Survey
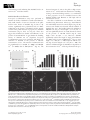
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Tissue engineering wikipedia , lookup
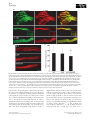
Cell growth wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cell culture wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Signal transduction wikipedia , lookup
Extracellular matrix wikipedia , lookup
List of types of proteins wikipedia , lookup
New Phytologist Research Arabidopsis VILLIN4 is involved in root hair growth through regulating actin organization in a Ca2+dependent manner Yi Zhang*, Yingyu Xiao*, Fei Du, Lijuan Cao, Huaijian Dong and Haiyun Ren Key Laboratory of Cell Proliferation and Regulation Biology of Ministry of Education and College of Life Science, Beijing Normal University, Beijing 100875, China Summary Author for correspondence: Haiyun Ren Tel: +86 10 58806090 Email: [email protected] Received: 18 October 2010 Accepted: 12 December 2010 New Phytologist (2011) 190: 667–682 doi: 10.1111/j.1469-8137.2010.03632.x Key words: actin-binding protein, actin bundle, actin cytoskeleton, cytoplasmic streaming, villin. • Villin is one of the major actin filament bundling proteins in plants. The function of Arabidopsis VILLINs (AtVLNs) is still poorly understood in living cells. In this report, the biochemical activity and cellular function of AtVLN4 were examined. • The biochemical property of AtVLN4 was characterized by co-sedimentation assays, fluorescence microscopy and spectroscopy of pyrene fluorescence. The in vivo function of AtVLN4 was analysed by ectopically expressing it in tobacco pollen and examining the phenotypes of its T-DNA insertional plants. • Recombinant AtVLN4 protein exhibited multiple activities on actin, including actin filament bundling, calcium (Ca2+)-dependent filament severing and barbed end capping. Expression of AtVLN4 in tobacco pollen induced the formation of supernumerary actin cables and reduced pollen tube growth. Loss of function of AtVLN4 resulted in slowing of root hair growth, alteration in cytoplasmic streaming routes and rate, and reduction of both axial and apical actin bundles. • Our results demonstrated that AtVLN4 is involved in root hair growth through regulating actin organization in a Ca2+-dependent manner. Introduction The actin cytoskeleton is a highly organized and dynamic component of eukaryotic cells. It has crucial functions in many physiological processes, including cell growth, cell morphology and motility. Its dynamics and organization are precisely controlled spatially and temporally by numerous actin-binding proteins (ABPs) (Dos Remedios et al., 2003). In plant cells, actin filaments are often organized into higher-order structures, such as bundles and cables, which serve as tracks for cytoplasmic streaming and intracellular transport of organelles and vesicles (Shimmen et al., 1995; Ye et al., 2009). Actin filament cross-linking and bundling proteins are thought to be responsible for actin bundle formation (Thomas et al., 2009). Villin is one of the major proteins responsible for organizing actin filaments into bundles (Dos Remedios et al., 2003; Thomas et al., 2009). It belongs to a multifunctional superfamily of actin-binding proteins called the villin ⁄ *These authors contributed equally to this work. 2011 The Authors New Phytologist 2011 New Phytologist Trust gelsolin ⁄ fragmin family (Friederich et al., 1999; Su et al., 2007). This superfamily is a group of proteins sharing three or six tandem 125–150-aa gelsolin homology domains, designated G1–G6. Gelsolin, the founding member of the family, is composed of six gelsolin homology domains that have distinct properties, endowing this protein with various activities, including severing, capping and nucleating actin filaments (Vandekerckhove, 1990; Burtnick et al., 1997; Su et al., 2007). In addition to the gelsolin-like core domain, typical villin contains an additional extension at its C-terminus, termed the headpiece (VHP), which allows each molecule of villin to arrange actin filaments into bundles (Friederich et al., 1990; Hartwig & Kwiatkowski, 1991). Most, but not all, villins sever actin filaments and cap their barbed ends at micromolar calcium (Ca2+) concentrations (Hesterberg & Weber, 1983; Northrop et al., 1986; Janmey & Matsudaira, 1988; Otto, 1994; Ferrary et al., 1999). The first two villin homologs identified in plants were P-135-ABP and P-115-ABP, which were isolated from lily (Lilium longiflorum) pollen tubes by biochemical fractionation (Nakayasu et al., 1998; Yokota et al., 1998). Both of New Phytologist (2011) 190: 667–682 667 www.newphytologist.com New Phytologist 668 Research them are able to generate actin bundles with uniform polarity in a Ca2+ ⁄ calmodulin (CaM)-dependent manner in vitro (Yokota et al., 1998, 2000, 2003; Yokota & Shimmen, 1999). Both P-135-ABP and P-115-ABP colocalize with actin filament bundles in the transvacuolar strands and the subcortical regions within the root hair cells of Hydrocharis dubia (Tominaga et al., 2000; Yokota et al., 2003). Microinjection of the antiserum against P-115-ABP or P-135-ABP into living root hair cells disintegrates actin filament bundles, destroys transvacuolar strands and alters cytoplasmic streaming routes, thus demonstrating that these villin homologs are involved in maintenance of actin bundles in root hairs (Tominaga et al., 2000; Yokota et al., 2003). Moreover, biochemical experiments have shown that P-135-ABP can form a complex with G-actin and accelerate the polymerization and depolymerization of actin filaments in the presence of Ca2+ ⁄ CaM (Yokota et al., 2005). The Arabidopsis genome contains five villin-like genes (AtVLN1-5) that are abundantly expressed in a wide range of tissues, with elevated expression levels in certain types of cells (Klahre et al., 2000). The full length of AtVLN3 and the headpiece domains of AtVLN1 to AtVLN3, when fused with green fluorescent protein (GFP), decorate actin filaments in plant and animal cells, thus demonstrating that these AtVLNs bind to actin filaments in vivo (Klahre et al., 2000). The biochemical activities and cellular functions of several AtVLNs have recently been studied. Recombinant AtVLN1 binds with high affinity to F-actins, bundles actin filaments in a Ca2+- and CaM-insensitive manner, and protects actin filaments from ADF-mediated depolymerization, but does not sever, cap or nucleate actin filaments (Huang et al., 2005). AtVLN3 has overlapping and distinct activities with AtVLN1 (Khurana et al., 2010). It not only bundles actin filaments in a Ca2+-independent manner, but also severs actin filaments and bundles when Ca2+ is elevated to micromolar levels (Khurana et al., 2010). Loss of function of AtVLN5 sensitizes actin filaments in pollen grains and tubes to latrunculin B, demonstrating that it is a regulator of actin filament stability in pollen (Zhang et al., 2010). As both AtVLN3 and AtVLN5 can sever actin filaments at physiological Ca2+ concentrations in vitro, they have been proposed to be involved in regulating actin filament turnover in response to alteration of Ca2+ levels in plant cells, such as in pollen tubes (Khurana et al., 2010; Staiger et al., 2010; Zhang et al., 2010). However, more cytological evidence is still needed to support this hypothesis. We examined the biochemical properties of a villin isovariant, AtVLN4, and investigated its function within root hairs of Arabidopsis. Our results demonstrate that AtVLN4 maintains all the typical activities of villin family members, regulates the organization of long axial and short apical actin bundles in root hairs, and is essential for normal root hair growth and cytoplasmic streaming within root hairs. New Phytologist (2011) 190: 667–682 www.newphytologist.com Materials and Methods Plant materials and growth conditions Arabidopsis thaliana (L.) Heynh. ecotype Columbia seeds were surface sterilized and grown vertically on Murashige and Skoog (MS) agar plates supplemented with 3.0% sucrose in a controlled growth room with 16 h light : 8 h dark cycles at 22C ± 2C. Sequence alignment and phylogenetic analysis Alignments were performed with CLUSTALX version 1.83 (http://bips.u-strasbg.fr/fr/Documentation/ClustalX/) using default settings, and phylogenetic trees were constructed using the neighbor-joining algorithm of MEGA version 4 (Tamura et al., 2007). A bootstrap test of the phylogeny was performed with 1000 replications. Plasmid construction and protein production A full-length cDNA clone (U60239) for AtVLN4 was kindly donated by the ABRC (Ohio State University, Columbus, OH, USA). The coding sequence for AtVLN4 was subcloned into pBI121 under the control of a LAT52 promoter or in frame with 6· His into pET-30a(+) (Novagen, Madison, WI, USA), and confirmed by sequencing. The error-free AtVLN4-pET-30a(+) construct was overexpressed in Escherichia coli strain BL21 (DE3). Cells were grown to an OD600 of 0.6 at 37C in Luria–Bertani medium and induced with 0.5 mM isopropylthio-b-galactoside at 22C for 12 h. Cultures were collected by centrifugation and resuspended in binding buffer (400 mM NaCl, 40 mM phosphate buffer saline, pH 8.0) supplemented with a 1 : 200 dilution of a stock solution of protease inhibitors (Ren et al., 1997). This was followed by purification using a Ni-NTA His Bind Resin following the protocol in the manufacturer’s manual (Novagen). The purified proteins were dialysed overnight against buffer G (2 mM TrisHCl, 200 lM CaCl2, 0.5 mM DTT, 200 lM ATP, pH 7.5) and stored in aliquots in liquid nitrogen. Protein concentrations were determined with the Bradford reagent (Bio-Rad), using BSA as a standard. Actin was isolated from rabbit skeletal muscle acetone powder using the method described by Pardee & Spudich (1982) and labeled on Cys-374 with pyrene iodoacetamide (Pollard, 1983) and Oregon Green 488 iodoacetamide (Kuhn & Pollard, 2005). Co-sedimentation assays High-speed co-sedimentation assays were used to determine the F-actin binding and depolymerizing activity of AtVLN4. All proteins were preclarified at 200 000 g for 1 h before use. Actin was polymerized in the presence of 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist 1 · F buffer (buffer G with the addition of 50 mM KCl, 2.5 mM MgCl2 and 0.1 mM ATP) for 16 h at 4C. AtVLN4 was incubated with 3.0 lM preformed F-actin in the presence of various concentration of Ca2+ for 1 h at room temperature (RT). To determine whether Ca2+ affects the binding ability of AtVLN4 to actin filaments, AtVLN4 was incubated with phalloidin-stabilized-F-actin (3.0 lM) in the presence of various concentrations of Ca2+. Free Ca2+ concentrations in the presence of ethylene glycol-bis(betaaminoethyl ether)-N,N,N ¢,N ¢-tetraacetic acid (EGTA) were calculated using MAXC programs (available at http:// www.stanford.edu/~cpatton/maxc.html). According to our calculations, 2.0 mM EGTA was equivalent to the volume of 0.04 lM free Ca2+, 0.4 mM EGTA was equal to 0.5 lM Ca2+, 0.2 mM EGTA was equal to 5 lM Ca2+ and 0.15 mM EGTA was equal to 50 lM Ca2+ in all of our experimental systems. After centrifugation at 200 000 g for 1 h in a TLA-110 rotor (Beckman, Brea, CA, USA) at 4C, equal amounts of supernatants and pellets were subjected to 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and stained with Coomassie Brilliant Blue R 250 (DingGuo, Beijing, China). To determine Kd, increasing amounts of AtVLN4 (0, 0.2, 0.4, 0.8, 1.6, 2.4 and 3.2 lM) were incubated with 5.0 lM preformed F-actin in the presence of 2.0 mM EGTA for 1 h at RT. Samples were analysed as described earlier, and the amount of AtVLN4 in the pellets or supernatants was quantified using QUANTITY ONE v4.6.5 software (Bio-Rad). The Kd value for AtVLN4 bound to F-actin was calculated by fitting the data of bound protein vs free protein to a hyperbolic function using PRISM 5 software (GraphPad Software, Inc., San Diego, CA, USA). Low-speed co-sedimentation assays were used to determine the actin bundling activity of AtVLN4. F-actin (3.0 lM) was incubated with AtVLN4 in the presence of various concentration of Ca2+ for 30 min at RT. Samples were centrifuged at 13 500 g for 30 min at 4C, analysed by SDSPAGE and the percentage of actin in each pellet was calculated. The experiments described earlier and the functional assays described later were all repeated independently at least three times using separate new preparations of each sample. Actin filament depolymerization assays Preassembled F-actin (3.0 lM, 50% pyrene-labeled) was incubated with AtVLN4 in the presence of various concentration of Ca2+ for 5 min at RT and the sample was diluted 10-fold with 1· F buffer. The change of pyrene fluorescence intensity accompanying the F-actin depolymerization was monitored for 600 s after dilution. Research for 5 min at RT. Then, 1.0 lM G-actin saturated with 4.0 lM human profilin I and one-tenth volume of 10· F buffer were added to initiate actin polymerization at the barbed end of actin filaments. The increase of pyrene fluorescence was monitored after the actin elongation was initiated. Fluorescence microscopy visualization of actin filaments and bundles F-actin (3.0 lM) was incubated with 0.4 lM AtVLN4 in the presence of 200 lM Ca2+ or 2.0 mM EGTA for 30 min at RT and labeled with equimolar amount of Alexa 488-phalloidin (Molecular Probes, Eugene, Oregon, USA). F-actin was diluted to 50 nM with 1· F buffer and observed using a microscope (Carl Zeiss 200M, Germany) equipped with a 100 · ⁄ 1.5-numerical aperture Planapo objective. Digital images were collected with an Axio CamMR charge-coupled device camera using AxioVision software (Carl Zeiss). Fluorescence microscopy to determine the severing activity of AtVLN4 Oregon Green labeled actin (75% labeled) was polymerized in the presence of 1· KMEI buffer (50 mM KCl, 1 mM MgCl2, 1 mM EGTA and 10 mM imidazole, pH 7.0) at RT for at least 1 h. Glass flow cells, with a capacity of c. 10 ll, were prepared each day as described in Kuhn & Pollard (2005). Flow cells were blocked with two volumes of 1% BSA for at least 2 min, and washed with a fluorescence buffer (10 mM imidazole pH 7.0, 50 mM KCl, 1 mM MgCl2, 100 mM dithiothreitol (DTT), 0.2 mM ATP, 15 mM glucose, 20 lg ml)1 catalase, 100 lg ml)1 glucose oxidase, 0.5% methylcellulose). Actin filaments (200 lM) in the fluorescence buffer were introduced into the chambers and allowed to settle for 5 min. Fluorescence buffer alone, or a mixture of AtVLN4 at different concentrations of Ca2+, was perfused into the flow cells. Actin filaments were observed under an Observer Z1 microscope (Carl Zeiss) equipped with an alphaPlanApo 100 · ⁄ 1.46-numerical aperture oil objective. The time-course of actin filament severing was photographed at 2 s or 5 s intervals using an AxioCam Hsm camera (Carl Zeiss) and AxioVision software. Exposure time was 900 ms. To quantify the severing activity of AtVLN4, ‡ 20 filaments with lengths > 10 lm were chosen, and the average severing frequency was calculated from the numbers of breaks, per unit of filament length, per unit time (i.e. breaks lm)1 s)1) (Andrianantoandro & Pollard, 2006; Khurana et al., 2010; Zhang et al., 2010). Seeded elongation assays Particle bombardment-mediated transient expression in tobacco pollen Freshly prepared F-actin seeds (0.4 lM) were mixed with AtVLN4 in the presence of various concentrations of Ca2+ Particle bombardment-mediated transient expression of AtVLN4 in tobacco pollen was performed as described by 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist (2011) 190: 667–682 www.newphytologist.com 669 670 Research Fu et al. (2001). Bombarded pollen grains were washed into Petri dishes with 1.5 ml of germination medium (5 lM CaCl2, 5 lM Ca(NO3)2, 1 mM MgSO4, 0.01% H3BO3, 15% sucrose, pH 6.5) and shaken at 80 rpm and 28C. After incubation for 3 h, the pollen tubes were photographed to measure tube length. The untransformed pollen tubes were used as a wild-type control. Reverse-transcription polymerase chain reaction (RT-PCR) analysis Total RNA was isolated from 9-d-old Arabidopsis seedlings using the plant RNA extraction kit according to the manufacturer’s instructions (AutoLab Biotechnology Co. Ltd., Beijing, China). Two micrograms of total RNA was added to M-MLV reverse transcription kit (Promega Co., Madison, WI, USA) with Oligo-dT primers (Takara Biotechnology (Dalian) Co., Ltd., Dalian, China) for the synthesis of cDNA. Four microliters of the reaction product was used as templates to amplify a 2131 bp fragment of AtVLN4 in order to detect the expression level of AtVLN4 in wild type and mutants. Primers used were: 5¢-GCTATGTTTTCCAGTATTCTTATCCCG-3¢ (4-1f) and 5¢-AAAAACTCAATCCTCAACTTCTGCAA-3¢ (4-2r). ACTIN2 ⁄ 8 (forward primer 5¢-GGTAACATTGTGCTCAGTGGTGG-3¢ and reverse primer 5¢-AACGACCTTAATCTTCATGCTGC-3¢) genes were used as an internal control. To detect the upstream product, a 176 bp fragment of AtVLN4 was amplified using the following primers: 5¢-AAGAATGAAGAAATAAACAGTTGATGGG-3¢ (v4uf) and 5¢-AGTGAAAGAGAGAGAATTGAATTGAGAG-3¢ (v4ur). To further determine the expression level of AtVLN4 in wild-type and mutant plants, quantitative real-time RTPCR was performed to amplify a 156 bp fragment using the 7500 Real-Time PCR System (ABI, Carlsbad, CA, USA) according to the manufacturer’s instructions. The reactions were performed in a 20-ll volume containing Power SYBR Green PCR Master Mix (ABI). Primers used were: 5¢-GATTGTTTCTTCTTCAGGTCACAGGTT-3¢ (4-2f) and 4-2r. The level of eIF4A transcript (forward primer 5¢-GGGTATCTATGCTTACGGTTTCG-3¢ and reverse primer 5¢-CAGAGAACACTCCAACCTGAATC3¢) was amplified as an internal control. To detect the expression of AtVLN4 in root hair cells, a RT-PCR assay was performed according to the method described by Zhou et al. (2005), with some modifications. Root hairs were pulled using fine forceps and immersed directly in lysis buffer containing 50 mM DTT and 10 units ll)1 RNase Inhibitor (Takara). After freezing and thawing, the root hairs were digested with RNase-free DNase (TaKaRa), and the product was used as templates for the transcription of cDNA. Forty-five PCR cycles were performed with the root hair cell-specific cDNA template and 30 cycles New Phytologist (2011) 190: 667–682 www.newphytologist.com New Phytologist with the whole root cDNA template. AtVLN4-specific primers were designed to include introns so that the PCR products amplified from cDNA (738 bp) could be distinguished from PCR products amplified from genomic DNA (1400 bp). The primers for AtVLN4 were 5¢-CCAGTCAAAGCGAGCCCG-3¢ and 5¢-GAAACCAAATTTCCTCAATTTGATACA-3¢. Two other genes, GLABRA2 and EXPANSINA7, were used as controls (Lee & Cho, 2006; Lee et al., 2008). Measurement of root hair length and growth rate Root hairs were observed in 4-d-old seedlings. To measure the length of root hairs, all the hairs in the same root region (1.7–2.0 mm from the tip of each primary root) were measured using IMAGEJ software (http://rsb.info.nih.gov/ij). To examine the growth rate, root hairs were photographed twice with an interval of 2 h. The increase in length of each root hair was measured and growth rate calculated. Quantitative analysis of cytoplasmic streaming in root hairs Cytoplasmic streaming in root hairs was observed in 4-dold seedlings using an Observer D1 microscope (Carl Zeiss) equipped with a 63 · ⁄ 1.25-numerical oil immersion objective. Images were captured at 1-s intervals over a period of 2 min using an MRm Rev CCD camera (Carl Zeiss) and AxioVision 4.2 software. Particles exhibiting continuous movement were selected at random to measure and calculate the direction and velocity of particle movement. Labeling of actin filaments in tobacco pollen and Arabidopsis root hairs Actin filaments in tobacco pollen tubes were labeled according to the method described by Xiang et al. (2007). The stained pollen was observed using a confocal laser scanning microscope (Olympus FV-300, Tokyo, Japan) mounted on an inverted microscope (Olympus IX-70). Serial confocal optical sections were taken at a step size of 0.8 lm using Olympus Fluoview 4.0 software. Actin filaments in Arabidopsis root hair cells were labeled as described previously (Miller et al., 1999; Ketelaar et al., 2002, 2003), with some modifications. Four-d-old Arabidopsis roots were prefixed for 2 min with 100 lM m-maleimido benzoyl N-hydroxysuccinimide ester (Sigma) in 1% freshly prepared paraformaldehyde and 0.025% glutaraldehyde in actin-stabilizing buffer (ASB, 50 mM Pipes, 0.5 mM MgCl2, 0.5 mM CaCl2 and 37 mM KCl, pH 6.8).This was followed by immersion in 200 lM ester, 2% paraformaldehyde and 0.05% glutaraldehyde in ASB for 20 min. Roots were then fixed in a final concentration of 4% paraformaldehyde and 0.1% glutaraldehyde in ASB for 20 min. Roots were washed three times in ASB, followed by permeabilizing with 100 lg 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist ml)1 L-alpha-lysophosphatidylcholine (Sigma) in ASB for 5 min. Actin filaments were stained with 0.33 lM Alexa 488phalloidin (Molecular Probes) for 20 min, washed three times in ASB and observed as already described. Quantitative analysis of actin filament bundling in root hairs Skewness analysis was performed according to the method described by Higaki et al. (2010). The z-series stacks of all optical sections covering the whole root hair actin filament images were filtered using Gaussian blur to reduce background noise and then skeletonized using the procedure of ThinLine (a JAVA plug-in procedure; see Higaki et al., 2010). The actin filament pixels were collected into a single image using maximum-intensity projections and the skewness values were calculated. To analyse the actin organization in different regions of the root hairs, quantification of the fluorescence intensity of actin cables was performed as described by Martin et al. (2007), with small modifications. Lines were drawn perpendicularly to the long axis of the cell, across actin cables at apical (5 lm and 10 lm from the tip), subapical (12 lm and 15 lm from the tip) and shank (20 lm, 40 lm and 60 lm from the tip) regions of the root hairs. The number of fluorescence intensity peaks, as well as the peak values, along these lines was measured. The percentages of these peak values at different intensity ranges were calculated. Results AtVLN4 binds to actin filaments Based on phylogeny, plant villins were grouped into three subclasses containing varying numbers of type 2 Ca2+-bind- Research ing sites (Choe et al., 2002) (see the Supporting Information, Fig. S1). AtVLN4, along with AtVLN5, belonged to clade III and possessed three type 2 Ca2+-binding sites in G1, G2 and G4 domains. Sequence alignment further revealed that the overall structure and most of the key amino acids were conserved in AtVLN4 (Fig. S2). The results of phylogenetic analysis and sequence alignments were consistent with the results of Huang et al. (2005) and Khurana et al. (2010). These data imply that AtVLN4 possesses the general biochemical properties of villin family members. The ability of AtVLN4 to bind to F-actin was examined using high-speed co-sedimentation assays. When AtVLN4 was centrifuged alone, little AtVLN4 was detected in the pellet fraction (Fig. 1a, lanes 1 and 5). By contrast, in the presence of 200 lM Ca2+ or 2.0 mM EGTA (Fig. 1a, lanes 3 and 7), a significant amount of AtVLN4 was found in the pellet together with polymerized actin. These results indicate that AtVLN4 can bind to F-actin at both high and low concentrations of Ca2+ in vitro. To determine the affinity of AtVLN4 for binding to F-actin, increasing concentrations of AtVLN4 were incubated with actin filaments in the presence of 2.0 mM EGTA. After centrifugation, the amount of AtVLN4 in the supernatant and pellet fractions was quantified by densitometric analysis. Fig. 1(b) shows a representative experiment in which the concentration of F-actin-bound AtVLN4 was plotted against the concentration of free AtVLN4, and the data were fitted with a hyperbolic function. The average dissociation constant (Kd) for AtVLN4 binding to F-actin was 0.24 ± 0.05 lM (mean ± SD, n = 3). We also tested whether the binding activity of AtVLN4 to actin filaments was affected by Ca2+. As shown in Fig. 1(c), the percentage of AtVLN4 that cosedimented with phalloidin-stabilized-actin filaments was not statistically different (P > 0.05) at any of the Ca2+ Fig. 1 AtVLN4 binds to F-actin. (a) A high-speed co-sedimentation assay was used to determine the binding activity of AtVLN4 to F-actin. Lanes 1 and 5, AtVLN4 alone in the pellet; lanes 2 and 6, AtVLN4 alone in the supernatant; lanes 3 and 7, AtVLN4 plus actin in the pellet; lanes 4 and 8, AtVLN4 plus actin in the supernatant. Lanes 1–4 show the samples in the presence of 200 lM calcium (Ca2+), and lanes 5–8 show the samples in the presence of 2.0 mM ethylene glycol-bis(beta-aminoethyl ether)-N,N,N‘,N‘-tetraacetic acid (EGTA). (b) Increasing concentrations of AtVLN4 (0.2–3.2 lM) were incubated and co-sedimented with F-actin in the presence of 2.0 mM EGTA. The concentration of F-actin-bound AtVLN4 was plotted against the concentration of free AtVLN4 and fitted with a hyperbolic function. For this representative experiment, the Kd was 0.27 lM. (c) To determine whether Ca2+ affects the binding activity of AtVLN4 to actin filaments, AtVLN4 was incubated with phalloidin-stabilized-F-actin (3.0 lM) in the presence of various concentrations of Ca2+ and the samples were subjected to high speed co-sedimentation assays. Gels were analysed to determine the percentage of AtVLN4 in the pellet. Error bars indicate SE (n = 3). 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist (2011) 190: 667–682 www.newphytologist.com 671 New Phytologist 672 Research concentrations tested, indicating that AtVLN4 binds to Factin in a Ca2+-insensitive manner. AtVLN4 bundles actin filaments Low-speed co-sedimentation assays were performed to examine the ability of AtVLN4 to bundle actin filaments. Only small amounts of actin were detected in the pellet fraction in the absence of AtVLN4 (Fig. 2a, lane 1). By contrast, significantly more polymerized actin (P < 0.01) sedimented in the presence of AtVLN4, and the amount of actin in the pellet increased in proportion to AtVLN4 concentration (Fig. 2a, lanes 2–4; Fig. 2b). These data suggest that AtVLN4 can bundle actin filaments in the presence of 2.0 mM EGTA. To test whether the bundling activity of AtVLN4 was Ca2+-dependent, low-speed co-sedimentation assays were performed with fixed concentration of AtVLN4 in the presence of different concentrations of Ca2+. The amount of sedimented actin was significantly less at 50 lM (P = 0.006) and 200 lM Ca2+ (P = 0.003) than at 0.04 lM Ca2+ (Fig. 2c). The (a) (d) (b) (e) decreased amounts of actin in the pellet at high concentrations of Ca2+ could result from alterations in bundling, severing or monomer-binding activities, or a combination of these properties. It can be concluded, however, that AtVLN4 bundles actin filaments at both high and low concentrations of Ca2+. The effect of AtVLN4 on actin filaments was directly visualized using fluorescence microscopy. Actin filaments were individually scattered in control samples (Fig. 2d). By contrast, actin bundles were detected with the addition of AtVLN4 (Fig. 2e,f). The bundles induced by AtVLN4 at 200 lM Ca2+ (Fig. 2e, mean length = 5.7 ± 4.1 lm, n = 94) were significantly shorter (P < 0.01) than those formed in the presence of 2.0 mM EGTA (Fig. 2f, mean length = 38.6 ± 15.8 lm, n = 37). Moreover, AtVLN4 significantly reduced the length of single actin filaments (P < 0.01) in the presence of 200 lM Ca2+ (Fig. 2e, mean length = 1.6 ± 0.6 lm, n = 100) compared with the control (Fig. 2d, mean length = 8.8 ± 4.3 lm, n = 100). The reduction in filament length by AtVLN4 was dependent on the concentration of Ca2+: reactions performed in the pres(c) (f) Fig. 2 AtVLN4 bundles actin filaments at different concentrations of calcium (Ca2+). (a) A low-speed co-sedimentation assay was used to determine the bundling activity of AtVLN4 in the presence of 2.0 mM ethylene glycol-bis(beta-aminoethyl ether)-N,N,N‘,N‘-tetraacetic acid (EGTA). Lane 1, actin alone in the supernatant and pellet; lanes 2–4, actin plus 0.4 lM, 0.8 lM or 1.6 lM AtVLN4 in the supernatant and pellet. (b) Statistical analysis of actin bundling activity of AtVLN4. Increasing concentrations of AtVLN4 (0.2–2.4 lM) were incubated with F-actin in the presence of 2.0 mM EGTA and samples were subjected to low speed co-sedimentation assays. Gels were analysed to determine the percentage of actin in the pellet. Error bars indicate SE (n = 3). (c) The effect of Ca2+ on bundling activity of AtVLN4 was determined by low-speed co-sedimentation assays in the presence of different free Ca2+concentrations. Error bars indicate SE (n = 3). Asterisks represent values that are statistically different (P < 0.05 by Student’s t-test) from the percentage of actin in the pellet at 0.04 lM Ca2+. (d–f) The ability of AtVLN4 to generate actin bundles was visualized by fluorescence microscopy. F-actin (3.0 lM) was incubated with 0.4 lM AtVLN4 protein in the presence of 200 lM Ca2+ (e) or 2.0 mM EGTA (f) at room temperature for 30 min and labeled with Alexa 488-phalloidin. A sample with actin alone was used as a control (d). Bar, 5 lm. New Phytologist (2011) 190: 667–682 www.newphytologist.com 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist ence of 2.0 mM EGTA had a mean filament length of 8.3 ± 4.5 lm (Fig. 2f, n = 100). These observations confirm that AtVLN4 generates actin bundles at both high and low concentrations of Ca2+. AtVLN4 severs actin filaments in a Ca2+-dependent manner High-speed co-sedimentation assays were performed to examine the effect of AtVLN4 on actin filaments. In the presence of 200 lM Ca2+, AtVLN4 retained substantially more actin (P < 0.05) in the supernatant (Fig. S3a, lanes 2–4; Fig. S3b) than did the controls (Fig. S3a, lane 1; Fig. S3b). However, when the Ca2+ concentration was chelated to 0.5 lM or 0.04 lM, the amount of actin in the supernatant was not significant (P > 0.05) compared with the actin alone control (Fig. S3b,c). AtVLN4 also promoted dilution-mediated actin filament depolymerization Research in a Ca2+-dependent manner (Fig. S3d,e). These data suggest that AtVLN4 can depolymerize or sever actin filaments in a Ca2+-dependent manner. Time-lapse fluorescence microscopy was employed to directly determine whether AtVLN4 severs actin filaments. Few breaks were observed in the control sample (Video S1). By contrast, AtVLN4 induced breaks along actin filaments over time (Fig. 3a; Video S3). The number of breaks per unit of filament length per second (breaks lm)1 s)1) was calculated as the ‘severing frequency’ to quantify the severing activity of AtVLN4 (Andrianantoandro & Pollard, 2006; Khurana et al., 2010; Zhang et al., 2010). AtVLN4 significantly raised the severing frequency over that of the actin alone control (P < 0.01). The severing frequency increased in proportion to AtVLN4 concentration (Fig. 3b; Videos S1– S5). The severing was markedly faster at concentrations > 10 nM, making it difficult to assess quantitatively. To determine whether the severing activity of AtVLN4 was Fig. 3 AtVLN4 severs actin filaments in a calcium (Ca2+)-dependent manner. (a) Severing activity of AtVLN4 is directly observed by fluorescence microscopy. Oregon Green-labeled actin filaments (200 lM) were introduced into a flow cell, and 1.0 nM AtVLN4 in the presence of 200 lM Ca2+ was applied at time zero. Actin filaments showed breaks (arrows) as time elapsed. The time in seconds following addition of AtVLN4 is shown in the bottom right corner of each panel. See also the Supporting Information, Video S3. Bar, 10 lm. (b) Statistical analysis of severing activity. Different concentrations of AtVLN4 were perfused into the flow cells containing Oregon Green actin filaments in the presence of 200 lM Ca2+. Severing frequency was calculated as the number of breaks per unit of filament length, per unit time. At least 20 filaments for each experimental treatment were counted. Error bars represent SE (n = 3). Asterisks represent values that are statistically different (P < 0.05 by Student’s t-test) from that of the control. (c) The severing activity of AtVLN4 is Ca2+ dependent. AtVLN4 (5.0 nM) in the presence of various concentrations of Ca2+ was introduced into flow cells containing Oregon Green actin filaments. At least 20 filaments for each experimental treatment were selected to calculate the average severing frequency. Error bars represent SE (n = 3). Asterisks represent values that are statistically different (P < 0.05 by Student’s t-test) from that of the control. 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist (2011) 190: 667–682 www.newphytologist.com 673 674 Research Ca2+-dependent, similar assays were performed with fixed concentrations of AtVLN4 in the presence of different concentrations of Ca2+. As shown in Fig. 3(c), the severing frequency reduced with the decreasing of Ca2+ concentration (Videos S5–S9). When the Ca2+ concentration was 0 or 0.5 lM, the average severing frequency was not statistically different (P = 0.86) from that of the actin alone control. These data indicate that AtVLN4 can sever actin filaments in a Ca2+-dependent manner. After severing, villin ⁄ gelsolin ⁄ fragmin family member remains attached to the barbed ends of the actin filaments as a cap, thereby preventing actin fragments from reannealing. Seeded elongation assays were also employed to determine the capping activity of AtVLN4. The results demonstrate that AtVLN4 caps the barbed ends of actin filaments in a Ca2+-dependent manner (Fig. S4). AtVLN4 colocalizes with actin filaments, induces the formation of actin cables and decreases tube growth in tobacco pollen We next sought to determine the effect of AtVLN4 overexpression on actin organization in plant cells. Full-length AtVLN4 fused with GFP was expressed under the control of a pollen-specific promoter Lat52 in tobacco pollen. The GFP fluorescence was distributed evenly throughout the whole cell (Fig. S5a,b). By contrast, AtVLN4-GFP was detected associated with a longitudinally arrayed filamentous network (Figs 4a, S5c,d). Alexa 568-phalloidin labeling of the transformed cells revealed that AtVLN4 colocalized with actin filaments, especially the thick actin cables, in the apical, subapical, shank and basal regions of the pollen tube and the pollen grain (Fig. 4a). When treated with Latrunculin B, an inhibitor of actin polymerization that causes the actin network to disassemble (Morton et al., 2000), both the AtVLN4-GFP organization and actin networks were disrupted (Fig. 4a). To examine the effect of AtVLN4 overexpression on the actin cytoskeleton, we compared the actin organizations of wild type, GFP and AtVLN4-GFP transformed pollen tubes. As shown in Fig. 4(b), confocal imaging of Alexa 568-phalloidin fluorescence revealed that the actin filament network in wild-type pollen tubes consisted of thick and fine, often longitudinally oriented, cables. Overexpression of GFP did not affect actin organization (Fig. 4b). By contrast, AtVLN4-GFP tubes showed more thick actin cables than did wild-type or GFP cells (Fig. 4b); indicating that AtVLN4 induces the formation of more actin cables in tobacco pollen. Moreover, pollen tube growth was greatly altered following transformation with AtVLN4. As shown in Fig. 4(c), the length of pollen tubes growing for 3 h was significantly reduced when AtVLN4 was overexpressed. The average length, 292.5 ± 90.9 lm (n > 150), was significantly shorter than that of the untransformed tubes (350.6 ± New Phytologist (2011) 190: 667–682 www.newphytologist.com New Phytologist 133.2 lm, n > 150; P < 0.01) or the tubes transformed with GFP (340.6 ± 100.1 lm, n > 150; P < 0.01). T-DNA insertion mutants of the AtVLN4 gene exhibit a short-root-hair phenotype To gain an insight into the possible functions of AtVLN4 in Arabidopsis, we used RT-PCR analysis to investigate its expression pattern within different tissues. As shown in Fig. S6(a), AtVLN4 had higher expression levels in roots than in aerial tissues. The data from the Affymetrix ATH1 GeneChip arrays in the Genevestigator database further indicate that AtVLN4 (At4g30160) is more abundant in the root hair zone than in the root tip and elongation zone (Zimmermann et al., 2004) (Fig. S6b). The root map for AtVLN4 expression generated from the AREX database suggests more directly that AtVLN4 might be expressed in root hair cells (Birnbaum et al., 2003; Brady et al., 2007; Cartwright et al., 2009) (Fig. S6c). To verify this, the expression of AtVLN4 in root hair cells was examined. As shown in Fig. 5(a), the AtVLN4 transcript was present in root hair cells. To analyse the function of AtVLN4 in Arabidopsis, we obtained two T-DNA insertion lines, SALK_049058 (atvln4-1) and SAIL_517_A03 (atvln4-2), from the ABRC and isolated homozygous plants (Fig. S7a,b). Sequencing analysis of the T-DNA borders revealed that atvln4-1 and atvln4-2 have T-DNA insertion at 88 bp downstream of the start of the 10th intron and at 416 bp downstream of the start of 3¢-UTR, respectively (Fig. 5b). In RT-PCR analysis, using total RNA prepared from whole seedlings, both the TDNA insertion lines prevented accumulation of full-length transcripts in the mutational plants (Fig. 5(d), inset), which was further confirmed by quantitative real-time RT-PCR (Fig. S7d). Neither of the mutant lines showed phenotypic changes in their primary root length (Fig. S8). However, both the mutant lines had shorter root hairs than those of wild-type plants (Fig. 5c). As shown in Fig. 5(d), the length of root hairs was significantly reduced in atvln4-1 (167.6 ± 25.0 lm; P < 0.01) and atvln4-2 (201.9 ± 33.1 lm; P < 0.01) plants compared with wild-type plants (336.7 ± 43.1 lm). The growth rate of root hairs also significantly decreased in atvln4-1 (0.75 ± 0.34 lm min)1; P < 0.01) and atvln4-2 (0.77 ± 0.32 lm min)1; P < 0.01) plants compared with wild-type plants (1.03 ± 0.34 lm min)1). These data suggest that AtVLN4 is required for normal root hair growth. Direction and velocity of cytoplasmic streaming are altered in atvln4 root hairs We then observed cytoplasmic streaming within atvln4 root hairs. As shown in Video S10, wild-type root hairs presented a known pattern of reverse fountain streaming: particles rap- 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist (a) Research AtVLN4-GFP F-actin (b) Merge (c) Fig. 4 AtVLN4 colocalizes with actin filaments, induces the formation of actin cables and slows tube growth of tobacco pollen. (a) Tobacco pollen was transformed with Lat52:AtVLN4-GFP plasmid (3.0 lg) by microprojectile bombardment, and stained with Alexa 568-phaloidin (red signal). AtVLN4-GFP and actin filaments were colocalized in the pollen grain (the top panel) and the pollen tube (the middle panel), as judged by the presence of yellow filaments in the merged images. Inset showed the localization of AtVLN4-GFP and actin filaments in the tip region of pollen tube. When treated with 100 nM Latrunculin B (LatB), both the structures of AtVLN4-GFP and actin filaments were disrupted (the bottom panel). Bars, 10 lm. (b) Comparison of actin organization in untransformed (WT), GFP and AtVLN4-GFP (AtVLN4) transformed tobacco pollen. Staining conditions and imaging parameters were identical between pollen samples. Images shown are z-series stacks of all optical sections. For each group, > 10 pollen tubes were observed and showed similar staining patterns. Bar, 10 lm. (c) Length of pollen tubes (mean ± SE, n > 150) after germination in culture medium for 3 h. Asterisk represents values that are statistically different (P < 0.05 by Student’s t-test) from that of the control. These data were obtained from three independent experiments. idly moved to the apex along the cortex and moved back in the center of the cell after reaching the tip region. However, the pattern of cytoplasmic streaming appeared disturbed and irregular in atvln4-1 (Video S11) and atvln4-2 (Video S12) root hairs. Cytoplasmic streaming routes became transverse to the axis of root hair growth (Fig. 6a). In order to quantify the alteration in streaming routes, we measured the angles between the tracks of particle movement and the axis of root hair growth. As shown in Fig. 6(b), the tracks of particles were always longitudinal (angle from 0 to 10) or 2011 The Authors New Phytologist 2011 New Phytologist Trust slightly oblique (angle from 10 to 30) to the growth axis in wild-type root hairs. By contrast, c. 50% particles moved in a severe oblique (angle from 30 to 80) or even transverse direction (angle from 80 to 90) in atvln4 root hairs. We also measured the velocity of cytoplasmic streaming in root hairs for each genotype. As shown in Fig. 6(c), the average velocity of cytoplasmic streaming was significantly reduced in atvln4-1 (0.82 ± 0.23 lm s)1, n = 40; P < 0.01) and atvln4-2 (0.98 ± 0.20 lm s)1, n = 40; P < 0.01) root hairs compared with that of wild type root hairs (1.36 ± New Phytologist (2011) 190: 667–682 www.newphytologist.com 675 New Phytologist 676 Research Fig. 5 AtVLN4 is expressed in root hairs and its T-DNA insertion mutants exhibit a short-root-hair phenotype. (a) Reverse-transcription polymerase chain reaction (RT-PCR) analysis of AtVLN4 transcript in Arabidopsis root hair cells. EXPANSINA7 (E7) and GLABRA2 (GL2) were used as controls for hair cell-specific and nonhair-cell-specific amplification. Gene-specific primer sets were designed to include introns to distinguish amplification from cDNA vs genomic DNA. (b) Location of T-DNA insertions. The structure of AtVLN4 is shown schematically, with coding (gray) and non-coding (white) regions. Closed triangles indicate the sites of T-DNA insertion in the mutant lines. Open triangles indicate the positions of primers used in the RT-PCR analysis. (c) Root hair phenotype of atvln4 mutants. Bar, 0.5 mm. (d) Root hair length (closed columns, mean ± SE, n > 500 from at least 30 individual roots) and growth rate (open columns, mean ± SE, n > 60 from at least 10 individual roots) of wild-type (WT) plants and atvln4 mutants. Asterisks represent values that are statistically different (P < 0.01 by Student’s t-test) from that of the control. These data were obtained from three independent experiments. Inset: RT-PCR analysis of AtVLN4 transcript in wild-type and T-DNA insertional mutants. The ACTIN2 ⁄ 8 gene was amplified as an internal control. 0.24 lm s)1, n = 40). Together, these data suggest that AtVLN4 is essential for normal cytoplasmic streaming in root hairs. Actin organization is affected in atvln4 root hairs To visualize actin organization, root hairs of wild-type and atvln4-1 plants were subjected to actin staining with Alexa488 phalloidin. As shown in Fig. 7(a), actin cables were arranged longitudinally in the shank of the wild-type root hair, and some short actin filaments were present in the tip. However, the prominent long actin cables were almost completely absent in the shank of atvln4 root hairs and were replaced by some discontinuous short cables (Fig. 7b). Two methods were employed to quantitatively evaluate actin organization in root hairs. First, a statistical parameter called skewness, which has been successfully applied to examine the extent of actin filament bundling in living guard cells of Arabidopsis (Higaki et al., 2010), was calculated for wildtype and atvln4 root hairs. The skewness value became higher when high fluorescence intensity pixel numbers were increased by actin filament bundling. As shown in Fig. 7(e), the average skewness value was significantly decreased for atvln4 root hairs (1.20 ± 0.21, n = 14; P < 0.01) compared with that of wild-type root hairs (1.98 ± 0.62, n = 14). New Phytologist (2011) 190: 667–682 www.newphytologist.com These data indicate that fewer actin polymers were organized into high-order cables in atvln4 root hairs. Second, the fluorescence intensities of actin cables were further analysed in different regions of root hair cells. Lines were drawn perpendicularly to the long axis of the root hair cell and the number of fluorescence intensity peaks, as well as the peak values, along the lines were measured. As shown in Fig. 7(f– h), the histogram of fluorescence intensity distributions for actin cables showed that the percentages of actin cables with low fluorescence intensity (< 600 au) significantly increased (P < 0.01 by v2-test) in atvln4 root hairs compared with those in wild-type hairs. These data suggest that atvln4 root hairs had consistently fewer high-order actin cables in the shank, subapical and apical regions than did the wild-type root hairs. Collectively, these data indicate that AtVLN4 is responsible not only for the formation and ⁄ or maintenance of long axial actin cables in the shank region, but also for the short actin cables in the subapical and apical regions of Arabidopsis root hairs. Discussion In this paper, we have characterized the biochemical activities of AtVLN4 in vitro and analysed its function in plant cells. Sequence alignment reveals that the key amino acids 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist Research Fig. 6 Direction and velocity of cytoplasmic streaming are altered in atvln4 root hairs. (a) Cytoplasmic streaming routes in wild-type and atvln4 root hairs. The tracks of several particles were followed over a few seconds within root hairs. The arrowhead indicates the current position of a single particle, while the recent position of the same particle is recorded by colored dots. The time in seconds following imaging is shown in the top right corner of each panel. See also the Supporting Information, Videos S10–S12. Bar, 10 lm. (b) Quantitative analysis of streaming direction in wild-type and atvln4 root hairs. The angles between the tracks of particle movement and the axis of root hair growth were measured and grouped into four types: longitudinal (angles from 0 to 10), oblique (angles from 10 to 30), severe oblique (angles from 30 to 80) and transverse (angles from 80 to 90). The percentage of particles within each group was analysed and presented. Forty cytoplasmic streaming particles from several independent root hairs were measured for each genotype. (c) Velocity of cytoplasmic streaming (mean ± SE, n = 40) in wild-type and atvln4 root hairs. Asterisks represent values that are statistically different (P < 0.01 by Student’s t-test) from the velocity of cytoplasmic streaming in wild-type root hairs. These data were obtained from three independent experiments. are conserved in AtVLN4 (Fig. S2), implying that AtVLN4 maintains the general biochemical activities of villin family members. Indeed, AtVLN4 exhibits multiple effects on actin filaments. First, low-speed co-sedimentation assays and direct visualization of actin filaments by fluorescence microscopy show that AtVLN4 efficiently bundles actin filaments at both low and high concentrations of Ca2+ (ranging from 0.04 lM to 200 lM), although bundles are much shorter at 200 lM Ca2+ than at 0.04 lM Ca2+. These data imply that AtVLN4 may contribute to the generation of long and short actin filament bundles under the regulation of Ca2+ in the plant cells. Second, time-lapse fluorescence microscopy using Oregon Green actin filaments reveals that AtVLN4 exhibited a dose-dependent increase in severing 2011 The Authors New Phytologist 2011 New Phytologist Trust frequency with increasing Ca2+. This has similarity to the behavior of AtVLN3 (Khurana et al., 2010) and AtVLN5 (Zhang et al., 2010). AtVLN4, as well as AtVLN3 and AtVLN5, severs actin filaments at micromolar Ca2+ concentrations, making it a potential candidate for actin filament severing in plant cells. Finally, seeded actin elongation assays show that AtVLN4 can effectively cap the barbed ends of actin filaments in a Ca2+-dependent manner, with increased concentration of Ca2+ enhancing the capping activity. Thus, as stated above, AtVLN4 retains all the activities of villin family members, and is similar to human villin (Walsh et al., 1984) and AtVLN5 (Zhang et al., 2010), but is markedly different from AtVLN1 (Huang et al., 2005). New Phytologist (2011) 190: 667–682 www.newphytologist.com 677 New Phytologist 678 Research (a) (b) (c) (d) (e) (f) (g) (h) Fig. 7 Actin organization is affected in atvln4 root hairs. (a–d) The actin cytoskeleton of a wild-type root hair (a) and an atvln4-1 root hair (b) was visualized by confocal laser scanning microscopy after staining with Alexa 488-phalloidin. (c) and (d) show the corresponding bright-field images for (a) and (b) root hairs, respectively. Staining conditions and imaging parameters were identical between cells. Images shown are zseries stacks of all optical sections. For each line, > 20 root hairs were observed and showed similar staining patterns. Bar, 10 lm. (e) The actin cable level was substantially decreased in atvln4 root hairs based on skewness analysis. The skewness value of each root hair analysed is shown (n = 14). Dashed lines indicate the mean value of each group. Asterisks represent values that are statistically different (P < 0.01 by Student’s ttest) between the wild type and the atvln4 line. (f–h) To quantify the fluorescence intensity of actin cables in different regions of wild-type and atvln4 root hair cells, we measured the peaks of the fluorescence profiles along lines drawn across actin cables. A histogram of the peak values in the shank (f), subapical (g) and apical (h) regions of the root hairs is shown. Error bars indicate ± SE. Wild type (n = 25), closed bars; atvln4 (n = 20), open bars. These data were obtained from three independent experiments. Wild type and atvln4 root hairs show statistically different values in the shank (P < 0.01 by v2-test), subapical (P < 0.01 by v2-test) and apical (P < 0.01 by v2-test) regions. Root hair cells are specialized root epidermal cells with tubular extensions, whose development, together with pollen tubes, serves as an attractive working model for investigation of the highly complex tip growth of plant cells. In these cells, actin filament cables orientated parallel to the long axis of the tube are well characterized, and are believed to play a prominent role as tracks for the intracellular transport of organelles and vesicles (Miller et al., 1999; Sheahan et al., 2004; Lovy-Wheeler et al., 2005; Ye et al., 2009). Villin, one of the major actin bundling proteins in animal cells, has been indicated to be involved New Phytologist (2011) 190: 667–682 www.newphytologist.com in organizing and maintaining actin bundles in plant cells. Lily villins colocalize with actin cables in pollen tubes and root hairs and are responsible for bundle formation and maintenance in these cells (Yokota et al., 1998, 2003; Tominaga et al., 2000). AtVLN5 loss-of-function retards pollen tube growth and sensitizes actin filaments in pollen grains and tubes to Latrunculin B, demonstrating its role in regulating actin filament stability in pollen (Zhang et al., 2010). Injection of an anti-lily villin antibody into growing root hairs of Arabidopsis leads to actin filament unbundling (Ketelaar et al., 2002), implying that AtVLNs may also 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist participate in actin bundle formation and maintenance in root hairs. AtVLN4 bundles actin filaments in vitro and ectopic expression of AtVLN4 in tobacco pollen causes the formation of excessive actin cables. AtVLN4 is expressed in root hairs. Moreover, knock-out of AtVLN4 eliminates the long axial actin cables and causes the formation and ⁄ or existence of fewer actin cables in root hairs. This alters the direction and velocity of cytoplasmic streaming in root hairs and leads to reduced root hair growth. These data demonstrate that AtVLN4 is responsible for the formation and ⁄ or maintenance of long axial actin cables in the shank regions of root hairs. Overexpression and loss of function of AtVLN4 have contrary effects on actin organization, but both result in impaired growth rate of pollen tubes and root hairs, suggesting that actin organization is precisely controlled and regulated in plant cells. Our results also provide evidence that intact, longitudinal actin cables are essential for cytoplasmic streaming in root hairs and that AtVLN4 is a significant regulator of this process. All the AtVLNs studied, including AtVLN1 (Huang et al., 2005), AtVLN3 (Khurana et al., 2010) and AtVLN5 (Zhang et al., 2010), efficiently bundle actin filaments in vitro. These villin isovariants may cooperate to generate actin bundles within plant cells. AtVLN1 and AtVLN3 coexist in many cell types and have overlapping and distinct activities in actin bundle formation and turnover (Khurana et al., 2010). In addition to the villin proteins, several other actin-bundling proteins have been identified, including formin (Cheung & Wu, 2004; Ye et al., 2009), fimbrin (Kovar et al., 2000) and LIM proteins (Thomas et al., 2006; Papuga et al., 2010). There may be cooperative action among these actin bundling proteins to elaborate actin bundles in plant cells. Similar to pollen tubes, the actin cytoskeleton is subjected to significant rearrangement in the subapical regions and tips of root hairs. The long actin bundles in the shank flare out into short and fine bundles, whose precise conformation has not yet been clearly defined, at the subapical regions of root hairs (Miller et al., 1999; Ketelaar et al., 2002, 2003). Debate surrounds the presence of filamentous actin in the extreme apical areas of growing root hairs (Baluska et al., 2000; Ketelaar et al., 2003; Sheahan et al., 2004; Wang et al., 2004; Voigt et al., 2005). Root hairs, as well as pollen tubes, have an oscillatory, tip-high Ca2+ gradient that is considered responsible for regulating and modulating the dynamics and arrangement of the actin cytoskeleton by the activation ⁄ inactivation of Ca2+-dependent ABPs (Ren & Xiang, 2007). With evidence of P-135-ABP accelerating the polymerization and depolymerization of actin in a Ca2+dependent manner, as well as AtVLN3 and AtVLN5 severing actin filaments in the physiological range of Ca2+ (1– 10 lM), villins have been implicated to be Ca2+-regulated proteins involved in modifying actin organization and dynamics in the apical regions of root hairs and pollen tubes (Yokota et al., 2005; Khurana et al., 2010; Staiger et al., 2011 The Authors New Phytologist 2011 New Phytologist Trust Research 2010; Zhang et al., 2010). AtVLN4 possesses severing activities on actin filaments at physiological Ca2+ levels. Moreover, AtVLN4 caps the barbed ends of actin filaments in a Ca2+-dependent manner. Thus, it is plausible to predict that AtVLN4 may participate in regulating actin organization in the regions close to the tips of root hairs. Indeed, loss of function of AtVLN4 results in fewer actin cables in the subapical and apical regions of root hairs. Along with the result that AtVLN4 generates relatively short actin bundles at high concentrations of Ca2+in vitro, we provide evidence that AtVLN4 is involved in organizing actin filaments into fine and short bundles, probably through its actin filament bundling, severing and ⁄ or capping activities, in the subapical and apical regions of root hairs. Recently, Staiger et al. (2009) directly examined the behavior of cortical actin in epidermal cells and revealed that actin filament dynamics are dominated by rapid growth and severing activity. Further work directly measuring dynamic parameters of the actin cytoskeleton in atvln4 root hairs is required to verify and illustrate whether and how villin isoforms regulate actin dynamics in the apical regions in response to tip-high Ca2+ fluxes. Acknowledgements We thank Dr David Kovar and Yujie Li (University of Chicago, IL, USA) for their helpful suggestions on actin filament severing assays. We also thank Dr Jinxing Lin and Dr Yinglang Wan (Institute of Botany, Chinese Academy of Sciences) for their friendly help with microscope use. This work was supported by the National Basic Research Program of China (2007CB108700), the National Natural Science Foundation of China (30630005, 30870211, 30970174) and the Chinese Transgenic Project (2009 ZX08009-059B) to H.R. References Andrianantoandro E, Pollard TD. 2006. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF ⁄ cofilin. Molecular Cell 24: 13–23. Baluska F, Salaj J, Mathur J, Braun M, Jasper F, Samaj J, Chua NH, Barlow PW, Volkmann D. 2000. Root hair formation: F-actindependent tip growth is initiated by local assembly of profilin-supported F-actin meshworks accumulated within expansin-enriched bulges. Developmental Biology 227: 618–632. Birnbaum K, Shasha DE, Wang JY, Jung JW, Lambert GM, Galbraith DW, Benfey PN. 2003. A gene expression map of the Arabidopsis root. Science 302: 1956–1960. Brady SM, Orlando DA, Lee JY, Wang JY, Koch J, Dinneny JR, Mace D, Ohler U, Benfey PN. 2007. A high-resolution root spatiotemporal map reveals dominant expression patterns. Science 318: 801–806. Burtnick LD, Koepf EK, Grimes J, Jones EY, Stuart DI, McLaughlin PJ, Robinson RC. 1997. The crystal structure of plasma gelsolin: implications for actin severing, capping, and nucleation. Cell 90: 661–670. New Phytologist (2011) 190: 667–682 www.newphytologist.com 679 680 Research Cartwright DA, Brady SM, Orlando DA, Strumfels B, Benfey PN. 2009. Reconstructing spatiotemporal gene expression data from partial observations. Bioinformatics 25: 2581–2587. Cheung AY, Wu HM. 2004. Overexpression of an Arabidopsis formin stimulates supernumerary actin cable formation from pollen tube cell membrane. Plant Cell 16: 257–269. Choe H, Burtnick LD, Mejillano M, Yin HL, Robinson RC, Choe S. 2002. The calcium activation of gelsolin: insights from the 3A structure of the G4–G6 ⁄ actin complex. Journal of Molecular Biology 324: 691–702. Dos Remedios CG, Chhabra D, Kekic M, Dedova IV, Tsubakihara M, Berry DA, Nosworthy NJ. 2003. Actin binding proteins: regulation of cytoskeletal microfilaments. Physiological Reviews 83: 433–473. Ferrary E, Cohen-Tannoudji M, Pehau-Arnaudet G, Lapillonne A, Athman R, Ruiz T, Boulouha L, El Marjou F, Doye A, Fontaine JJ et al. 1999. In vivo, villin is required for Ca2+-dependent F-actin disruption in intestinal brush borders. Journal of Cell Biology 146: 819–830. Friederich E, Pringault E, Arpin M, Louvard D. 1990. From the structure to the function of villin, an actin-binding protein of the brush border. Bioessays 12: 403–408. Friederich E, Vancompernolle K, Louvard D, Vandekerckhove J. 1999. Villin function in the organization of the actin cytoskeleton. Correlation of in vivo effects to its biochemical activities in vitro. Journal of Biological Chemistry 274: 26751–26760. Fu Y, Wu G, Yang ZB. 2001. Rop GTPase-dependent dynamics of tiplocalized F-Actin controls tip growth in pollen tubes. Journal of Cell Biology 152: 1019–1032. Hartwig JH, Kwiatkowski DJ. 1991. Actin-binding proteins. Current Opinion in Cell Biology 3: 87–97. Hesterberg LK, Weber K. 1983. Demonstration of three distinct calciumbinding sites in villin, a modulator of actin assembly. Journal of Biological Chemistry 258: 365–369. Higaki T, Kutsuna N, Sano T, Kondo N, Hasezawa S. 2010. Quantification and cluster analysis of actin cytoskeletal structures in plant cells: role of actin bundling in stomatal movement during diurnal cycles in Arabidopsis guard cells. Plant Journal 61: 156–165. Huang S, Robinson RC, Gao LY, Matsumoto T, Brunet A, Blanchoin L, Staiger CJ. 2005. Arabidopsis VILLIN1 generates actin filament cables that are resistant to depolymerization. Plant Cell 17: 486–501. Janmey PA, Matsudaira PT. 1988. Functional comparison of villin and gelsolin. Effects of Ca2+, KCl, and polyphosphoinositides. Journal of Biological Chemistry 263: 16738–16743. Ketelaar T, de Ruijter NC, Emons AM. 2003. Unstable F-actin specifies the area and microtubule direction of cell expansion in Arabidopsis root hairs. Plant Cell 15: 285–292. Ketelaar T, Faivre-Moskalenko C, Esseling JJ, de Ruijter NC, Grierson CS, Dogterom M, Emons AM. 2002. Positioning of nuclei in Arabidopsis root hairs: an actin-regulated process of tip growth. Plant Cell 14: 2941–2955. Khurana P, Henty JL, Huang S, Staiger AM, Blanchoin L, Staiger CJ. 2010. Arabidopsis VILLIN1 and VILLIN3 have overlapping and distinct activities in actin bundle formation and turnover. Plant Cell 22: 2727–2748. Klahre U, Friederich E, Kost B, Louvard D, Chua NH. 2000. Villin-like actin-binding proteins are expressed ubiquitously in Arabidopsis. Plant Physiology 122: 35–48. Kovar DR, Staiger CJ, Weaver EA, McCurdy DW. 2000. AtFim1 is an actin filament crosslinking protein from Arabidopsis thaliana. Plant Journal 24: 625–636. Kuhn JR, Pollard TD. 2005. Real-time measurements of actin filament polymerization by total internal reflection fluorescence microscopy. Biophysical Journal 88: 1387–1402. New Phytologist (2011) 190: 667–682 www.newphytologist.com New Phytologist Kumar N, Khurana S. 2004. Identification of a functional switch for actin severing by cytoskeletal proteins. Journal of Biological Chemistry 279: 24915–24918. Kumar N, Tomar A, Parrill AL, Khurana S. 2004. Functional dissection and molecular characterization of calcium-sensitive actin-capping and actin-depolymerizing sites in villin. Journal of Biological Chemistry 279: 45036–45046. Lee SH, Cho HT. 2006. PINOID positively regulates auxin efflux in Arabidopsis root hair cells and tobacco cells. Plant Cell 18: 1604–1616. Lee Y, Bak G, Choi Y, Chuang WI, Cho HT, Lee Y. 2008. Roles of phosphatidylinositol 3-kinase in root hair growth. Plant Physiology 147: 624–635. Lovy-Wheeler A, Wilsen KL, Baskin TI, Hepler PK. 2005. Enhanced fixation reveals the apical cortical fringe of actin filaments as a consistent feature of the pollen tube. Planta 221: 95–104. Martin SG, Rincon SA, Basu R, Perez P, Chang F. 2007. Regulation of the formin for3p by cdc42p and bud6p. Molecular Biology of the Cell 18: 4155–4167. Miller DD, De Ruijter NC, Bisseling T, Emons AM. 1999. The role of actin in root hair morphogenesis: studies with lipochito-oligo-saccharide as a growth stimulator and cytochalasin as an actin perturbing drug. Plant Journal 17: 141–154. Morton WM, Ayscough KR, McLaughlin PJ. 2000. Latrunculin alters the actin-monomer subunit interface to prevent polymerization. Nature Cell Biology 2: 376–378. Nakayasu T, Yokota E, Shimmen T. 1998. Purification of an actinbinding protein composed of 115-kDa polypeptide from pollen tubes of lily. Biochemical and Biophysical Research Communications 249: 61–65. Northrop J, Weber A, Mooseker MS, Franzini-Armstrong C, Bishop MF, Dubyak GR, Tucker M, Walsh TP. 1986. Different calcium dependence of the capping and cutting activities of villin. Journal of Biological Chemistry 261: 9274–9281. Otto JJ. 1994. Actin-bundling proteins. Current Opinion in Cell Biology 6: 105–109. Papuga J, Hoffmann C, Dieterle M, Moes F, Moreau F, Tholl S, Steinmetz A, Thomas C. 2010. Arabidopsis LIM Proteins: a family of actin bundlers with distinct expression patterns and modes of regulation. Plant Cell 22: 3034–3052. Pardee JD, Spudich JA. 1982. Purification of muscle actin. Methods in Cell Biology 24: 271–289. Pollard TD. 1983. Measurement of rate constants for actin filament elongation in solution. Analytical Biochemistry 134: 406–412. Ren H, Gibbon BC, Ashworth SL, Sherman DM, Yuan M, Staiger CJ. 1997. Actin purified from maize pollen functions in living plant cells. Plant Cell 9: 1445–1457. Ren H, Xiang Y. 2007. The function of actin-binding proteins in pollen tube growth. Protoplasma 230: 171–182. Sheahan MB, Staiger CJ, Rose RJ, McCurdy DW. 2004. A green fluorescent protein fusion to actin-binding domain 2 of Arabidopsis fimbrin highlights new features of a dynamic actin cytoskeleton in live plant cells. Plant Physiology 136: 3968–3978. Shimmen T, Hamatani M, Saito S, Yokota E, Mimura T, Fusetani N, Karaki H. 1995. Roles of actin filaments in cytoplasmic streaming and organization of transvacuolar strands in root hair cells of Hydrocharis. Protoplasma 185: 188–193. Staiger CJ, Poulter NS, Henty JL, Franklin-Tong VE, Blanchoin L. 2010. Regulation of actin dynamics by actin-binding proteins in pollen. Journal of Experimental Botany 61: 1969–1986. Staiger CJ, Sheahan MB, Khurana P, Wang X, McCurdy DW, Blanchoin L. 2009. Actin filament dynamics are dominated by rapid growth and severing activity in the Arabidopsis cortical array. Journal of Cell Biology 184: 269–280. 2011 The Authors New Phytologist 2011 New Phytologist Trust New Phytologist Su H, Wang T, Dong HJ, Ren HY. 2007. The villin ⁄ gelsolin ⁄ fragmin superfamily proteins in plants. Journal of Integrative Plant Biology 49: 1183–1191. Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24: 1596–1599. Thomas C, Hoffmann C, Dieterle M, Troys MV, Ampe C, Steinmetza A. 2006. Tobacco WLIM1 is a novel F-actin binding protein involved in actin cytoskeleton remodeling. Plant Cell 18: 2194–2206. Thomas C, Tholl S, Moes D, Dieterle M, Papuga J, Moreau F, Steinmetz A. 2009. Actin bundling in plants. Cell Motility and the Cytoskeleton 66: 940–957. Tominaga M, Yokota E, Vidali L, Sonobe S, Hepler PK, Shimmen T. 2000. The role of plant villin in the organization of the actin cytoskeleton, cytoplasmic streaming and the architecture of the transvacuolar strand in root hair cells of Hydrocharis. Planta 210: 836–843. Vandekerckhove J. 1990. Actin-binding proteins. Current Opinion in Cell Biology 2: 41–50. Voigt B, Timmers AC, Samaj J, Muller J, Baluska F, Menzel D. 2005. GFP-FABD2 fusion construct allows in vivo visualization of the dynamic actin cytoskeleton in all cells of Arabidopsis seedlings. European Journal of Cell Biology 84: 595–608. Walsh TP, Weber A, Higgins J, Bonder EM, Mooseker MS. 1984. Effect of villin on the kinetics of actin polymerization. Biochemistry 23: 2613–2621. Wang YS, Motes CM, Mohamalawari DR, Blancaflor EB. 2004. Green fluorescent protein fusions to Arabidopsis fimbrin 1 for spatio-temporal imaging of F-actin dynamics in roots. Cell Motility and the Cytoskeleton 59: 79–93. Xiang Y, Huang X, Wang T, Zhang Y, Liu Q, Hussey PJ, Ren H. 2007. ACTIN BINDING PROTEIN 29 from Lilium pollen plays an important role in dynamic actin remodeling. Plant Cell 19: 1930–1946. Ye J, Zheng Y, Yan A, Chen N, Wang Z, Huang S, Yang Z. 2009. Arabidopsis formin3 directs the formation of actin cables and polarized growth in pollen tubes. Plant Cell 21: 3868–3884. Yokota E, Muto S, Shimmen T. 2000. Calcium-calmodulin suppresses the filamentous actin-binding activity of a 135-kilodalton actin-bundling protein isolated from lily pollen tubes. Plant Physiology 123: 645–654. Yokota E, Shimmen T. 1999. The 135-kDa actin-bundling protein from lily pollen tubes arranges F-actin into bundles with uniform polarity. Planta 209: 264–266. Yokota E, Takahara K, Shimmen T. 1998. Actin-bundling protein isolated from pollen tubes of lily. Biochemical and immunocytochemical characterization. Plant Physiology 116: 1421–1429. Yokota E, Tominaga M, Mabuchi I, Tsuji Y, Staiger CJ, Oiwa K, Shimmen T. 2005. Plant villin, lily P-135-ABP, possesses G-actin binding activity and accelerates the polymerization and depolymerization of actin in a Ca2+-sensitive manner. Plant Cell Physiology 46: 1690–1703. Yokota E, Vidali L, Tominaga M, Tahara H, Orii H, Morizane Y, Hepler PK, Shimmen T. 2003. Plant 115-kDa actin-filament bundling protein, P-115-ABP, is a homologue of plant villin and is widely distributed in cells. Plant Cell Physiology 44: 1088–1099. Zhang H, Qu X, Bao C, Khurana P, Wang Q, Xie Y, Zheng Y, Chen N, Blanchoin L, Staiger CJ, Huang S. 2010. Arabidopsis VILLIN5, an actin filament bundling and severing protein, is necessary for normal pollen tube growth. Plant Cell 22: 2749–2767. Zhou B, Sauter M, Sun M. 2005. Single microspore reverse transcriptionpolymerase chain reaction: dynamic change of gene expression during microsporogenesis. Bioscience Biotechnology and Biochemistry 69: 2240–2243. Zimmermann P, Hirsch-Hoffmann M, Hennig L, Gruissem W. 2004. GENEVESTIGATOR. Arabidopsis microarray database and analysis toolbox. Plant Physiology 136: 2621–2632. 2011 The Authors New Phytologist 2011 New Phytologist Trust Research Supporting Information Additional supporting information may be found in the online version of this article. Fig. S1 Phylogenetic analysis of villins from angiosperms. Fig. S2 Multiple sequence alignment of plant villin proteins. Fig. S3 AtVLN4 severs or depolymerizes actin filaments in a calcium (Ca2+)-dependent manner. Fig. S4 AtVLN4 inhibits actin filament elongation from barbed ends in a calcium (Ca2+)-dependent manner. Fig. S5 Subcellular localization of GFP, AtVLN4-GFP and GFP-fABD2 in tobacco pollen. Fig. S6 AtVLN4 is abundant in the Arabidopsis root hair zone. Fig. S7 Molecular analysis of AtVLN4 T-DNA insertion lines. Fig. S8 Comparison of primary root length (mean ± SE, n > 20) in wild-type and atvln4 plants. Video S1 Time-lapse fluorescence microscopy series of 200 lM Oregon Green-actin filaments treated with fluorescence buffer only. Video S2 Actin filaments severed by 0.5 nM AtVLN4 in the presence of 200 lM calcium (Ca2+). Video S3 Actin filaments severed by 1 nM AtVLN4 in the presence of 200 lM calcium (Ca2+). Video S4 Actin filaments severed by 2.5 nM AtVLN4 in the presence of 200 lM calcium (Ca2+). Video S5 Actin filaments severed by 5 nM AtVLN4 in the presence of 200 lM calcium (Ca2+). Video S6 Actin filaments show minimal breakage when treated with 5 nM AtVLN4 in the absence of calcium (Ca2+). Video S7 Actin filaments show minimal breakage when treated with 5 nM AtVLN4 in the presence of 0.5 lM calcium (Ca2+). Video S8 Actin filaments severed by 5 nM AtVLN4 in the presence of 5 lM calcium (Ca2+). New Phytologist (2011) 190: 667–682 www.newphytologist.com 681 682 Research Video S9 Actin filaments severed by 5 nM AtVLN4 in the presence of 50 lM calcium (Ca2+). Video S10 This movie corresponds to the pattern of cytoplasmic streaming of a wild-type root hair (shown in Fig. 6a, left panels). Video S11 This movie corresponds to the pattern of cytoplasmic streaming of an atvln4-1 root hair (shown in Fig. 6a, middle panels). New Phytologist (2011) 190: 667–682 www.newphytologist.com New Phytologist Video S12 This movie corresponds to the pattern of cytoplasmic streaming of an atvln4-2 root hair (shown in Fig. 6a, right panels). Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office. 2011 The Authors New Phytologist 2011 New Phytologist Trust