Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
No-SCAR (Scarless Cas9 Assisted Recombineering) Genome Editing wikipedia , lookup
History of genetic engineering wikipedia , lookup
Genetic drift wikipedia , lookup
Oncogenomics wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
Genome evolution wikipedia , lookup
Point mutation wikipedia , lookup
Population genetics wikipedia , lookup
The Evolution of Sex
Many groups of organisms, notably the majority of animals and plants, reproduce
sexually. Why?
Dandelion
Taraxacum officinale
Bdelloid rotifer
Two-fold Cost of Sex
f=2
Sexual Population
Asexual Population
Population with two sexes, only one
of which is capable of bearing young
Population with one sex capable
of bearing young
t
n/(2N+n)
N
kS
t+1
n
kS
N
kS
½
½
1
1
n/(N+n)
This cost arises because if females contribute all resources to gametes, asexual females
can produce the same number of offspring as sexual females, but avoid ‘diluting’ their
genome with paternal genetic material when producing offspring. Thus, in the absence
of strong selection for sex through recombination, a clonally reproducing mutant is
expected to spread rapidly in a sexual population of males and females.
A Molecular Machinery Enabling Recombination
Sex and recombination through meiosis are confined to eukaryotes
However, a complex molecular machinery enabling homologous recombination
between different DNA molecules was already present in prokaryotes long
before the first eukaryotes evolved. The original function of this machinery lies
in DNA repair
Three types of DNA damages are repaired in bacteria through mechanisms
that involve homologous recombination:
• Double strand breaks
• Stalled replication forks
• Single stranded DNA that arose from incomplete replication
A Molecular Machinery Enabling Recombination
Recombinatorial repair of double strand breaks is the most relevant mechanism
with respect to the evolution of sex in eukaryotes, where double strand breaks
are induced during Meiosis I to initiate crossovers
A Molecular Machinery Enabling Recombination
Even though the main function of the recombination machinery in bacteria is to
repair DNA damages, it should be stressed that already in bacteria this
machinery is sometimes employed to effect sexual processes
Three such mechanisms of recombination have been identified:
• Conjugation: the exchange of plasmids
• Transduction: the transfer of DNA mediated by phages
• Transformation: the uptake and integration of free DNA from the
environment
Meiosis and the Alternation of Generations
Most eukaryotes are characterized by a life-cycle termed the alternation of
generations, in which a diploid phase of cell divisions alternates with a
haploid phase
Meiosis mediates the transition from a diploid generation to a haploid
generation
Meiosis:
• Reduces the genomic content of cells by one half
• Produces haploid cells with unique combinations of genes.
Syngamy mediates the transition from a haploid generation back to a diploid
generation
Meiosis and the Alternation of Generations
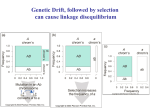
From a purely population genetics perspective, all that recombination does is
to reduce statistical associations –linkage disequilibrium– between alleles at
different loci
Thus, the problem of why recombination is so prevalent in natural
populations boils down to the questions of what forces generate LD and
under what conditions there is selection to destroy LD.
Meiosis and the Alternation of Generations
Many hypotheses have been proposed to account for the advantage of
recombination:
• The Deterministic Mutational Theory
• The Hill-Robertson effect
• Red Queen Theory
Meiosis and the Alternation of Generations
The Deterministic Mutational Theory
• The majority of non-neutral mutations are deleterious
• If a mutation has a deleterious effect, it will then usually be removed from
the population by the process of natural selection
• Sexual reproduction is believed to be more efficient than asexual
reproduction in removing those mutations from the genome
There are two main hypotheses which explain how sex may act to remove
deleterious genes from the genome.
1.
Maintenance of mutation-free individuals
2.
Removal of deleterious genes
Meiosis and the Alternation of Generations
The Deterministic Mutational Theory
1. Maintenance of Mutation-free Individuals
In a finite asexual population under the pressure of deleterious mutations, the
random loss of individuals without such mutations is inevitable. In a sexual
population, however, mutation-free genotypes can be restored by
recombination of genotypes containing deleterious mutations.
Meiosis and the Alternation of Generations
The Deterministic Mutational Theory
2. Removal of Deleterious Genes
It assumes that the majority of deleterious mutations are only slightly
deleterious, and affect the individual such that the introduction of each
additional mutation has an increasingly large effect on the fitness of the
organism. This relationship between number of mutations and fitness is
known as synergistic epistasis
An organism may be able to cope with a few defects, but the presence of
many mutations could overwhelm its backup mechanisms.
The slightly deleterious nature of mutations means that the population will
tend to be composed of individuals with a small number of mutations. Sex
will act to recombine these genotypes, creating some individuals with fewer
deleterious mutations, and some with more. Because there is a major
selective disadvantage to individuals with more mutations, these individuals
die out
Meiosis and the Alternation of Generations
The Deterministic Mutational Theory
There has been much criticism of this theory, since it relies on two key restrictive
conditions:
1. The rate of deleterious mutation should exceed one per genome per generation in
order to provide a substantial advantage for sex.
While there is some empirical evidence for it, there is also strong evidence against it
2. There should be strong interactions among loci (synergistic epistasis), a mutationfitness relation for which there is only limited evidence
Meiosis and the Alternation of Generations
The Hill-Robertson Effect
If two advantageous alleles A and B occur at random. The two alleles are recombined
rapidly in a sexual population but in an asexual population the two alleles must arise
independently
Sex could be a method by which novel genotypes are created. Since sex combines
genes from two individuals, sexually reproducing populations can combine
advantageous genes while asexual populations can’t
Meiosis and the Alternation of Generations
The Hill-Robertson Effect
If, in a sexual population, two different advantageous alleles arise at different loci on a
chromosome in different members of the population, a chromosome containing the
two advantageous alleles can be produced within a few generations by recombination.
However, should the same two alleles arise in different members of an asexual
population, the only way that one chromosome can develop the other allele is to
independently gain the same mutation, which would take much longer
Meiosis and the Alternation of Generations
The Hill-Robertson Effect
But these explanations depend upon the rate of mutation. If favorable mutations are
so rare that each will become fixed in the population before the next arises, then
sexual and asexual populations would evolve at the same rate
Additionally, these explanations depend upon group selection, which most theorists
maintain is a weak selective force relative to individual selection – sex is still
disadvantageous to the individual due to the twofold cost of sex
Meiosis and the Alternation of Generations
The Red-Queen Theory
One of the most widely accepted theories to explain the persistence of sex is the Red
Queen Hypothesis which argues that sex is maintained to assist sexual individuals in
resisting parasites
When an environment changes, previously neutral or deleterious alleles can become
favorable. If the environment changed sufficiently rapidly, these changes in the
environment can make sex advantageous for the individual. Such rapid changes in
environment are caused by the co-evolution between hosts and parasites
Imagine, for example that there is one gene in parasites with two alleles p and P
conferring two types of parasitic ability, and one gene in hosts with two alleles h and H,
conferring two types of parasite resistance, such that parasites with allele p can attach
themselves to hosts with the allele h, and P to H. Such a situation will lead to cyclic
changes in allele frequency -as p increases in frequency, h will be disfavored
In reality, there will be several genes involved in the relationship between hosts and
parasites. In an asexual population of hosts, offspring will only have the different
parasitic resistance if a mutation arises. In a sexual population of hosts, however,
offspring will have a new combination of parasitic resistance alleles
Meiosis and the Alternation of Generations
The Red-Queen Theory
In other words, like Lewis Carroll's Red Queen, sexual hosts are continually adapting in
order to stay ahead of their parasites
Evidence for this explanation for the evolution of sex is provided by comparison of the
rate of molecular evolution of genes for kinases and immunoglobulins in the immune
system with genes coding other proteins. The genes coding for immune system proteins
evolve considerably faster
Critics of the Red Queen hypothesis question whether the constantly-changing
environment of hosts and parasites is sufficiently common to explain the evolution of sex
Mating Types
Mating types are the different types of gametes that can fertilize other
gametes in a sexually reproducing organism. Most species show two different
mating types (male and female, + and -, a and ), but some species of fungi can
present several thousands
Mating Types
Mating types might be the outcome of selection to:
• Facilitate finding mates
In order for gametes to fertilize other gametes, they need to attract
and/or be attracted by other gametes. Evolutionary models show that
there may be selection for some cells to specialize in attracting gametes
(by producing pheromones), while others specialize in becoming
attracted (by expressing pheromone receptors)
• Coordinate the inheritance of cytoplasmic genomes (for example
mitochondrial genes) so as to limit competition between unrelated
cytoplasmic genomes
Fusion of isogamous gametes brings together cytoplasmic genes from
different lineages that may compete to favor their own transmission to
the next generation. Intragenomic conflict reduces the fitness of the
organism and creates the context for the invasion of a nuclear gene that
enforces the inheritance of cytoplasm from a single mother cell
Anisogamy and Mobile Gametes
Anisogamy refers to the production of gametes that differ (generally in size) as
opposed to them being identical (isogamy)
One way to explain the evolution of this asymmetry is assuming that there is a
trade-off between productivity (more gametes are better than few gametes)
and survival of zygotes (bigger zygotes have greater survival than smaller)
This creates the context for the evolution of sexual antagonism with one of
the sexes acting as a cheater that withholds resources to produce more
gametes and the other sex contributing the resources withheld by the first
gamete to preserve zygotic viability. This sexual conflict results in males
producing small gametes that are viable only because of the resources
contributed by females
Anisogamy and Mobile Gametes
The previous model however does not take into consideration how gamete
density affects the probability of fertilization. When all gametes do not find a
partner to fuse with and when small gametes have a higher motility – thus
increasing encountering rates with large gametes – there can be additional
selective pressure for anisogamy. Thus anisogamy does not necessarily
respond to the logic of sexual conflict but might be beneficial for both sexes
Secondary Loss of Sex
Most multicellular organisms —especially animals—have a genetic system
that involves obligate sex as well as male and female
Because of the twofold cost of sex, this near ubiquity of sex is even more
difficult to explain than explaining how sex and recombination evolved in the
first place. Some species, however, have re-evolved the ability to reproduce
asexually either partially or completely, thus offering important opportunities
to investigate the evolutionary forces that maintain sex within populations
and to test hypotheses for the advantage of recombination
Secondary Loss of Sex
Partial loss of sexual reproduction is relatively common in multicellular
organisms and characteristic of several large taxa. Some groups of animals –
for example aphids, waterfleas and monogonont rotifers – reproduce mainly
asexually, but under certain conditions males and females are produced and
mate (facultative sex or cyclic parthenogenesis). In other groups, sexual and
obligatorily asexual individuals coexist, although often with different
geographical distributions ('geographical parthenogenesis').
Secondary Loss of Sex
By contrast, complete abandonment of sexual reproduction is rare among
multicellular organisms. For example, there are fewer than 100
parthenogenetic vertebrate species. The genetic basis for parthenogenetic
reproduction varies among groups and includes single mutations,
hybridization (possibly the only cause of parthenogenesis in vertebrates), and
maternally inherited bacteria. Moreover, there is a great diversity in
cytogenetic mechanisms by which offspring are produced asexually