Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
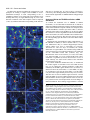
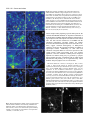
3691 Journal of Cell Science 112, 3691-3702 (1999) Printed in Great Britain © The Company of Biologists Limited 1999 JCS0835 The suppression of testis-brain RNA binding protein and kinesin heavy chain disrupts mRNA sorting in dendrites W. L. Severt1, T. U. L. Biber1, X.-Q. Wu2, N. B. Hecht2, R. J. DeLorenzo3 and E. R. Jakoi1,* 1Department of Physiology, Medical College of Virginia/Virginia Commonwealth University, Richmond, VA 23298, USA 2Center for Research on Reproduction and Women’s Health, and Department of Obstetrics and Gynecology, University of Pennsylvania School of Medicine, Philadelphia, PA 19104, USA 3Department of Neurology, Medical College of Virginia/Virginia Commonwealth University, Richmond, VA 23298, USA 4Department of Cell Biology, Duke University Medical Center, Durham, NC 27710, USA *Author for correspondence (e-mail: [email protected]) Accepted 17 August; published on WWW 18 October 1999 SUMMARY Ribonucleoprotein particles (RNPs) are thought to be key players in somato-dendritic sorting of mRNAs in CNS neurons and are implicated in activity-directed neuronal remodeling. Here, we use reporter constructs and gel mobility shift assays to show that the testis brain RNAbinding protein (TB-RBP) associates with mRNPs in a sequence (Y element) dependent manner. Using antisense oligonucleotides (anti-ODN), we demonstrate that blocking the TB-RBP Y element binding site disrupts and mislocalizes mRNPs containing α-calmodulin dependent kinase II (α-CAMKII) and ligatin mRNAs. In addition, we show that suppression of kinesin heavy chain motor protein alters only the localization of α-CAMKII mRNA. Thus, differential sorting of mRNAs involves multiple mRNPs and selective motor proteins permitting localized mRNAs to utilize common mechanisms for shared steps. INTRODUCTION and trans-acting proteins that confer stability, govern translation, and provide a means for movement. To date, most studies have focused on the sequences and structural features of mRNAs which are sufficient to direct localization when introduced as reporter constructs (Mayford et al., 1996b; MacDonald and Kerr, 1998; Muslimov et al., 1997; for review see Bassell et al., 1999). While these can be limited to a particular mRNA, others are present in a subclass of mRNAs and thus may be candidates mediating common mechanisms for shared steps. It is anticipated that many different transacting proteins will be involved including factors that direct regional (dendrite versus soma) distribution and more specific sorting to individual synapses, as well as those controlling translation, stability and movement. One of the structural features common to several localized mRNAs in hippocampal neurons is the Y element which permits formation of RNPs containing the TB-RBP (Translin) (Han et al., 1995; Wu et al., 1999; Muramatsu et al., 1998; Kobayashi et al., 1998). The TB-RBP is a microtubuleassociated protein (Han et al., 1995) that binds DNA and mRNA in a sequence specific manner (Han et al., 1995; Kwon and Hecht, 1991). In the mammalian testis, TB-RBP appears to have the dual functions of translational suppression of stored germ cell mRNAs (Kwon and Hecht, 1991, 1993) and of mRNA transport through the intercellular bridges of male germ cells, thereby, distributing mRNA between haploid cells (Morales et al., 1998; Hecht, 1998). In neurons, the activity of Cellular differentiation necessitates the regional distribution of protein to establish polarity and functional microdomains. In the polarized neuronal cell, definition of axonal-dendritic axes and of individual synapses occurs early in development and is subjected to structural reorganization throughout life in response to synaptic activity. Although spatial heterogeneity of protein can arise by the directed targeting of polypeptides, regional localization of mRNA for localized translation has recently emerged as an energetically and dynamically advantageous alternative. Several mRNAs, RNPs, and polyribosomes have been localized within distal dendrites and synaptic areas of CNS neurons by biochemical fractionation (Chicurel et al., 1993; Tiedge et al., 1991, 1993; Miyashiro et al., 1994; Crino and Eberwine, 1996) and by in situ hybridization (Garner et al., 1988; Kleinman et al., 1990; Burgin et al., 1990; Benson et al., 1992; Bian et al., 1996; Gao and Keene, 1996; Antic and Keene, 1998). These findings provide support for the concept that structural organization may be translationally regulated locally within synapses (Steward, 1997; Comery et al., 1997; Weiler et al., 1997; Mohr, 1999). However, the proteins needed for regional distribution of specific mRNAs and the functional significance of this spatial heterogeneity are not well defined. Localization of mRNA involves two classes of factors: cisacting regions of the mRNA that encode spatial information Key words: mRNA sorting, Testis-brain RNA binding protein, Kinesin, Hippocampal neuron 3692 W. L. Severt and others the Y element may function in an analogous manner permitting formation of TB-RBP associated RNPs required for mRNA sorting. How RNPs associate with the cytoskeleton and the identity of the motor protein(s) involved are largely not known. In neurons, directed movement of the RNP occurs as an energy dependent process (Davis et al., 1987) directed by microtubules. Because microtubules are bi-directionally oriented within the dendritic compartment of CNS neurons (Baas et al., 1988), members of the kinesin and/or dynein motor protein families may be involved. Several studies report microtubule-mediated movement of mRNAs into dendrites at rates (10-21 µm/hour, Davis et al., 1987; 360 µm/hour, Knowles et al., 1996; 250-400 µm/hour, Muslimov et al., 1997; 300 µm/hour, Wallace et al., 1998) consistent with calculated velocities of members of the kinesin family of motor proteins (Hirokawa, 1998). Whether these differences reflect the properties of a single motor protein or of several members of the superfamily of kinesin (or dynein) motor proteins is not understood. To fully understand the mechanisms that govern mRNA localization and its functional significance in neurons, identification and characterization of the trans-acting proteins involved are necessary. Identification of trans-acting proteins is typically done using gel mobility shift assays. To characterize their function, transfection studies in cultured cells using reporter constructs and chimeric/deletion mutants are often performed. In such studies, over-expression and/or introduction of a foreign mRNA encoding the trans-acting protein may alter the localization process. An alternate approach to understanding the role(s) of cis-acting elements and trans-acting proteins is to perturb their function. To achieve this, inactivation of mRNA by antisense oligonucleotides (anti-ODN) is widely used (Phillips and Gyurko, 1997). Typically, synthesis suppression of the cognate protein by anti-ODN results from either ribonuclease H degradation of the mRNA or inhibition of its translation. In this report, we investigate the role(s) of cellular factors important for sorting of mRNAs in hippocampal neurons. We use gel mobility shift assays to identify the trans-acting RNAbinding protein and we use antisense oligonucleotides (antiODN) to disrupt formation of RNP complexes either by blocking the cis-acting sequence or by suppressing the transacting motor protein. Subsequently, the location of the targeted mRNA and its cognate protein are determined by fluorescence in situ hybridization (FISH), confocal microscopy and immuno-cytochemistry. We reason that if differential sorting of dendritically targeted mRNA plays a key role in the distribution of a given protein, then missorting of its encoding mRNA will alter cognate protein expression. We study two dendritically targeted mRNAs: one encoding the fatty-acylated membrane-bound protein, ligatin; the other the cytosolic protein, α-calmodulin dependent kinase II (α-CAMKII). Our studies identify the microtubule-associated TB-RBP and implicate kinesin heavy chain as trans-acting proteins important for distribution of dendritically localized mRNAs containing the Y element, but not for somata-restricted mRNAs. Moreover, we show that differential sorting of multiple mRNPs is involved, whereas mRNAs for cytosolic protein(s) utilize different motor protein(s) from those used sorting membrane-bound protein(s) mRNAs. MATERIALS AND METHODS Neuronal cell cultures Rat hippocampi from neonate day 2 were trypsinized, triturated, and plated (104 cells per coverslip) onto a confluent glia feeder bed as described (Jakoi et al., 1992). Cells were maintained in defined medium for 15-18 days before use. Pyramidal neurons were identified morphologically. Neurons in culture were stained with specific antiserum for α-CAMKII (Chemicon MAB1213) and for ligatin (Jakoi et al., 1987); low levels of ligatin immuno-reactivity were found in glia. Recombinant β-galactosidase (βGAL) plasmid and adenovirus The protocols to obtain recombinant adenovirus have been described in detail by Nevins et al. (1997). Briefly, the AdCMVβGAL is a replication defective recombinant adenovirus (E1a/E1b minus) obtained by in vivo recombination between the adenovirus mutant and the pAdCMVβGAL. After isolation, the recombinant adenoviral DNA was amplified in HEK293 cells, a trans-complementing cell line for E1 function. Plaques were detected within 5 days post-infection and viruses harvested 5 days later. Recombinant clones were amplified in HEK293 cells to obtain a titer of ~1010 p.f.u./ml and triple plaque purified to insure that viral suspensions used for the in vitro experiments are free of wild-type virus. Two constructs were used: pAdCMVβGAL and pAdCMVβGAL-3′UTR500. For pAdCMVβGAL, the entire coding region of Escherichia coli lacZ was excised from pGEMβGAL (Promega) and inserted into the HindIIIBamHI site of pAdCMV (gift of Drs Okatani and Nevins, HHMI Duke University). For pAdCMVβGAL-3′UTR500, a 500-nt cDNA fragment of ligatin 3′UTR was ligated onto the 3′ end of the lacZ insert. This construct was then inserted into the HindIII-NotI region of pAdCMV. Rat hippocampal neuronal cells (2×104 per coverslip) in culture were infected with 2×105 p.f.u./µl of viral suspension in 200 µl culture medium and incubated for 30 minutes at 37°C. Culture medium (400 µl) was then added to each well and the coverslips incubated for 60 minutes at 37°C. Subsequently, the media were replaced and the cells were maintained for 4 hours at 37°C. Expression of βgalactosidase mRNA was detected by FISH and confocal microscopy using a complementary oligonucleotide probe (βGAL, 5′CAGTGAATC CGTAATCAT-3′) (Operon) tailed with biotinylateddUTP (Clontech) as described (Panchision et al., 1995). Constructs and RNA transcripts Constructs derived from ligatin cDNA were cloned into pGEM-3Z (-) (Promega) and screened by restriction analysis. The plasmids were linearized with HinfI. Radiolabeled and unlabeled RNA transcripts were synthesized in vitro with SP6 polymerase using the Riboprobe In Vitro Transcription System (Promega). The single stranded template of the UTR500 consisting of 500-nt and 59-nt of polylinker was used to synthesize transcripts for gel shift assays. Sense αCAMKII transcripts were transcribed from 29-nt template consisting of 17 nt of coding region (315-332) and 12-nt of polylinker. For control transcripts, EcoRI linearized Riboprobe pGEM-11Z (-) polylinker (58-nt) was used. For competition with the TB-RBP RNAbinding motif, sense RNA transcripts were synthesized from a 67-nt protamine 3′UTR template (pGEMc) consisting of 42-nt of 3′UTR and 25-nt of polylinker (Kwon and Hecht, 1991) using SP6 polymerase. RNA gel mobility shift assays and UV cross-linking Gel mobility shift assays were performed according to the method of Gillis and Malter (1991). Hippocampal neuronal cell lysates were prepared by repetitive (3×) freeze-thaw lysis in 25 mM Tris-HCl, pH 7.9, 0.5 mM ethylene glycol-bis (β-amino ethyl ether)-N,N,N′,N′-tetra acetic acid (EDTA), and 0.1 mM phenylmethylsulfonyl fluoride Disrupted mRNA sorting in CNS neurons 3693 (PMSF), followed by centrifugation at 15,000 g (4°C) for 15 minutes. The supernatant was stored at −70°C. Cell lysate (12 µg) and uniformly labeled [32P]RNA encoding a fragment of ligatin 3′UTR (150,000-250,000 cpm) were incubated in the absence or presence of non-radiolabeled competitors. Binding reaction mixes were incubated at 30°C for 10 minutes in a buffer containing 10% glycerol, 20 mM Tris-base, pH 7.9, 50 mM KCl, 0.3 mM EDTA, 1.1 mM dithiothreitol, 0.5 mM PMSF, 1% polyvinyl alcohol, 4mM ATP, 0.7 mM MgCl2, and 0.5 µg tRNA in a total volume of 25 µl. Heparin (40 µg) was added and the reaction mixture was incubated in ice (10 minutes) prior to digestion with RNase T1 (37°C, 30 minutes). Shift bands were analyzed by electrophoresis on a 7% nondenaturing polyacrylamide gel (80:1 acrylamide:bisacrylamide) in Tris-borate-EDTA buffer. The gel was dried and RNA-protein complexes were detected by autoradiography. For competition experiments, unlabeled competitor RNAs were synthesized as above except no radiolabeled NTPs were added to the reaction. Competitor RNAs were incubated with cell lysate at 30°C for 10 minutes prior to addition of [32P]-3′UTR probe. Competitor ODN [specific anti-ODN sequence 5′-AGCCCAGAGCTTG-3′; specific sense-ODN sequence 5′-CAAGCTCTGGGCT-3′; non-specific sequence 5′-TCTGTATAGACATGATGGCTG3′), was annealed to the radiolabeled probe (10 minutes at 65°C then slow cooled to room temperature) before incubation with the reaction mixture. Super shift assays were performed as described for shift assays except that specific antibody (anti-α-CAMKII, Chemicon MAB1213; anti-tubulin, Chemicon MAB065; anti-actin, ICN C4; anti-68 kDa intermediate filament, gift of Dr Bigbee, Medical College of Virginia; and anti-TB-RBP, Wu et al., 1997) was added to the homogenate prior to the addition of radiolabeled probe. For UV cross-linking, RNA-protein complexes were UV irradiated (5 minutes) in ice under a germicidal lamp (Phillips G15T80) located 5 cm above the reaction mixture. Irradiated shift complexes were then heparinized and digested with RNase T1. For some experiments, RNA-protein complexes were irradiated directly in the gel and the excised band then subjected to SDS-PAGE. Prior to electrophoresis, irradiated samples were suspended in SDS sample buffer without β-mercaptoethanol. SDS-PAGE was performed according to the method of Laemmli (1970) using 8% polyacrylamide gels. ODN incubation conditions Disruption of RNP formation was performed as described by Jakoi and Severt (1999). Sterile, non-derivatized, phosphodiester TB-RBP antisense motif ODN [5′-AGCCCAGAGCTTG-3′](1 µM, OPERON) was administered daily to cultured neurons for 3-4 days. As controls, TB-RBP sense motif ODN [5′-CAAGCTCTGGGCT-3′] was given as described. To suppress kinesin heavy chain function, sterile, nonderivatized phosphodiester ODN was administered to neurons in culture (50 µM initially, then 25 µM every 12 hours for 3 days) as described by Ferreira et al. (1992). For this purpose, ODN encoding the following sequences were synthesized (Operon): antisense KHC Table 1. The TB-RBP binding Y element on protamine, ligatin and α-CAMKII mRNAs mRNA Protamine 2 Ligatin α-CAMKII Sequence CTGAGCCCTGAGCT CCAAGCTCTGGGCT AGAAGCCCTATGCT Location Bases 442-455 (3′UTR) Bases 1099-1112 (3′UTR) Bases 316-329 (coding region) The Y elements found in two dendritically targeted mRNAs share strong homology with the first identified Y element from testicular protamine 2 mRNA (Kwon and Hecht, 1991). Bases printed in bold underline are invariant and compose 71% of the total sequence. (5′-CCGGGTCCGCCATCTTTCTGGCAG-3′) (5′TGCCAGAAAGATGGCGGACCCGG-3′). and sense KHC FISH and confocal microscopy FISH was performed as described (Panchision et al., 1995). Briefly, cells were fixed in 5% paraformaldehyde in phosphate buffered saline (PBS, 30 minutes), permeabilized with 70% ethanol, and prehybridized in 50% formamide for 60 minutes (42°C). Complementary oligonucleotide probe (ligatin, 5′-GTCTTCTGGGGCTTCTGAGAG-3′; α-CAMKII, 5′-GGTAGCATCCTGGCACT-3′; NSE, 5′-TCTGTATAGACATGATGGCTG-3′) (Operon) was tailed with biotinylated-dUTP (Clontech) and added to the hybridization mix. α-Tubulin cDNA probe was synthesized by SP6 polymerase using the RIBOPROBE kit and pBR322kα1(Gift of Dr Cowan, New York Medical Center). Cells were hybridized for 60 minutes at 37°C, washed 3 times with 0.5× SSC, 0.1% SDS at 37°C for 30 minutes and then blocked with 0.25% gelatin-0.1% saponin in phosphate buffered saline (PBS) for 60 minutes. Bound probe was detected with Ultra avidin-Texas red conjugate (Leinco Technologies) (60 minutes). Quantitation and image analysis Neurons were selected for analysis if they exhibited: (1) a characteristic pyramidal morphology (diamond- or triangular-shaped soma of 20-30 µm diameter, bifurcated apical dendrite, 2-3 basal dendrites) and (2) planar dendrites (continuous baseline fluorescence within dendrites) at distances >200 µm from the soma. Cells were scanned with a Zeiss LSM410 (Zeiss Inc., PA) using a C-Apo 40× objective (1.2 NA). To permit an accurate comparison in a given experimental set, the experimental and control images were obtained under exactly the same scanning conditions (laser power, photomultiplier sensitivity, filter settings, scanning duration, averaging, pinhole settings, etc) that optimized the sense-ODN treated specimens. Where indicated the soma signal of the experimental image was normalized to that of the control image by changing the photomulitplier sensitivity. Full frame images were analyzed with the Carl Zeiss Laser Scanning Microscope System Software version 3.84. The digitally recorded intensities of pixels (1 to 256) from selected Fig. 1. Expression of β-galactosidase mRNA in hippocampal neurons infected with 2×105 p.f.u. AdCMVβGAL (A) and AdCMVβGAL-3′UTR500 (B). Cultures (n=2 per group) were fixed 4 hours after infection. Specific hybridization signal was detected by FISH and confocal microscopy as described in Materials and Methods. Micrographs are representative of 3 independent experiments. Insert depicts pseudocolor intensity scale (1-256). Arrowhead denotes soma. 3694 W. L. Severt and others regions were averaged and expressed as change in intensity per unit area. Because the auto-fluorescence intensity and the relative change in fluorescence from individual neurons within a given experiment and among independent experiments did not vary significantly, no correction for auto-fluorescence (background) was applied. Data are given as the mean ± s.e.m. Statistical analysis was performed using Student’s t-test. Values of <0.05 are considered significant. The images presented are encoded with a pseudo-color table to permit discrimination between different gray levels. Two color tables were used: a standard glow color table and a customized color table (Tbrain, see Fig. 5). For illustration, the experimental and control images in a given set were printed in exactly the same manner. Immunocytochemistry. Immunocytochemistry was performed as described by Jakoi et al. (1992). Briefly, cells were fixed in 5% paraformaldehyde in PBS for 30 minutes, permeabilized in 0.01% saponin-0.25% gelatin-PBS for 60 minutes and subsequently incubated with immune serum (rabbit anti-ligatin, Jakoi et al., 1987; mouse anti-α-CAMKII, Chemicon MAB1213; or mouse anti-SUK4, Developmental Studies Hybridoma Bank, University of Iowa) for 60 minutes. Bound primary antibody was detected indirectly with either biotinylated goat anti-rabbit serum or biotinylated rat affinity purified goat anti-mouse serum (Vector Labs) and Texas red-ultra avidin as described (Jakoi et al., 1992). Cells were photographed with a Zeiss epi-fluorescence microscope using a 25× objective (0.8 NA). RESULTS Reporter construct containing the Y element sorts into dendrites Our initial interest was to identify proteins that act as transacting factors to localize mRNAs in hippocampal neurons. The discovery that the TB-RBP binds localized mRNAs encoding tau and myelin basic protein (Han et al., 1995), led us to ask whether dendritically localized mRNAs contain the TB-RBP consensus sequence and whether these mRNAs associate with the TB-RBP. Using the GenBank database, we found ligatin and α-CAMKII mRNAs contain sequences with high homology (>70%) to a TB-RBP binding sequence (Y element) (Table 1). This Y element resides within the 3′UTR of ligatin mRNA, in α-CAMKII mRNA, it is within the coding region. A chimeric βGAL construct was tested to determine whether a 500-nt region of ligatin mRNA containing the Y element was active in localizing mRNA within dendrites. For these experiments, we used a gene transfer vector from adenovirus (Nevins et al., 1997) that efficiently delivers genes into the post-mitotic neurons with no adverse effects on cell morphology, viability, or function (Moriyoshi et al., 1996; Caillaud et al., 1993). The molecular chimera was constructed by inserting the ligatin mRNA 3′UTR containing the Y element between the vector lacZ coding region and the SV40 3′UTR polyadenylation signal. The adenovirus constructs were infected at 2×105 p.f.u. for 90 minutes at 37°C. Subsequently, the virus was removed, and four hours later the encoding mRNAs were located by FISH and confocal microscopy. Both glia and neurons were infected under these conditions; the efficiency of infection was ~40%. The construct containing the 500-nt region of ligatin mRNA 3′UTR was effective in localizing β-galactosidase mRNA within dendrites of cultured neurons (Fig. 1B). In the absence of this 3′UTR region (control construct), the β-galactosidase mRNA remained confined to the somata (Fig. 1A). These data establish that a 500 nt region of ligatin 3′UTR is sufficient for sorting. Formation of RNA-protein complexes in lysates of hippocampal neurons To identify the RNA binding protein(s) involved, we next studied the in vitro formation of RNA-protein complexes using this 500-nt region in RNA gel mobility shift and UV crosslinking assays. Hippocampal cell lysates were incubated with 32P-labeled transcripts containing the same 3′UTR region of ligatin mRNA used in Fig. 1. To reduce formation of nonspecific RNA protein complexes, reaction mixtures were incubated in the presence of heparin and then digested with RNase T1. Radiolabeled complexes were detected by polyacrylamide gel electrophoresis (PAGE) under nondenaturing conditions. Transcripts generated from the distal 500-nt sequence of this message formed a single shift band (Fig. 2A, lanes 2-5). This RNA-protein complex could be competed (>50%) by unlabeled 3′UTR-500 transcript at 10fold excess (Fig. 2A, lanes 3 and 5). Non-specific competitor RNAs encoding pGEM11z(-) polylinker (58-nt) or tRNA did not compete (see below, Fig. 4A, lane 5 and Fig. 4B, lane 1, respectively). The apparent molecular mass of the trans-acting protein(s) involved in formation of this RNA-protein shift complex was determined by UV cross-linking and SDS-PAGE. RNA-protein complexes were generated, UV irradiated, excised from the gel and then analyzed by SDS-PAGE and autoradiography. A Fig. 2. Formation of a specific RNA-protein complex between 32Plabeled ligatin 3′UTR region and hippocampal neuronal cell lysate. (A) RNA mobility shift assays of cell extracts from untreated neurons were performed as described in Materials and Methods. The binding mixture was analyzed by electrophoresis on a 7% nondenaturing polyacrylamide gel. Lane 1 contains the 32P-labeled ligatin 3′UTR probe alone. Lanes 2-5 contain the ligatin 3′UTR region probe and cellular extracts from two different preparations alone (lanes 2 and 4) and in the presence of unlabeled 10-fold excess specific competitor RNA (lanes 3 and 5). Arrowhead denotes the position of the band shift complex. (B) Identification of polypeptide interacting with the ligatin 3′UTR transcript by UV cross-linking and SDS PAGE. RNA-protein binding reactions using 32P-labeled ligatin 3′UTR region as substrate were carried out as in A. 32P-labeled RNA-protein shift complexes were UV irradiated for 5 minutes and the shift bands excised. Cross-linked products were resolved directly by SDS PAGE on an 8% polyacrylamide gel. Lane 1 contains radiolabeled probe incubated with cell lysate. Lane 2 contains radiolabeled probe alone. Arrow denotes single polypeptide resolved. Prestained molecular mass standards (Pharmacia) were used as size markers. Disrupted mRNA sorting in CNS neurons 3695 Fig. 3. Effect of specific antibody on RNA-protein shift complex. Shift assays of ligatin 3′UTR region with cell lysate of hippocampal neurons were performed as described in the presence or absence of specific antibody. Lanes 1-5 contain radiolabeled probe and cellular extract. Lanes 1-4 contain radiolabeled probe and specific antibody to TB-RBP (1), tubulin (2), actin (3), 68 kDa intermediate filament (4), respectively. Lane 5 contains RNA-protein complex alone (control). Migration of the single shift band is retarded (super shift) in lane 1. Lower arrow, specific RNAprotein complex. Upper arrow, supershifted complex. single 32 kDa polypeptide was resolved (Fig. 2B, lane 1). In the absence of cell lysate, no radiolabeled polypeptides were found (Fig. 2B, lane 2). In other experiments, RNA-protein complexes were irradiated in solution prior to RNase T1 digestion and, again, a single 32 kDa polypeptide was resolved (data not shown). Because the efficiency of UV cross-linking is low, these results provide a minimal estimate of the transacting proteins that bind to this region of the 3′UTR of ligatin mRNA. Identification of TB-RBP as trans-acting protein To investigate the possibility that the 32 kDa protein in the ligatin RNA-protein complexes was TB-RBP, we used affinity purified TBP antibody in super shift mobility assays (Wu et al., 1997). In these assays, the RNA-protein complexes were incubated with the specific antibody, digested with RNase T1 and then analyzed by PAGE. In the absence of added antibody, one band was detected (Fig. 3, lane 5, lower band). In the presence of TB-RBP antibody, a slower migrating shift band (super shift band) was detected (Fig. 3, lane 1). This super shift was specific, since antibodies to cytoskeletal proteins, tubulin (Fig. 3, lane 2), α-actin (Fig. 3, lane 3), and a 68 kDa intermediate filament (Fig. 3, lane 4) did not alter the migration of the RNA-protein complex. Collectively, our findings strongly suggest that the TB-RBP of hippocampal neuronal lysates interacts with a 500 nt region of ligatin mRNA (Fig. 3, lane 5). To confirm the necessity of having an accessible Y element on the mRNA for formation of the mRNA-protein complexes, we performed competition assays as described in Fig. 2 in which neuronal cell lysates were preincubated with unlabeled RNA and ODN competitors (10 minutes, 30°C). Compared to the control (Fig. 4A, lanes 2 and 8), RNA-protein complex formation was disrupted by unlabeled transcripts encoding the testis protamine 2 Y element (Fig. 4A at 10- and 50-fold excess, lane 3 and 4) and by unlabeled antisense ODN encoding the ligatin Y element (Fig. 4A at 1:5000, lane 6). Neither tRNA (1:50) nor the TB-RBP Y element sense-ODN (1:5000) competed (Fig. 4A non-specific competitors, lanes 5 and 7). When the unlabeled competitor RNA encoding the Y element of testis protamine 2 mRNA was added to the reaction mixture without preincubation with the cell lysate, no inhibition of the RNA-protein complex was seen (data not shown). The RNA-protein complexes were also diminished by hybridizing the 32P-labeled transcript with unlabeled anti-ODN (complementary to the TB-RBP Y element of ligatin) at a molar ratio of 1:5 (specific competitor) (Fig. 4B, lane 3). However, hybridization with unlabeled RNA encoding pGEM11z (-) polylinker (Fig. 4B, lane 1) or ODN complementary to the N-methyl-D-aspartate receptor (NMDAR1, non-specific competitor) (Fig. 4B at 1:5, lane 4) did not interfere with RNA-protein interactions. These results show that RNA encoding a sequence >70% homologous to the ligatin Y element and ODN complementary to the ligatin Y element can disrupt RNA-protein complex formation. Fig. 4. Formation of RNA-protein complex involves TBRBP Y element. (A) Gel mobility shift assays using radiolabeled ligatin 3′UTR region were performed as described in Fig. 2 and formed complexes resolved by electrophoresis in non-denaturing polyacrylamide gels (80:1, acrylamide: bisacrylamide). To test for competition of RNP formation, hippocampal neuronal lysates were preincubated with unlabeled competitor for 10 minutes at 30°C. Lane 1 contains radiolabeled transcript alone. Lanes 2 and 8 contain lysate and radiolabeled probe alone (control). Lanes 3-7 show competition of RNA-protein complex by RNA and ODN. Unlabeled RNA transcript encoding TB-RBP Y element sense sequence (lanes 3 and 4, specific competitor of 67 nt) and tRNA (lane 5, unrelated competitor) were added at 1:10, 1: 50, and 1:50, respectively. Unlabeled ODN encoding anti-sense TB-RBP Y element ODN (lane 6, specific sequence competitor) was added at 1:5000. Sense TB-RBP motif ODN (lane 7, non-specific DNA competitor) was added at 5000-fold excess. (B) The gel mobility shift assay using 32P-labeled ligatin 3′UTR region and hippocampal cell lysates was performed as described in Fig. 2. The control RNA-protein complex is seen in lane 2. In lane 1, pGEM11(-) polylinker of 58 nt (non-specific competitor) was added with radio-labeled transcript. In lanes 3 and 4, unlabeled ODN encoding 13 nt of Y element anti-sense (specific competitor of 13 nt) at 1:5 (lane 3) or 21 nt of N-methyl-D-aspartate receptor subunit 1 (NMDAR1, non-specific competitor) at 1:5 (lane 4). (C) RNA-protein shift complex with TB-RBP Y element of α-CAMKII coding region. The gel mobility shift assay using 32P-labeled mRNA encoding the TBP Y element (17-nt coding and 12-nt polylinker) and hippocampal cell lysates was performed as described in Fig. 2. Lane 1 contains radiolabeled probe and cellular lysate. Unlabeled TB-RBP anti-ODN (lane 2, 13-nt sequence competitor) and TB-RBP sense ODN (lane 3, 13-nt competitor) were added at 1:5000. Arrowhead denotes specific RNA-protein complexes. 3696 W. L. Severt and others To address the question of whether the Y element per se can participate in forming a complex with TB-RBP, we used a radiolabeled transcript of 29-nt corresponding to the αCAMKII Y element (17-nt coding and 12-nt polylinker) in gel mobility shift assays. A single shift band (Fig. 4C, lane 1) was detected. Formation of this shift complex could be competed (>60%) by unlabeled anti-ODN corresponding to the antisense and sense Y elements (Fig. 4C, lanes 2 and 3, respectively). Collectively, these findings identify the TB-RBP as a common factor bound to ligatin and α-CAMKII mRNAs in hippocampal neurons. Antisense ODN to the TB-RBP motif alters mRNA distribution To evaluate the functional role of TB-RBP in mRNA localization, we used anti-ODN to inactivate the Y element of endogenous mRNA and thereby perturb in situ RNP formation. If the somato-dendritic transport of RNPs was disrupted, then the affected mRNAs would be expected to either (1) increase within the soma relative to the dendrites or (2) decrease within the soma due to selective degradation of the misrouted mRNA. In the first instance, the soma to dendrite ratio would therefore increase; in the second case; this ratio may decrease or remain the same depending upon the turnover rate of the mRNA within the dendrites. Non-derivatized, phosphodiester ODN complementary to the TB-RBP Y element (5′-AGCCCAGAGCTTG-3′) (1 µM) was administered daily to cultured hippocampal neurons; control cultures were given a sense-ODN of 5′-CAAGCTCTGGGCT-3′. After 3-4 days of treatment, the cellular distribution of ligatin mRNA was examined using FISH and confocal microscopy. The scanning parameters were optimized for the sense-ODN treated cells (controls) (Fig. 5A). Measurements from several neurons (n=7 total, sense-ODN, n=4; antisense-ODN, n=3) were pooled and the mean values calculated. A significant (P=0.015) mean increase of 32% in signal intensity was found in the somata of the anti-ODN treated neurons (Fig. 5B). To evaluate further the ODN-treated cells, we normalized the somata signals of anti-ODN versus sense-ODN neurons (Fig. 5C) by altering the scanning parameters. Under these conditions, the dendrite signal decreased 66% (P=0.001) at a distance of 40-50 µm (sense-ODN, n=11, anti-ODN, n=8) (Fig. 6); at a distance of 100-110 µm (sense-ODN, n=14, anti-ODN, n=8), the relative signal intensity decreased by 72% (P=0.001). Thus the ratio of soma to dendrite signal increased ~3.0-fold in the anti-ODN versus sense-ODN treated neurons. These data indicate that the increased signal within the soma was not due to a general increase in transcription but involved changes in mRNA distribution. These findings suggest that the Y element Fig. 5. Pseudocolor micrographs depicting subcellular distribution of ligatin mRNA in hippocampal neurons treated with anti-ODN and sense-ODNs to the Y element. (A-C) Distribution of ligatin mRNA. Neurons (15 days old) were treated for 4 days with 1 µM sense-ODN (A) or 1 µM anti-ODN (B) complementary to the TB-RBP Y element. Levels of ligatin mRNA were measured by FISH and confocal microscopy. In anti-ODN treated neurons, increased mRNA levels are found in the somata (**) and decreased levels within dendrites (*). (C) The signal intensity of the soma in B normalized to that in A. When the soma intensities are equalized, the dendritic signals in the anti-ODN treated neurons are significantly reduced (P= 0.001) (I). (D) No probe. (E and F) Distribution of α-CAMKII mRNA in sense-ODN (E) and anti-ODN (F) treated neurons. Confocal microscopic analysis of representative cells of 2 experiments is shown. Specific labeling of α-CAMKII mRNA is altered by anti-ODN within the somata (**) and dendrites (*). (G and H) Distribution of α-tubulin mRNA in sense-ODN (G) and antiODN (H) treated cells. Disrupted mRNA sorting in CNS neurons 3697 Fig. 6. Quantitation of ligatin and αCAMKII mRNA levels of ODN treated neurons described in Fig. 5. Left panel, representative images showing relative area (box) measured along dendrite. Right panel, mean amounts of specific signal within soma or dendrite. n=7 cells for each mRNA. Insert, pseudocolor intensity scale (1-256). Bar, 25 µm. is involved in delivery of ligatin mRNA within dendrites. To investigate the role of the Y element in a second mRNA known to be transported (Benson et al., 1992; Chicurel et al., 1993), we analyzed the effect of the Y element anti-ODN on the cellular distribution of αCAMKII mRNA. We found decreased hybridization signal throughout the neurons, especially within the dendritic arbors (compare Fig. 5E to F). The measurements from several neurons (n=7 total, sense-ODN, n=3; antisense-ODN, n=4) were pooled and the mean values calculated (Fig. 6). Significant (P=0.001) decreases in α-CAMKII mRNA signal intensities were found with a 58% decrease in the somata, 67% within dendrites at 30-40 µm distant from the somata, and 70% at 40-50 µm. The ratio of soma to dendrite signal for αCAMKII mRNA was increased 1.6-fold. To assess whether the anti-ODN depleted and/or altered the distribution of control mRNAs that do not contain the Y element, we examined α-tubulin mRNA which lacks Y and H elements and is restricted to the somata of hippocampal neurons. Administering oligonucleotides as described above, no effect on the subcellular distribution of α-tubulin mRNAs was found (Fig. 5G and H). This establishes that the anti-ODN was specific for localized mRNAs containing the Y element and suggests that inactivation of the Y element alters the subcellular distribution of the ligatin and α-CAMKII mRNAs. Antisense ODN to the TB-RBP binding motif suppresses protein expression The consequences of blocking mRNA transport by anti-ODN were examined using immunofluorescence to quantitate ligatin and α-CAMKII proteins in ODN treated cells. Following treatment with anti-ODNs, marked reductions in ligatin and αCAMKII protein levels were found (2 separate experiments, n=11 and 19, respectively) (Fig. 7B and D). In sense-ODN treated neurons (controls) (ligatin, n=15; CAMKII, n=25), expected amounts of these proteins were detected throughout the somato-dendritic compartment (Fig. 7A and C). Although the reduced levels of α-CAMKII protein reflected the decreased levels of its mRNA and ligatin protein levels were reduced, ligatin mRNA was increased (32%) under these conditions within the soma. These data indicate that anti-sense ODN binding to the Y element and/or mis-localization of ligatin mRNA may affect translation of the mRNA. Kinesin heavy chain motor is required for localization of select mRNAs To determine whether specific motor proteins could be involved with TB-RBP in mRNA movement, we used the procedure of Ferreira et al. (1992) to suppress kinesin heavy chain (KHC). Initially, an anti-ODN of 24-nt targeted to the initiator AUG region of KHC was used and the cellular distribution of ligatin mRNA and of α-CAMKII mRNAs were examined in neurons. Although no changes were seen in ligatin mRNA distribution (compare Fig. 8A and B) with anti-sense ODN treatment, there was a marked decrease in the cytosolic level of α-CAMKII mRNA throughout the somatic-dendritic compartment of these cells (compare Fig. 8C and D, E and F). Quantitation of the decline in α-CAMKII mRNA revealed significant decreases (36.6%, P=0.002) in somata. Measurements from four separate experiments were pooled and mean values obtained for somata (sense-ODN, n=9, antiODN, n=8) and within dendrites at 30-40 µm (sense-ODN, n=24, anti-ODN, n=20) and at 40-50 µm (sense-ODN, n=24, anti-ODN, n=20) from the soma (43.3% and 42.5%, P<0.001 and P<0.001, respectively). No significant decrease was found when neurons were treated with a KHC sense-ODN (Fig. 8C and E). To ascertain whether the dendritic localization of αCAMKII mRNAs was perturbed, we normalized the intensities of the somata of KHC sense-ODN and KHC anti-ODN treated cells from 2 separate experiments. Under these scanning conditions, significant decreases (27%, P=0.01 and 49%, P<0.001, respectively) in signal intensity were found at 80-90 µm (sense-ODN, n=11; anti-ODN, n=13) and at 120130 µm (sense-ODN, n=12, anti-ODN, n=11) from the somata. To control for specificity of the anti-KHC ODN effects, we targeted a second non-overlapping site located immediately downstream from our first sequence with an anti-ODN of 3698 W. L. Severt and others 25 nt. Again, α-CAMKII mRNA levels were reduced throughout the somato-dendritic compartment (data not shown), suggesting that the effect was due to suppression of KHC. Moreover, the levels and distribution of the somatarestricted mRNA encoding neuron specific enolase (NSE) were unaffected by the ODNs used to target KHC (Fig. 8G and H). To evaluate whether the KHC anti-ODN altered the accumulation of α-CAMKII mRNA directly, we performed FISH at 24 hours, a time before KHC protein levels decrease from the ODN treatment (50% suppression is seen at 36 hours). No significant decrease in the level of α-CAMKII mRNA was found (data not shown). Whereas several sense ODNs (encoding KHC and Y element) had no effect on α-CAMKII mRNA levels and specific anti-ODNs complementary to the Y element and to KHC led to decreased α-CAMKII mRNA levels, we propose that mis-localization of α-CAMKII mRNA, rather than transcriptional inhibition, leads to decreased levels of this transcript. Depletion of KHC from neurons after the specific antiODN treatments was assayed by using immunocytochemistry with a specific KHC antibody (SUK4) (sense-ODN, n=27; anti-ODN, n=44; 4 independent experiments) (Fig. 9A and B). Under identical conditions of staining and exposure, the specific immuno-labeling of neurons incubated with antisense ODN was reduced. The cytoarchitecture and viability of the ODN treated cells were not affected as previously reported by Ferreira et al. (1992). The partial suppression of KHC protein by KHC anti-ODN is consistent with previous reports of a 50-80% reduction in KHC (Ferreira et al., 1992) and is likely the reason we detect a residual localization of α-CAMKII mRNA. DISCUSSION Spatial heterogeneity of mRNAs in hippocampal neurons has been previously reported using isolated nerve terminal (synaptoneurosomes) preparations (Chicurel et al., 1993; Tiedge et al., 1991, 1993; Miyashiro et al., 1994) and in situ hybridization (Garner et al., 1988; Kleinman et al., 1990; Burgin et al., 1990; Steward and Wallace, 1995; Steward et al., 1998). These earlier studies lend support to the hypothesis that sorting of RNA plays a vital role in the establishment of axonal-dendritic polarity (Garner et al., 1988; Bruckenstein et al., 1990; Litman et al., 1993; Behar et al., 1995; Kanai and Hirokawa, 1995; Marsden et al., 1996) and of synapses (Steward, 1997; Steward and Banker, 1992; Miyashiro et al., 1994; Chircurel et al., 1993; Tiedge et al., 1993; Weiler et al., 1997; Steward et al., 1998). Transfection studies performed in primary cultured somatic cells, including neurons, have used reporter constructs and chimeric/deletion mutants to show that cis-acting element(s) residing within the 3′UTRs of mRNAs encode spatial information. Moreover, in vivo expression of reporter constructs in transgenic mice demonstrate that cis-acting elements within the 3′UTR of αCAMKII mRNA are sufficient for dendritic localization (Mayford et al., 1996b). In this study, we utilized neuronal cell cultures for analysis of the differential distribution of dendritically localized mRNAs to identify elements important for this process. We were able to alter the distribution of two localized mRNAs by inactivating a common cis-acting Fig. 7. Cellular distribution of ligatin and α-CAMKII proteins in neurons treated with ODNs complementary to the TB-RBP motif. Neurons are treated with ODN as described in Fig. 5. Following 4 days of ODN treatment, specific anti-ligatin staining is seen throughout the somato-dendritic compartment of sense-ODN (A) and anti-ODN (B) treated neurons. Specific staining of antiserum to the α subunit of CAMKII in Y element sense-ODN (C) treated neurons and anti-ODN (D) treated cells. n=2 independent experiments. Bar, 25 µm. element. Administration of anti-ODN complementary to the TB-RBP Y element disrupted RNA-protein interactions (Fig. 4) and sorting of α-CAMKII and ligatin mRNAs within dendrites (Fig. 5). Although the anti-ODN treatment is highly selective for its complementary mRNA (Phillips and Gyruko, 1997), sequence-independent toxic effects have been reported to occur with this experimental approach. In most instances where toxic effects are seen, phosphothioate derivatized ODN and/or high dosages of ODN have been used. Because the hippocampal neurons are grown in defined media in the absence of serum, we use unmodified ODN given daily at concentrations below toxic levels. During treatment, neurons maintained well-differentiated morphology and did not decrease in cell number. Moreover, the ODN-induced effect is reversed with removal of the ODN. Because the use of ODNs to perturb cell function necessitates numerous controls, we provide the following to show sequence specificity of the anti-ODN we have used: (1) the corresponding sense ODN has no effect (Figs 4, 5 and 7), (2) anti-ODN does not affect either the distribution or the amount of mRNAs lacking the Y element, indicating that cellular toxicity is not involved (Fig. 5), (3) several different ODN Disrupted mRNA sorting in CNS neurons 3699 give the same inhibitory effect for KHC, and (4) anti-ODN directed against the Y element disrupt RNA-protein complex formation in vitro (Fig.4). Moreover, we show that a reporter construct containing this Y element encodes spatial information sufficient to sort mRNA to dendrites in cultured cells (Fig. 1). Our findings support a general role for the TBRBP in sorting mRNAs within the somato-dendritic compartment. Additionally, our results suggest that TB-RBP does not mediate stability of these mRNAs. The specific RNA binding protein, TB-RBP, is a microtubule-associated protein that binds mRNA in a sequence specific manner (Han et al., 1995; Kwon and Hecht, 1993). In the mammalian testis, TB-RBP appears to have the dual functions of translational suppression and of intracellular and intercellular mRNA transport (Morales et al., 1998). In brain, the TB-RBP is primarily expressed in neurons (Wu et al., 1999) and forms 11.5S RNPs with untranslated BC1 and BC200 RNAs (Muramatsu et al., 1998; Kobayashi et al., 1998) which are localized within dendrites. Here we show that the TB-RBP binds localized mRNAs encoding ligatin and α-CAMKII and that disruption of these interactions alters somato-dendritic sorting. We suspect that the TB-RBP is involved in tethering specific mRNAs to microtubules and hence permitting their movement. Moreover, we propose the TB-RBP in association with other proteins suppress translation of bound mRNAs in an analogous manner to its role in testis. In vitro and in vivo assays have demonstrated that these proteins, the transitional endoplasmic reticulum ATPase, Trax and a cytoskeletal gamma actin, interact with TB-RBP in diverse tissues including brain (Wu et al., 1999). It is likely that posttranslational regulation of TB-RBP by synaptic activity may provide a mechanistic link important in regulating both spatial and temporal expression of specific proteins several hundreds of microns from the nucleus. Many studies have focused on the localization of mRNA at synaptic areas because of its implied role in local synthesis of protein. In particular, preparations of isolated nerve terminals (synaptoneurosomes) have been used to study neurotransmitter effects on local translation (Weiler et al., 1997). In synaptoneurosomal preparations, functional polyribosomes form in vitro in response to K+ depolarization and to glutamate (Weiler and Greenough, 1993). Activation of the metabotropic subtype of glutamatergic receptors initiates protein synthesis, while activation of the ionotropic NMDA receptor subtype attenuates it. These findings indicate that synaptic activity regulates translation of localized mRNAs locally and that distinct signaling cascades may be involved. In our studies, we directly tested whether local translation is a prerequisite for gene expression by disrupting mRNA delivery. We studied two localized mRNAs, αCAMKII and ligatin, that are post-transcriptionally regulated in models of glutamate-induced injury (Liang and Jones, 1997; Murray et al., 1995; Panchision et al., 1995; Jakoi et al., 1992). Using immunocytochemistry, we show that preventing TB-RBP binding to its Y element by anti-ODN decreases the amounts of α-CAMKII and ligatin proteins we can detect within hippocampal neurons. Although reduced levels of α-CAMKII and ligatin proteins in dendrites reflect directly the ODN-induced reduction in the encoding mRNAs, suppression of translation within the somata may result from either RNA mis-sorting or DNA binding to the cis-acting TB-RBP Y element on the α-CAMKII and ligatin mRNAs. Our data can not discriminate between these possibilities. Collectively, our findings implicate translational control as a critical regulatory site governing local expression of proteins important for synaptic activity. The movement of numerous mRNAs within cells including neurons has been documented. Both microfilament and microtubule mediated movement have been implicated. However, only the microfilament associated motor protein, Myo04, has been identified. This protein localizes ASH1 mRNA in budding Saccharomyces cerevisiae (Long et al., 1997). In neurons, microtubule associated movement of mRNA within dendrites may utilize either kinesin and/or dynein motors. Several studies have reported microtubule mediated movement of mRNAs into dendrites at rates consistent with kinesin mediated movement (Davis et al., 1987; Knowles et al., 1996; Muslimov et al., 1997; Wallace et al., 1998). Because KHC-null (knock out) animals are not viable due to neuronal death (Saxton et al., 1991), and no pharmacological agent is available to selectively inhibit movement by individual motor proteins, we have suppressed the function of a specific motor protein with an anti-ODN. Earlier studies by Ferreira et al. (1992) showed that after 36 hours of anti-ODN treatment, KHC protein levels are reduced 50% and the movement of organelles within the axon of newly plated hippocampal neurons is perturbed in a reversible manner. To perturb movement of mRNAs with an anti-ODN to KHC, we have used well-differentiated neonatal hippocampal neurons in culture. Using FISH and confocal microscopy, we have demonstrated that KHC anti-ODN has no effect on levels and distribution of α-CAMKII mRNA levels at 24 hours. However, at 72 hours, α-CAMKII mRNA levels were decreased throughout the somato-dendritic compartments, when the level of kinesin heavy chain protein is reduced. We conclude that this general decline in αCAMKII mRNA by KHC anti-ODN was a specific effect because (1) the corresponding sense-ODN had no effect, (2) two different ODNs complementary to KHC had the same effect, making suppression of a protein different from KHC unlikely, (3) no reduction in α-CAMKII mRNA was seen at 24 hours a time when KHC protein was not yet affected by ODN treatment, (4) ODN against the Y element (a nonrelated sequence to the KHC anti-ODN) disrupted αCAMKII mRNA sorting and also reduced its cellular levels, and (5) the stability of other mRNAs such as ligatin and neuron specific enolase was not affected, indicating a lack of cellular toxicity. Collectively, these findings indicate that ODN treatment did not alter the net accumulation of αCAMKII mRNAs. Our data indicate that the motor protein KHC plays a necessary role in the localization of at least one dendritically targeted mRNA although the roles of other motor proteins cannot be excluded. In situ hybridization and biochemical fractionation were used previously in hippocampal neurons to localize mRNAs (Chicurel et al., 1993; Tiedge et al., 1991, 1993; Miyashiro et al., 1994; Crino and Eberwine, 1996; Garner et al., 1988; Kleinman et al., 1990; Burgin et al., 1990; Steward and Wallace, 1995; Comery et al., 1997; Weiler et al., 1997; Steward et al., 1998). Here we alter the distribution of two transported mRNAs by inhibiting the formation of a RNA- 3700 W. L. Severt and others Fig. 8. Effect of KHC anti-ODN on the relative distributions of somato-dendritic mRNAs in hippocampal neurons. Neurons are given ODNs (50 µM initially then 25 µM every 12 hours for 3 days) as described. For FISH, cells are hybridized with biotinylated probe complementary to the mRNA of ligatin (A, B), α-CAMKII (C-F), and NSE (G, H). Bound probe is visualized with Texas red-ultra avidin conjugate. (A,C,E,G) FISH/confocal micrographs of neurons treated with KHC sense-ODN show somato-dendritic distribution of mRNAs. (B,D,F,H) Neurons treated with KHC anti-ODN show a marked decrease in labeling for α-CAMKII mRNAs (D and F). The regional distribution of ligatin (B) and NSE (H) mRNAs are not affected. * Dendrite; ** soma. Relative intensity scale as in Fig. 5. Bars: 50 µm (A-D,G,H); 20 µm (E and F). protein complex and by depleting a specific motor protein. We conclude that TB-RBP and KHC are important contributors in the localization of mRNA within the dendritic compartment of hippocampal neurons and implicate microtubules as the transit route. Our data and the interactions of TB-RBP with the transitional endoplasmic reticulum ATPase, the ATPase involved in intracellular transport and vesicle fusion (Wu et al., 1999), suggest structural heterogeneity of RNA-protein complexes involved in the regional localization of mRNA in hippocampal neurons. Additionally, we show that local translation of targeted mRNA is critical for expression of the cognate protein within post-synaptic areas. Our working hypothesis is that the formation of RNA-protein complexes important for mRNA sorting may be governed by synaptic activity and thus provides a mechanistic link by which neural differentiation and plasticity can be regulated locally within dendrites and post-synaptic areas of CNS neurons. The authors thank Drs J. Nevins, J. DeGregori, E. Ellis, J. Feher, E. Raff, L. Jakoi and M. Sheetz for editorial comments in the preparation of this manuscript. We thank Drs G. Leone, J. DeGregori, and J. Nevins (HHMI Duke University Medical Center, NC) for preparation of the recombinant adenoviruses. Appreciation is expressed to Dr Okatani (HHMI Duke University) for the pAdCMV, Dr Cowan (New York Medical Center, New York) for the pBR322kα1 encoding α-tubulin and Dr Bigbee (Virginia Commonwealth University, VA) for anti-intermediate filament antibody. We thank C. Gerwin for technical assistance. This work was supported by A. D. Williams grants 6-46917 (T.B.) and 6-46576 (E.R.J.) and AHAWilliam Randolph Hearst award 92009590 (E.R.J.), by NIH Center grant and Jacob Javits award (R.J.D.) and by the Sophie and Nathan Gumenick Neuroscience Research Endowment, and by NICHD grant HD28832 (N.B.H.). Fig. 9. Regional distribution of KHC protein in hippocampal neurons treated with KHC sense-ODN (A) and KHC antiODN (B). ODNs were administered as described in Fig. 8. After 72 hours, neurons were stained with specific antibody to KHC (1:10). Results are representative of 4 experiments. * dendrite; ** soma. Bar, 25 µm. Disrupted mRNA sorting in CNS neurons 3701 REFERENCES Antic, D. and Keene, J. D. (1998). Messenger ribonucleoprotein complexes containing human ELAV proteins: interactions with cytoskeleton and translational apparatus. J. Cell Sci. 111, 183-197. Baas, P. W., Deitch, J. S., Black, M. M. and Banker, G. A. (1988). Polarity orientation of microtubules in hippocampal neurons: uniformity in the axon and non-uniformity in the dendrite. Proc. Nat. Acad. Sci. USA 85, 83358339. Bassell, G. J., Oleynikov, Y. and Singer, R. H. (1999). The travels of mRNAs through all cells large and small. FASEB J. 13, 447-454. Behar, L., Marx, R., Sadot, E., Barg, J. and Ginzburg, I. (1995). Cisacting and trans-acting proteins are involved in tau mRNA targeting into neurites of differentiating neuronal cells. Int. J. Neurosci. 13, 113127. Benson, D. I., Gall, C. M. and Isackson, P. J. (1992). Dendritic localization of type II calcium calmodulin-dependent protein kinase mRNA in normal and reinnervated rat hippocampus. Neuroscience 46, 851-857. Bian, F., Chu, T., Schilling, K. and Oberick, J. (1996). Differential mRNA transport and the regulation of protein synthesis: selective sensitivity of purkinje cell dendritic mRNAs to translational inhibition. Mol. Cell. Neurosci. 7, 116-133. Bruckenstein, D. A., Lein, P. J., Higgins, D. and Fremeau, R. T., Jr. (1990). Distinct spatial localization of mRNAs in cultured sympathetic neurons. Neuron 5, 809-819. Burgin, K. E., Waxham, M. N., Rickling, S., Westgate, S. A., Mobley, W. C. and Kelly, P. T. (1990). In situ hybridization histochemistry of Ca+2/ calmodulin-dependent protein kinase in developing rat brain. J. Neurosci. 10, 1788-1798. Caillaud, C., Akli, S., Vigne, E., Kouklakoff, A., Perricaudet, M., Poenaru, L., Kahn, A. and Berwald-Netter, Y. (1993). Adenoviral vector as a gene delivery system into cultured rat neuronal and glial cells. European J. Neurosci. 5, 1287-1291. Chicurel, M., Terrain, D. M. and Potter, H. (1993). mRNA at the synapse: analysis of a synaptosomal preparation enriched in hippocampal dendritic spines. J. Neurosci. 13, 4054-4063. Comery, T. A., Harris, J. B., Willems, P. J., Oostra, B. A., Irwin, S. A., Weiler, I. J. and Greenough, W. T. (1997). Abnormal dendritic spines in fragile X knockout mice: maturation and pruning deficits. Proc. Nat. Acad. Sci. USA 94, 5401-5404. Crino, P. B. and Eberwine, J. (1996). Molecular characterization of the dendritic growth cone: regulated mRNA transport and local protein synthesis. Neuron 17, 1173-1187. Davis, L., Burger, B., Banker, G. A. and Steward, O. (1987). Dendritic transport: quantitative analysis of the time course of somato-dendritic transport of newly synthesized RNA. J. Neurosci. 10, 3056-3068. Ferreira, A., Nicolas, J., Vale, R. D., Banker, G. and Kosik, K. S. (1992). Suppression of kinesin expression in cultured hippocampal neurons using antisense oligonucleotides. J. Cell Biol. 117, 595-606. Gao, F. B. and Keene, J. D. (1996). Nel-N1/Hel-N2 proteins are bound to poly(A)+ mRNA in granular RNP structures and are implicated in neuronal differentiation. J. Cell Sci. 109, 579-589. Garner, C. C., Tucker, M. P. and Matus, A. (1988). Selective localization of messenger RNA for cytoskeletal protein MAP2 in dendrites. Science 336, 674-677. Gillis, P. and Malter, J. S. (1991). The adenosine-uridine binding factor recognizes the AU rich elements of cytokine, lymphokine and oncogene mRNAs. J. Biol. Chem. 266, 3176-3177. Han, J. R., Yiu, G. K. and Hecht, N. B. (1995). Testis/brain RNA binding protein attaches translationally repressed and transported mRNAs to microtubules. Proc. Nat. Acad. Sci. USA 92, 9550-9554. Han, J. R., Gu, W. and Hecht, N. B. (1995). Testis-brain RNA binding protein, a testicular translational regulatory RNA-binding protein, is present in the brain and binds to the 3′ untranslated regions of transported brain mRNAs. Biol. Reprod. 53, 707-717. Hecht, N. B. (1998). Molecular mechanisms of male germ cell differentiation. BioEssays 20, 555-561. Hirokawa, N. (1998). Kinesin and dynein superfamily proteins and the mechanism of organelle transport. Science 279, 519-526. Jakoi, E. R., Ross, P. E., Ting-Beall, H. P., Kaufman, B. and Vanaman, T. C. (1987). Ligatin: a peripheral membrane protein with covalently bound palmitic acid. J. Biol. Chem. 262, 1300-1304. Jakoi, E. R., Sombati, S., Gerwin, C. and DeLorenzo, R. J. (1992). Excitatory amino acid receptor produces a selective and long-lasting modulation of gene expression in hippocampal neurons. Brain Res. 582, 282-290. Jakoi, E. R. and Severt, W. L. (1999). Disruption of RNP formation and sorting in hippocampal neurons. Meth. Enzymol. 313, 456-466. Kanai, Y. and Hirokawa, N. (1995). Sorting mechanisms of tau and MAP2 in neurons: suppressed axonal transit of MAP2 and locally regulated microtubule binding. Neuron 14, 421-432. Kleinman, R., Banker, G. and Steward, O. (1990). Differential subcellular localization of particular mRNAs in hippocampal neurons in culture. Neuron 5, 821-830. Knowles, R. B., Sabry, J. H., Martone, M. E., Deerinck, T. J., Ellisman, M. H., Bassell, G. J. and Kosik, K. S. (1996). Translocation of RNA in granules in living neurons. J. Neurosci. 16, 7812-7820. Kobayashi, S., Takashima, A. and Anzai, K. (1998). The dendritic translocation of Translin protein in the form of BC1 RNA protein particles in developing rat hippocampal neurons in primary culture. Biochem. Biophys. Res. Commun. 253, 448-453. Kwon, Y. K. and Hecht, N. B. (1991). Cytoplasmic protein binding to highly conserved sequences in the 3′ untranslated region of mouse protamine 2 mRNA, a translationally regulated transcript of male germ cells. Proc. Nat. Acad. Sci. USA 88, 3584-3588. Kwon, Y. K. and Hecht, N. B. (1993). Binding of a phosphoprotein to the 3′ untranslated region of the mouse protamine 2 mRNA temporally represses its translation. Mol. Cell. Biol. 13, 6547-6567. Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685. Liang, F. and Jones, E. G. (1997). Differential and time-dependent changes in gene expression for type II calcium/calmodulin-dependent protein kinase, 67 kDa glutamic acid decarboxylase, and glutamate receptor subunits in tetanus toxin-induced focal epilepsy. J. Neurosci. 17, 21682180. Litman, P., Barg, J., Rindozoonski, L. and Ginzburg, I. (1993). Subcellular localization of tau mRNA in differentiating neuronal cell cultures: implications for neuronal polarity. Neuron 10, 627-638. Long, R. M., Singer, R. H., Meng, X., Gonzales, I., Nasmyth, K. and Jansen, R.-P. (1997). Mating type switching in yeast controlled by asymmetric localization of ASH1 mRNA. Science 277, 383-387. MacDonald, P. M. and Kerr, K. (1998). Mutational analysis of an RNA recognition element that mediates localization of bicoid mRNA. Mol. Cell Biol. 18, 3788-3795. Marsden, K., Ferralli, J., Botteri, F. and Matus, A. (1996). Transgenic expression of embryonic MAP2 in adult mouse brain: implications for neuronal polarity. J. Neurosci. 16, 3265-3273. Mayford, M., Bach, M. E., Huang, Y. Y., Wang, L. Hawkins, R. D. and Kandel E. R. (1996a). Control of memory formation through regulated expression of α-CAMKII transgene. Science 274, 1678-1683. Mayford, M., Baranes, D., Podsypanina, K. and Kandel, E. R. (1996b). The 3′-untranslated region of CAMKII alpha is a cis-acting signal for the localization and translation of mRNA in dendrites. Proc. Nat. Acad. Sci. USA 93, 13250-13255. Miyashiro, K., Dichter, M. and Eberwine, J. (1994). On the nature and differential distribution of mRNAs in hippocampal neurites: implications for neuronal functioning. Proc. Nat. Acad. Sci. USA 91, 10800-10804. Mohr, E. (1999). Subcellular RNA compartmentalization. Prog. Neurobiol. 57, 507-525. Morales, C. R., Wu, X. Q. and Hecht, N. B. (1998). The DNA/RNA-binding protein, TB-RBP, moves from the nucleus to the cytoplasm and through intercellular bridges in male germ cells. Dev. Biol. 200, 113-123. Moriyoshi, K., Richards, L. J., Aazawa, C., O’Leay, D. D. M. and Nakanishi, S. (1996). Labeling of neural cells using adenovirus gene transfer of membrane-targeted GFP. Neuron 16, 255-260. Murray, K. D., Gall, C. M., Benson, D. L., Jones, E. G. and Isackson, P. J. (1995). Decreased expression of the α subunit of calcium/calmodulin dependent kinase type II mRNA in the adult rat CNS following recurrent limbic seizures. Brain Res. Mol. Brain Res. 32, 221-232. Muramatsu, T., Ohmae, A. and Anzai, K. (1998). BC1 RNA protein particles in mouse brain contain two y-, h-element-binding proteins, Translin and a 37 kDa protein. Biochem. Biophys. Res. Commun. 247, 7-11. Muslimov, I. A., Sanit, E., Homel, P., Perini, S., Higgins, D. and Tiedge, H. (1997). RNA transport in dendrites: a cis-acting targeting element is contained within neuronal BC1 RNA. J. Neurosci. 17, 4722-4733. Nevins, J. R., DeGregori, J., Jakoi, L. and Leone, G. (1997). Functional analysis of E2F. Meth. Enzymol. 283, 205-219. Panchision, D. M., Gerwin, C. M., DeLorenzo, R. J. and Jakoi, E. R. 3702 W. L. Severt and others (1995). Glutamate receptor activation regulates mRNA at both transcriptional and post-transcriptional levels. J. Neurochem. 65, 969-977. Phillips, M. I. and Gyurko, R. (1997). Antisense oligonucleotides: new tools for physiology. NIPS 12, 99-105. Saxton, W. M., Hocks, M. E., Goldstein, L. S. B. and Raff, E. C. (1991). Kinesin heavy chain is essential for viability and neuromuscular functions in Drosophila, but mutants show no defects in mitosis. Cell 64, 1093-1102. Steward, O. and Banker, G. A. (1992). Getting the message from the gene to the synapse: sorting and intracellular transport of RNA in neurons. Trends Neurosci. 15, 180-186. Steward, O. and Wallace, C. S. (1995). mRNA distribution within dendrites: relationship to afferent innervation. J. Neurobiol. 26, 447-449. Steward, O. (1997). mRNA localization in neurons: a multipurpose mechanism? Neuron 18, 9-12. Steward, O., Wallace, C. S., Lyford, G. L. and Worley, P. F. (1998). Synaptic activation causes the mRNA for the IEG ARC to localize selectively near activated postsynaptic sites on dendrites. Neuron 21, 741-751. Tiedge, H., Fremeau, Jr., R. T, Weinstock, P. H., Arancio, O. and Brosius, J. (1991). Dendritic localization of neural BC1 RNA. Proc. Nat. Acad. Sci. USA 84, 7773-7777. Tiedge, H., Chen, W. and Brosius, J. (1993). Primary structure, neuralspecific expression, and dendritic location of human BC200 RNA. J. Neurosci. 13, 2382-2390. Wallace, C. S., Lyford, G. L., Worley, P. F. and Steward, O. (1998). Differential intracellular sorting of immediate early gene mRNAs depends on signals in the mRNA sequence. J. Neurosci. 18, 26-35. Weiler, I. J. and Greenough, W. T. (1993). Metabotropic glutamate receptors trigger postsynaptic protein synthesis. Proc. Nat. Acad. Sci. USA 90, 71687171. Weiler, I. J., Irwin, S. A., Klintsova, A. Y., Spencer, C. M., Brazelton, A. D., Miyashiro, K., Comery, T. A., Patel, B., Eberwine, J. and Greenough, W. T. (1997). Fragile X mental retardation protein is translated near synapses in response to neurotransmitter activation. Proc. Nat. Acad. Sci. USA 94, 5395-5400. Wu, X. Q., Gu, W., Meng, X. and Hecht, N. B. (1997). The RNA binding protein, TB-RBP, is the mouse homologue of translin, a recombination protein associated with chromosomal translocation. Proc. Nat. Acad. Sci. USA 94, 5640-5645. Wu, X. Q., Petrusz, P. and Hecht, N. B. (1999). Testis-brain RNA binding protein (Translin) is primarily expressed in neurons of the mouse brain. Brain Res. 819, 174-178.