Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Extracellular matrix wikipedia , lookup
Cellular differentiation wikipedia , lookup
Endomembrane system wikipedia , lookup
Cell culture wikipedia , lookup
Protein moonlighting wikipedia , lookup
Cell encapsulation wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
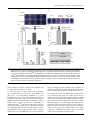
Journal of General Virology (2015), 96, 1795–1800 Short Communication DOI 10.1099/vir.0.000116 VP5 autocleavage is required for efficient infection by in vitro-recoated aquareovirus particles Shicui Yan,1,2 Jie Zhang,1 Hong Guo,1 Liming Yan,1,2 Qingxiu Chen,1,2 Fuxian Zhang1 and Qin Fang1 Correspondence Qin Fang [email protected] Received 23 January 2015 Accepted 4 March 2015 1 State Key Laboratory of Virology, Wuhan Institute of Virology, Chinese Academy of Sciences, Wuhan 430071, PR China 2 University of Chinese Academy of Sciences, Beijing, PR China Grass carp reovirus (GCRV) is a member of the genus Aquareovirus in the family Reoviridae, and contains five core proteins (VP1–VP4 and VP6) and two outer-capsid proteins (VP5 and VP7) in its particle. Previous studies have revealed that the outer-capsid proteins of reovirus are responsible for initiating infection, but the mechanism is poorly understood. Using baculovirusexpressed VP5 and VP7 to recoat purified cores, in vitro assembly of GCRV was achieved in this study. Recoated GCRV (R-GCRV) closely resembled native GCRV (N-GCRV) in particle morphology, protein composition and infectivity. Similar to N-GCRV, the infectivity of R-GCRV could be inhibited by treating cells with the weak base NH4Cl. In addition, recoated particles carrying an AsnAAla substitution at residue 42 of VP5 (VP5N42A/VP7 R-GCRV) were no longer infectious. These results provide strong evidence that autocleavage of VP5 is critical for aquareovirus to initiate efficient infection. The genus Aquareovirus is a tentative member of the family Reoviridae, which currently includes 15 proposed genera (King et al., 2011). Many aquareoviruses have been isolated from apparently healthy-looking fish, including the finfish and shellfish species. However, several members within the genus are identified as pathogens that are capable of causing serious disease in aquatic animals (Fang et al., 1989; Ke et al., 1990; Rangel et al., 1999). Grass carp reovirus (GCRV) has been recognized as the most pathogenic among the isolated aquareoviruses (Mohd Jaafar et al., 2008; Rangel et al., 1999). Hence, GCRV provides a useful model to understand the infection and pathogenesis of aquareoviruses in general. the inner capsid or core of the virion. Previous threedimensional structure studies have revealed that the inner capsid of GCRV is constructed mainly of two proteins, VP3 and VP6, whilst the other three proteins (VP1, VP2 and VP4) are RNA polymerase-related complex proteins that possess the enzymic activities necessary for viral transcription and replication (Cheng et al., 2008; Fang et al., 2005). Similar to the homologous proteins m1 and s3 of MRV in their structural organization, VP7 is a protective protein located on top of the VP5 subunits. In contrast to MRVs, there is no protein equivalent of s1, which sits on the fivefold vertices of the virion and functions as the cellattachment protein (Cheng et al., 2008; Fang et al., 2005). The struture of GCRV is a multilayered icosahedral particle with a diameter of approximately 75–85 nm enclosing a segmented dsRNA genome. The 11 genome segments encode seven structural proteins (VP1–VP7) and five nonstructural proteins. Genome sequence analysis and threedimensional structural reconstructions by cryo-electron microscopy have indicated that there are great similarities between aquareoviruses and mammalian reoviruses (MRVs) in protein structure and particle organization (Cheng et al., 2008, 2010; Fang et al., 2005). Similar to other reoviruses in the family Reoviridae, the GCRV particle is organized mainly into two layers: an outer capsid and an inner capsid. Two hundred trimeric heterodimers of the VP5 and VP7 proteins make up the outer capsid of the virus particle, whilst the other five structural proteins (VP1–VP4 and VP6) constitute Reoviruses are non-enveloped virions that lack a lipidbilayer membrane. Based on their particle organization, they utilize an entry pathway different from that of enveloped viruses. To initiate infection in a target cell, reoviruses must disrupt the host cell membrane through a mechanism other than the membrane fusion route seen for enveloped viruses. Previous studies have revealed that MRVs need to be activated for infection by undergoing stepwise disassembly of the outer capsid to generate infectious subvirion particles (ISVP) (Chandran & Nibert, 1998; Liemann et al., 2002; Sturzenbecker et al., 1987). When virions are activated to infect host cells, the outermost capsid protein s3 of MRV is degraded by endosomal proteases, leaving m1 exposed on the surface. The m1 protein is found predominantly in the form of two fragments, m1N 000116 G 2015 The Authors Printed in Great Britain Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 17:34:50 1795 S. Yan and others (4 kDa) and m1C (72 kDa), following putative autocleavage between residues Asn42 and Pro43 near the N terminus of the intact protein (Nibert et al., 1991, 2005). In addition, infection with intact virions, but not ISVPs, can be blocked by treating cells with a weak base, such as NH4Cl, that inhibits the proteolysis of m1 within cells (Chandran & Nibert, 1998; Jané-Valbuena et al., 1999). Studies on the characteristics of infectious GCRV particles have demonstrated that complete digestion of VP7 and partial cleavage of VP5 can lead to enhanced infectivity of the virus, indicating that VP7 and VP5 may play important roles in GCRV entry into host cells during infection (Fang et al., 2008; Zhang et al., 2010). Recent three-dimensional structural studies of GCRV ISVP at 3.3 Å resolution revealed that autocleavage of the VP5 protein, which is myristoylated at the N terminus, occurs during conversion from the dormant to the primed state at an early stage of viral infection (Zhang et al., 2010). To gain insight into the molecular mechanism of virus infection and particle assembly, in vitro assembly of GCRV-like particles by recoating baculovirus-expressed VP5 and VP7 onto purified cores was achieved in this study. In addition, the infection characteristics of recoated WT (R-GCRV or VP5/ VP7 R-GCRV) and mutant (VP5N42A/VP7 R-GCRV) particles were investigated. Previous studies reported that the outer-capsid proteins VP5 and VP7 are required for cell entry during aquareovirus infection, and that putative autocleavage of outercapsid protein VP5 is likely to be critical for virus entry into host cells (Fang et al., 2008; Zhang et al., 2010). To determine whether VP5 and/or VP7 were responsible for aquareovirus entry, a dual-expressed VP5 and VP7 (vAcGCRV-VP5/VP7) and singly expressed VP5 or VP7 (vAcGCRV-VP5 or vAcGCRV-VP7; data not shown) recombinant baculoviruses were constructed, respectively. In addition, to determine whether the N42 residue was responsible for autocleavage of VP5, a mutated VP5 containing an AsnAAla substitution at residue 42 was also generated (Fig. 1a) and expressed in Spodoptera frugiperda (Sf9) cells using a recombinant baculovirus (vAcGCRVVP5N42A/VP7). An immunofluorescence assay was carried out to confirm the subcellular location of VP5 or VP5N42A with VP7 during recombinant baculovirus infection. As shown in Fig. 1(b), compared with vAcGCRV-VP5/VP7, the co-localization of mutant VP5N42A with VP7 in infected Sf9 cells was not changed. To estimate the levels of protein expression and VP5N/VP5C cleavage, Sf9 cell lysates containing VP5 or VP5N42A co-expressed with VP7 were analysed by Western blotting (WB). The data revealed that the expression of WT VP5 resulted in two VP5-derived bands, namely intact VP5 and cleaved C-terminal VP5 (termed VP5C). By contrast, in vAcGCRV-VP5N42A/VP7infected cells, no cleaved VP5C could be detected (Fig. 1c). Moreover, the interactions between VP5 or VP5N42A and VP7 were investigated by a co-immunoprecipitation assay. The results showed that WT VP5 or mutant VP5N42A could efficiently co-immunoprecipitate with VP7 using anti-VP7 1796 polyclonal antibodies, and VP7 was efficiently co-immunoprecipitated with VP5 or VP5N42A by anti-VP5 polyclonal antibodies (Fig. 1d) but not by control antibody. These results indicated that the vAcGCRV-VP5/VP7 and vAcGCRVVP5N42A/VP7 recombinant baculoviruses could express VP5 or VP5N42A and VP7 at high levels in Sf9 cells. Moreover, both VP5 and VP5N42A could interact with VP7. These results suggested that the VP5/VP7 and VP5N42A/VP7 recombinant proteins co-expressed in Sf9 cells were suitable for recoating GCRV cores in vitro. To achieve the assembly in vitro, GCRV cores with protruding turrets were purified by CsCl gradient centrifugation and their structure confirmed using transmission electron microscopy (TEM) (Fig. 2a). Cytoplasmic extracts were also prepared from Sf9 cells infected with recombinant baculoviruses co-expressing proteins VP5 and VP7 or singly expressing VP5 or VP7. These extracts were incubated with purified GCRV cores at 28 uC for 5 h and the recoated particles purified by CsCl gradient centrifugation. Comparison by TEM of these recoated particles (RGCRV) with native particles (N-GCRV), also purified by CsCl-based centrifugation, showed that the two types of particle had indistinguishable morphologies (Fig. 2b, c). No recoated particles were obtained when purified cores were incubated with VP5 or VP7 alone, strongly suggesting that a complex of VP5 and VP7 is required for binding to and recoating viral cores. To confirm the protein composition of R-GCRV particles, samples were analysed using SDS-PAGE (Fig. 2d). Seven structural protein components were identified whose molecular masses were consistent with those seen in similar analysis performed on N-GCRV particles. In addition, WB analysis confirmed that VP5 and VP7 had specifically bound to the cores and remained bound throughout the CsCl centrifugation (Fig. 2e). To evaluate the infectivity of recoated particles, they were titred on Ctenopharyngodon idellus kidney (CIK) cells. As shown in Fig. 2(f), in common with N-GCRVs, RGCRVs were 105-fold more infectious than cores on a perparticle basis. In contrast to these results, when VP5 or VP7 cell extracts (infected by vAcGCRV-VP5 or vAcGCRVVP7) were singly incubated with cores (VP5 R-GCRV or VP7 R-GCRV), no infectivity was detected. These results indicated that infectious R-GCRV could only be produced by recoating core particles with both VP5 and VP7 proteins. Cell entry of MRV or aquareovirus has been reported to be a multistep process involving proteolytic disassembly of virions into ISVPs (Chandran et al., 1999; Fang et al., 2008). This proteolysis of intact virions could be inhibited by treating cells with weak bases, such as NH4Cl (Chandran et al., 1999). To prove the biological similarity between RGCRVs and N-GCRVs, cells were infected with or without 20 mM NH4Cl treatment. At 24 h post-infection (p.i.), obvious viral plaques were observed in the mock-treated cells but not in NH4Cl-treated cells (Fig. 3a). Cell supernatants were also harvested and virus titres were assayed. The results showed that more than a 105-fold Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 17:34:50 Journal of General Virology 96 VP5 autocleavage is critical for aquareovirus infection (a) VP5 VP5N VP5C Native VP5 N42-P43 Mutated VP5 A42-P43 (b) VP5 VP7 Hoechst Merger VP5N42A VP7 Hoechst Merger (c) vAcGCRVVP5/VP7 GCRV Mock vAcGCRVVP5N42A/VP7 VP5 VP5C VP7 (d) vAcGCRV-VP5/VP7infected cells Lysate IP Mock Lysate IP: VP7 Mock infected cells WB: VP5 VP5 VP5C WB: VP7 VP7 IP: VP5 IP vAcGCRV-VP5N42A/VP7- WB: VP7 VP7 WB: VP5 VP5 VP5C reduction in virus titre was observed with N-GCRV or RGCRV when the cells were treated with NH4Cl (Fig. 3b). These results indicated that NH4Cl could inhibit the infection with both N-GCRV and R-GCRV, confirming the similarity between them. The infectivity of mutant VP5N42A/VP7 R-GCRV was evaluated on CIK cells. At 24 h p.i., a large number of plaques was observed in VP5/VP7 R-GCRV-infected cells but not in core- or VP5N42A/VP7 R-GCRV-infected cells (Fig. 3c). To further confirm that recoated particles with mutant VP5N42A were defective in infection, the virus titres were examined using both TCID50 and real-time quantitative PCR (qPCR) assays. As shown in Fig. 3(d), the infectivity of WT R-GCRV was similar to that of N-GCRV. http://vir.sgmjournals.org Fig. 1. Identification of recombinant protein expression in infected cells. (a) Schematic representation of the VP5 protein and its mutation at the autocleavage site. The arrow indicates the putative autocleavage site. (b) Immunofluorescence assays of VP5 or VP5N42A and VP7 in vAcGCRV-VP5/VP7- or vAcGCRVVP5N42A/VP7-infected cells. Hoechst dye was used to stain nuclei and a merged image is shown on the right. (c) WB analyses of VP5 or VP5N42A and VP7 in vAcGCRV-VP5/VP7 or vAcGCRV-VP5N42A/VP7 infected cells. GCRV infected CIK cells and uninfected Sf9 cells served as positive and negative controls respectively. (d) Co-immunoprecipitation assays of VP5 or VP5N42A with VP7 in infected cells. In contrast, little or no infectivity was detected with either control virus cores or mutant VP5N42A/VP7 R-GCRV particles. In the qPCR assay, the mRNAs of the GCRV core proteins VP3 and VP6 were detected to assess their expression levels. High mRNA levels were detected in WT R-GCRV-infected cells but not in cells infected with mutant VP5N42A/VP7 R-GCRV, or with virus cores or cores mixed with mock Sf9 cell lysates (Fig. 3e). In addition, WB analysis showed that the expression of VP3 and VP6 could be detected in WT R-GCRV-infected cells but not in mutant VP5N42A/VP7 R-GCRV-infected cells (Fig. 3f). These results strongly indicated that the WT RGCRVs have similar infectivity to N-GCRV but that infectivity is lost when the N42 residue is substituted to Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 17:34:50 1797 S. Yan and others (a) Core 100 nm (b) (c) R-GCRV N-GCRV 100 nm 100 nm (d) e or C V CR P7 V G c V CR vA P5/ G N-V V CR -G (f) R VP1–VP3 8 VP4 VP5 VP5C VP7 log10 p.f.u. ml–1 VP6 Core 6 R-GCRV N-GCRV 4 VP5 R-GCRV VP7 R-GCRV 2 (e) VP5 VP5C 0 VP7 Fig. 2. Characteristics and infectivity assays of in vitro-recoated particles. (a) TEM image of purified GCRV cores. The boxed area in the image is enlarged to show details. The arrow indicates the turret. (b, c) TEM images of purified R-GCRV and N-GCRV. (d, e) SDS-PAGE analysis of protein components and WB analysis of VP5 and VP7 in purified R-GCRV and N-GCRV. Purified core and vAcGCRV-VP5/VP7 served as controls. (f) Infectious titre of R-GCRV determined by a p.f.u. assay. The core, N-GCRV, VP5 R-GCRV and VP7 R-GCRV served as controls. Each bar represents the mean±SD derived from three independent experiments. A (VP5N42A), suggesting that the autocleavage of VP5 at aa 42 is indispensable for GCRV to initiate infection. The baculovirus expression system was used for the highlevel expression of recombinant VP5 and VP7 proteins, and in vitro assembly using the expressed proteins to recoat purified GCRV cores was achieved. Infectious R-GCRV could only be produced by recoating core particles with 1798 both VP5 and VP7 proteins, and not with VP5 or VP7 singly, indicating that VP7 is required for particle assembly and cell entry. These findings not only suggest that a VP5– VP7 complex can be spontaneously assembled onto purified core particles in stoichiometric amounts, but also support a proposed structure-based mechanism whereby VP7 removal triggers a VP5 conformational change and Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 17:34:50 Journal of General Virology 96 VP5 autocleavage is critical for aquareovirus infection (a) (c) – NH4Cl + N-GCRV VP5/VP7 R-GCRV Core VP5N42A/VP7 R-GCRV R-GCRV (b) (d) N-GCRV N-GCRV+NH4Cl R-GCRV 8 R-GCRV+NH4Cl 8 log10 TCID50 ml–1 log10 p.f.u. ml–1 10 6 4 2 0 Core VP5/VP7 R-GCRV N-GCRV VP5N42A/VP7 R-GCRV 6 4 2 0 (e) (f) 107 Relative expression of core proteins 106 105 VP3 104 1 2 4 VP3 VP6 103 3 102 VP6 101 100 Actin 10–1 1 2 3 4 Fig. 3. VP5 autocleavage is indispensable for efficient infection of aquareovirus. (a) Plaque assays of R-GCRV and N-GCRV in the absence (”) or presence (+) of NH4Cl. (b) Infectious titres of supernatants harvested at 24 h p.i. in (a) were determined by a p.f.u. assay. (c) Plaque assay of VP5N42A/VP7 R-GCRV. Core and VP5/VP7 R-GCRV served as controls. (d) Infectious titre of VP5N42A/VP7 R-GCRV. Core, VP5/VP7 R-GCRV and N-GCRV served as controls. (e, f) Relative expression of VP3 and VP6 in mRNA (e) or protein (f) level calculated by qPCR or WB assay, respectively. Lanes: 1, core; 2, core mixed with Sf9; 3, VP5/ VP7 R-GCRV; 4, VP5N42A/VP7 R-GCRV. Actin was used as a loading control in (f). Data in (b), (d) and (e) represent the mean±SD derived from three independent experiments. autoproteolysis, converting a virion from a dormant state to a primed ISVP (Zhang et al., 2010). To enter host cells, a reovirus virion needs to transform into an ISVP that can penetrate the cell membrane, and this process is acid dependent (Chandran et al., 1999; Kothandaraman et al., 1998). In this study, the infectivity of N-GCRV or R-GCRV could be blocked by the weak base NH4Cl, suggesting that N-GCRVs and R-GCRVs utilize a similar route to enter host cells. More importantly, RGCRVs with a VP5N42A mutation were shown to be defective in infectivity and progeny protein expression in infected cells, indicating that an N42A substitution almost completely blocked the VP5 autocleavage. This result is consistent with a previous report in MRV where cores recoated with s3 http://vir.sgmjournals.org and m1 bearing the N42A mutation were defective for infectivity (Odegard et al., 2004), suggesting that this amino acid of m1 plays an important role in the cleavage process. In summary, an in vitro assembly model of aquareovirus was generated in this study. The results suggest that RGCRV can serve as a tool for investigating virus entry. Based on the in vitro assembly model of GCRV, the functions of the outer-capsid proteins VP5 and VP7 in virus entry could be fully investigated. To our knowledge, this study has also provided the first experimental evidence for the indispensability of aquareovirus VP5 autocleavage in cell entry by using cores recoated with a mutated VP5 protein. These results lay a foundation for further efforts to reveal the detailed entry mechanism of aquareoviruses. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 17:34:50 1799 S. Yan and others Acknowledgements Ke, L. H., Fang, Q. & Cai, Y. Q. (1990). Characteristics of a novel We express our sincere gratitude to Professor Peng Gong for editing our manuscript, Juan Liu, Wen Dai and Lanlan Zhang for their work in constructing recombinant VP5 and/or VP7 baculoviruses, and Ling Shao for her work in generating mutant VP5 recombinant and the related VP5N42A and VP7 baculoviruses. This work was supported in part by grants from the National Natural Science Foundation of China (31172434, 31372565 and 31370190). isolate of grass carp hemorrhage virus. Acta Hydrobiol Sin 14, 153– 159. King, A. M. Q., Adams, M. J., Carstens, E. B. & Lefkowitz, E. J. (editors) (2011). Virus Taxonomy. Ninth Report of the International Committee on Taxonomy of Viruses. San Diego: Elsevier. Kothandaraman, S., Hebert, M. C., Raines, R. T. & Nibert, M. L. (1998). No role for pepstatin-A-sensitive acidic proteinases in reovirus infections of L or MDCK cells. Virology 251, 264–272. Liemann, S., Chandran, K., Baker, T. S., Nibert, M. L. & Harrison, S. C. (2002). Structure of the reovirus membrane-penetration protein, m1, in a complex with is protector protein, s3. Cell 108, 283–295. References Chandran, K. & Nibert, M. L. (1998). Protease cleavage of reovirus capsid protein m1/m1C is blocked by alkyl sulfate detergents, yielding a new type of infectious subvirion particle. J Virol 72, 467–475. Mohd Jaafar, F., Goodwin, A. E., Belhouchet, M., Merry, G., Fang, Q., Cantaloube, J. F., Biagini, P., de Micco, P., Mertens, P. P. C. & Attoui, H. (2008). Complete characterisation of the American grass carp Chandran, K., Walker, S. B., Chen, Y., Contreras, C. M., Schiff, L. A., Baker, T. S. & Nibert, M. L. (1999). In vitro recoating of reovirus cores with baculovirus-expressed outer-capsid proteins m1 and s3. J Virol reovirus genome (genus Aquareovirus: family Reoviridae) reveals an evolutionary link between aquareoviruses and coltiviruses. Virology 373, 310–321. 73, 3941–3950. Nibert, M. L., Schiff, L. A. & Fields, B. N. (1991). Mammalian Cheng, L., Fang, Q., Shah, S., Atanasov, I. C. & Zhou, Z. H. (2008). reoviruses contain a myristoylated structural protein. J Virol 65, 1960–1967. Subnanometer-resolution structures of the grass carp reovirus core and virion. J Mol Biol 382, 213–222. Cheng, L., Zhu, J., Hui, W. H., Zhang, X., Honig, B., Fang, Q. & Zhou, Z. H. (2010). Backbone model of an aquareovirus virion by cryo- electron microscopy and bioinformatics. J Mol Biol 397, 852–863. Fang, Q., Ke, L. H. & Cai, Y. Q. (1989). Growth characterization and high titre culture of GCHV. Virol Sin 4, 315–319. Fang, Q., Shah, S., Liang, Y. Y. & Zhou, Z. H. (2005). 3D reconstruc- tion and capsid protein characterization of grass carp reovirus. Sci China C Life Sci 48, 593–600. Nibert, M. L., Odegard, A. L., Agosto, M. A., Chandran, K. & Schiff, L. A. (2005). Putative autocleavage of reovirus m1 protein in concert with outer-capsid disassembly and activation for membrane permeabilization. J Mol Biol 345, 461–474. Odegard, A. L., Chandran, K., Zhang, X., Parker, J. S., Baker, T. S. & Nibert, M. L. (2004). Putative autocleavage of outer capsid protein m1, allowing release of myristoylated peptide m1N during particle uncoating, is critical for cell entry by reovirus. J Virol 78, 8732–8745. Fang, Q., Seng, E. K., Ding, Q. Q. & Zhang, L. L. (2008). Rangel, A. A. C., Rockemann, D. D., Hetrick, F. M. & Samal, S. K. (1999). Identification of grass carp haemorrhage virus as a new Characterization of infectious particles of grass carp reovirus by treatment with proteases. Arch Virol 153, 675–682. Sturzenbecker, L. J., Nibert, M., Furlong, D. & Fields, B. N. (1987). Jané-Valbuena, J., Nibert, M. L., Spencer, S. M., Walker, S. B., Baker, T. S., Chen, Y., Centonze, V. E. & Schiff, L. A. (1999). Reovirus virion- Intracellular digestion of reovirus particles requires a low pH and is an essential step in the viral infectious cycle. J Virol 61, 2351–2361. like particles obtained by recoating infectious subvirion particles with baculovirus-expressed s3 protein: an approach for analyzing s3 functions during virus entry. J Virol 73, 2963–2973. Zhang, X., Jin, L., Fang, Q., Hui, W. H. & Zhou, Z. H. (2010). 3.3 Å Cryo-EM structure of a nonenveloped virus reveals a priming mechanism for cell entry. Cell 141, 472–482. 1800 genogroup of aquareovirus. J Gen Virol 80, 2399–2402. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 17:34:50 Journal of General Virology 96