Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
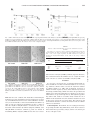
0090-9556/05/3303-336–340$20.00 DRUG METABOLISM AND DISPOSITION Copyright © 2005 by The American Society for Pharmacology and Experimental Therapeutics DMD 33:336–340, 2005 Vol. 33, No. 3 2923/1196079 Printed in U.S.A. HUMAN CYTOCHROME P450 2S1: LACK OF ACTIVITY IN THE METABOLIC ACTIVATION OF SEVERAL CIGARETTE SMOKE CARCINOGENS AND IN THE METABOLISM OF NICOTINE Shou-Lin Wang, Xiao-Yang He, and Jun-Yan Hong School of Public Health/Environmental and Occupational Health Sciences Institute (S.-L.W., X.-Y.H., J.-Y.H.), University of Medicine and Dentistry of New Jersey, Piscataway, New Jersey; and School of Public Health, Nanjing Medical University, Nanjing, People’s Republic of China (S.-L.W.) Received November 4, 2004; accepted December 15, 2004 ABSTRACT: 1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP). The full-length CYP2S1 cDNA was amplified by nested polymerase chain reaction from a human lung cDNA library and was expressed in both Chinese hamster ovary (CHO) cells and Sf9 insect cells. In contrast to the positive controls, i.e., CHO cells expressing human CYP2A13 (for NNK activation) or human CYP1A1 (for BaP activation), there was no increase in NNK- or BaP-induced toxicity in the CHO cells expressing CYP2S1. The heterologously expressed CYP2S1 proteins showed no detectable activity in metabolizing nicotine and PhIP. These results clearly demonstrate that CYP2S1 does not catalyze the metabolism of nicotine and the metabolic activation of these lung carcinogens. Cytochrome P450 (P450) enzymes are monooxygenases. Together with their redox partner NADPH-cytochrome P450 reductase, these enzymes catalyze the oxidative metabolism of a wide variety of drugs and other xenobiotics. P450s are the major enzymes responsible for the metabolic activation of environmental carcinogens and toxicants (Gibson and Skett, 1994). Although many P450 enzymes have the highest expression level in the liver, some P450 enzymes are expressed predominantly in extrahepatic tissues. Since the toxicity of a given compound is tightly linked to its metabolic fate in the target tissue, it is believed that the metabolic activation in situ of carcinogens/toxicants by extrahepatic P450 enzymes is critical for tissuespecific carcinogenicity/toxicity (Ding and Kaminsky, 2003). Recently, a novel P450 enzyme, CYP2S1, has been identified. The CYP2S1 gene is localized in the CYP2 gene cluster on chromosome 19q13.2, encoding protein that consists of 504 amino acid residues with a derived molecular mass of 55.8 kDa. CYP2S1 is predominantly expressed in human extrahepatic tissues including trachea and lung, as well as in the stomach, small intestine, spleen, and skin (Rylander et al., 2001; Rivera et al., 2002; Smith et al., 2003). It is inducible by dioxin in a human lung epithelial cell line, A549 (Rivera et al., 2002). Except for all-trans retinoic acid (Smith et al., 2003), the substrates of CYP2S1 are unknown. The high level of CYP2S1 expression in human respiratory tissues suggests that this enzyme may play a role in the metabolism of environmental chemicals that humans are exposed to through the airways. Cigarette smoking is the most important etiological factor of human lung cancer (Hoffmann et al., 1994; Spivack et al., 1997). It has been reported that about 87% of human lung cancers are caused by cigarette smoking (American Cancer Society, 2001). Whereas nicotine, a major tobacco component, is responsible for smoking addiction and may also be involved in lung carcinogenesis, many carcinogens have been identified in cigarette smoke (Hecht, 1999). These include potent lung carcinogens 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK; a tobacco-specific nitrosamine), benzo[a]pyrene (BaP; a polycyclic aromatic hydrocarbon), and 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP; a heterocyclic amine) (Hecht, 1999, 2002). All these carcinogens require P450-mediated metabolic activation to exert their carcinogenicity and toxicity. This study was supported by National Institutes of Health Grant R01-ES10048 (J.-Y.H.) and National Institute of Environmental Health Sciences Center Grant ES-05022. Article, publication date, and citation information can be found at http://dmd.aspetjournals.org. doi:10.1124/dmd.104.002923. ABBREVIATIONS: P450, cytochrome P450; NNK, 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone; BaP, benzo[a]pyrene; PhIP, 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine; CHO, Chinese hamster ovary; PCR, polymerase chain reaction; CHO-2S1, CYP2S1-expressing CHO cells; DMSO, dimethyl sulfoxide; MTS, 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfopheny)-2H-tetrazolium; LC/MS, liquid chromatography/mass spectrometry; FRT, Flp recombination target; CHO-2A13, CYP2A13-expressing CHO cells; CHO-1A1, CYP1A1-expressing CHO cells; AF, aflatoxin. 336 Downloaded from dmd.aspetjournals.org at ASPET Journals on March 4, 2016 Cytochrome P450 (P450) enzymes play a critical role in the metabolic activation of a wide variety of environmental carcinogens. Recently, a novel human P450 enzyme, CYP2S1, has been identified. It is inducible by dioxin and other classical aryl hydrocarbon receptor ligands. However, little is known regarding the substrates and the functional role of CYP2S1. Since CYP2S1 is predominantly expressed in human lung and trachea, it is reasonable to speculate that CYP2S1 may play an important role in metabolizing the environmental chemicals to which human respiratory tissues are exposed. In the present study, we examined the activity of human CYP2S1 in the metabolism of nicotine and in the activation of three potent carcinogens in cigarette smoke, 4-(methylnitrosamino)-1(3-pyridyl)-1-butanone (NNK), benzo[a]pyrene (BaP), and 2-amino- CYP2S1 IN NICOTINE/CIGARETTE SMOKE CARCINOGEN METABOLISM To understand the toxicological and physiological roles of CYP2S1, it is critical to identify its substrates. As part of our effort to explore the metabolic role of CYP2S1 for environmental carcinogens/ toxicants, the present work examined the activity of CYP2S1 in the metabolic activation of NNK and BaP by using the Chinese hamster ovary (CHO) cells that stably express human CYP2S1 cDNA. We also determined the activities of heterologously expressed CYP2S1 proteins in metabolizing nicotine and PhIP. Our results demonstrate that human CYP2S1 is not involved in the metabolism of these chemicals present in cigarette smoke. Materials and Methods with horseradish peroxidase as the secondary antibody (1:2400 dilution). The immunoblot was visualized by ECL detection according to the manufacturer’s protocol (Amersham Biosciences Inc., Piscataway, NJ). Cytotoxicity Assay. The CYP2S1-expressing CHO cells (CHO-2S1) were added into a 24-well plate (1 ⫻ 105 cells per well) with F-12 complete medium containing 800 g/ml hygromycin B and grown under 95% humidity and 5% CO2 at 37°C for 2 to 3 h to allow for the cell attachment. The cells were then treated with NNK or BaP (all dissolved in DMSO) at different concentrations for 24 h. The Flp-In CHO cells transfected with pcDNA5 vector alone (containing no cDNA insert) were used as a negative control. The Flp-In CHO cells expressing CYP2A13 (known to activate NNK) and CYP1A1 (known to activate BaP) were used as positive controls. Cytotoxicity was determined by a modified MTS [3-(4,5-dimethylthiazol2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfopheny)-2H-tetrazolium] assay using the Cell Titer 96 AQueous nonradioactive cell proliferation assay kit. For each 24-well plate, the MTS mixture was prepared by mixing 1 ml of MTS (inner salt), 50 l of phenazine methosulfate, and 11 ml of Ham’s F-12 nutrient mixture. After NNK or BaP treatment, the medium in each well was removed and 0.5 ml of MTS mixture was added. The cells were incubated with MTS mixture at 37°C in the dark for 30 min. The plate was then read at 490-nm wavelength by a Quant plate reader (Bio-Tek Instruments, Winooski, VT). In each group, the viability of cells incubated with DMSO vehicle only was set at 100%. For selected samples, the cell survival status was also determined by microscopic examination. Heterologous Expression of Human CYP2S1 Proteins. The BaculoDirect baculovirus expression system was used to express human CYP2S1 proteins according to the manufacturer’s protocol. The CYP2S1 cDNA was inserted into the pENTR-TOPO vector with Histag tail using the pENTR/D-TOPO cloning kit. After the correct DNA sequence was confirmed, CYP2S1 cDNA was transferred from pENTR vector to the BaculoDirect linear DNA; then, the construct was directly used to infect Sf9 cells for the production of CYP2S1 proteins. Hemin stock solution (2 mg/ml) was prepared by dissolving hemin in 50% ethanol and 0.2 M NaOH, and the stock solution was added to the culture medium for the final concentration of 5 g/ml. Microsomal proteins were prepared from the infected Sf9 cells by sonication and differential centrifugation as described (He et al., 2004) and stored at ⫺80°C before use. The CYP2S1 protein level in the microsomes was determined by immunoblot analysis as described previously. Microsomal P450 content was determined by reduced CO-difference spectrum after solubilization (Omura and Sato, 1964). The CO-difference spectra were recorded using a UV/visible spectrophotometer (Shimadzu UV 160U; Shimadzu, Kyoto, Japan). Metabolism of Nicotine and PhIP by CYP2S1 and LC/MS Analysis. Except for the use of CYP2S1, the methods used for the metabolism study and LC/MS analysis have been well established in our laboratory (Prabhu et al., 2001; Xu et al., 2004; Bao et al., 2005). The incubation mixture (final volume, 200 l for nicotine metabolism and 230 l for PhIP metabolism) consisted of 50 mM Tris-HCl buffer (pH 7.4), 1 mM EDTA, CYP2S1 enzyme (20 pmol), NADPH-P450 reductase (P450/reductase ⫽ 10 pmol:30 units), and nicotine (10 or 100 M) or PhIP (200 M). For nicotine metabolism, the incubation mixture included rat liver cytosol as a source of cytosolic aldehyde oxidase for cotinine formation. Human CYP2A6 and CYP2A13 proteins (obtained from our baculovirus/Sf9 cell expression system) were used as positive controls for nicotine metabolism. Human CYP1A2 (also produced by the baculovirus/Sf9 system and obtained from BD Gentest, Woburn, MA) was used as a positive control for PhIP metabolism. The amount of these P450 enzymes used in the incubation was 5 pmol. The reaction was carried out at 37°C for 30 min and terminated by the addition of an equal volume of ice-cold acetone or methanol to quench the metabolism of nicotine or PhIP, respectively. Cotinine-methyld3, as an internal standard of cotinine, was added into the reaction solution immediately after the reaction was stopped. The mixture was then centrifuged to remove the protein precipitates. Prior to the metabolite analysis by LC/MS, the supernatant from nicotine metabolism was extracted using an Oasis HLB SPE cartridge (Waters, Milford, MA) and the supernatant from PhIP metabolism was filtered using a 0.4-M polytetrafluoroethylene syringe filter (Sartorius AG, Goettingen, Germany). A TSQ Quantum triple-quadrupole mass spectrometer (Thermo Finnigan, San Jose, CA) equipped with an ion electrospray interface was used for the analysis of cotinine, a major metabolite of nicotine, and of N-hydroxy PhIP, a Downloaded from dmd.aspetjournals.org at ASPET Journals on March 4, 2016 Chemicals and Reagents. NNK (99% purity) was purchased from Chemsyn (Lenexa, KS). PhIP (98% purity) was purchased from Toronto Research Chemicals (North York, Ontario, Canada). BaP (⬎97% purity), (⫺)-nicotine (99% purity), (⫺)-cotinine (99% purity), and cotinine-methyl-d3 (99 atom % D) were obtained from Sigma-Aldrich (St. Louis, MO). Human lung firststrand cDNA library was obtained from BD Biosciences Clontech (Palo Alto, CA). CYP2S1 cDNA PCR primers were synthesized by Integrated DNA Technologies (Coralville, IA). The Penta His Antibody was purchased from QIAGEN (Valencia, CA). PfuUltra HF DNA Polymerase was obtained from Stratagene (La Jolla, CA). The Cell Titer 96 AQueous nonradioactive cell proliferation assay kit was purchased from Promega (Madison, WI). Restriction enzymes and Quick T4 DNA ligase were obtained from New England Biolabs (Beverly, MA). BaculoDirect baculovirus expression system, TOPO cDNA TA cloning kit, Flp-In CHO cells, pcDNA5/FRT, pOG44, LipofectAMINE 2000, Ham’s F-12 nutrient mixture, fetal bovine serum, penicillinstreptomycin-glutamine, and hygromycin B were purchased from Invitrogen (Carlsbad, CA). Cloning of CYP2S1 cDNA. A full-length CYP2S1 cDNA was amplified from the human lung first-strand cDNA library by a reported nested PCR method with the same outer primers (Rylander et al., 2001). The nested PCR primers were designed using the RightPrimer 1.2.5 software (BioDisk, San Francisco, CA), and the primer sequences were: forward, 5⬘-ATGGAGGCGACCGGCACCTGGGCGC-3⬘; reverse, 5⬘-TCAATGGTGATGGTGATGATGTCTGGTCTGCGTGGTGGAGTGAAG-3⬘. For the reverse primer, a 6⫻ polyhistidine tag (6⫻ Histag) sequence (underlined) was inserted immediately before the stop codon of CYP2S1 cDNA. PfuUltra HF DNA polymerase was used in the PCR amplification with the following conditions: denaturation at 94°C for 5 min, followed by 30 thermal cycles (94°C for 30 s; 64°C for 30 s, 72°C for 90 s) and extension at 72°C for 10 min. The PCR amplification generated a single major band with the predicted size of 1.5 kb after gel electrophoresis. The cDNA was subsequently cloned into a pCR 2.1-TOPO vector using the TOPO TA cloning kit, and the full-length CYP2S1 cDNA was sequenced to ensure no PCR-induced mutations. Human CYP1A1, CYP2A6, and CYP2A13 cDNAs were cloned previously by our laboratory with the same approach. Transfection and Stable Expression of CYP2S1 cDNA in CHO Cells. A Flp-In CHO line was used to establish the stable transfectant cells. The CYP2S1 cDNA inserted in the TOPO vector was transferred into a Flp-In expression vector pcDNA5/FRT using BamHI and XhoI and Quick T4 DNA ligase. This construct (pcDNA5/FRT/CYP2S1) was then used to cotransfect Flp-In CHO cells with a pOG44 vector (Flp recombinase expression vector) in a 1:9 ratio by LipofectAMINE 2000 in a six-well plate containing 3 ⫻ 105 Flp-In CHO cells per well. Forty-eight hours after the transfection, the cells were transferred into 60-mm dishes and cultured with the F-12 complete medium (Ham’s F-12 nutrient mixture, 10% fetal bovine serum, and 1% penicillin-streptomycin-glutamine) containing 800 g/ml hygromycin B. The selection medium was changed every 2 days until the foci could be identified. The well growing foci were selected to expand in a 12-well plate for 7 to 10 days. The cells were then trypsinized and transferred into a 96-well plate at 1 cell per well for monoclonal selection. After the formation of foci, 6 to 10 well growing colonies were selected for further expansion in the dishes. The presence of CYP2S1 cDNA and its protein expression in these stable transfectant cells were confirmed by PCR and immunoblotting, respectively. The immunoblot analysis used a monoclonal antibody against 6⫻ Histag as the primary antibody (1:10,000 dilution) and a goat anti-mouse IgG conjugated 337 338 WANG ET AL. carcinogenic metabolite of PhIP. The supernatant obtained from the incubation was chromatographed on a Supelco Discovery HS F5 column (Thermo Finnigan) under isocratic conditions with methanol/10 mM ammonium acetate (90:10, v/v, pH ⬃5). Cotinine, cotinine-methyl-d3, and N-hydroxy PhIP were measured by performing selected reaction monitoring. The most abundant ion transitions, m/z 177 ⬎ m/z 80 for cotinine and m/z 180 ⬎ m/z 80 for cotinine-methyl-d3, as well as m/z 241 for N-hydroxy PhIP, were selected for the identification and quantification. Results and Discussion FIG. 1. Expression of human CYP2S1 proteins in Flp-In CHO cells and in Sf9 insect cells. The expression of human CYP2S1 proteins was determined by immunoblot analysis (A and B) with a monoclonal antibody against the 6⫻ Histag that was fused with the C-terminal of CYP2S1 protein. In A and B, Neg refers to negative controls from the cells transfected with expression vectors without CYP2S1 cDNA. A, cell lysate (30 g of protein/lane) prepared from different CHO cell colonies that were transfected with CYP2S1 cDNA; B, microsomal proteins (5 g) prepared from Sf9 cells that were infected with the baculovirus-containing CYP2S1 cDNA; C, microsomal proteins (2 mg/ml) were used for P450 determination by reduced CO-difference spectrum. Downloaded from dmd.aspetjournals.org at ASPET Journals on March 4, 2016 Expression of Human CYP2S1 Proteins in Flp-In CHO Cells and in Sf9 Insect Cells. We used the Flp-In CHO cells to generate the transfectants that stably express CYP2S1. These cells contain an Flp recombination target (FRT) site in their genome and allow for the integration of the pcDNA5/FRT expression vector at the FRT site via Flp recombinase-mediated DNA recombination (O’Gorman et al., 1991). This process results in the generation of isogenic stable cell lines and ensures that the different colonies obtained from the transfection will express the same level of the transgene. In our study, colonies of stable transfectants that were resistant to hygromycin B were selected, and the integration of CYP2S1 cDNA in their genomic DNA was confirmed by PCR with CYP2S1-specific primers (data not shown). Immunoblotting with a monoclonal antibody against 6⫻ Histag, which was fused with the C-terminal of CYP2S1 protein, further demonstrated that CYP2S1 proteins were successfully expressed in these stable transfectants (Fig. 1A). There was no significant difference in the levels of CYP2S1 protein expression among different antibiotic-resistant colonies (Fig. 1A). This result is consistent with the working mechanism of the Flp-In system since, in all the stable transfectants, the transgenes are integrated at the same FRT site. To produce a sufficient amount of CYP2S1 proteins for the metabolism study, the recombinant baculoviruses containing CYP2S1 cDNA were used to infect Sf9 insect cells. Immunoblot analysis of the microsomal proteins prepared from the infected Sf9 cells showed a single protein band with the predicted molecular weight (⬃55,000) of human CYP2S1 protein (Fig. 1B). In addition, microsomes prepared from the CYP2S1-expressing Sf9 cells displayed a distinctive absorp- tion peak at 450 nm, a characteristic of P450 enzymes, in reduced CO-difference spectrum analysis (Fig. 1C). These results demonstrate that CYP2S1 proteins were also successfully expressed in the Sf9 cells. NNK- and BaP-Induced Toxicity in CHO Cells Expressing CYP2S1. To assess the potential role of CYP2S1 in the metabolic activation of NNK and BaP, the Flp-In CHO cells expressing CYP2S1 (CHO-2S1) were treated with different concentrations of BaP (0, 0.05, 0.1, 0.5, 1.0, and 2.0 M) or NNK (0, 0.1, 1.0, 10, 100, and 200 M). The selection of these concentrations was based on either a reported study (Townsend et al., 2002) or our pilot experiments. Cells transfected with the pcDNA5 vector alone (containing no P450 cDNA) were used as a negative control. Since CYP2A13 and CYP1A1 are known to be the major enzymes for the metabolic activation of NNK and BaP, respectively (Doehmer et al., 1999; Su et al., 2000), the Flp-In CHO cells expressing either CYP2A13 (CHO-2A13) or CYP1A1 (CHO-1A1) were used as corresponding positive controls. The cytotoxicity was determined by a widely used MTS assay that measures the cell viability. The working principle of the MTS assay is that the mitochondrial dehydrogenase released during cell death can reduce MTS to a colored formazan product that can be measured directly by a plate reader for absorption at 490 nm (O’Toole et al., 2003). There was no significant cell death induced by NNK and BaP treatment in the cells transfected with the pcDNA5 vector alone. This is consistent with the fact that these cells do not express the P450 enzymes that metabolically activate NNK and BaP. NNK and BaP caused a dose-dependent decrease in cell viability in the CHO-2A13 and CHO-1A1 cells, respectively. In contrast, there was no change in the viability in the CHO-2S1 cells treated with either NNK or BaP, even at the highest concentrations (Fig. 2). The result of the MTS assay was confirmed by the microscopic examination. As shown in Fig. 3, whereas the surviving cell number was significantly reduced in the CHO-2A13 cells treated with NNK (200 M) or the CHO-1A1 cells treated with BaP (1 M), there was no significant cell death in the CHO-2S1 cells treated with the same concentration of NNK or a 5-fold higher concentration of BaP. These results clearly indicate that CYP2S1 plays an insignificant role in the metabolic activation of NNK and BaP. Metabolism of Nicotine and PhIP by CYP2S1. The major pathway of nicotine metabolism in humans is to form cotinine through C-oxidation. The first step is catalyzed by the P450 system to produce the intermediate nicotine-⌬⫺1⬘(5⬘)-imminium ion, which is further oxidized to cotinine by cytosolic aldehyde oxidase (Gorrod and Hibberd, 1982; Peterson et al., 1987; Williams et al., 1990). Previous studies have shown that CYP2A6 is the major enzyme responsible for metabolizing nicotine to cotinine in humans (Cashman et al., 1992). Recently, we have demonstrated that CYP2A13 is at least as efficient as CYP2A6 for this reaction (Bao et al., 2005). On the other hand, human CYP1A2 is the key enzyme catalyzing the activation of PhIP to form N-hydroxy PhIP, which is a major and carcinogenic metabolite (Wallin et al., 1990). To determine whether CYP2S1 is active in the metabolism of nicotine and in the metabolic activation of PhIP, heterologously expressed CYP2S1 was incubated with either nicotine or PhIP. The expressed CYP2S1 proteins are catalytically active, since the enzymes from the same batch of preparation were able to metabolize aflatoxin B1 (AFB1) to its several metabolites, including AFM1-diol, AFB1-diol, AFM1, AFQ1, and AFP1 (J.-S. Wang and J.-Y. Hong, unpublished results). CYP2A6 and CYP2A13 were used as the positive controls for nicotine metabolism, and CYP1A2 as a positive control for PhIP metabolism. They were obtained by the same heterologous expression system and were incubated with nicotine or 339 CYP2S1 IN NICOTINE/CIGARETTE SMOKE CARCINOGEN METABOLISM TABLE 1 Activities of different human P450 enzymes in the metabolism of nicotine and PhIP The activities were expressed as pmol of metabolite formation/min/pmol of P450. For nicotine metabolism, the activity was determined by cotinine formation, and the values are mean ⫾ S.D. of three separate experiments. For PhIP metabolism, the activity was determined by N-hydroxy PhIP formation, and the results are the average of duplicate experiments. Substrate Concentration Nicotine P450 2A6 2A13 1A2 2S1 10 M 100 M 0.70 ⫾ 0.12 5.15 ⫾ 1.29 — N.D. 4.01 ⫾ 1.25 18.97 ⫾ 2.25 — N.D. PhIP (200 M) — — 6.57 N.D. N.D., not detectable; —, not determined. able formation of N-hydroxy PhIP by CYP2S1 (20 pmol). Therefore, these results demonstrate that CYP2S1 does not catalyze the metabolism of nicotine to cotinine and of PhIP to N-hydroxy PhIP. Conclusion FIG. 3. Morphological changes in the CHO cells treated with NNK or BaP. Upper panel, CHO-2S1 and CHO-2A13 cells were treated with 200 M NNK; lower panel, CHO-2S1 and CHO-1A1 cells were treated with 5 M and 1 M BaP, respectively. The control CHO-2S1 cells were treated with the vehicle (DMSO) only. After 24 h, a significant decrease in cell numbers was observed in the NNK-treated CHO-2A13 and BaP-treated CHO-1A1 cells, but not in the CHO-2S1 cells. PhIP under the same conditions. The metabolites formed during the incubation were then determined by the LC-tandem mass spectrometry or LC/MS methods that have been well established in our laboratory. As shown in Table 1, whereas both CYP2A6 and CYP2A13 (5 pmol of each) were efficient in metabolizing nicotine to cotinine at either the 10 M or 100 M substrate concentration, there was no detectable cotinine formation by CYP2S1 (20 pmol). Similarly, at the 200 M PhIP concentration, CYP1A2 (5 pmol) was efficient in metabolizing PhIP to N-hydroxy PhIP, whereas there was no detect- To assess the toxicological roles of individual human P450 enzymes, identification of their environmental substrates is a necessary step. Until now, all-trans retinoic acid was the only known substrate of CYP2S1 (Smith et al., 2003). The predominant expression of CYP2S1 in human respiratory tissues and its induction by dioxin in mouse lung and human lung cell line A549 lead to the speculation that CYP2S1 is involved in the metabolism of a number of carcinogens to which humans are exposed through airways. Although this hypothesis may still be valid, results of our studies clearly exclude the role of CYP2S1 in the metabolism of nicotine and in the metabolic activation of three major carcinogens in cigarette smoke. The endogenous and environmental substrates as well as the functional roles of CYP2S1 remain to be determined. Acknowledgments. We thank Dr. G. F. You (Rutgers University) for providing the human lung cDNA library, Dr. X. Xu for assistance in LC-tandem mass spectrometry analysis, and Dr. A.-N. T. Kong for the use of the plate reader. Downloaded from dmd.aspetjournals.org at ASPET Journals on March 4, 2016 FIG. 2. NNK- or BaP-induced toxicity in the Flp-In CHO cells expressing human CYP2S1. Cell viability was determined by MTS assay. The Flp-In CHO cells expressing CYP2S1 were treated with different concentrations of NNK or BaP and cultured at 95% humidity and 5% CO2 at 37°C for 24 h. The Flp-In CHO cells transfected with CYP2A13 (for NNK activation) and CYP1A1 (for BaP activation) were used as positive controls; Flp-In CHO transfected with pcDNA5 alone was used as negative control in both NNK and BaP treatments. The values are mean ⫾ S.D. of triplicate samples. 340 WANG ET AL. References Address correspondence to: Dr. Jun-Yan Hong, School of Public Health, University of Medicine and Dentistry of New Jersey, Room 385, 683 Hoes Lane West, Piscataway, NJ 08854. E-mail: [email protected] Downloaded from dmd.aspetjournals.org at ASPET Journals on March 4, 2016 American Cancer Society (2001) Tobacco use, in Cancer Facts & Figures 2001, pp 29 –32, American Cancer Society, Atlanta. Bao Z, He X-Y, Ding X, Prabhu S, and Hong J-Y (2005) Metabolism of nicotine and cotinine by human cytochrome P450 2A13. Drug Metab Dispos 33:258 –261. Cashman JR, Park SB, Yang ZC, Wrighton SA, Jacob P 3rd, and Benowitz NL (1992) Metabolism of nicotine by human liver microsomes: stereoselective formation of transnicotine N⬘-oxide. Chem Res Toxicol 5:639 – 646. Ding X and Kaminsky LS (2003) Human extrahepatic cytochromes P450: function in xenobiotics metabolism and tissue-selective chemical toxicity in the respiratory and gastrointestinal tracts. Annu Rev Pharmacol Toxicol 43:149 –173. Doehmer J, Buters JT, Luch A, Soballa V, Baird WM, Morisson H, Stegeman JJ, Townsend AJ, Greenlee WF, Glatt HR, et al. (1999) Molecular studies on the toxifying effects by genetically engineered cytochromes P450. Drug Metab Rev 31:423– 435. Gibson GG and Skett P (1994) Introduction to Drug Metabolism, 2nd ed, Blackie Academic and Professional, Glasgow. Gorrod JW and Hibberd AR (1982) The metabolism of nicotine-delta 1⬘(5⬘)-iminium ion, in vivo and in vitro. Eur J Drug Metab Pharmacokinet 7:293–298. He XY, Shen J, Hu WY, Ding X, Lu AY, and Hong JY (2004) Identification of Val117 and Arg372 as critical amino acid residues for the activity difference between human CYP2A6 and CYP2A13 in coumarin 7-hydroxylation. Arch Biochem Biophys 427:143–153. Hecht SS (1999) Tobacco smoke carcinogens and lung cancer. J Natl Cancer Inst 91:1194 –1210. Hecht SS (2002) Tobacco smoke carcinogens and breast cancer. Environ Mol Mutagen 39:119 – 126. Hoffmann D, Brunnemann KD, Prokopczyk B, and Djordjevic MV (1994) Tobacco-specific N-nitrosamines and Areca-derived N-nitrosamines: chemistry, biochemistry, carcinogenicity and relevance to humans. J Toxicol Environ Health 41:1–52. O’Gorman S, Fox DT, and Wahl GM (1991) Recombinase-mediated gene activation and site-specific integration in mammalian cells. Science (Wash DC) 251:1351–1355. Omura T and Sato R (1964) The carbon monoxide-binding pigment of liver microsomes. J Biol Chem 239:2379 –2385. O’Toole SA, Sheppard BL, McGuinness EP, Gleeson NC, Yoneda M, and Bonnar J (2003) The MTS assay as an indicator of chemosensitivity/resistance in malignant gynaecological tumours. Cancer Detect Prev 27:47–54. Peterson LA, Trevor A, and Castagnoli N Jr (1987) Stereochemical studies on the cytochrome P-450 catalyzed oxidation of (S)-nicotine to the (S)-nicotine delta 1⬘(5⬘)-iminium species. J Med Chem 30:249 –254. Prabhu S, Lee MJ, Hu WY, Winnik B, Yang I, Buckley B, and Hong JY (2001) Determination of 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) and its metabolite 2-hydroxyamino-PhIP by liquid chromatography/electrospray ionization-ion trap mass spectrometry. Anal Biochem 298:306 –313. Rivera SP, Saarikoski ST, and Hankinson O (2002) Identification of a novel dioxin-inducible cytochrome P450. Mol Pharmacol 61:255–259. Rylander T, Neve EP, Ingelman-Sundberg M, and Oscarson M (2001) Identification and tissue distribution of the novel human cytochrome P450 2S1 (CYP2S1). Biochem Biophys Res Commun 281:529 –535. Smith G, Wolf CR, Deeni YY, Dawe RS, Evans AT, Comrie MM, Ferguson J, and Ibbotson SH (2003) Cutaneous expression of cytochrome P450 CYP2S1: individuality in regulation by therapeutic agents for psoriasis and other skin diseases. Lancet 361:1336 –1343. Spivack SD, Fasco MJ, Walker VE, and Kaminsky LS (1997) The molecular epidemiology of lung cancer. Crit Rev Toxicol 27:319 –365. Su T, Bao Z, Zhang Q, Smith TJ, Hong JY, and Ding X (2000) Human cytochrome P450 CYP2A13: predominant expression in the respiratory tract and its high efficiency in metabolic activation of a tobacco-specific carcinogen, 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone. Cancer Res 60:5074 –5079. Townsend AJ, Kabler SL, Doehmer J, and Morrow CS (2002) Modeling the metabolic competency of glutathione S-transferases using genetically modified cell lines. Toxicology 181-182: 265–269. Wallin H, Mikalsen A, Guengerich FP, Ingelman-Sundberg M, Solberg KE, Rossland OJ, and Alexander J (1990) Differential rates of metabolic activation and detoxification of the food mutagen 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine by different cytochrome P450 enzymes. Carcinogenesis 11:489 – 492. Williams DE, Shigenaga MK, and Castagnoli N Jr (1990) The role of cytochromes P-450 and flavin-containing monooxygenase in the metabolism of (S)-nicotine by rabbit lung. Drug Metab Dispos 18:418 – 428. Xu X, Iba MM, and Weisel CP (2004) Simultaneous and sensitive measurement of anabasine, nicotine and nicotine metabolites in human urine by liquid chromatography/tandem mass spectrometry. Clin Chem 50:1– 8.




![[4-20-14]](http://s1.studyres.com/store/data/003097962_1-ebde125da461f4ec8842add52a5c4386-150x150.png)













