Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
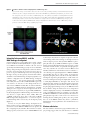
S.J. Boulton1 DNA Damage Response Laboratory, Cancer Research UK, The London Research Institute, Clare Hall Laboratories, South Mimms EN6 3LD, U.K. Colworth Medal Lecture Delivered at the SECC, Glasgow, on 26 July 2006 Simon Boulton Abstract Inherited germline mutations in either BRCA1 or BRCA2 confer a significant lifetime risk of developing breast or ovarian cancer. Defining how these two genes function at the cellular level is essential for understanding their role in tumour suppression. Although BRCA1 and BRCA2 were independently cloned over 10 years ago, it is only in the last few years that significant progress has been made towards understanding their function in cells. It is now widely accepted that both genes play critical roles in the maintenance of genome stability. Evidence implicates BRCA2 as an integral component of the homologous recombination machinery, whereas BRCA1 is an E3 ubiquitin ligase that has an impact on DNA repair, transcriptional regulation, cell-cycle progression and meiotic sex chromosome inactivation. In this article, I will review the most recent advances and provide a perspective of potential future directions in this field. Introduction Breast cancer is the second leading cause of cancer-related deaths in women and represents the leading cause of cancer Key words: BRCA1, BRCA2, DNA repair, DNA-damage sensing, tumour suppressor. Abbreviations used: ATM, ataxia telangiectasia mutated; ATR, ATM- and Rad3-related; BAP, BRCA1-associated protein; BARD1, BRCA1-associated RING domain 1; BRCT, BRCA1 C-terminus; BRIP, BRCA1-interacting protein; Cdk, cyclin-dependent kinase; CtIP, C-terminal-binding-proteininteracting protein; DSB, double-strand break; dsDNA, double-stranded DNA; FA, Fanconi’s anaemia; FANC, FA complementation group; GST, glutathione S-transferase; HR, homologous recombination; MRN, Mre11–Rad50–Nbs1; MSCI, meiotic sex chromosome inactivation; NHEJ, non-homologous end joining; OB, oligonucleotide/oligosaccharide-binding; SCF, Skp1/cullin/Fbox; siRNA, small interfering RNA; SSA, single-strand annealing; ssDNA, single-stranded DNA; Ub, ubiquitin; Xi, inactive X chromosome; XIST, Xi-specific transcript. 1 email [email protected] incidence. Of the total number of woman diagnosed with breast cancer, approx. 10% have a reported history of the disease. The fact that the disease has a high prevalence in certain families led to the notion that this subset of breast cancer cases may be attributed to inheritance of predisposing mutations in cancer-susceptibility genes [1–3]. Indeed, in 1991, Mary-Claire King and co-workers used linkage analysis to assign BRCA1, the first breast cancer-susceptibility gene, to chromosome 17 [4]. Then, 3 years later, Mark Skolnick and colleagues cloned the BRCA1 gene [5]. BRCA1 was subsequently shown to harbour truncating mutations in a large number of familial breast cancer cases, and over 300 missense variants of BRCA1 have been identified in patients to date [5]. Search for a second breast cancer-susceptibility gene was prompted by the absence of BRCA1 mutations in families with a high incidence of male breast cancer [6]. Linkage analysis mapped BRCA2 to chromosome 13, and the identity of the gene was reported in 1995 [7,8]. It is now widely accepted that individuals that inherit a germline mutation in BRCA1 or BRCA2 have a significantly increased lifetime risk of developing breast or ovarian cancer. Approx. 80% of individuals with a BRCA1 or BRCA2 mutation develop breast cancer by the age of 70 years, whereas 40% of carriers with BRCA1 mutations and 20% with BRCA2 mutations develop ovarian cancer by a similar age. Consistent with the paradigm of tumour-suppressor genes, tumours arising in individuals with BRCA1 or BRCA2 mutations consistently exhibit LOH (loss-of-heterozygosity) of the wild-type second allele [1–3]. Colworth Medal Lecture Cellular functions of the BRCA tumour-suppressor proteins BRCA1 and BRCA2 function in the maintenance of genome integrity Since their identification over a decade ago, the challenge of defining the cellular functions of BRCA1 and BRCA2 has received considerable attention. Primarily, through the analysis of chicken DT40 knockout cell lines, knockout mice and tumour-derived human cell lines, it has become widely established that both BRCA1 and BRCA2 perform roles in the maintenance of genome stability, where they function as ‘caretakers’ of the genome [9,10]. That is, cells that are defective for either BRCA1 or BRCA2 exhibit extensive genome instability. This phenotype is strikingly demonstrated through the use of spectral karyotyping, a chromosome painting technique that reveals profound chromosomal translocation, duplications and aberrant fusion events between C 2006 Biochemical Society 633 634 Biochemical Society Transactions (2006) Volume 34, part 5 non-homologous chromosomes in BRCA1- and BRCA2deficient cells [11–15]. The genome instability phenotype associated with the loss of BRCA1 or BRCA2 is a hallmark of a number of human diseases, including FA (Fanconi’s anaemia) and Bloom’s syndromes [16,17]. These diseases share a common underlying defect in DNA-repair or DNA-damagesensing processes. Indeed, the observation that BRCA1 and BRCA2 proteins form nuclear foci upon treatment with DNA-damaging agents led to the hypothesis that both genes function in the maintenance of genome stability, potentially through a direct role in DNA repair. This possibility received further support by the findings that BRCA1 and BRCA2 foci co-localize with the DNA-repair protein Rad51 that is essential for the accurate repair of DNA DSBs (double-strand breaks) [18–20]. Eukaryotic cells possess at least three distinct pathways for DSB repair including NHEJ (non-homologous end joining), SSA (single-strand annealing) and HR (homologous recombination) [21,22]. NHEJ involves the direct re-ligation of the broken DNA ends and can be error prone in nature owing to the loss of terminal bases that may be removed in order for ligation to occur efficiently. Also, NHEJ does not rely on extensive sequence recognition for repair to proceed so has the capacity to ligate DNA ends from non-homologous chromosomes. This type of aberrant repair may result in acentric or dicentric chromosomes that will pose further problems to the cell during chromosome segregation at mitosis. In the case of acentric chromosomes, the remaining chromosome fragment may be lost, whereas dicentric chromosomes may undergo breakage at mitosis, followed by repair and further aberrant rearrangements. Indeed, inappropriate utilization of NHEJ is thought to be one of the major causes of DNA rearrangements and translocation in cells. SSA, on the other hand, promotes DSB repair by annealing short stretches of sequence homology that flank the break site that are uncovered following DSB resection. Repair between short stretches of sequence homology results in the deletion of intervening sequences and loss of genetic information. In contrast with NHEJ and SSA that are error-prone in nature, HR is the predominant mechanism employed to accurately repair DSBs in S-phase and G2 -phase cells. HR is initiated by nucleolytic processing of the DSB to generate recombination-proficient ssDNA (single-stranded DNA) overhangs that are substrates for the recombinase enzyme Rad51, the eukaryotic counterpart of RecA. In the presence of ATP, Rad51 forms a helical nucleoprotein filament on ssDNA. Within the context of the nucleoprotein filament, Rad51 is able to search for an intact homologous template and then catalyse invasion of the ssDNA into a donor sister chromatid or homologous chromosome to form a joint molecule. The resulting joint molecule acts as a primer for DNA synthesis to extend the heteroduplex DNA that, following further processing and resolution of the joint DNA molecules, leads to repair of the DSB and restoration of DNA integrity [21,22]. The co-localization of Rad51, BRCA1 and BRCA2 at nuclear foci implied that these proteins function together in DNA-repair processes [19,20,23]. Although BRCA1 and C 2006 Biochemical Society BRCA2 have both been shown to associate with Rad51 in pull-down experiments [19,20,23], the interaction between BRCA1 and Rad51 appears to be indirect and the biological relevance of this interaction has not been forthcoming. However, BRCA2 does interact directly with Rad51 and is an integral component of the HR machinery in human cells (see below). Although a direct involvement for BRCA1 in HR processes has remained elusive, BRCA1-defective cells are compromised for homology-directed repair of DSBs [24], although this defect is not as pronounced as that observed in BRCA2-defective cells, such as CAPAN-1 [11]. One of the complicating issues with defining the cellular function of BRCA1 is that its loss in cells confers a pleiotropic phenotype that cannot be exclusively attributed to a defect in DNA-repair processes. In fact, BRCA1 has been implicated in a number of diverse cellular processes [25]. Xu et al. [26] demonstrated that BRCA1 is required for S-phase and G2 /M checkpoint arrest in response to DNA damage. ATM (ataxia telangiectasia mutated)-dependent phosphorylation of BRCA1 at Ser1423 is important for its role in the G2 /M checkpoint, but not for its role in the S-phase checkpoint, as mutation of this site compromised its G2 function [26]. Current evidence suggests that BRCA1 functions as a checkpoint mediator, similar to Mdc1, 53BP1 and claspin, that collectively facilitates interactions between ATM/ATR (ATM- and Rad3-related) and their substrates by mediating the spatiotemporal assembly of damage-specific multiprotein complexes at and around repair sites [27]. In BRCA1 exon 11 isoform-deficient cells, centrosome amplification is also observed that leads to the formation of multiple spindle poles in a single cell [14]. BRCA1 also has an impact on transcriptional activation. Hartman and Ford [28] showed that BRCA1 could enhance the expression of XPC (xeroderma pigmentosum C), DDB2 (damage-specific DNA-binding protein 2) and GADD45 (growth-arrest and DNA-damageinducible protein 45), independent of p53, but it is currently unclear whether this effect is direct or an indirect consequence of its roles in DNA repair and checkpoint signalling [28]. Links with transcription have also been proposed based on the observation that BRCA1 can associate with the RNA polymerase II holoenzyme complex via RNA helicase A and by the finding that the BRCT (BRCA1 C-terminus) domains at the C-terminus can function as a transcriptional activator when fused to the DNA-binding domain of Gal4 [29–33]. BRCA1 is clearly dispensable for basal transcription function and is not an essential component of RNA polymerase II, so it remains to be seen whether the association of BRCA1 with transcription is biologically relevant. Ganesan et al. [34] also identified a role for BRCA1 in supporting XIST (inactive X chromosome-specific transcript) RNA concentration required for X chromosome inactivation. They demonstrated that BRCA1 co-localizes with markers on the inactive X chromosome (Xi) and binds to XIST RNA by chromatin immunoprecipitation. In BRCA1-deficient cells, the Xi chromatin structure is altered and leads to expression of an otherwise Xi-silenced transgene tagged with GFP (green fluorescent protein) [34]. A specialized role for BRCA1 on X Cellular functions of the BRCA tumour-suppressor proteins Figure 1 BRCA1: domains, interacting proteins, activities and functions Schematic diagram of the BRCA1 protein depicting the position of the RING, nuclear localization signal (NLS) and BRCT domains. The indicated BRCA1-interacting proteins are shown under the region of BRCA1 required for their association. Interacting proteins important for BRCA1 function in vivo are shown in blue. Interacting proteins whose relevance to BRCA1 function remains unclear are shown in black. aa, amino acids. HDAC, histone deacetylase; pRB, retinoblastoma family protein. and Y chromosomes has also been suggested by Turner et al. [35]. During the pachytene stage of meiosis I, the X and Y chromosomes form a condensed chromatin domain termed the sex of XY body that is transcriptionally silent through a process referred to as MSCI (meiotic sex chromosome inactivation). ATR localization to the XY body is important for the establishment of MSCI through H2AX histone phosphorylation. Both ATR localization and H2AX phosphorylation were shown to be abolished in BRCA1-deficient cells, revealing a role for BRCA1 in MSCI [35]. The current challenge to the field is to understand how BRCA1 has an impact functionally on these different cellular processes. The domain structure of BRCA1 The human BRCA1 gene comprises 24 exons and encodes an 1863-amino-acid protein (Figure 1). Sequence conservation between mammalian species is weak, with the exception of two highly conserved domains located in the N- and Cterminal regions of the protein, which include a RING domain located in the N-terminus and two tandem BRCT motifs at the extreme C-terminal end (Figure 1). The N-terminal RING domain was identified as soon as the BRCA1 gene was cloned on the basis of homology with similar domains found in proteins that interact directly or indirectly with DNA. The RING domain of BRCA1 encompasses the first 109 amino acids. Within this region of the protein a characteristic core of approx. 50 amino acids contains a conserved pattern of seven cysteine residues and one histidine residue arranged in an interleaved fashion to form a structure responsible for co-ordinating the binding of two Zn2+ ions [36]. Unlike some RING domains, this motif in BRCA1 does not bind directly to DNA, rather it forms an interaction surface responsible for heterodimerization with a structurally related protein BARD1 (BRCA1-associated RING domain 1) [37]. Like BRCA1, BARD1 also possesses an N-terminal RING domain and two BRCT motifs at the C-terminus, in addition to three ankyrin repeats of unknown function. Of the many protein interaction partners identified with BRCA1, the significance of its association with BARD1 is beyond question and is supported by numerous in vitro and in vivo observations. Both proteins co-localize at repair and S-phase foci and require each other for focus formation [19]. BRCA1 and BARD1 mutant cell lines and knockout C 2006 Biochemical Society 635 636 Biochemical Society Transactions (2006) Volume 34, part 5 mice exhibit indistinguishable phenotypes indicative of a shared functional role in vivo [13,38,39]. Three-dimensional structural analysis of the BRCA1–BARD1 RING heterodimer complex indicates that these proteins interact through an extensive four-helix bundle formed by helices that flank the core RING motif (amino acids 24–64) [36,40]. Although mutations in the BARD1 gene associated with breast cancer predisposition are very infrequent compared with the frequency of BRCA1 mutations, a number of tumour-derived mutations have been found [41]. Despite the surprisingly low frequency of BARD1 mutations found in tumours, the interaction surface responsible for BRCA1–BARD1 heterodimerization is targeted for mutation in BRCA1. Indeed, missense mutation in five of the critical Zn2+ binding residues in the BRCA1 RING have been found in tumours and functional analysis has shown that many of these missense mutations reduce or abolish heterodimerization in vitro [9]. A number of studies also indicate that BRCA1– BARD1 heterodimerization is important for stability of the two proteins in vivo [42,43]. Collectively, these observations indicate that BRCA1 functions as a heterodimer with BARD1 in vivo and this association and the structural integrity of the RING domain is of critical importance for the tumoursuppression function of BRCA1. Cancer-predisposing missense and truncating mutations in BRCA1 are also found within the two BRCT motifs at the C-terminus of the protein indicating that, like the RING domain, the function of the BRCT motifs is also of critical importance for tumour suppression [44]. The BRCT motif is an approx. 100-amino-acid domain that is present in a number of other DNA–repair and DNA-damage-response proteins, including 53BP1, MDC1, XRCC1 and budding yeast Rad9 [45]. Structural analysis of the two BRCT motifs in BRCA1 reveals that the individual motifs form a similar structure to each other that are packed together in a head-to-tail configuration. Many of the tumour-derived missense mutations in this region of BRCA1 map to the interface between the two BRCT motifs and result in destabilization of the BRCT structure [45]. A number of BRCA1–interacting factors, such as CtIP (C-terminal-binding-protein-interacting protein), BRIP (BRCA1-interacting protein) and p300, interact via the BRCT, supporting the notion that this part of the protein functions as a multipurpose protein–protein interaction module [46,47]. Indeed, recent studies have demonstrated that BRCT motifs can function as phosphopeptide-binding sites that can mediate protein–protein interactions with phosphoproteins [48,49]. Furthermore, BRCA1 is known to bind to phosphorylated CtIP and BRIP/FANCJ (FA complementation group J) via its BRCT motifs [47]. The presence of BRCT motifs in BRCA1 and BARD1 therefore raises the possibility that binding to phosphoproteins may have important functional implications. BRCA1 function in a simple eukaryote The importance of BRCA1 to DNA-damage-repair processes is also highlighted by the conservation of certain cellular C 2006 Biochemical Society functions in the simple metazoan, Caenorhabditis elegans [50]. The nematode orthologues of BRCA1 and BARD1 (CeBRC-1 and CeBRD-1 respectively) share extensive sequence and domain conservation with their counterparts in higher eukaryotes, including RING, ankyrin and BRCT domains. Although CeBRC-1 is shorter than human BRCA1, it is highly related to the major human BRCA1 splice variant, exon delta-11. CeBRC-1 and CeBRD-1 interact to form a heterodimer via their respective RING domains [50]. As is the case in Xenopus laevis and in humans [42,43], heterodimerization of CeBRC-1 and CeBRD-1 is required for stable expression of each protein [51]. Cebrc-1 or Cebrd-1 mutants exhibit indistinguishable phenotypes consistent with a shared role in vivo that are highly reminiscent of human cell lines compromised for BRCA1. Specifically, Cebrc-1 or Cebrd-1 mutants display enhanced p53-dependent germ cell death, radiation sensitivity and chromosome fragmentation, consistent with roles in DNA-damage-repair processes [50]. CeBRC-1 and CeBRD-1 also form foci at sites of DNA damage where they co-localize with Rad51 [51]. The ability of CeBRC-1 and CeBRD-1 to form repair foci is dependent on the C. elegans MRN (Mre11–Rad50–Nbs1) complex, but independent of Rad51 and CeBRC-2 (C. elegans BRCA2) [51]. It is of notable interest that ionizing radiationinduced CeBRC-1/CeBRD-1 foci are restricted to the mitotic compartment of the C. elegans germline, the only region of the adult animal that is actively proliferating. This implies that CeBRC-1 and CeBRD-1 do not simply respond to DSBs, rather they are most likely to be recruited to sites of ionizing radiation-induced replication stress. The distinction between responding to replication stress and to DSBs is supported further by the fact that CeBRC-1 and CeBRD-1 do not form discrete foci at sites of SPO-11 induced meiotic DSBs and are dispensable for meiotic recombination and the generation of crossover products that are evident at diakinesis (results not shown). This behaviour contrasts with CeBRC-2 and Rad51 that form foci at meiotic DSBs, are recruited to ionizing radiation-induced repair foci throughout the germline (rather than being restricted to nuclei in the mitotic region), and are both essential for generating crossover products by recombination [52,53]. Perhaps surprisingly, Cebrc-1 or Cebrd-1 mutants are dispensable for checkpoint-dependent cell-cycle arrest following DNA damage. This contrasts with the failure to induce cell-cycle arrest in atl-1 (C. elegans ATR) mutants, a known component of the S-phase checkpoint, following hydroxyurea or ionizing radiation treatments [54]. The fact that Cebrc-1 or Cebrd-1 is dispensable for checkpoint functions indicates that the sensitivity of Cebrc-1 or Cebrd-1 mutants to DNA-damaging agents is due to DNA-repair defects and not due to a defect in checkpoint signalling. This observation also indicates that the checkpoint function of human BRCA1 has been acquired late on in evolution. Finally, a role for BRCA1 on the X chromosome may also be conserved, as Cebrc-1 or Cebrd-1 mutants exhibit a Him phenotype that arises due to non-disjunction of the X chromosome, indicative of a problem in meiotic chromosome segregation Cellular functions of the BRCA tumour-suppressor proteins [50]. The precise function of CeBRC-1 and CeBRD-1 in meiosis is currently unknown. BRCA1 interaction proteins: true and false It is possible that the diverse functions of BRCA1 manifest through its ability to interact with many different proteins (Figure 1). Indeed, BRCA1 has been reported to interact with tumour-suppressor genes, oncogenes, transcriptional activators and repressors, DNA-damage checkpoint components, cell-cycle regulators and ubiquitylation factors [9,10,25,47]. However, the biological significance of many of the reported interactions are as yet unclear. BARD1 and BRCA1 and their homologues in other species interact to form a heterodimer, co-localize at repair foci, require each other for protein stability and function together as an E3 Ub (ubiquitin) ligase (see below). BARD1- and BRCA1-deficient cells or animal knockouts exhibit strikingly similar phenotypes. BARD1 is essential for BRCA1 function in vivo. UbcH5c associates with a region that encompasses the RING domain of BRCA1 and co-operates with BRCA1– BARD1 in ubiquitylation reactions in vitro [42,55–59]. Depletion of UbcH5c in C. elegans or human cells abolished BRCA1-mediated ubiquitylation in vivo [51]. UbcH5c is therefore of critical importance to the ubiquitylation function of BRCA1. BAP (BRCA1-associated protein) is a C-terminal Ub hydrolase reported to interact with the N-terminal region of BRCA1 and is predicted to function as a de-ubiquitylation enzyme [60]. Although BRCA1 and BARD1 undergo autopolyubiquitylation in vitro, BAP1 does not catalyse their deubiquitylation [58]. Evidence that BAP performs a cellular role that impacts on BRCA1 function has not been forthcoming. The MRN complex is one of the primary sensors of DNA damage in eukaryotic cells and is implicated in DNA repair and checkpoint signalling through its ability to tether broken DNA ends in manner analogous to ‘molecular Velcro’ [61]. The MRN complex is essential for BRCA1 function as it forms a stable association with BRCA1 in response to DNA damage [62], its C. elegans counterpart is required for efficient recruitment of CeBRC-1 and CeBRD-1 to repair foci, and both C. elegans and human MRN complexes are required for BRCA1-mediated ubiquitylation events at sites of DNA damage in vivo [51]. The BRCC complex contains BRCA1, BARD1, BRCA2, Rad51, BRCC36 and BRCC45 [56]. The significance of the interaction between BRCA1, BRCA2 and Rad51 is not known, although these proteins do co-localize at repair foci and similar interactions have been reported for their C. elegans counterparts [20,51,63]. BRCC36 and BRCC45 have been reported to simulate BRCA1–BARD1-mediated ubiquitylation reactions in vitro, but the relevance of these proteins in vivo has not been explored in any great detail [56]. BRIP/FANCJ and CtIP are phosphoproteins that interact with the BRCT domains of BRCA1 [47]. CtIP and BRCA1 are both required for efficient activation of the S-phase checkpoint in response to DNA damage, whereas BRIP/FANCJ are not [62]. The relevance of the association between BRIP/FANCJ and BRCA1 has been called into question by experiments in chicken DT40 cells [64]. It is possible that this interaction is only relevant to BRCA1 function in human cells [46]. Phosphorylation of γ H2AX is one of the earliest events that occurs in response to DNA damage and is important for the recruitment and/or retention of BRCA1 at repair foci [27,65,66]. It is believed that this association is mediated through the binding of phospho-γ H2AX by the BRCT domain in BRCA1. Consistent with this role, cell lines deficient for γ H2AX are compromised for BRCA1-mediated ubiquitylation events at DNA-damage sites [51]. RNA polymerase II is essential for transcription and is reported to interact with BRCA1 and is ubiquitylated by BRCA1–BARD1 in vitro [25,47]. Although, RNA polymerase II is ubiquitylated in vivo, this modification still occurs in BRCA1-deficient cells, and evidence to support an important biological connection between these proteins remains ambiguous. E2F1, pRB, p53, p300 and HDAC1/2 (histone deacetylase 1/2) are reported to interact with BRCA1, but the functional relevance of their association is not known at present [25,47] (Figure 1). BRCA1–BARD1: an E3 Ub ligase The RING domain present in a subset of proteins in the RING domain family confers E3 Ub-ligase activity. Ubiquitylation is the stepwise process by which a target protein is modified by covalent attachment of mono-Ub or polyUb chains [67]. This process occurs in an ATP-dependent manner and is initiated by the formation of a thiol-ester bond between a cysteine residue in the Ub-activating enzyme (E1) and the C-terminal glycine residue of Ub. The second step involves the transfer of Ub from the E1 enzyme to an E3 Ub-conjugating enzyme, with the formation of a new thiolester bond. Finally, an E3 Ub ligase catalyses the transfer of Ub from the E2 to a lysine residue in the target protein. Ubiquitylation is most commonly known for its role in proteolysis. In such cases, proteins modified with Lys48 poly-Ub chains are recognized by the 26 S proteasome and targeted for degradation. However, a role in protein turnover is not the only function of ubiquitylation, as it is now known that Ub chains can be formed by linkage through any one of the seven lysine residues in Ub [68]. For example, Lys63 poly-Ub chains impinge on DNA repair, translation and cellular transport independent of proteolysis. The consequence of ubiquitylation to form Lys6 , Lys11 , Lys27 , Lys29 , Lys33 or Lys63 chains on a given target protein remains to be defined, but such modifications may alter cellular localization, change enzymatic activity, regulate protein–protein interactions or participate in signal transduction. The presence of a RING domain in BRCA1 therefore prompted Lorick et al. [69] to test for an associated E3 Ub-ligase activity. This study revealed that an N-terminal fragment of BRCA1 comprising the first 788 amino acids possesses this activity when a recombinant fragment of BRCA1 fused to glutathione S-transferase was incubated in the presence of E1 Ub-activating enzyme, Escherichia coli lysate containing the human E2 Ub-conjugating enzyme, UbcH5b, and ATP [69]. A subsequent study showed that a minimal region of 78 amino acids including the C 2006 Biochemical Society 637 638 Biochemical Society Transactions (2006) Volume 34, part 5 RING domain was sufficient for E3 Ub-ligase activity [59]. But of physiological importance, it was shown that this activity is abolished by a tumour-derived mutation (C61G) in one of the conserved cysteine residues within the RING domain [59]. The fact that BRCA1 exists as a heterodimer with BARD1 prompted subsequent studies of the ubiquitylation function of BRCA1 in combination with BARD1. Hashizume et al. [57] found that the associated E3 Ub-ligase activity of BRCA1 is dramatically enhanced when fragments of BRCA1 (amino acids 1–304) and BARD1 (amino acids 25–189) proteins were combined in vitro [57]. The enhanced E3 Ub-ligase activity of the BRCA1-BARD1 RING heterodimer is also abolished by the tumour-derived C61G mutation [57]. Collectively these studies demonstrated that the BRCA1–BARD1 heterodimer catalyses ubiquitylation reactions in vitro when combined with E1 and E2 enzymes that is dependent on the structural integrity of the BRCA1 RING domain. In contrast with HECT (homologous with the E6-associated protein C-terminus) domain E3 ligases that form a ubiquitylated intermediate as part of the transfer of Ub to a substrate, RING domain E3 ligases, such as BRCA1–BARD1, are believed to stimulate the transfer of Ub from the E2 directly to the substrate [70,71]. In the initial studies of BRCA1-mediated ubiquitylation, the ubiquitylated species observed in the reaction was most likely to be the autoubiquitylation of BRCA1 and/or BARD1. Indeed, Mallery et al. [58] demonstrated using purified full-length BRCA1– BARD1 proteins that their associated E3 Ub-ligase activity catalyses the formation of multiple poly-Ub chains on itself. This auto-ubiquitylation reaction was found to stimulate the E3 Ub-ligase activity of the heterodimer approx. 20-fold [58]. This suggested one mechanism through which this activity may be regulated, but how auto-ubiquitylation is able to simulate catalysis and whether this activation occurs in vivo remains unclear. It has also been shown that BRCA1–BARD1 auto-ubiquitylation occurs through Lys6 in Ub, suggesting that this enzyme may conjugate Ub through less conventional linkages in vivo [72,73]. However, the Ub linkage(s) conjugated by BRCA1–BARD1 remains somewhat controversial, as a number of independent studies have reached very different conclusions. The majority of studies to determine the lysine residue on Ub that is conjugated by a given E3 enzyme rely on a panel of seven Ub mutants mutated for all but one of the lysine residues able to form Ub chains. It is currently unclear whether all of these seven different Ub mutants fold correctly, so interpretation of data obtained with these mutant proteins needs to be treated with caution. The relevance of BRCA1-mediated ubiquitylation reactions in vivo was supported further by Dong et al. [56] who demonstrated that a holoenzyme complex, termed BRCC, that was purified from H1299 or HEK-293 (human embryonic kidney)-derived cell lines by virtue of a FLAG-tagged BARD1, contained BRCA1, BRCA2, Rad51 and two uncharacterized proteins (BRCC36 and BRCC45) and possessed E3 Ub-ligase activity. Comparison of the recruitment of recombinant BRCA1–BARD1 with that of the purified C 2006 Biochemical Society BRCC complex for E2 Ub-conjugating enzymes revealed a preference for the E2 UbcH5c in vitro [56]. Through the utilization of an antibody that specifically recognizes conjugated Ub, but not the free form of Ub, Morris and Solomon [73] were able to visualize ubiquitylation events in vivo. In this important study, it was shown that conjugated ubiquitylation events are readily detected at stalled replication forks in S-phase cells following treatment with hydroxyurea, an inhibitor of ribonucleotide reductase, and at sites of DSB repair following exposure to ionizing radiation. Strikingly, ubiquitylation events at DNA-damage sites co-localized with BRCA1 repair foci and through the use of siRNA (small interfering RNA) technology, it was shown that depletion of BRCA1 or BARD1 proteins abolished ubiquitylation events at stalled replication forks and DSB repair sites in vivo [73] (Figure 2). Furthermore, ectopic expression of the K6R mutant of Ub also abolished ubiquitylation events at stalled replication forks and DSB repair sites in vivo, consistent with the fact that activation of BRCA1–BARD1 by autopolyubiquitylation requires Lys6 Ub chain conjugation [73]. This study established an assay to monitor BRCA1-mediated ubiquitylation events at repair foci, but how this activity is activated following DNA damage and the identity of the ubiquitylated proteins at repair sites remains unknown. Polanowska et al. [51,74] recently examined the role of C. elegans BRCA1–BARD1 (CeBRC-1 and CeBRD-1; CeBCD) in DNA-repair processes through the biochemical characterization of the CeBCD complex purified from whole C. elegans extracts before and after exposure to ionizing radiation. Analogous to the ubiquitylation activity of the human BRCC complex, the CeBCD complex possessed E3 Ub-ligase activity in vitro and utilizes the E2 Ub-conjugating enzyme Ubc5(LET-70), the C. elegans orthologue of human UbcH5c. The biological relevance to the DNA-damage repair of CeBCD dependent ubiquitylation activity and Ubc5(LET70) was demonstrated by analysing ubiquitylation events induced by damage in different genetic backgrounds in a manner analogous to the study by Morris and Solomon [73]. Consistent with a conserved response in C. elegans and substantiating previous work in human cells, ionizing radiation treatment was found to induce the formation of nuclear conjugated Ub foci specifically in the mitotic compartment of the germline, the only region in the adult animal that is actively proliferating. Importantly, these foci were abolished in Cebrc-1, Cebrd-1 and ubc5(let-70) mutants, indicating that ubiquitylation events at DNA-damage sites are dependent on CeBCD [51]. The latter also provided the first in vivo evidence that UbcH5c homologues, including UBC5(LET70), are the bona fide E2 Ub-conjugating enzymes required for CeBCD-dependent ubiquitylation events. Consistent with these data in C. elegans, siRNA knockdown for three different E2 Ub-conjugating enzymes revealed that UbcH5c, and not UbcH2 or Ube2K, is required for BRCA1-dependent ubiquitylation at DNA-damage sites in human cells. Collectively, these data established UbcH5c homologues as the E2 Ub-conjugating enzymes that function with BRCA1 in vivo [51] (Figure 2). Cellular functions of the BRCA tumour-suppressor proteins Figure 2 Regulation of BRCA1-mediated ubiquitylation at DNA-damage sites Representative images of fixed MCF7 cells before and 1 h after 2 Gy of ionizing radiation, and then immunostained with antibodies specific to conjugated Ub (FK2) and counterstained with DAPI (4 ,6-diamidino-2-phenylindole), are shown on the left. Conjugated (Conj.) Ub foci are induced at DNA-damage sites. The pathway responsible for activating BRCA1-mediated ubiquitylation events is shown on the right. Cells deficient for the MRN complex, ATM, ATR, γ H2AX, BRCA1 and UbcH5c (E2 Ub-conjugating enzyme) are defective for the formation of conjugated Ub foci are induced at DNA-damage sites [51]. Once recruited and activated, BRCA1/BARD1–UbcH5c catalyses ubiquitylation of unknown factors at sites of DNA damage that are predicted to function in DNA-damage signalling and/or repair. Interplay between BRCA1 and the DNA-damage checkpoint Of particular interest was the finding that a stable complex between CeBCD and Ubc5(LET-70) in C. elegans and between BRCA1 and UbcH5c in human cells was detected specifically after DNA damage [51,74]. A role for the DNAdamage checkpoint in promoting this association was inferred from the observation that the association between the respective C. elegans and human proteins, and consequently E3 Ub-ligase activity, was abrogated by treatment with caffeine, an inhibitor of the DNA-damage checkpoint [51]. Moreover, cell lines compromised for the DNA-damage checkpoint, including Nbs1−/− , A-T (ataxia telangiectasia) and ATRSeckel cells (defective for Nbs1, ATM and ATR respectively) were found to be defective for the damage-induced association between BRCA1 and UbcH5c. Consistent with these observations, CeBCD-mediated ubiquitylation at DNAdamage sites in C. elegans was shown to depend on the checkpoint genes mre-11 and atl-1 (C. elegans ATR) and BRCA1mediated ubiquitylation at DNA-damage sites in human cells was shown to require Mre11, Nbs1, ATM, ATR and γ -H2AX. Collectively, these data revealed that CeBCD/ BRCA1-dependent ubiquitylation is a conserved response to DNA damage that is activated by the checkpoint [51,74] (Figure 2). At least one way the DNA-damage checkpoint has an impact on this response to DNA damage is by regulating the association between BRCA1–BARD1 and its specific E2 Ub-conjugating enzyme, UbcH5c. The formation of a stable, damage-specific, interaction between CeBCD/BRCA1 and its E2 enzyme may therefore depend on a protein conformational change induced after damage or through association with an adaptor protein. Indeed, human BRCA1 is phosphorylated directly by ATM and ATR in response to DNA damage [75] and, although the functional consequence of this interaction remains unclear, it is plausible that phosphorylation may stabilize the association between BRCA1 and UbcH5c. Stabilization of the E2–E3 association could also occur following auto-polyubiquitylation of BRCA1–BARD1 [58]. UbcH5c may also be a direct target for checkpointdependent phosphorylation, but this is currently unknown. Checkpoint-dependent phosphorylation could mark substrates for binding and subsequent ubiquitylation, analogous to SCF (Skp1/cullin/F-box) Ub ligases that interact and ubiquitylate their substrates in a phosphorylation-dependent manner [76]. Substrate–E3 interaction could be mediated by the phosphopeptide-binding activity of the BRCT domains that would also bring the substrate into close juxtaposition with the ubiquitylation activity of the RING domain. With a total of four BRCT domains, it is tempting to speculate that the BRCA1–BARD1 heterodimer may bind to more than one phosphorylated protein at a time or have two interfaces for defining substrate binding and specificity. It is also possible that one of the BRCT domains may bind to γ -H2AX at DNA-damage sites [65], while the other domain binds to its substrate(s). Elusive substrates The data of Morris and Solomon [73], and Polanowska et al. [51] strongly suggest that bona fide substrates for CeBCD/ C 2006 Biochemical Society 639 640 Biochemical Society Transactions (2006) Volume 34, part 5 Figure 3 BRCA2: domains, interacting proteins, activities and functions Schematic diagram of the BRCA2 protein depicting the position of the eight BRC motifs (blue), the DNA-binding region [helical domain (yellow)], OB-folds (red) and the single Rad51-binding site (green) at the extreme C-terminus, regulated by Cdk phosphorylation. The indicated BRCA2-interacting proteins are shown under the corresponding region of BRCA1 required for their association. aa, amino acids; NLS, nuclear localization signal. BRCA1-mediated ubiquitylation reside or are recruited to sites of DNA damage and may therefore function in DNA repair, checkpoint signalling and/or regulation of chromatin dynamics [51,73] (Figure 2). Although the BRCA1–BARD1 E3 Ub ligase is able to ubiquitylate FANCD2 (FA complementation group D2), RNA polymerase II, nucleoplasmin, p53, H2AX and the nucleosome core histones H2A, H2B, H3 and H4 in vitro, bona fide targets in vivo remain elusive [56,58]. Indeed, the BRCA1–BARD1 E3 Ub ligase is very promiscuous in vitro in that it is able to ubiquitylate many proteins that are highly unlikely targets in vivo. Therefore the ability of BRCA1–BARD1 to mediate ubiquitylation of a protein in vitro should be treated with caution, particularly as none of the proteins so far shown to be ubiquitylated in vitro by BRCA1–BARD1 have been shown to be substrates in vivo. Analogous to SCF Ub ligases, BRCA1– BARD1 substrates are likely to be phosphorylated and in this modified form may associate with the BRCT domains. Thus substrates may be identified in screens designed to pull out phosphoproteins that bind to the BRCT domains. Alternatively, comparative proteomic approaches to identify all of the proteins that are ubiquitylated on chromatin before and after DNA damage has the potential to identify BRCA1 targets that should be distinguishable from alternative E3 enzyme targets by analysing the ubiquitylation status in BRCA1-defective cells. Until a bona fide substrate is found, it is impossible to predict the effect of BRCA1-mediated ubiquitylation without knowing the type of lysine chain linked to the substrate and mutation of the target lysine C 2006 Biochemical Society residue(s) in the substrate is the only conceivable way that the effect of BRCA1-mediated ubiquitylation will be inferred. In contrast with BRCA1, a wealth of evidence has implicated BRCA2 as an integral component of the HR pathway, where it directly regulates the key recombinase Rad51 [9]. BRCA2 and Rad51 not only co-localize at DNA repair foci, but also interact directly with one another in vitro and by co-immunoprecipitation [18,23,63,77]. BRCA2-deficient cells, such as CAPAN-1, exhibit predominantly cytoplasmic Rad51 and are defective for recruitment of Rad51 to repair foci. These data indicate that BRCA2 is required for nuclear localization of Rad51 and for its efficient recruitment to sites of DSB repair. The domain structure of BRCA2 The human BRCA2 gene encodes a 3418-amino-acid protein that is one of the largest polypeptides in the human proteome (Figure 3). When the BRCA2 gene was cloned in 1995, no obvious domain similarities were identified [7]. However, a repeated motif termed the BRC domain was identified by sequence comparison of the BRCA2 protein to itself [78]. The BRC motif is approx. 70 amino acids in length with a core sequence spanning 26 amino acids and is repeated eight times over a region of 1000 amino acids within the second third of the BRCA2 protein (Figure 3). The sequence of the BRC repeats in BRCA2 are somewhat diverged from one another with BRC1, BRC3, BRC4, BRC7 and BRC8 being most related to one another. Of critical importance to the Cellular functions of the BRCA tumour-suppressor proteins understanding of the role of BRCA2 in DSB repair processes was the finding that the BRC domains mediate direct binding to the Rad51 recombinase [18,79]. Although all of the eight BRC motifs in BRCA2 are able to bind to Rad51, in either yeast two-hybrid or in vitro pull-down assays, they have very different affinities for Rad51, with BRC3 and BRC4 having high affinity, whereas BRC5 and BRC6 have very low affinity [18,79]. It is currently believed that six of the eight mostly highly conserved BRC motifs amongst mammalian species bind to Rad51 in vivo, including BRC1–BRC4, BRC7 and BRC8. Structural insight into the way the BRC motif binds to Rad51 came from Pellegrini et al. [80] who solved the cocrystal structure of the BRC4 motif bound to the RecA homology domain of Rad51. Since Rad51 has a tendency to form large heterogeneous multi-oligomeric aggregates, this study required the generation of a covalent linkage between BRC5 and Rad51 to facilitate purification of the two proteins as a single fusion. A number of hydrophobic residues within a stretch of 28 amino acids within the BRC4 motif ensure close contact with Rad51. Gly1523 and the sequence F1524 HTASGK1530 , which forms a block of highly conserved amino acids, form critical hydrophobic and polar interactions with Rad51. Indeed, cancer-predisposing mutations in BRC4 corresponding to T1526A blocks the ability of BRC4 to bind to Rad51. Similar tumour-derived mutations in equivalent positions in BRC1 (T1011R), BRC2 (S1221P) and BRC7 (T1980I) have been reported. It was proposed that the BRC motif functions as a Velcro strip in the way it binds to Rad51, in that it possesses a large number of contacts that are relatively independent of one another. Comparison of the BRC4–Rad51 structure to the crystallographic RecA filament revealed that BRC4 mimics the interface that is formed between adjacent RecA molecules. That is, BRCA2 interacts with Rad51 by mimicking a structural motif that enables Rad51 to oligomerize [80]. It has been proposed that BRCA2 binding maintains Rad51 in a monomeric state that is primed for deposition on to DNA to form a nucleoprotein filament functional for DNA strand exchange. In addition to tumourderived mutations, the importance of the BRC domain to the role of BRCA2 in HR was illustrated by the finding that in vivo overexpression of the BRC4 motif blocked the formation of Rad51 foci at DNA-damage sites [63]. Yang et al. [81] identified an 800-amino-acid region in the final third of the BRCA2 protein that was predicted to form extensive secondary structure. This region was shown previously to associate with the highly acidic 70-aminoacid polypeptide DSS1 [82] that was originally identified as one of three genes that map to a 1.5 Mb locus deleted in the inherited developmental malformation syndrome, split hand/split foot [83]. Co-expression of BRCA2 C-terminal fragment with DSS1 was found to significantly improve solubility and subsequent protein purification that enabled the generation of co-crystals with and without bound oligo(dT) ssDNA [81]. Structural analysis revealed five distinct domains in the BRCA2 C-terminal fragment (Figure 3). The first domain comprises 190 amino acids consisting of mainly α-helices, termed the helical domain. This is followed by the three structurally related domains of approx. 110 amino acids (OB1, OB2 and OB3) that exhibit homology and structural similarity to the OB (oligonucleotide/oligosaccharide-binding)-fold that is present in most prokaryotic and eukaryotic ssDNA-binding proteins, including SSB (ssDNA-binding protein) and RPA (replication protein A). A 130-amino-acid insertion is also found in OB2, that is absent from other OBfold-containing proteins, which adopts a tower-like structure (tower domain) sticking out from the OB-fold. DSS1 was found to bind to the helical domain, OB1 and OB2 through extensive electrostatic interactions. The acidic nature of DSS1 coupled with its association with the tower, OB1 and OB2, also led to the suggestion that DSS1 might play a role as a DNA mimetic that could regulate the DNA-binding activity of BRCA2. This possibility currently remains unclear. Consistent with the presence of three OBfold domains, the BRCA2 C-terminal fragment was shown to bind with high affinity to ssDNA, whereas the tower domain was found to bind to dsDNA (double-stranded DNA) [81]. This led to the idea that BRCA2 could target Rad51 (via interaction with the BRC motifs) to the dsDNA/ssDNA junction at processed DSBs (through the combined DNAbinding affinities of the tower and OB-folds). In addition to binding to Rad51 via the BRC motifs, a second Rad51-binding site has also been identified at the extreme C-terminus of the protein [84,85] (Figure 3). Using a series of nine overlapping GST (glutathione S-transferase)fusion proteins spanning the entire coding region of BRCA2, Esashi et al. [86] found that the extreme C-terminal GST– BRCA2 fusion protein that is capable of binding Rad51 is also phosphorylated in asynchronous HeLa extracts. Phosphorylation site mapping revealed that Ser3291 is the site modified in BRCA2, the location of Rad51. This site is located in a Cdk (cyclin-dependent kinase) consensus sequence and was subsequently shown to be phosphorylated by Cdk. Strikingly, it was shown that a non-phosphorylated peptide containing the Rad51-binding sites efficiently bound Rad51, whereas phosphorylation at Ser3291 abolished binding. Analysis of the phosphorylation status during the cell cycle revealed that phosphorylation at Ser3291 is low in S-phase when recombination is active and is high as cells enter mitosis. Moreover, under DNA-damaging conditions, phosphorylation at Ser3291 is rapidly removed, thus allowing Rad51 association. The biological relevance of this site is highlighted by the existence of four independent tumour-derived mutations in the Ser3291 -Phe3292 Cdk target sequence. Furthermore, expression of a non-phosphorylated fragment in cells reduced recombination rates. Collectively these results suggested that phosphorylation at Ser3291 could act as a molecular switch to regulate HR by modulating the interaction of BRCA2 with Rad51 [86]. So far no obvious domains or functions have been assigned to the first third of the BRCA2 protein. It is of notable interest that Esashi et al. [86] also found that the first 906 amino acids of BRCA2 contain at least two phosphorylation sites. However, the exact position of these phosphorylation sites and C 2006 Biochemical Society 641 642 Biochemical Society Transactions (2006) Volume 34, part 5 their relevance to the function of BRCA2 remains to be determined. This region is clearly of great interest and will be of considerable focus in the future. BRCA2 homologues in simple eukaryotes The presence of BRC and OB-fold motifs is a hallmark of BRCA2-related proteins and has proved to be a useful method for the identification of BRCA2 orthologues in other species [87]. Surprisingly, the number of BRC motifs in BRCA2-related proteins is highly variable with 15 BRC motifs found in Trypanosoma brucei BRCA2 ranging to a single BRC motif in Ustilago maydis BRCA2 (Brh2) and C. elegans BRCA2 (CeBRC-2) [87]. Kojic et al. [88] were the first to describe a non-mammalian BRCA2. Their analysis of U. maydis Brh2 revealed an essential role in DNA repair, allelic recombination and meiosis analogous to Rad51. Consistent with the presence of a single BRC domain, Brh2 was also found to bind to Rad51 directly [88]. Subsequently, Siaud et al. [89] described Arabidopsis thaliana BRCA2 with four BRC motifs that was essential for meiotic recombination. In the absence of Atbrca2, plants were sterile owing to a failure to repair meiotic DSBs. Indeed, the aberrant recombination events observed in Atbrca2 deficient plants were eliminated by a mutation in spo11, the enzyme responsible for the generation of meiotic DSBs [89]. Martin et al. [53] also described arguably the simplest BRCA2-related protein, which was identified in C. elegans. CeBRC-2 is a 394-amino-acid protein, a little over one-tenth of the size of human BRCA2, which contains a single BRC, a single OB-fold domain and a nuclear localization signal. CeBRC-2 interacts directly with Rad51 through its BRC motif and binds preferentially to ssDNA through its OB-fold domain. It was shown that Cebrc-2 mutants are defective for meiotic and ionizing radiation-induced DSB repair owing to an inability to target Rad51 to the nucleus and a failure to recruit Rad51 to sites of DSBs [53]. Collectively, these studies substantiated the view that BRCA2 and its related proteins function in DSB repair via HR by directly regulating Rad51 functions. BRCA2-interacting proteins In addition to Rad51, a number of other BRCA2-associated proteins have been described (Figure 3). DSS1 was originally identified by Marston et al. [82] in a yeast two-hybrid screen that bound to the region spanning amino acids 2472 and 2957 in BRCA2 (see above for details). The first convincing demonstration that DSS1 is functionally related to BRCA2 came from studies of the DSS1 homologue in U. maydis. UmDss1 was found to bind to Brh2 and mutants in this gene conferred radiation sensitivity, defects in recombination and genome instability [90]. Gudmundsdottir et al. [91] subsequently demonstrated that depletion of DSS1 in human cells compromises DNA-damage-induced Rad51 focus formation and genome maintenance, analogous to the defects observed in the absence of BRCA2. Collectively, these data indicate that DSS1 is a conserved component of the HR pathway that somehow functions with BRCA2 to efficiently target Rad51 C 2006 Biochemical Society to sites of DSBs. FANCD2, a component of the FA DNArepair pathway, also binds directly to BRCA2 in both yeast two-hybrid assays and co-immunoprecipitation studies [92]. Moreover, biallelic inactivation of BRCA2 was found to result in FA and was assigned to FANCD1 (FA complementation group D1) [93]. Current evidence supports a role for FANCD2, FANCD1/BRCA2 and the FA pathway in orchestrating lesion repair via HR and/or translesion bypass pathways. Indeed, a number of studies have reported that FA-deficient cell lines are compromised for HR-mediated DSB repair [94,95]. However, the precise mechanism through which the FA pathway promotes DNA repair remains unclear. EMSY was identified by Hughes-Davis et al. [96] in a yeast two-hybrid screen as a factor that associates with BRCA2 within the region encoded by exon 3. Of particular significance was the finding that EMSY is amplified in breast and ovarian cancers. Consistent with a role in DNA repair, EMSY is recruited to DNA-damage sites; however, its precise role in repair and its relevance to the role of BRCA2 in HR repair processes remains unclear. BRCA2: functional insights from biochemistry Assessing the influence of human BRCA2 on Rad51 activities in vitro has been hampered by its low expression levels and proteolytic degradation that have precluded the purification of full-length BRCA2 protein. Consequently, biochemical studies were initially restricted to the use of fragments. In vitro, the BRC domain alone is capable of inhibiting the formation of the Rad51 nucleoprotein filaments, whereas a C-terminal fragment of BRCA2 containing the three OBfold domains that confer ssDNA binding was shown to stimulate the strand exchange activity of Rad51 [81,97]. It was not clear whether these effects were direct and reflected the true biochemical function of human BRCA2 in vivo. However, a recent study using the purified full-length U. maydis BRCA2 homologue (Brh2) has shown that, when present at substoichiometric amounts relative to Rad51, Brh2 facilitates Rad51 nucleation and filament formation from a dsDNA/ssDNA junction and stimulates Rad51-mediated recombination [98]. Subsequently, San Filippo et al. [99] have reported that a hybrid recombinant BRCA2 fusion protein consisting of BRC3/BRC4 and its DNA-binding domain is able to stimulate Rad51 activities in vitro. A similar effect has also been observed using purified C. elegans proteins (M.I.R. Petalcorin and S.J. Boulton, unpublished work). Collectively, these studies support a conserved role for BRCA2 proteins as a recombination mediator. The current model for BRCA2 function in DSB repair by HR indicates that it is required for efficient nuclear localization of Rad51, it mediates the recruitment of Rad51 to sites of DSBs, it binds to the ssDNA/dsDNA junction at a processed DSB and promotes the nucleation of the Rad51 nucleprotein filament, and, once the Rad51 filament has formed, BRCA2 stimulates Rad51-mediated strand exchange and D-loop formation (Figure 4). Cellular functions of the BRCA tumour-suppressor proteins Figure 4 Model for the action of BRCA2 in DSB repair by HR (1) BRCA2 is required for the recruitment of Rad51 to DSBs. (2) Following nucleolytic resection of the DSB to generate 3 ssDNA overhangs, BRCA2–DSS1 binds to the ssDNA/dsDNA junction and offloads Rad51 on to the ssDNA overhang, thereby facilitating the nucleation of the Rad51 filament. (3) Once the Rad51 nucleoprotein filament has formed, BRCA2 stimulates Rad51-mediated strand exchange and D-loop formation. Conclusions In the last decade, considerable progress has been made in elucidating the cellular functions of BRCA1 and BRCA2. On the basis of a number of recent studies, it is reasonable to suggest that the role of BRCA1–BARD1 in such diverse cellular processes as DNA repair, checkpoint signalling, transcription and meiotic sex chromosome inactivation may be achieved through ubiquitylation of specific target proteins. This is clearly the case for DNA damage. Identification of BRCA1 substrates will be the next major development in this field, and their finding and characterization remains crucial to our full understanding of how BRCA1 contributes to cancer predisposition. On the other hand, the role for BRCA2 in HR processes is well established, although the one area that remains to be explored in detail is how BRCA2 function is modulated in response to DNA damage. Research in my laboratory is supported by Cancer Research UK and grants from Breast Cancer Campaign. References 1 Narod, S.A. (2002) Nat. Rev. Cancer 2, 113–123 2 Narod, S.A. and Foulkes, W.D. (2004) Nat. Rev. Cancer 4, 665–676 3 Turner, N., Tutt, A. and Ashworth, A. (2004) Nat. Rev. Cancer 4, 814–819 4 Hall, J.M., Lee, M.K., Newman, B., Morrow, J.E., Anderson, L.A., Huey, B. and King, M.-C. (1990) Science 250, 1684–1689 5 Miki, Y., Swensen, J., Shattuck-Eidens, D., Futreal, P.A., Harshman, K., Tavtigian, S., Liu, Q., Cochran, C., Bennett, L.M., Ding, W. et al. (1994) Science 266, 66–71 6 Stratton, M.R., Ford, D., Neuhasen, S., Seal, S., Wooster, R., Friedman, L.S., King, M.-C., Egilsson, V., Devilee, P., McManus, R. et al. (1994) Nat. Genet. 7, 103–107 7 Wooster, R., Bignell, G., Lancaster, J., Swift, S., Seal, S., Mangion, J., Collins, N., Gregory, S., Gumbs, C. and Micklem, G. (1995) Nature 378, 789–792 8 Wooster, R., Neuhausen, S.L., Mangion, J., Quirk, Y., Ford, D., Collins, N., Nguyen, K., Seal, S., Tran, T., Averill, D. et al. (1994) Science 265, 2088–2090 9 Jasin, M. (2002) Oncogene 21, 8981–8993 10 Venkitaraman, A.R. (2002) Cell 108, 171–182 11 Moynahan, M.E., Pierce, A.J. and Jasin, M. (2001) Mol. Cell 7, 263–272 12 Patel, K.J., Yu, V.P., Lee, H., Corcoran, A., Thistlethwaite, F.C., Evans, M.J., Colledge, W.H., Friedman, L.S., Ponder, B.A. and Venkitaraman, A.R. (1998) Mol. Cell 1, 347–357 13 Shen, S.X., Weaver, Z., Xu, X., Li, C., Weinstein, M., Chen, L., Guan, X.Y., Ried, T. and Deng, C.X. (1998) Oncogene 17, 3115–3124 14 Xu, X., Weaver, Z., Linke, S.P., Li, C., Gotay, J., Wang, X.W., Harris, C.C., Ried, T. and Deng, C.X. (1999) Mol. Cell 3, 389–395 15 Sharan, S.K., Pyle, A., Coppola, V., Babus, J., Swaminathan, S., Benedict, J., Swing, D., Martin, B.K., Tessarollo, L., Evans, J.P. et al. (2004) Development 131, 131–142 16 Kennedy, R.D. and D’Andrea, A.D. (2005) Genes Dev. 19, 2925–2940 17 Mankouri, H.W. and Hickson, I.D. (2004) Biochem. Soc. Trans. 32, 957–958 18 Chen, P.L., Chen, C.F., Chen, Y., Xiao, J., Sharp, Z.D. and Lee, W.H. (1998) Proc. Natl. Acad. Sci. U.S.A. 95, 5287–5292 19 Scully, R., Chen, J., Ochs, R.L., Keegan, K., Hoekstra, M., Feunteun, J. and Livingston, D.M. (1997) Cell 90, 425–435 C 2006 Biochemical Society 643 644 Biochemical Society Transactions (2006) Volume 34, part 5 20 Scully, R., Chen, J., Plug, A., Xiao, Y., Weaver, D., Feunteun, J., Ashley, T. and Livingston, D.M. (1997) Cell 88, 265–275 21 Jeggo, P.A. (1998) Adv. Genet. 38, 185–218 22 West, S.C., Chappell, C., Hanakahi, L.A., Masson, J.Y., McIlwraith, M.J. and Van Dyck, E. (2000) Cold Spring Harbor Symp. Quant. Biol. 65, 315–321 23 Chen, J., Silver, D.P., Walpita, D., Cantor, S.B., Gazdar, A.F., Tomlinson, G., Couch, F.J., Weber, B.L., Ashley, T., Livingston, D.M. and Scully, R. (1998) Mol. Cell 2, 317–328 24 Moynahan, M.E., Chiu, J.W., Koller, B.H. and Jasin, M. (1999) Mol. Cell 4, 511–518 25 Scully, R. and Livingston, D.M. (2000) Nature 408, 429–432 26 Xu, B., Kim, S. and Kastan, M.B. (2001) Mol. Cell. Biol. 21, 3445–3450 27 Kastan, M.B. and Bartek, J. (2004) Nature 432, 316–323 28 Hartman, A.R. and Ford, J.M. (2002) Nat. Genet. 32, 180–184 29 Anderson, S.F., Schlegel, B.P., Nakajima, T., Wolpin, E.S. and Parvin, J.D. (1998) Nat. Genet. 19, 254–256 30 Monteiro, A.N., August, A. and Hanafusa, H. (1996) Proc. Natl. Acad. Sci. U.S.A. 93, 13595–13599 31 Neish, A.S., Anderson, S.F., Schlegel, B.P., Wei, W. and Parvin, J.D. (1998) Nucleic Acids Res. 26, 847–853 32 Scully, R., Anderson, S.F., Chao, D.M., Wei, W., Ye, L., Young, R.A., Livingston, D.M. and Parvin, J.D. (1997) Proc. Natl. Acad. Sci. U.S.A. 94, 5605–5610 33 Chapman, M.S. and Verma, I.M. (1996) Nature 382, 678–679 34 Ganesan, S., Silver, D.P., Greenberg, R.A., Avni, D., Drapkin, R., Miron, A., Mok, S.C., Randrianarison, V., Brodie, S., Salstrom, J. et al. (2002) Cell 111, 393–405 35 Turner, J.M., Aprelikova, O., Xu, X., Wang, R., Kim, S., Chandramouli, G.V., Barrett, J.C., Burgoyne, P.S. and Deng, C.X. (2004) Curr. Biol. 14, 2135–2142 36 Brzovic, P.S., Rajagopal, P., Hoyt, D.W., King, M.-C. and Klevit, R.E. (2001) Nat. Struct. Biol. 8, 833–837 37 Wu, L.C., Wang, Z.W., Tsan, J.T., Spillman, M.A., Phung, A., Xu, X.L., Yang, M.C., Hwang, L.Y., Bowcock, A.M. and Baer, R. (1996) Nat. Genet. 14, 430–440 38 Deng, C.X. (2002) Environ. Mol. Mutagen 39, 171–177 39 McCarthy, E.E., Celebi, J.T., Baer, R. and Ludwig, T. (2003) Mol. Cell. Biol. 23, 5056–5063 40 Meza, J.E., Brzovic, P.S., King, M.-C. and Klevit, R.E. (1999) J. Biol. Chem. 274, 5659–5665 41 Thai, T.H., Du, F., Tsan, J.T., Jin, Y., Phung, A., Spillman, M.A., Massa, H.F., Muller, C.Y., Ashfaq, R., Mathis, J.M. et al. (1998) Hum. Mol. Genet. 7, 195–202 42 Brzovic, P.S., Keeffe, J.R., Nishikawa, H., Miyamoto, K., Fox, 3rd, D., Fukuda, M., Ohta, T. and Klevit, R. (2003) Proc. Natl. Acad. Sci. U.S.A. 100, 5646–5651 43 Joukov, V., Chen, J., Fox, E.A., Green, J.B. and Livingston, D.M. (2001) Proc. Natl. Acad. Sci. U.S.A. 98, 12078–12083 44 Friedman, L.S., Ostermeyer, E.A., Szabo, C.I., Dowd, P., Lynch, E.D., Rowell, S.E. and King, M.-C. (1994) Nat. Genet. 8, 399–404 45 Williams, R.S., Green, R. and Glover, J.N. (2001) Nat. Struct. Biol. 8, 838–842 46 Cantor, S.B., Bell, D.W., Ganesan, S., Kass, E.M., Drapkin, R., Grossman, S., Wahrer, D.C., Sgroi, D.C., Lane, W.S., Haber, D.A. and Livingston, D.M. (2001) Cell 105, 149–160 47 Deng, C.X. and Brodie, S.G. (2000) BioEssays 22, 728–737 48 Manke, I.A., Lowery, D.M., Nguyen, A. and Yaffe, M.B. (2003) Science 302, 636–639 49 Yu, X., Chini, C.C., He, M., Mer, G. and Chen, J. (2003) Science 302, 639–642 50 Boulton, S.J., Martin, J.S., Polanowska, J., Hill, D.E., Gartner, A. and Vidal, M. (2004) Curr. Biol. 14, 33–39 51 Polanowska, J., Martin, J.S., Garcia-Muse, T., Petalcorin, M.I. and Boulton, S.J. (2006) EMBO J. 25, 2178–2188 52 Alpi, A., Pasierbek, P., Gartner, A. and Loidl, J. (2003) Chromosoma 112, 6–16 53 Martin, J.S., Winkelmann, N., Petalcorin, M.I., McIlwraith, M.J. and Boulton, S.J. (2005) Mol. Cell. Biol. 25, 3127–3139 54 Garcia-Muse, T. and Boulton, S.J. (2005) EMBO J. 24, 4345–4355 55 Brzovic, P.S., Meza, J.E., King, M.-C. and Klevit, R.E. (2001) J. Biol. Chem. 276, 41399–41406 56 Dong, Y., Hakimi, M.A., Chen, X., Kumaraswamy, E., Cooch, N.S., Godwin, A.K. and Shiekhattar, R. (2003) Mol. Cell 12, 1087–1099 57 Hashizume, R., Fukuda, M., Maeda, I., Nishikawa, H., Oyake, D., Yabuki, Y., Ogata, H. and Ohta, T. (2001) J. Biol. Chem. 276, 14537–14540 C 2006 Biochemical Society 58 Mallery, D.L., Vandenberg, C.J. and Hiom, K. (2002) EMBO J. 21, 6755–6762 59 Ruffner, H., Joazeiro, C.A., Hemmati, D., Hunter, T. and Verma, I.M. (2001) Proc. Natl. Acad. Sci. U.S.A. 98, 5134–5139 60 Jensen, D.E., Proctor, M., Marquis, S.T., Gardner, H.P., Ha, S.I., Chodosh, L.A., Ishov, A.M., Tommerup, N., Vissing, H., Sekido, Y. et al. (1998) Oncogene 16, 1097–1112 61 Moreno-Herrero, F., de Jager, M., Dekker, N.H., Kanaar, R., Wyman, C. and Dekker, C. (2005) Nature 437, 440–443 62 Greenberg, R.A., Sobhian, B., Pathania, S., Cantor, S.B., Nakatani, Y. and Livingston, D.M. (2006) Genes Dev. 20, 34–46 63 Chen, C.F., Chen, P.L., Zhong, Q., Sharp, Z.D. and Lee, W.H. (1999) J. Biol. Chem. 274, 32931–32935 64 Bridge, W.L., Vandenberg, C.J., Franklin, R.J. and Hiom, K. (2005) Nat. Genet. 37, 953–957 65 Celeste, A., Fernandez-Capetillo, O., Kruhlak, M.J., Pilch, D.R., Staudt, D.W., Lee, A., Bonner, R.F., Bonner, W.M. and Nussenzweig, A. (2003) Nat. Cell Biol. 5, 675–679 66 Nakamura, T.M., Du, L.L., Redon, C. and Russell, P. (2004) Mol. Cell. Biol. 24, 6215–6230 67 Weissman, A.M. (2001) Nat. Rev. Mol. Cell Biol. 2, 169–178 68 Peng, J., Schwartz, D., Elias, J.E., Thoreen, C.C., Cheng, D., Marsischky, G., Roelofs, J., Finley, D. and Gygi, S.P. (2003) Nat. Biotechnol. 21, 921–926 69 Lorick, K.L., Jensen, J.P., Fang, S., Ong, A.M., Hatakeyama, S. and Weissman, A.M. (1999) Proc. Natl. Acad. Sci. U.S.A. 96, 11364–11369 70 Huibregtse, J.M., Scheffner, M., Beaudenon, S. and Howley, P.M. (1995) Proc. Natl. Acad. Sci. U.S.A. 92, 2563–2567 71 Joazeiro, C.A. and Weissman, A.M. (2000) Cell 102, 549–552 72 Wu-Baer, F., Lagrazon, K., Yuan, W. and Baer, R. (2003) J. Biol. Chem. 278, 34743–34746 73 Morris, J.R. and Solomon, E. (2004) Hum. Mol. Genet. 13, 807–817 74 Polanowska, J., Martin, J.S., Fisher, R., Scopa, T., Rae, I. and Boulton, S.J. (2004) BioTechniques 36, 778–780, 782 75 Tibbetts, R.S., Cortez, D., Brumbaugh, K.M., Scully, R., Livingston, D., Elledge, S.J. and Abraham, R.T. (2000) Genes Dev. 14, 2989–3002 76 Spruck, C.H. and Strohmaier, H.M. (2002) Cell Cycle 1, 250–254 77 Chen, J.J., Silver, D., Cantor, S., Livingston, D.M. and Scully, R. (1999) Cancer Res. 59, 1752s-1756s 78 Bork, P., Blomberg, N. and Nilges, M. (1996) Nat. Genet. 13, 22–23 79 Wong, A.K., Pero, R., Ormonde, P.A., Tavtigian, S.V. and Bartel, P.L. (1997) J. Biol. Chem. 272, 31941–31944 80 Pellegrini, L., Yu, D.S., Lo, T., Anand, S., Lee, M., Blundell, T.L. and Venkitaraman, A.R. (2002) Nature 420, 287–293 81 Yang, H., Jeffrey, P.D., Miller, J., Kinnucan, E., Sun, Y., Thoma, N.H., Zheng, N., Chen, P.L., Lee, W.H. and Pavletich, N.P. (2002) Science 297, 1837–1848 82 Marston, N.J., Richards, W.J., Hughes, D., Bertwistle, D., Marshall, C.J. and Ashworth, A. (1999) Mol. Cell. Biol. 19, 4633–4642 83 Crackower, M.A., Scherer, S.W., Rommens, J.M., Hui, C.C., Poorkaj, P., Soder, S., Cobben, J.M., Hudgins, L., Evans, J.P. and Tsui, L.C. (1996) Hum. Mol. Genet. 5, 571–579 84 Mizuta, R., LaSalle, J.M., Cheng, H.L., Shinohara, A., Ogawa, H., Copeland, N., Jenkins, N.A., Lalande, M. and Alt, F.W. (1997) Proc. Natl. Acad. Sci. U.S.A. 94, 6927–6932 85 Sharan, S.K., Morimatsu, M., Albrecht, U., Lim, D.S., Regel, E., Dinh, C., Sands, A., Eichele, G., Hasty, P. and Bradley, A. (1997) Nature 386, 804–810 86 Esashi, F., Christ, N., Gannon, J., Liu, Y., Hunt, T., Jasin, M. and West, S.C. (2005) Nature 434, 598–604 87 Lo, T., Pellegrini, L., Venkitaraman, A.R. and Blundell, T.L. (2003) DNA Repair 2, 1015–1028 88 Kojic, M., Kostrub, C.F., Buchman, A.R. and Holloman, W.K. (2002) Mol. Cell 10, 683–691 89 Siaud, N., Dray, E., Gy, I., Gerard, E., Takvorian, N. and Doutriaux, M.P. (2004) EMBO J. 23, 1392–1401 90 Kojic, M., Yang, H., Kostrub, C.F., Pavletich, N.P. and Holloman, W.K. (2003) Mol. Cell 12, 1043–1049 91 Gudmundsdottir, K., Lord, C.J., Witt, E., Tutt, A.N. and Ashworth, A. (2004) EMBO Rep. 5, 989–993 92 Hussain, S., Wilson, J.B., Medhurst, A.L., Hejna, J., Witt, E., Ananth, S., Davies, A., Masson, J.Y., Moses, R., West, S.C. et al. (2004) Hum. Mol. Genet. 13, 1241–1248 93 Howlett, N.G., Taniguchi, T., Olson, S., Cox, B., Waisfisz, Q., De Die-Smulders, C., Persky, N., Grompe, M., Joenje, H., Pals, G. et al. (2002) Science 297, 606–609 Cellular functions of the BRCA tumour-suppressor proteins 94 Nakanishi, K., Yang, Y.G., Pierce, A.J., Taniguchi, T., Digweed, M., D’Andrea, A.D., Wang, Z.Q. and Jasin, M. (2005) Proc. Natl. Acad. Sci. U.S.A. 102, 1110–1115 95 Yamamoto, K., Ishiai, M., Matsushita, N., Arakawa, H., Lamerdin, J.E., Buerstedde, J.M., Tanimoto, M., Harada, M., Thompson, L.H. and Takata, M. (2003) Mol. Cell. Biol. 23, 5421–5430 96 Hughes-Davies, L., Huntsman, D., Ruas, M., Fuks, F., Bye, J., Chin, S.F., Milner, J., Brown, L.A., Hsu, F., Gilks, B. et al. (2003) Cell 115, 523–535 97 Davies, A.A., Masson, J.Y., McIlwraith, M.J., Stasiak, A.Z., Stasiak, A., Venkitaraman, A.R. and West, S.C. (2001) Mol. Cell 7, 273–282 98 Yang, H., Li, Q., Fan, J., Holloman, W.K. and Pavletich, N.P. (2005) Nature 433, 653–657 99 San Filippo, J., Chi, P., Sehorn, M.G., Etchin, J., Krejci, L. and Sung, P. (2006) J. Biol. Chem. 281, 11649–11657 Received 6 June 2006 C 2006 Biochemical Society 645