Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Cytokinesis wikipedia , lookup
Extracellular matrix wikipedia , lookup
Cellular differentiation wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cell culture wikipedia , lookup
Tissue engineering wikipedia , lookup
Cell encapsulation wikipedia , lookup
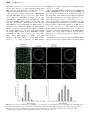
Journal of Experimental Botany, Vol. 62, No. 8, pp. 2949–2957, 2011 doi:10.1093/jxb/err007 Advance Access publication 28 February, 2011 This paper is available online free of all access charges (see http://jxb.oxfordjournals.org/open_access.html for further details) RESEARCH PAPER Differential effects of the brefeldin A analogue (6R)-hydroxy-BFA in tobacco and Arabidopsis Markus Langhans1, Sebastian Förster2, Günter Helmchen2 and David G. Robinson1,* 1 2 Department of Cell Biology, Heidelberg Institute for Plant Sciences, University of Heidelberg, D-69120 Heidelberg, Germany Institute for Organic Chemistry, University of Heidelberg, D-69120 Heidelberg, Germany * To whom correspondence should be addressed. E-mail: [email protected] Abstract The effects of two brefeldin A (BFA) analogues (BFA lactam; 6(R)-hydroxy-BFA) on plant cells were tested. Although these two compounds elicited BFA-like effects in mammalian cells, the lactam analogue failed to elicit a response in plant cells. By contrast, while the 6(R)-hydroxy-BFA analogue gave rise to a classic BFA response in tobacco mesophyll protoplasts and true leaves of Arabidopsis (redistribution of Golgi enzymes into the ER), it failed to cause the formation of BFA-compartments in Arabidopsis root cells and cotyledonary leaves. Even when the GNL1-LM mutant of Arabidopsis, which has a cis-Golgi located BFA-sensitive ARF-GEF, was used, the 6(R)-hydroxy analogue failed to elicit a response at conventional BFA concentrations. Only at concentrations of over 200 mM did 6(R)-hydroxy-BFA elicit a BFA-like effect. These differences are interpreted in terms of the different properties of the respective TGN- (Arabidopsis roots) and cis-Golgi- (tobacco mesophyll) localized BFA-sensitive ARF-GEFs. Key words: 6(R)-hydroxy-BFA, ARF-GEFs, Arabidopsis, BFA, tobacco mesophyll. Introduction One of the most useful tools to study vesicle-mediated trafficking in eukaryotic cells is the fungal macrocyclic lactone brefeldin A (Klausner et al., 1992; Gaynor et al., 1998; Nebenführ et al., 2002). It causes the rapid release of vesicle coat proteins such as coatomer and clathrin triskelions and adaptors into the cytosol, and prevents further vesicle formation (Donaldson et al., 1990; Robinson and Kreis, 1992; Ritzenthaler et al., 2002). As a consequence, the integrity of several subcellular compartments is severely perturbed. Early observations on mammalian cells pointed to two intracellular sites of BFA action: one situated in the early Golgi leading to a redistribution of Golgi membranes and luminal contents into the ER (Lippincott-Schwartz et al., 1989; Sciaky et al., 1997), the other lying in the transGolgi network (TGN) leading, together with endosomal membranes, to the formation of a tubular-vesicular aggregate called the ‘BFA-compartment’ (Chege and Pfeffer, 1990; Lippicott-Schwartz et al., 1991; Wood et al., 1991). Despite these heavily publicized effects of BFA, other BFA-induced perturbations, for example, in membrane lipids (Mérigout et al., 2002), cannot be ruled out. A BFA-induced resorption of Golgi cisternae into the ER was first recorded some 12 years ago for tobacco leaf epidermal cells (Boevink et al., 1998), and later confirmed in suspension-cultured tobacco BY-2 cells (Ritzenthaler et al., 2002). On the other hand, the induction of a BFAcompartment in plants was most prominently observed in root cells of maize (Henderson et al., 1994; Baluska et al., 2002) and Arabidopsis thaliana (Geldner et al., 2003; Grebe et al., 2003). In both of these cases, and in contrast to the situation with tobacco, a loss of Golgi cisternae after BFA treatment was not observed. It is now generally thought that the apparent discrepancy in BFA action on plant cells is a consequence of the subcellular location of ARF-GEFs, which can be either BFA-resistant or BFA-sensitive (Richter et al., 2007; Teh and Moore, 2007). Since both types of ARF-GEFs are equally capable of nucleotide exchange, there appears to be no evolutionary advantage in Abbreviations: ARF-GEFs, guanine-nucleotide exchange factors for ADP-ribosylation factor GTPases; CHX, cycloheximide; GNL1, GNOM-like-1; Man1, a-mannosidase 1; ST, sialyl transferase; SYP61, syntaxin of plants 61; VHA-a1, the a1 subunit of the vacuolar H+-ATPase. ª 2011 The Author(s). This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/bync/2.0/uk/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. 2950 | Langhans et al. expressing either one or the other form (Anders and Jürgens, 2008; Richter et al., 2009), and indeed Golgi stack BFA resistance or sensitivity may vary from tissue to tissue within the same plant (Robinson et al., 2008a). The Arabidopsis genome encodes 9–12 ARF-GTPases and eight ARF-GEFs (Jürgens and Geldner, 2002). Unlike other eukaryotes, ARF-GEFs in plants are all large (170–200 kDa), and fall into two subfamilies: the GGG class with GNOM, GNL1, GNL2, and the BIG class with five representatives. Interestingly, whereas all large ARF-GEFs in yeast and mammals are BFA-sensitive, in Arabidopsis two (GNL1 and BIG3) are BFA-resistant (Anders and Jürgens, 2008). Resistance to BFA is conferred by specific residues in a 40 amino acid stretch within the catalytic SEC7 domain of these ARF-GEFs (Cherfils and Melancon, 2005; Anders and Jürgens, 2008). Thus, L696 confers BFA-resistance to GNL1, and when mutated to M causes GNL1 to become BFA-sensitive (Richter et al., 2007). Conversely, the BFA-sensitive GNOM can be made resistant in the M696L mutant (Geldner et al., 2003). Of the eight ARF-GEFs in Arabidopsis, only GNOM and GNL1 (GNOM-like-1) have been localized. GNOM was discovered as an essential factor mediating the recycling of the auxin efflux carrier PIN1 from an otherwise undefined endosomal compartment to the plasma membrane in root cells of Arabidopsis (Geldner et al., 2003; Kleine-Vehn et al., 2008). The recycling compartment is presumably an early endosome (EE), and as such may be a domain of the TGN which, in plants, assumes the function of an EE (Richter et al., 2007; Robinson et al., 2008b). On the other hand, GNL1 localizes to the Golgi stack (Richter et al., 2007; Teh and Moore, 2007), and when mutated to the BFA-sensitive L696M form, causes Golgi resorption into the ER in Arabidopsis root cells upon exposure to BFA (Teh and Moore, 2007). In addition to its effects on protein trafficking in the secretory and endocytic pathways, BFA also shows strong Fig. 1. Effects of BFA on TGN markers in Arabidopsis roots and tobacco mesophyll protoplasts. An Arabidopsis line stably expressing the TGN marker SYP61-CFP (a, b) and tobacco mesophyll protoplasts transiently expressing SYP61-YFP and the cis-Golgi marker Man1-RFP (c–h; e and h are merge images) were used. (i–j) Measurements of the sizes of TGN punctae in control protoplasts (i) and protoplasts after BFA treatment (j). Magnification: bars¼10 lm (a, b), 5 lm (c–h). BFA analogues in tobacco and Arabidopsis | 2951 Fig. 2. Golgi stack and TGN located ARF-GEFs in Arabidopsis and tobacco. Comparison of sensitivities towards BFA and (6R)-hydroxy-BFA. cytostatic activity against many cancer cell lines (Betina et al., 1962). However, because of its poor availability and poor pharmacological properties, BFA has not established itself as an anti-cancer drug. Thus, several analogues have been synthesized in order to improve solubility (Anadu et al., 2006; Förster et al., 2011). One of these: (6R)-hydroxy-BFA, which elicits BFA-like effects in mammalian cells (Förster et al., 2011), is shown here to generate similar responses to BFA in tobacco protoplasts. However, this compound was found to be incapable of causing the formation of a BFA-compartment in Arabidopsis root cells. Even with the GNL1 LM mutant the (6R)-hydroxy analogue is much less effective than BFA in eliciting a response. Materials and methods Stable transformed plant material Plants of Arabidopsis thaliana stably transformed with VHA-a1-GFP (Dettmer et al., 2006), VHA-a1-RFP/N-ST-YFP (Dettmer et al., 2006), GNL1-VHA-a1-GFP (Richter et al., 2007), GNL1(LM)VHA-a1-GFP, GNL1(LM)-N-ST-YFP (Richter et al., 2007) were grown from surface-sterilized seeds in half-strength Murashige and Skoog medium (Murashige and Skoog, 1962) 2952 | Langhans et al. lM, 180 lM, and 360 lM) or BFA lactam or (6R)-hydroxy-BFA (36 lM, 90 lM, 180 lM, and 360 lM). BFA and BFA analogues were dissolved in a stock solution in DMSO. In these experiments, plant material was pre-treated with BFA or BFA-analogues (36 lM, 90 lM, 180 lM, and 360 lM) for 30 min before observing in the CLSM. In special cases, plant material was also treated 1 h with cycloheximide (70 lM) for 30 min before treatment with BFA (36 lM or 90 lM). Details concerning the synthesis and structure of the BFA analogues used in this paper are described in Förster et al. (2011). Confocal microscopy 80 ll protoplast solution of N. tabacum was pipetted in an area (10315 mm) bordered with a frame of 100 lm thick plastic isolating tape on a slide to protect the protoplasts from external pressure. The area was covered with a cover slip (24332 mm). In addition, A. thaliana roots and leaves (cotyledons and true leaves) were transferred to slides. Cells or plant material were observed under a Zeiss Axiovert LSM510 Meta microscope using a CApochromat 633/1.2 W corr water immersion objective for protoplasts and a Plan-Neofluoar 253/0.8 Imm corr DIC for Arabidopsis roots and leaves. Special settings were designed for observing single or double expression with different XFPconstructs. Fluorescence was detected by the Metadetector using main beam splitters HFT 488/543 and HFT 458/514. The following fluorophores (excited and emitted by frame switching in the singleor multi-tracking mode) were used: GFP (488 nm/496–518 nm), YFP (514 nm/529–550 nm), and RFP (543 nm/593–625 nm). Pinholes were adjusted to 1 Airy Unit for each wavelength. Postacquisition image processing was performed using the Zeiss LSM 510 image Browser (4.2.0.121) and CorelDrawX4 (14.0.0.567). Fig. 3. Effects of BFA and BFA-analogues on Arabidopsis and tobacco. Arabidopsis root cells stably expressing the fluorescent TGN marker VHA-a1-GFP, and tobacco mesophyll protoplasts transiently expressing the cis-Golgi marker Man1-RFP were tested with BFA, (6R)-hydroxy-BFA, and BFA-lactam at 90 lM for 30 min before observing in the CLSM. Magnification: bars¼10 lm (a–d), 5 lm (e–h). with 1% (w:v) sucrose in a controlled climate room at 22 C with a 16 h day length. Transient expression in protoplasts Plants of Nicotiana tabacum cv. Petit Havana were grown from surface-sterilized seeds in Murashige and Skoog medium with 2% sucrose in a controlled room at 24 C with a 16 h day length at a light irradiance of 200 mE m2 s1. Preparation of tobacco leaf protoplasts was done exactly as described in Foresti et al. (2006). A total volume of 500 ll of the obtained protoplasts mix was pipetted into a disposable 1 ml plastic cuvette and mixed with an appropriate amount of plasmid DNA (Man1-RFP, YFP-Syp61; Bubeck et al., 2008; Langhans et al., 2008) dissolved in 100 ll of electroporation buffer. The protoplasts were electroporated with stainless steel electrodes at a distance of 3.5 mm, using a complete exponential discharge of a 1000 lF capacitor charged at 160 V. After 30 min of absolute rest, electroporated protoplasts were removed from the cuvettes and transferred to 5 cm Petri dishes with 2 ml of TEX buffer. Protoplasts were then incubated for 24 h at 25 C in a dark chamber. Treatments with BFA and BFA analogues Experiments were performed with 0 lM BFA or BFA analogues, as a control or with different concentrations of BFA (36 lM, 90 Results BFA compartments are produced in both tobacco and Arabidopsis, but of different sizes Whereas, in mammalian cells, the TGN and EE are separate organelles, in higher plants the TGN also functions as an EE (Dettmer et al., 2006; Lam et al., 2009; Otegui and Spitzer, 2008). In addition, the TGN is not permanently fixed at the trans pole of the Golgi stack, but can detach and move separately from the stack (Viotti et al., 2010). The application of BFA emphasizes this subdivision of the Golgi apparatus (‘Golgi split’, Nebenführ et al., 2002), since BFAsensitive ARF-GEFs reside either at the TGN or, additionally, in the cisternae of the Golgi stack. In the former case, as in Arabidopsis root cells and in epidermal cells of the cotyledons, the Golgi stack appears to be unaffected and the release of the TGN continues. Since BFA prevents vesiculation (presumably of clathrin-coated vesicles) at the TGN, the released malfunctioning TGNs accumulate to form a ‘BFA-compartment’. This can easily be demonstrated with any bona fide fluorescent TGN marker, for example, SYP61 (Fig. 1a, b). However, when a BFAsensitive ARF-GEF is also present in the Golgi stack, as in tobacco, the cisternae of the stack are resorbed into the ER (Fig. 1, compare d and g). Accordingly, there will be no ongoing replenishment of TGN, and, as a result, large BFA-compartments are not formed. Instead, the TGN spots only enlarge roughly 2–3-fold (Fig. 1, compare c and f, and i and j). This observation probably reflects the BFA analogues in tobacco and Arabidopsis | 2953 Fig. 4. Comparison of the effects of different concentrations of BFA and (6R)-hydroxy-BFA on Arabidopsis root cells expressing VHA-a1-GFP and tobacco protoplasts expressing Man1-RFP. Standard treatment period of 30 min. Magnification: bars¼10 lm (a–i), 5 lm (j–r). temporary continuation of cisternal maturation and TGN release until the drug has achieved its inhibitory effect on the rest of the Golgi stack. The location of BFA-sensitive ARF-GEFs in the Golgi apparatus of different higher plants together with the two basic morphological responses after BFA addition are summarized in Fig. 2. BFA lactam does not elicit a BFA-like effect on Arabidopsis roots or tobacco protoplasts Although reported to have a BFA-like effect on mammalian cells (Förster et al., 2011), the BFA lactam analogue was unable to induce BFA compartments in Arabidopsis root cells (Fig. 3, compare a and b with c and d). In addition, it did not cause a redistribution of the fluorescent cis-Golgi marker Man1-RFP into the ER in tobacco mesophyll protoplasts (Fig. 3, compare e with h). Thus the BFA- lactam analogue, at the concentrations tested here, seems to be without effect on plant cells. (6R)-hydroxy-BFA elicits a BFA-like effect on tobacco protoplasts but not on Arabidopsis (6R)-hydroxy-BFA, which shows BFA-like effects on mammalian cells (Förster et al., 2011), also caused a typical BFA-phenotype when applied to tobacco mesophyll protoplasts (Fig. 3, compare f and g). However, it was less effective than BFA at lower concentrations (below 50 lM), with punctate signals for the Golgi marker Man1-RFP still being visible in addition to an ER-labelling (Fig. 4, compare k and o with l and p). By contrast, as shown in Fig. 3 (compare b and c), (6R)-hydroxy-BFA failed to induce BFA compartments in Arabidopsis roots. This was not a question of concentration. Even when presented at the 2954 | Langhans et al. extremely high concentration of 180 lM this BFA analogue failed to generate the production of a BFA compartment (Fig. 4, compare b–e with f–i). This was also not achieved with prolonged (up to 4 h) treatment periods (data not shown). To determine whether the failure to elicit a BFA-effect was not restricted to the roots of Arabidopsis, the (6R)-hydroxy-BFA analogue was tested on epidermal cells in cotyledons and true leaves of Arabidopsis plants stably expressing the TGN marker VHA-a1-RFP and the transGolgi marker ST-YFP (Dettmer et al., 2006). Surpisingly, in response to BFA, the cotyledon epidermal cells behaved like root cells producing typical BFA-compartments (Fig. 5a, b). By contrast, and as previously reported by Robinson et al. (2008a), BFA caused a redistribution of the Golgi marker ST-YFP into the ER in the epidermal cells of true leaves (Fig. 5d, e). However, in neither type of epidermal cell did the (6R)-hydroxy-BFA analogue elicit any change in the TGN/Golgi marker proteins (Fig. 5c, f). (6R)-hydroxy-BFA fails to elicit a BFA effect in the Arabidopsis GNL1-LM- mutant Fig. 5. Comparison of the effects of BFA and (6R)-hydroxy-BFA on cotyledon leaves and real leaves of Arabidopsis plants stably expressing VHA-a1-RFP and ST-YFP. Squares of leaf tissue were excised and incubated in aqueous solutions containing 90 lM drug for 30 min. The inset in (b) is a high magnification view of a single BFA compartment with a red core derived from the TGN marker surrounded by yellow Golgi stacks. The arrows in (e) point to the redistribution of the Golgi marker ST-YFP into the nuclear envelope (¼ER). Magnification: bars¼10 lm. In wild-type Arabidopsis roots, BFA induces the formation of the BFA compartment, but does not affect Golgi stacks which lie outside of this structure (Fig. 6a, b). However, in the GNL1-LM mutant of Arabidopsis the Golgi stacklocated ARF-GEF also becomes BFA-sensitive, leading to a redistribution of the trans-Golgi marker ST-YFP into the ER upon BFA treatment (Fig. 6i, j). In addition, in the GNL1-LM mutant plants large BFA compartments are not formed. This can be clearly seen in mutant lines expressing the TGN marker VHA-a1-GFP, the signal for which enlarges some 2-fold over a 20 min treatment period (see Fig. 6. Comparison of BFA (90 lM) effects in root cells of wt and GNL1-LM mutant Arabidopsis lines. (a–d) Formation of the BFA compartment (b) is prevented by cycloheximide (c). (e–h) BFA causes the TGN marker VHA-a1-GFP to be detected in the ER after 30 min (f). This is prevented by cycloheximide (g). BFA causes the trans-Golgi marker ST-YFP to redistribute into the ER (j). This is only partially prevented by cycloheximide (k). Control treatments with cycloheximide alone are presented in (c), (g), and (k). Magnification: bars¼10 lm. BFA analogues in tobacco and Arabidopsis | 2955 rather the new synthesis of this protein which cannot exit the ER. This was established in control experiments where the roots were pretreated with the protein synthesis inhibitor cycloheximide This inhibitor prevented the appearance of an ER signal in the GNL1-LM line expressing VHA-a1-GFP (Fig. 6g), and only partially prevented the BFA-induced redistribution of ST-YFP into the ER (Fig. 6k). The different effects of BFA on the trans-Golgi and TGN-based fluorescent markers in the GNL1-LM mutant prompted us to examine the effect of the (6R)hydroxy-BFA analogue on this mutant line. As shown in Fig. 7, the (6R)-hydroxy-BFA analogue did not elicit any changes in the punctate appearance of the TGN-marker VHA-a1-GFP (Fig. 7a–d). In contrast, the analogue did cause the redistribution of the trans-Golgi marker ST-YFP into the ER, but only at concentrations much higher than 100 lM (Fig. 7f–i). The inclusion of cycloheximide caused no change in these responses (Fig. 7e, j). The effects of (6R)-hydroxy-BFA on tobacco and Arabidopsis are summarized in Fig. 2. Discussion BFA and cisternal maturation in the Golgi stack BFA-induced Golgi stack disassembly probably provides the best evidence for cisternal maturation in a cis/trans direction. Working with tobacco BY-2 cells expressing the cis- Golgi marker Man1-GFP, Ritzenthaler et al. (2002) noticed that this signal continued to remain in the stack after BFA had caused the cisternae to be reduced in number to 2 or 3. Since the remaining cisternae had a trans-like morphology, it was considered that the cis-cisternae were being gradually transformed as they moved upwards through the stack. This observation has recently been confirmed by Schoberer et al. (2010) who, in addition to providing convincing evidence for a stepwise trans-directed cisternal transport in tobacco epidermal cells, also showed that Golgi stack disassembly begins with the loss of translocated proteins. Most significantly, and as demonstrated in our experiments with the GNL1 mutant, once a Golgi cisterna has established itself as a TGN its proteins cannot be redistributed into the ER after BFA treatment. BFA analogues and modes of BFA action Fig. 7. Examination of the effects of (6R)-hydroxy-BFA on root cells of GNL1-LM Arabidopsis lines expressing either VHA-a1-GFP (a–e) or ST-YFP (a–i) or marker proteins. Treatment time¼30 min. Magnification: bars¼10 lm. Supplementary Fig. S1 at JXB online). Interestingly, an ER distribution for the VHA-a1-GFP does appear after 30 min of BFA treatment (Fig. 6f). However, this signal does not represent the relocation of the TGN-based protein, but It has long been held that BFA has two different modes of action in mammalian cells: one causing an inhibition of vesicle-mediated protein trafficking, and the other leading to apoptosis (Nojiri et al., 1995). However, it has been shown that stressing the ER through prolonged BFA treatment can also lead to cell death via caspase activation (Rao et al., 2001; Murakami et al., 2007). Nevertheless, there are some BFA analogues which do not induce apoptosis but do affect protein trafficking and Golgi morphology (Anadu et al., 2006). There are also other BFA analogues which effect neither secretion or Golgi 2956 | Langhans et al. structure (Brüning et al., 1992; Klausner et al., 1992; Förster et al., 2011). Thus, a BFA analogue which, in some way, leads to a modified BFA response or is active in mammals but not plants (or vice versa) is obviously a useful tool to have in terms of understanding BFA/ARF-GEF interactions. One such example is a BFA lactam analogue, which is active in mammalian cells (Förster et al., 2011), but is without effect on plants (this study). Another example is the naturally occurring 7-dehydro-brefeldin A (7-oxo-BFA), which has been shown to be much more potent than BFA in perturbing Golgi-based secretion in suspension-cultured sycamore cells (Driouich et al., 1997). There is no obvious reason why the BFA-lactam analogue should elicit BFA-like effects in mammalian cells but not in plants. According to molecular modelling, the lactam analogue exists in two conformations, one of which is very similar to BFA, the other less so. In solution, sufficient amounts of the former should be present to allow for the generation of BFA-like effects (Förster et al., 2011). Structural studies on the (6R)-hydroxy BFA analogue also indicate that it has almost or basically the same conformation as BFA and fits equally well into the SEC7 BFA-binding domain (Förster et al., 2011). In principle, it should therefore work equally well as BFA on all plants, but as we have seen it has a differential effect. In wild-type Arabidopsis roots, whose cells have a BFA-sensitive ARFGEF (GNOM) only at the TGN, it is without effect. Even in the GNL1-L696M mutant, (6R)-hydroxy-BFA was unable to elicit this effect, unless presented at concentrations 5–10 times higher than is normally required to elicit a BFA effect. This suggests that the presence of the amino acid L696 (in Arabidopsis GNOM) is not the only factor in an ARF-GEF which determines its BFA resistance. This is supported by the fact that tobacco, which has a naturally occurring BFA-sensitive GNL1 homologue (with M683 in the SEC7 domain; Wang et al., 2008), first responds to (6R)-hydroxy BFA at three times higher concentrations than BFA. Obviously, an explanation for the lack of effect of the BFA-lactam analogue on plants as a whole, and the differential sensitivities of GNOM and GNL1 towards the (6R)-hydroxy BFA analogue, will only be possible after a careful comparison of the SEC7 domains in the various organisms has been carried out. Supplementary data Supplementary data can be found at JXB online. Supplementary Fig. S1. Time-course of BFA action on root cells of a GNL1-LM Arabidopsis line expressing VHA-a1-GFP. Acknowledgements We greatly appreciate the financial support of the German Research Council (DFG grant RO 440/ 11-4). References Anadu NO, Davisson VJ, Cushman M. 2006. Synthesis and anticancer activity of brefeldin A ester derivatives. Journal of Medicinal Chemistry 49, 3897–3905. Anders N, Jürgens G. 2008. Large ARF guanine nucleotide exchange factors in membrane trafficking. Cellular and Molecular Life Sciences 65, 3433–3445. Baluska F, Hlavacka A, Samaj J, Palme K, Robinson DG, Matoh T, McCurdy DW, Menzel D, Volkmann D. 2002. F-actin-dependent endocytosis of cell wall pectins in meristematic root cells. Insights from brefeldin A-induced compartments. Plant Physiology 130, 422–431. Betina V, Horáková K, Baráth Z. 1962. Anti-HeLa cell effect of cyanein. Naturwissenschaften 49, 241. Boevink P, Oparka K, Santa Cruz S, Martin B, Betteridge A, Hawes C. 1998. Stacks on tracks: the plant Golgi apparatus traffics on an actin/ER network. The Plant Journal 15, 441–447. Brüning A, Karrenbauer A, Schnabel E, Wieland FT. 1992. Brefeldin A-induced increase of sphingomyelin synthesis. Assay for the action of the antibiotic in mammalian cells. Journal of Biological Chemistry 267, 5052–5025. Bubeck J, Scheuring D, Hummel E, Langhans M, Viotti C, Foresti O, Denecke J, Banfield DK, Robinson DG. 2008. The syntaxins SYP31 and SYP81 control ER-Golgi trafficking in the plant secretory pathway. Traffic 9, 1629–1652. Chege NW, Pfeffer SR. 1990. Compartmentation of the Golgi complex: brefeldin-A distinguishes trans-Golgi cisternae from the trans-Golgi network. Journal of Cell Biology 111, 893–899. Cherfils J, Melancxon P. 2005. On the action of Brefeldin A on Sec7-stimulated membrane-recruitment and GDP/GTP exchange of Arf proteins. Biochemical Society Transactions 33, 635–638. Dettmer J, Hong-Hermesdorf A, Stierhof YD, Schumacher K. 2006. Vacuolar H+-ATPase activity is required for endocytic and secretory trafficking in Arabidopsis. The Plant Cell 18, 715–730. Donaldson JG, Lippincott-Schwartz J, Bloom GS, Kreis TE, Klausner RD. 1990. Dissociation of a 110-kD peripheral membrane protein from the Golgi apparatus is an early event in brefeldin A action. Journal of Cell Biology 111, 2295–2306. Driouich A, Jauneau A, Staehelin LA. 1997. 7-Dehydrobrefeldin A, a naturally occurring brefeldin A derivative, inhibits secretion and causes a cis-to-trans breakdown of Golgi stacks in plant cells. Plant Physiology 113, 487–492. Foresti O, daSilva LL, Denecke J. 2006. Overexpression of the Arabidopsis syntaxin PEP12/SYP21 inhibits transport from the prevacuolar compartment to the lytic vacuole in vivo. The Plant Cell 18, 2275–2293. Förster S, Persch E, Tverskoy O, Rominger F, Helmchen G, Klein C, Gönen B, Brügger B. 2011. Syntheses and biological properties of brefeldin analogues. European Journal of Organic Chemistry doi:10.1002/ejoc200. Gaynor EC, Graham TR, Emr SD. 1998. COPI in ER/Golgi and intraGolgi transport: do yeast COPI mutants point the way? Biochimica et Biophysica Acta 1404, 33–51. BFA analogues in tobacco and Arabidopsis | 2957 Geldner N, Anders N, Wolters H, Keicher J, Kornberger W, Muller P, Delbarre A, Ueda T, Nakano A, Jürgens G. 2003. The Arabidopsis GNOM ARF-GEF mediates endosomal recycling, auxin transport, and auxin-dependent plant growth. Cell 112, 219–230. Grebe M, Xu J, Möbius W, Ueda T, Nakano A, Geuze HJ, Rook MB, Scheres B. 2003. Arabidopsis sterol endocytosis involves actin-mediated trafficking via ARA6-positive early endosomes. Current Biology 13, 1378–1387. Henderson J, Satiat-Jeunemaitre B, Napier R, Hawes C. 1994. Brefeldin A-induced disassembly of the Golgi apparatus is followed by disruption of the endoplasmic reticulum in plant cells. Journal of Experimental Botany 45, 1347–1351. Jürgens G, Geldner N. 2002. Protein secretion in plants: from the trans-Golgi network to the outer space. Traffic 3, 605–613. Klausner RD, Donaldson JG, Lippincott-Schwartz J. 1992. Brefeldin A: insights into the control of membrane traffic and organelle structure. Journal of Cell Biology 116, 1071–1080. Kleine-Vehn J, Dhonukshe P, Sauer M, Brewer PB, Wiśniewska J, Paciorek T, Benková E, Friml J. 2008. ARF GEFdependent transcytosis and polar delivery of PIN auxin carriers in Arabidopsis. Current Biology 18, 526–531. Lam SK, Cai Y, Tse YC, Wang J, Law AH, Pimpl P, Chan HY, Xia J, Jiang L. 2009. BFA-induced compartments from the Golgi apparatus and trans-Golgi network/early endosome are distinct in plant cells. The Plant Journal 60, 865–681. Langhans M, Marcote MJ, Pimpl P, Virgili-López G, Robinson DG, Aniento F. 2008. In vivo trafficking and localization of p24 proteins in plant cells. Traffic 9, 770–785. Lippincott-Schwartz J, Yuan L, Tipper C, Amherdt M, Orci L, Klausner RD. 1991. Brefeldin A’s effects on endosomes, lysosomes, and the TGN suggest a general mechanism for regulating organelle structure and membrane traffic. Cell 67, 601–616. Lippincott-Schwartz J, Yuan LC, Bonifacino JS, Klausner RD. 1989. Rapid redistribution of Golgi proteins into the ER in cells treated with brefeldin A: evidence for membrane cycling from Golgi to ER. Cell 56, 801–813. Mérigout P, Képès F, Perret AM, Satiat-Jeunemaitre B, Moreau P. 2002. Effects of brefeldin A and nordihydroguaiaretic acid on endomembrane dynamics and lipid synthesis in plant cells. FEBS Letters 518, 88–92. Nojiri H, Hori H, Nojima S. 1995. Induction of cell differentiation and apoptosis into human malignant tumour cell lines by drugs affecting ganglioside biosynthesis. Glycoconjugate Journal 12, 459. Otegui MS, Spitzer C. 2008. Endosomal functions in plants. Traffic 9, 1589–1598. Rao RV, Hermel E, Castro-Obregon S, del Rio G, Ellerby LM, Ellerby HM, Bredesen DE. 2001. Coupling endoplasmic reticulum stress to the cell death program. Mechanism of caspase activation. Journal of Biological Chemistry 276, 33869–33874. Richter S, Geldner N, Schrader J, Wolters H, Stierhof YD, Rios G, Koncz C, Robinson DG, Jürgens G. 2007. Functional diversification of closely related ARF-GEFs in protein secretion and recycling. Nature 448, 488–492. Richter S, Voss U, Jürgens G. 2009. Post-Golgi traffic in plants. Traffic 10, 819–828. Ritzenthaler C, Nebenführ A, Movafeghi A, Stussi-Garaud C, Behnia L, Pimpl P, Staehelin LA, Robinson DG. 2002. Reevaluation of the effects of brefeldin A on plant cells using tobacco Bright Yellow 2 cells expressing Golgi-targeted green fluorescent protein and COPI antisera. The Plant Cell 14, 237–261. Robinson MS, Kreis TE. 1992. Recruitment of coat proteins onto Golgi membranes in intact and permeabilized cells: effects of brefeldin A and G protein activators. Cell 69, 129–138. Robinson DG, Langhans M, Saint-Jore-Dupas C, Hawes C. 2008a. BFA effects are tissue and not just plant specific. Trends Plant Science 13, 405–408. Robinson DG, Jiang L, Schumacher K. 2008b. The endosomal system of plants: charting new and familiar territories. Plant Physiology 147, 1482–1492. Schoberer J, Runions J, Steinkellner H, Strasser R, Hawes C, Osterrieder A. 2010. Sequential depletion and acquisition of proteins during Golgi stack disassembly and reformation. Traffic [Epub ahead of print 18 Aug 2010.] Sciaky N, Presley J, Smith C, Zaal KJ, Cole N, Moreira JE, Terasaki M, Siggia E, Lippincott-Schwartz J. 1997. Golgi tubule traffic and the effects of brefeldin A visualized in living cells. Journal of Cell Biology 139, 1137–1155. Teh OK, Moore I. 2007. An ARF-GEF acting at the Golgi and in selective endocytosis in polarized plant cells. Nature 448, 493–496. Murakami Y, Aizu-Yokota E, Sonoda Y, Ohta S, Kasahara T. 2007. Suppression of endoplasmic reticulum stress-induced caspase activation and cell death by the overexpression of Bcl-xl or Bcl-2. The Journal of Biochemistry 141, 401–410. Viotti C, Bubeck J, Stierhof YD, et al. 2010. Endocytic and secretory traffic in Arabidopsis merge in the trans-Golgi network/early endosome, an independent and highly dynamic organelle. The Plant Cell 22, 1344–1357. Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 15, 473–497. Wang L, Liao FL, Zhu L, Peng XB, Sun MX. 2008. NtGNL1 is involved in embryonic cell division patterning, root elongation, and pollen tube growth in tobacco. New Phytologist 179, 81–93. Nebenführ A, Ritzenthaler C, Robinson DG. 2002. Brefeldin A: deciphering an enigmatic inhibitor of secretion. Plant Physiology 130, 1102–1108. Wood SA, Park JE, Brown WJ. 1991. Brefeldin A causes a microtubule-mediated fusion of the trans-Golgi network and early endosomes. Cell 67, 591–600.