Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
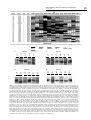
Oncogene (1998) 16, 555 ± 559 1998 Stockton Press All rights reserved 0950 ± 9232/98 $12.00 SHORT REPORT A novel 4 cM minimal deletion unit on chromosome 6q25.1-q25.2 associated with high grade invasive epithelial ovarian carcinomas Cristiano V Colitti1, Kerry J Rodabaugh1, William R Welch2, Ross S Berkowitz1, and Samuel C Mok1 1 Laboratory of Gynecologic Oncology, Department of Obstetrics, Gynecology and Reproductive Biology; 2Department of Pathology, Brigham and Women's Hospital; Harvard Medical School, Boston, Massachusetts 02115, USA Detailed deletion mapping of chromosome 6q has shown that the highest percentage of loss of heterozygosity (LOH) is located at 6q25-q27 and suggested that an ovarian cancer associated tumor suppressor gene may reside in this region. To further de®ne the smallest region of common loss, we used 12 tandem repeat markers spanning a region no more than 18 cM, located between 6q25.1 and 6q26, to examine allelic loss in 54 fresh and paran embedded invasive ovarian epithelial tumor tissues. Loss of heterozygosity was observed more frequently at the loci de®ned by marker D6S473 (14 of 32 informative cases, 44%) and marker D6S448 (17 of 40 informative cases, 43%). Detailed mapping of chromosome 6q25-q26 in these tumor samples identi®ed a 4 cM minimal region of LOH between markers D6S473 and D6S448 (6q25.1-q25.2). Loss of heterozygosity at D6S473 correlated signi®cantly both with serous versus non-serous ovarian tumors (P=0.040) and with high grade versus low grade specimens (P=0.023). The results suggest that a 4 cM deletion unit located at 6q25.1-q25.2 may contain the putative tumor suppressor gene which may play a role in the development and progression of human invasive epithelial ovarian carcinomas (IEOC). Keywords: ovary; cancer; loss of heterozygosity; chromosome 6 Ovarian carcinoma persists to be the leading cause of mortality among gynecological malignancies in the Western world. The high mortality rate is due to the late diagnosis of the disease, usually occurring during stages III and IV. Even though several oncogenes and tumor suppressor genes have been identi®ed in recent years, the genetic mechanisms involved in the initiation and progression of ovarian carcinoma remain largely unknown. Growing evidence suggests that a broad number of tumor suppressor genes might be involved in the development of dierent tumor types. Functional inactivation of these genes would deprive a constraint for cell proliferation. Although frequent LOH has been detected on chromosomes 1p, 3p, 4p, 5q, 6q, 7p, 8p, 9, 11p, 12, 13q, 16, 17p, 19p, 21q in IEOC (Lee et al., 1996; Ehlen Dubeau, 1990; Leary et al., 1993; Merlo et Correspondence: SC Mok Received 29 April 1997; revised 27 August 1997; accepted 27 August 1997 al., 1994; Zheng et al., 1991; Sato et al., 1991; Cliby et al., 1993; Cooke et al., 1996b; Lee et al., 1990; Foulkes et al., 1993; Orphanos et al., 1995b; Saito et al., 1992; Rodabaugh et al., 1995a; Wertheim et al., 1996), no tumor suppressor genes has yet been identi®ed. In recent years, chromosome 6q has been under a lot of scrutiny in ovarian cancer studies because of earlier evidence of high LOH at the estrogen receptor (ESR) site (Cooke et al., 1996b; Lee et al., 1990). Furthermore, frequent LOH on chromosome 6 has also been observed in gastric carcinoma (Queimado et al., 1995), breast cancer (Devilee et al., 1991; Iwase et al., 1995; Orphanos et al., 1995a; Sheng et al., 1996), small cell lung cancer (Merlo et al., 1994), and malignant melanoma (Millikin et al., 1991). Although frequent allelic deletion on chromosome 6q has been reported in the region 6q21-23.3, with a 43% allelic imbalance at the D6S287 locus (Orphanos et al., 1995b), more studies have been focused on the terminal region 6q24-27. (Lee et al., 1990) reported high frequencies (64%) of LOH at the ESR locus at 6q24-27. (Saito et al., 1992), using Southern blot hybridization, located a 1.9 cM commonly deleted region between loci D6S195 and D6S149, at 6q27. Furthermore, using Southern blot analysis and microsatellite markers, (Rodabaugh et al., 1995b) observed the highest rate of LOH at the 6q25-27 region, with locus D6S255 having the highest frequency of allelic loss of 43%. Recently, (Cooke et al., 1996b) has used 20 polymorphic microsatellite repeat markers to perform a more detailed investigation of LOH on 6q, particularly in the 6q27 region. Thirteen of thirty informative tumor cases (56%) showed allele loss at the estrogen receptor locus, located on 6q25, and twentythree of thirty-eight (62%) revealed LOH at locus D6S193, which falls on the 6q27 region. In this report, a set of twelve microsatellite markers, on chromosome 6q25-q26, were used to perform LOH studies on 54 invasive ovarian epithelial cancers. All tumors were informative (heterozygous) for at least one marker. The map positions and genetic distance between primers are based on previously published human genetic maps of Chromosome 6 (Cooke et al., 1996a; Gyapay et al., 1994; Volz et al., 1994), and they are shown in Figure 1. The frequency of LOH for each locus is also summarized in Figure 1. Twenty-seven of the 54 (50%) informative ovarian tumors exhibited LOH in at least one locus. Figure 2a shows the allelic deletion map for the selected cases that displayed LOH in at least one locus. Five out of 28 samples (cases 341, 353, 357, 530, ES3) showed loss at all informative loci studied, suggesting loss of the entire region. Analysis of Deletion mapping of chromosome 6q25 ± q26 in ovarian tumors CV Colitti et al 556 the pattern of LOH for the other samples illustrated a novel minimal deletion region of 4 cM ¯anked by, but not including, markers D6S441 and D6S442. This unit is de®ned by LOH at D6S448 and/or D6S473 with retention of heterozygosity at D6S441 and/or D6S442 in tumors 332, 403, 443, 498, 508, B17. Tumors 351 and 516 also show LOH for loci D6S415 and D6S437, with retention of heterozygosity at the ¯anking markers. This may suggest a second region of allelic loss, but it was not considered signi®cant because of the low LOH percentage at the two loci. Examples of LOH at four dierent markers on chromosome 6q25.1-q25.2 are shown in Figure 2b. The highest frequency of allelic loss occurred on loci D6S448 (43%) and D6S473 (44%) at 6q25.1-q25.2. Other loci that revealed high frequencies were D6S442 (36%) and D6S1007 (35%), also located at 6q25.1q25.2. Our data supports earlier studies showing a high frequency of allelic loss on the terminal portion of chromosome 6q. Several of these studies have described high frequencies of LOH at 6q27. Saito et al. (Saito et al., 1992) reported a 1.9 cM region of common allelic loss between loci D6S193 (11 of 23 informative serous cases, 48%), and D6S149 (10 of 22 informative serous cases, 45%). A 3 cM minimal region of allelic loss located between D6S264 and D6S297 at 6q27 was also documented by Cooke et al. (Cooke et al., 1996b). Other published data also point to chromosome 6q27 for the site of a possible tumor suppressor gene (Ehlen Dubeau, 1990; Zheng et al., 1991; Foulkes et al., 1993). However we identi®ed another area in the terminal region of 6q other than 6q27 which is distal to the ESR locus, at 6q25. The smallest common region of allelic loss was narrowed to a novel 4 cM segment between, but not including, loci D6S441 and D6S442, corresponding to the map position 6q25.1-q25.2. This ®nding will facilitate the identi®cation of a tumor suppressor gene(s) in the region by the positional cloning approach. Among all the markers used, loci ESR and D6S437 both showed the lowest percentage of LOH rate at 17%. Our results for ESR are in accordance with the ®ndings of both (Gallion et al., 1992) and (Leary et al., 1993), which reported LOH frequencies of 15% and Figure 1 Ideogram of chromosome 6q summarizing the 12 loci studied, map position and physical distance for each locus (Cooke et al., 1996a; Gyapay et al., 1994; Volz et al., 1994), and frequency of loss of heterozygosity in ovarian invasive tumors for each locus studied. LOH, number of tumors that showed allelic loss at each marker. Inf, number of informative samples (heterozygous). Surgical specimens of human ovarian tissue were obtained from 54 patients following a protocol approved by the Human Subjects Committee of Brigham and Women's Hospital. Archival material consisted of 11 sets of paran blocks. Control tissues consisted of segments of normal fallopian tube, uninvolved round ligament, or peripheral blood lymphocytes. All histopathological diagnoses of the invasive epithelial tumors were con®rmed and graded by a gynecological pathologist, and all cases were surgically staged according to the FIGO criteria. The invasive tumors consisted of 41 serous adenocarcinomas, ®ve mucinous cystadenocarcinomas, four endometroid adenocarcinomas, and four mixed epithelial carcinomas of the ovary. DNA was extracted from fresh specimens of 43 invasive cancers. In 11 cases DNA was extracted from archival material which had previously been ®xed in formalin and embedded in paran. Prior to the extraction, blocks containing predominantly tumor tissue were marked and selectively trimmed for tumor concentration. Paran sections were deparanized with xylene followed by absolute ethanol. All selected samples contained at least 80% of tumor cells. DNA extraction was performed using previously published methods (Edelson et al., 1997) Deletion mapping of chromosome 6q25 ± q26 in ovarian tumors CV Colitti et al 557 498 528 T N T N T N T N T N B17 T N 508 T N T N 498 443 B17 B27 T N T N D6S442 d T N T N 403 443 T N 351 T N 508 443 403 T N T N 498 D6S448 b 351 T N T N 332 403 T N 508 332 T N 498 B17 T N D6S473 c D6S441 321 a T N T N Figure 2 (a) Schematic representation of allelic deletion patterns of selected tumor cases that displayed LOH in at least one locus. The pattern of allelic loss de®ned a 4 cM minimal deletion unit ¯anked by, but not including, markers D6S441 and D6S442. (b) Autoradiographs of LOH results obtained at four of the markers used. For each panel, the marker name is shown above the patient identi®cation numbers. Tumor (T) and normal (N) tissues are matched for each patient. (a) At D6S441, patient B17 showed LOH at the tumor sample. Patients 332, 403, 498, and 508 all showed retention of heterozygosity. (b) At D6S448, allelic loss is observed in patients 403, 443, 498, 508, B17, and B27. (c) At D6S473, depicting LOH is shown in patients 332, 351, 443, 498, and 528. Tumor 403 showed retention of heterozygosity. (d) At D6S442, patients 321, 351, and 443 showed LOH at the tumor sample. Patients 498 and B17 are informative cases with no LOH. Patient 508 is an uninformative case. LOH studies were carried out by performing PCR ampli®cations of tandem repeats on chromosome 6q25-q26. Twelve oligonucleotide primer pairs were used (Research Genetics, Huntsville, AL). The dierent primers and their corresponding map positions are summarized in Figure 1. The forward primer of each set was end radio-labeled with Polynucleotide kinase (Boehringer Mannheim, Indianapolis, IN) and gamma [32P]ATP (ICN, Irvine, CA). After a 30 min incubation period, the reaction mix was diluted to a ®nal volume of 320 ml of primer-PCR mixture containing 40 ml of 10X PCR buer (0.1 M Tris-HCl, 0.5 M KCl, pH 8.3), 20-50 mM MgCl2, 20 ml of 1.25 mM deoxynucleotide triphosphate mixture, and 2 ml (10 units) of Taq polymerase (Perkin-Elmer Cetus, Norwalk, CT). Ampli®cation was carried out with 50 ng of genomic DNA using 30-35 cycles of PCR with denaturation at 958C for 1 min, annealing at 458 ± 658C for 1 min, and elongation at 728C for 1 min. 45 ml of loading buer consisting of 95% formamide, 20 mM EDTA, 0.05% bromophenol blue, and 0.05% xylene cyanol FF (Sigma, St. Louis, MO) was added to 5ml of the PCR product, and 3 ml of this mixture was loaded onto a 6% polyacrylamide gel. The electrophoresis gel was then run at a constant 1700 V, transferred onto 3 MM chromatography paper, dried, and exposed to X-ray ®lm with an intensifying screen, for a duration determined by the level of radioactivity count. The autoradiographs were then developed and analysed for indication of LOH. Cases were considered informative when heterozygosity was detected in the control tissue sample. LOH was de®ned as a visible reduction of 50% or more in the band intensity of one of the two alleles of the tumor tissue sample when compared to the normal tissue. Microsatellite instability (MSI) was de®ned as an electrophoretic mobility shift of equal intensity in tumor compared to normal tissue Deletion mapping of chromosome 6q25 ± q26 in ovarian tumors CV Colitti et al 558 Table 1 Correlation between histology, dierentiation, stage, and LOH in ovarian tumors for each locus Locus Histology Serous Mucinous Endometrioid Mixed Dierentiation Poor Moderate Well Stage I II III IV LOH/Inf (%) D6S442 D6S1007 D6S255 ESR D6S420 D6S441 D6S448 D6S473 4/19 (21) 1/4 (25) 0/3 (0) 0/3 (0) 4/20 (20) 0/2 (0) 1/2 (50) 0/2 (0) 8/28 (29) 0/2 (0) 1/4 (25) 0/3 (0) 15/29 (52) 1/4 (25) 1/4 (25) 0/3 (0) 13/24 (54) 1/4 (25) 0/2 (0) 0/2 (0) 12/28 (43) 0/2 (0) 1/3 (33) 0/3 (0) 10/22 (45) 1/4 (25) 0/1 (0) 0/4 (0) 2/14 (14) 2/10 (20) 1/5 (20) 5/18 (28) 0/6 (0) 0/2 (0) 8/24 (33) 1/8 (12) 0/5 (0) 13/23 (57) 2/11 (18) 2/6 (33) 11/17 (65) 2/10 (20) 1/5 (20) 10/24 (42) 2/10 (20) 1/2 (50) 0/1 (0) 0/2 (0) 4/24 (17) 1/2 (50) 0/3 (0) 0/1 (0) 4/19 (21) 1/3 (33) 0/3 (0) 0/4 (0) 7/27 (26) 2/3 (67) 2/4 (50) 0/5 (0) 14/28 (50) 1/3 (33) 0/3 (0) 0/2 (0) 14/26 (54) 0/1 (0) 1/4 (25) 0/3 (0) 11/26 (42) 1/3 (33) 23% respectively. We found a LOH occurrence of only 17% for the ESR locus. In contrast to our ®ndings, Cooke et al. demonstrated that LOH occurred at the ESR locus (6q25.1) at a high frequency of 56% (Cooke et al., 1996b). Another study that showed a high frequency of LOH at ESR was conducted by Lee et al., which showed that nine out of 14 informative cases (64%) exhibited allelic loss (Lee et al., 1990). The contrasting results may be due to the dierent number of informative cases and the number of cases of dierent histologic grades. While the ESR locus has been explored for allelic loss, with diering results, the region in the proximity of the ESR locus has not been examined. A greater quantity of closely-spaced markers in this region has recently become available, allowing us to further de®ne the minimally deleted segment. In this study, we mapped a region about 5 cM distal from the ESR locus, suggesting that a tumor suppressor gene may reside in the vicinity of the estrogen receptor gene but not involve it directly. The correlation between LOH and histology, dierentiation, and stage for all the ovarian tumors studied at each locus is shown in Table 1. Twenty-six of the 28 tumors showing LOH on chromosome 6q are serous adenocarcinomas, with only one endometrioid adenocarcinoma and one mucinous cystadenocarcinoma showing allelic loss in the same region. For each locus studied, chi-square P values were calculated for dierent clinicopathological parameters. P50.050 was considered signi®cant; P values between 0.050 and 0.100 were considered to suggest a dierence. The frequency of LOH at locus D6S473 (P=0.040) was signi®cantly higher for the serous cancers than of the other cancer types (Table 2). LOH was also signi®cantly higher at loci D6S255 (P=0.028) and D6S415 (P=0.033), while loci D6S448 (P=0.056) and D6S1007 (P=0.070) showed LOH to be suggestively more frequent in serous tumors than in non-serous D6S415 D6S437 D6S363 IGF2R 10/28 (36) 0/3 (0) 0/3 (0) 0/4 (0) 8/25 (32) 0/4 (0) 0/4 (0) 0/3 (0) 4/17 (24) 0/3 (0) 0/2 (0) 0/1 (0) 7/25 (28) 0/3 (0) 0/0 (0) 0/3 (0) 4/13 (31) 0/1 (0) 0/1 (0) 0/0 (0) 7/19 (37) 2/7 (29) 2/5 (40) 7/20 (35) 2/10 (20) 1/8 (13) 5/20 (25) 2/8 (25) 1/8 (13) 4/18 (22) 0/4 (0) 0/1 (0) 6/21 (29) 1/7 (14) 0/3 (0) 4/11 (36) 0/2 (0) 0/2 (0) 1/2 (50) 1/2 (50) 9/25 (36) 0/2 (0) 1/3 (33) 0/3 (0) 8/28 (29) 1/4 (25) 1/4 (25) 0/4 (0) 5/25 (20) 2/3 (67) 0/2 (0) 0/2 (0) 3/17 (18) 1/2 (50) 0/2 (0) 0/2 (0) 6/24 (25) 1/3 (33) 0/1 (0) 0/0 (0) 3/12 (25) 1/2 (50) Table 2 Relationship with clinicopathological parameters for locus D6S473 Clinicopathological characteristics Histology Serous Non-serousb Graded High Low D6S473 Total LOH (%) No LOH (%) Pa 24 8 13 (54) 1 (12) 11 (46) 7 (88) 0.040c 17 13 11 (65) 3 (23) 6 (35) 10 (77) 0.023c a LOH vs. No LOH. bNon-serous types include mucinous cystadenocarcinomas, endometrioid adenocarcinomas, and mixed epithelial carcinomas. cChi-square test. dHigh grade specimens include poorly dierentiated tumors. Low grade specimens include well and moderately dierentiated tumors ones. This ®nding seems to be in accordance with other reports that suggested that LOH in serous adenocarcinomas occurs more frequently than in other histological types (Sato et al., 1991; Foulkes et al., 1993). This suggests that losses of speci®c chromosomal regions on 6q may be involved with the development or progression of serous ovarian adenocarcinomas (Sato et al., 1991). Furthermore, a signi®cant correlation was observed between allele loss and low grade and high grade tumors (P=0.023) at locus D6S473. Loci D6S441 (P=0.083) and D6S448 (P=0.051) also showed LOH to be suggestively more frequent in high grade tumors. No other signi®cant correlation between LOH and other clinicopathological parameters was observed by Chi-Square analysis. In recent studies, allelic deletions on the terminal end of chromosome 6q have also been reported for dierent tumors. In small cell lung cancer, Merlo et al. used markers D6S255 and D6S264 on 6q25-q27 to report a 47% LOH frequency (Merlo et al., 1994). In Deletion mapping of chromosome 6q25 ± q26 in ovarian tumors CV Colitti et al breast cancer, Devilee et al. reported chromosome 6q being involved in allelic imbalance in more than 50% of informative cases. They also suggested that a potential tumor suppressor gene is thought to reside on 6q25.1 (Devilee et al., 1991). Furthermore, in malignant melanomas, Millikin et al. demonstrated that one of the markers de®ning the chromosomal region bearing the highest frequency of 6q allelic loss was ESR with 50% (Millikin et al., 1991). All these results may suggest the presence of a tumor suppressor gene in the 6q25 region that is common to ovarian and other carcinomas. On chromosome 6q25, only the superoxide dismutase 2 (SOD2) gene, which encodes the antioxidant enzyme manganese superoxide dismutase (MnSOD), has been suggested as a tumor suppressor in malignant melanoma (Volz et al., 1994; Bravard et al., 1995). Using expression vectors containing MnSOD cDNAs, Church et al. have shown that increased SOD2 gene expression modi®es the transformed phenotype of melanoma cells (Church et al., 1993). In ovarian cancer, although the SOD2 gene locus has previously been shown to have high frequencies of LOH (Cliby et al., 1993; Foulkes et al., 1993), there is no evidence that the gene has any involvement in ovarian carcinogenesis. Furthermore, the SOD2 gene locus has been located about 5 cM telomeric to D6S437 which is positioned outside our minimally deleted region (Gyapay et al., 1994; Volz et al., 1994). This suggests that there may be another novel tumor suppressor gene located in our candidate region which is involved in ovarian carcinogenesis. In conclusion, we report the identi®cation of a novel 4 cM minimal deletion unit on chromosome 6q25.1q25.2. The small size of our region may allow for the construction of a physical map of the minimally deleted region by yeast arti®cial chromosomes (YAC) cloning strategy, giving us an opportunity for direct cloning and identi®cation of a potential tumor suppressor gene associated with invasive epithelial ovarian cancer. Acknowledgements This work was supported by Public Health Service Grants R01CA63381, R01CA69453 and R01CA69291 from the National Institute of Health, Department of Health and Human Services. References Bravard A, Sabatier L, Hoschir F, Ricoul M, Luccioni C and Dutrillaux B. (1995). Int. J. Cancer, 51, 476 ± 480. Church SL, Grant JW, Ridnour LA, Oberley LW, Swanson PE, Meltzer PS and Trent JM. (1993). Proc. Natl. Acad. Sci. USA, 90, 3113 ± 3117. Cliby W, Ritland S, Hartmann L, Dodson M, Halling KC, Keeney G and Jenkins RB. (1993). Cancer Res., 53, 2393 ± 2398. Cooke IE, Cox SA, Shelling AN, Le Meuth VG, Spurr NK and Ganesan TS. (1996a). Mammal. Genome, 7, 157 ± 159. Cooke IE, Shelling AN, Le Meuth VG, Charnock ML and Ganesan TS. (1996b). Genes Chromosome & Cancer, 15, 223 ± 233. Devilee P, van Vliet M, van Sloun P, Kuipers Dijkshoorn N, Hermans J, Pearson PL and Cornelisse CJ. (1991). Oncogene, 6, 1705 ± 1711. Edelson MI, Scherer SW, Tsui LC, Welch WR, Bell DA, Berkowitz RS and Mok SC. (1997). Oncogene, 14, 2979 ± 2984. Ehlen T and Dubeau L. (1990). Oncogene, 5, 219 ± 223. Foulkes WD, Ragoussis J, Stamp GW, Allan GJ and Trowsdale J. (1993). Br. J. Cancer, 67, 551 ± 559. Gallion HH, Powell DE, Morrow JK, Pieretti M, Case E, Turker MS, Hunter JE and van Nagell JR, Jr. (1992). Gynecol. Oncol., 47, 137 ± 142. Gyapay G, Morissette J, Vignal A, Dib C, Fizames C, Millasseau P, Bernardi G, Lathrop M and Weissenbach J. (1994). Nat. Genet., 7, 246 ± 339. Iwase H, Greenman JM, Barnes DM, Bobrow L, Hodgson S and Mathew CG. (1995). Br. J. Cancer, 71, 448 ± 450. Leary JA, Doris CP, Boltz EM, Houghton CRS, Keord RF and Friedlander ML. (1993). Int. J. Gynecol. Cancer, 3, 293 ± 298. Lee JH, Kavanagh JJ, Wildrick DM, Wharton JT and Blick M. (1990). Cancer Res., 50, 2724 ± 2728. Lee WC, Balsara B, Liu Z, Jhanwar SC and Testa JR. (1996). Cancer Research, 56, 4297 ± 4301. Merlo A, Gabrielson E, Mabry M, Vollmer R, Baylin SB and Sidransky D. (1994). Cancer Res., 54, 2322 ± 2326. Millikin D, Meese E, Vogelstein B, Witkowski C and Trent J. (1991). Cancer Res., 51, 5449 ± 5453. Orphanos V, McGown G, Hey Y, Boyle JM and SantibanezKoref M. (1995a). Br. J. Cancer, 71, 290 ± 293. Orphanos V, McGown G, Hey Y, Thorncroft M, Santibanez-Koref M, Hickey I, Atkinson RJ and Boyle JM. (1995b). Br. J. Cancer, 71, 666 ± 669. Queimado L, Seruca R, Costa-Pereira A and Castedo S. (1995). Genes Chromosome & Cancer, 14, 28 ± 34. Rodabaugh KJ, Biggs RB, Qureshi JA, Barrett AJ, Welch WR, Bell DA, Berkowitz RS and Mok SC. (1995a). Oncogene, 11, 1249 ± 1254. Rodabaugh KJ, Blanchard G, Welch WR, Bell DA, Berkowitz RS and Mok SC. (1995b). Cancer Res., 55, 2169 ± 2172. Saito S, Saito H, Koi S, Sagae S, Kudo R, Saito J, Noda K and Nakamura Y. (1992). Cancer Res., 52, 5815 ± 5817. Sato T, Saito H, Morita R, Koi S, Lee JH and Nakamura Y. (1991). Cancer Res., 51, 5118 ± 5122. Sheng ZM, Marchetti A, Buttitta F, Champeme MH, Campani D, Bistocchi M, Lidereau R and Callahan R. (1996). Br. J. Cancer, 73, 144 ± 147. Volz A, Boyle JM, Cann HM, Cottingham RW, Orr HT and Ziegler A. (1994). Genomics, 21, 464 ± 472. Wertheim I, Tangir J, Muto MG, Welch WR, Berkowitz RS, Chen WY and Mok SC. (1996). Oncogene, 12, 2147 ± 2153. Zheng JP, Robinson WR, Ehlen T, Yu MC and Dubeau L. (1991). Cancer Res., 51, 4045 ± 4051. 559