular transport to physiologically and rapidly release their preformed stores of IL-4. Downloaded from http://www.jimmunol.org/ by guest on June 17, 2017 FIGURE 2. IL-5 accelerates the eotaxin-induced release of IL-4 from eosinophils, which is mediated by vesicular transport. With EliCell assays, IL-4 released extracellularly from eosinophils was captured with anti-IL-4 Ab bound to a gel matrix and detected with Alexa546-labeled anti-IL-4 mAb. IL-4 release was measured both as the average fluorescence intensities for immunoreactive IL-4 around 50 individual eosinophils (top) and the percentages of eosinophils exhibiting extracellularly released IL-4 (bottom). A, A representative (n ⫽ 4) time course of IL-4 release from eosinophils stimulated with medium (E), 6 nM eotaxin (䡺), 2 nM IL-5 (F), or both eotaxin and IL-5 (f). B, Eosinophils were pretreated for 30 min with actinomycin D (ActD), cycloheximide (Cycl), or brefeldin A (BFA) and then stimulated with 6 nM eotaxin and 2 nM IL-5 for 1 h. Results are means ⫾ SD from three donors. ⴱ and ⴱⴱ, p ⬍ 0.05 and p ⬍ 0.01, respectively, compared with no inhibitors. 1. Gleich, G. J. 2000. Mechanisms of eosinophil-associated inflammation. J. Allergy Clin. Immunol. 105:651. 2. Ohkawara, Y., K. G. Lim, Z. Xing, M. Glibetic, K. Nakano, J. Dolovich, K. Croituru, P. F. Weller, and M. Jordana. 1996. CD40 expression by human peripheral blood eosinophils. J. Clin. Invest. 97:1761. 3. Woerly, G., N. Roger, S. Loiseau, D. Dombrowicz, A. Capron, and M. Capron. 1999. Expression of CD28 and CD86 by human eosinophils and role in the secretion of type 1 cytokines (interleukin 2 and interferon ␥): inhibition by immunoglobulin A complexes. J. Exp. Med. 190:487. 4. Weller, P. F., T. H. Rand, T. Barrett, A. Elovic, D. T. Wong, and R. W. Finberg. 1993. Accessory cell function of human eosinophils: HLA-DR dependent, MHCrestricted antigen-presentation and interleukin-1␣ formation. J. Immunol. 150: 2554. 5. Lacy, P., and R. Moqbel. 2000. Eosinophil cytokines. Chem. Immunol. 76:134. 6. Brown, M. A., and J. Hural. 1997. Functions of IL-4 and control of its expression. Crit. Rev. Immunol. 17:1. 7. Noben-Trauth, N., J. Hu-Li, and W. E. Paul. 2000. Conventional, naive CD4⫹ T cells provide an initial source of IL-4 during Th2 differentiation. J. Immunol. 165:362. 8. Plaut, M., J. H. Pierce, C. J. Watson, J. Hanley-Hyde, R. P. Nordan, and W. E. Paul. 1989. Mast cell lines produce lymphokines in response to crosslinkage of Fc⑀RI or to calcium ionophore. Nature 339:64. 9. Schroeder, J. T., D. W. MacGlashan, Jr., A. Kagey-Sobotka, J. M. White, and L. M. Lichtenstein. 1994. IgE-dependent IL-4 secretion by human basophils: the relationship between cytokine production and histamine release in mixed leukocyte cultures. J. Immunol. 153:1808. 10. Yoshimoto, T., and W. E. Paul. 1994. CD4pos, NK1.1pos T cells promptly produce interleukin 4 in response to in vivo challenge with anti-CD3. J. Exp. Med. 179:1285. 11. Ferrick, D. A., M. D. Schrenzel, T. Mulvania, B. Hsieh, W. G. Ferlin, and H. Lepper. 1995. Differential production of interferon-␥ and interleukin-4 in response to Th1- and Th2-stimulating pathogens by ␥␦ T cells in vivo. Nature 373:255. 12. Sabin, E. A., M. A. Kopf, and E. J. Pearce. 1996. Schistosoma mansoni egginduced early IL-4 production is dependent upon IL-5 and eosinophils. J. Exp. Med. 184:1871. 13. Moqbel, R., S. Ying, J. Barkans, T. M. Newman, P. Kimmitt, M. Wakelin, L. Taborda-Barata, Q. Meng, C. J. Corrigan, S. R. Durham, and A. B. Kay. 1995. Identification of messenger RNA for IL-4 in human eosinophils with granule localization and release of the translated product. J. Immunol. 155:4939. 14. Bjerke, T., M. Gaustadnes, S. Nielsen, L. P. Nielsen, P. O. Schiotz, N. Rudiger, C. M. Reimert, R. Dahl, I. Christensen, and L. K. Poulsen. 1996. Human blood eosinophils produce and secrete interleukin 4. Respir. Med. 90:271. 15. Ying, S., M. Humbert, J. Barkans, C. J. Corrigan, R. Pfister, G. Menz, M. Larche, D. S. Robinson, S. R. Durham, and A. B. Kay. 1997. Expression of IL-4 and IL-5 mRNA and protein product by CD4⫹ and CD8⫹ T cells, eosinophils, and mast cells in bronchial biopsies obtained from atopic and nonatopic (intrinsic) asthmatics. J. Immunol. 158:3539. 16. Möller, G. M., T. A. de Jong, T. H. van der Kwast, S. E. Overbeek, A. F. Wierenga-Wolf, T. Thepen, and H. C. Hoogsteden. 1996. Immunolocalization of interleukin-4 in eosinophils in the bronchial mucosa of atopic asthmatics. Am. J. Respir. Cell Mol. Biol. 14:439. 17. Nakajima, H., G. J. Gleich, and H. Kita. 1996. Constitutive production of IL-4 and IL-10 and stimulated production of IL-8 by normal peripheral blood eosinophils. J. Immunol. 156:4859. 18. Bandeira-Melo, C., G. Gillard, I. Ghiran, and P. F. Weller. 2000. EliCell: a solid-phase dual antibody capture and detection assay to detect cytokine release by eosinophils. J. Immunol. Methods 244:105. 19. Barata, L. T., S. Ying, Q. Meng, J. Barkans, K. Rajakulasingam, S. R. Durham, and A. B. Kay. 1998. IL-4- and IL-5-positive T lymphocytes, eosinophils, and mast cells in allergen-induced late-phase cutaneous reactions in atopic subjects. J Allergy Clin. Immunol. 101:222. 20. Nonaka, M., R. Nonaka, K. Woolley, E. Adelroth, K. Miura, Y. Okhawara, M. Glibetic, K. Nakano, P. O’Byrne, J. Dolovich, and M. Jordana. 1995. Distinct immunohistochemical localization of IL-4 in human inflamed airway tissues: IL-4 is localized to eosinophils in vivo and is released by peripheral blood eosinophils. J. Immunol. 155:3234. 21. Fukuda, T., S. J. Ackerman, C. E. Reed, M. S. Peters, S. L. Dunnette, and G. J. Gleich. 1985. Calcium ionophore A23187 calcium-dependent cytolytic degranulation in human eosinophils. J. Immunol. 135:1349. 22. Weiler, C. R., H. Kita, M. Hukee, and G. J. Gleich. 1996. Eosinophil viability during immunoglobulin-induced degranulation. J. Leukocyte Biol. 60:493. 23. Karawajczyk, M., L. Seveus, R. Garcia, E. Bjornsson, C. G. Peterson, G. M. Roomans, and P. Venge. 2000. Piecemeal degranulation of peripheral blood eosinophils: a study of allergic subjects during and out of the pollen season. Am. J. Respir. Cell Mol. Biol. 23:521. 24. Dvorak, A. M., and P. F. Weller. 2000. Ultrastructural analysis of human eosinophils. Chem. Immunol. 76:1. 25. Lacy, P., S. Mahmudi-Azer, B. Bablitz, S. C. Hagen, J. R. Velazquez, S. F. P. Man, and R. Moqbel. 1999. Rapid mobilization of intracellularly stored RANTES in response to interferon-␥ in human eosinophils. Blood 94:23. 26. Fujisawa, T., Y. Kato, H. Nagase, J. Atsuta, A. Terada, K. Iguchi, H. Kamiya, Y. Morita, M. Kitaura, H. Kawasaki, et al. 2000. Chemokines induce eosinophil degranulation through CCR-3. J. Allergy Clin. Immunol. 106:507. The Journal of Immunology 27. Sabroe, I., A. Hartnell, L. A. Jopling, S. Bel, P. D. Ponath, J. E. Pease, P. D. Collins, and T. J. Williams. 1999. Differential regulation of eosinophil chemokine signaling via CCR3 and non-CCR3 pathways. J. Immunol. 162:2946. 28. Feng, D., R. Flaumenhaft, C. Bandeira-Melo, P. F. Weller, H. F. Dvorak, and A. M. Dvorak. 2001. Ultrastructural localization of vesicle-associated membrane protein-2 (VAMP-2) in human pericytes, vascular smooth muscle cells, endothelial cells, neutrophils and eosinophils. J. Histochem. Cytochem. 49:293. 29. Cohn, L., R. J. Homer, H. MacLeod, M. Mohrs, F. Brombacher, and K. Bottomly. 1999. Th2-induced airway mucus production is dependent on IL-4R␣, but not on eosinophils. J. Immunol. 162:6178. 4817 30. Mochizuki, M., J. Bartels, A. I. Mallet, E. Christophers, and J. M. Schroder. 1998. IL-4 induces eotaxin: a possible mechanism of selective eosinophil recruitment in helminth infection and atopy. J. Immunol. 160:60. 31. Shi, H., A. Humbles, C. Gerard, Z. Jin, and P. F. Weller. 2000. Lymph node trafficking and antigen presentation by endobronchial eosinophils. J. Clin. Invest. 105:945. 32. Throsby, M., A. Herbelin, J. M. Pléau, and M. Dardenne. 2000. CD11c⫹ eosinophils in the murine thymus: developmental regulation and recruitment upon MHC class I-restricted thymocyte deletion. J. Immunol. 165:1965. Downloaded from http://www.jimmunol.org/ by guest on June 17, 2017