Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
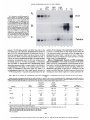
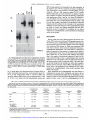
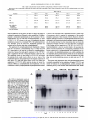
[CANCER RESEARCH 53. 2490-2497. June I. 19931 Cyclophosphamide Modulates Rat Hepatic Cytochrome P450 2C11 and Steroid 5tt-Reductase Activity and Messenger RNA Levels through the Combined Action of Acrolein and Phosphoramide Mustard1 Thomas K. H. Chang and David J. Waxman2 Department ¡ifBioluaical Chemistry and Molecular Phunnaciilogy ana Dana-Furher Cancer Institute, Hanard ABSTRACT Cyclophosphamide treatment of adult male rats leads to sustained de creases in several liver microsomal cytochrome P450 (CYP) activities, including CYP2C11-catalyzed Cyclophosphamide activation, via a process that is associated with a feminization of the overall pattern of liver enzyme expression (G. A. LeBlanc and D. J. Waxman, Cancer Res., 50: 5720-5726, 1990). The present study compares the effects of Cyclophosphamide and its isomerie analogue ifosphamide on the gender-dependent expression of hepatic CYP 2C11 and steroid Sa-reductase in adult male rats and also examines the role of the Cyclophosphamide metabolites acrolein and phosphoramide mustard in feminizing the expression of these liver enzymes. Ifosphamide (a) suppressed the male-specific CYP 2C11 mRNA and CYP 2C11-catalyzed liver microsomal testosterone 2a-hydroxylation and Cyclo phosphamide and ifosphamide 4-hydroxylation and (b) elevated the fe male-dominant liver enzyme steroid 5a-reducta.se and its mRNA 7-9 days after drug treatment, both occurring in a manner similar to that of cyclophosphamide, but requiring a 50% higher dose (180 mg/kg, single i.p. injection) to achieve these effects. This pattern of response could not be achieved by treatment of rats with acrolein or with Cyclophosphamide analogues that decompose to acrolein without formation of phosphoramiilf mustard. In contrast, phosphoramide mustard treatment (100 mg/ kg) did modulate microsomal CYP 2C11 and steroid 5a-reductase activ ities. Treatment with a lower dose (50 mg/kg) of phosphoramide mustard or with the acrolein precursor 4-hydroperoxydechlorocyclophosphamide (200 mg/kg) alone did not affect liver enzyme expression, whereas the combination of these agents produced an overall pattern of response that was similar to that conferred by Cyclophosphamide. These studies estab lish that ifosphamide is less potent than Cyclophosphamide in modulating the pattern of cytochrome P450 and steroid 5<x-reductase expression and that phosphoramide mustard is responsible for the modulation of liver enzyme expression by Cyclophosphamide, with acrolein potentiating the modulating activity of the mustard. Medical Schuol. Bustini. Massachusetts 02115 mustard possesses DNA-alkylating activity and is generally consid ered to be the therapeutically significant, cytotoxic metabolite of Cyclophosphamide (1,3). Acrolein. which is an electrophilic aldehyde, lacks antitumor activity (4. 5) but is highly reactive and binds covalently to proteins, including cytochrome P450 (6-9) and NADPHcytochrome P450 reducÃ-ase(10), leading to enzyme inactivation. The Cyclophosphamide metabolites 4-hydroxycyclophosphamide and al dophosphamide can also be metabolized by aldehyde dehydrogenases to yield inactive species (11, 12). Ifosphamide is an isomer of Cyclophosphamide (Fig. 1) that exhib its important quantitative differences in pharmacokinetics and metab olism, compared to Cyclophosphamide (13). Although Cyclophospha mide and ifosphamide are both activated by hepatic cytochrome P450 enzymes to form a 4-hydroxy metabolite that subsequently decom poses to yield acrolein plus a mustard derivative (isophosphoramide mustard in the case of ifosphamide), ¡fosphamide is activated at a lower rate than Cyclophosphamide (14). This appears to reflect both the lower catalytic efficiency for ifosphamide activation exhibited by individual cytochromes P450 and changes in the spectrum of cyto chrome P450 enzymes that can contribute to drug activation (15). In addition, a quantitatively important pathway of ifosphamide metabo lism is side-chain /V-dechloroethylation (16), which leads to the for mation of the therapeutically inactive but neurotoxic metabolite chloroacetaldehyde (17). In contrast. Cyclophosphamide is not subject to substantial A'-dechloroethylation (1, 18). Whereas Cyclophosphamide is known to interact with rat hepatic cytochromes P450 via a multi plicity of mechanisms (19). the potential effects of ifosphamide on hepatic cytochrome P450 enzyme profiles are not known. The precise mechanisms by which Cyclophosphamide alters rat hepatic cytochrome P450 protein levels and enzyme activities remain to be clarified. Early studies concluded that the effects were due to INTRODUCTION denaturation of cytochrome P450 by the Cyclophosphamide metabo lite acrolein (6. 7). In support of this proposal, sulfhydryl-containing Cyclophosphamide is a widely used anticancer alkylating agent prodrug that is bioactivated by the liver CYP3 monooxygenase system compounds prevent the decrease in cytochrome P450 enzyme activ ( 1). Three specific cytochrome P450 enzymes, forms CYP 2BI (phé ities observed 4 days after treatment of rats with cyclophosphamide nobarbital inducible). CYP 2C6 (constitutively expressed and gender (20). However, sulfhydryl compounds do not block the decreases independent), and CYP 2C11 (constitutively expressed and male spe observed 7 days after cyclophosphamide treatment (21 ). Rather, these cific),4 have been identified as major catalysts of Cyclophosphamide chronic cyclophosphamide-dependent decreases in liver cytochrome activation in rat liver (2). The primary metabolite formed by these P450 enzyme activities are associated with a feminization of liver enzymes. 4-hydroxycyclophosphamide. equilibrates with the ringenzyme profiles. Thus, cyclophosphamide suppresses the male-spe opened aldophosphamide. which undergoes spontaneous decomposi cific CYP 2A2, 2C11, and 3A2, while it induces the female-predom tion to yield phosphoramide mustard and acrolein ( 1). Phosphoramide inant enzymes CYP 2AI and steroid 5a-reductase (9). These effects of cyclophosphamide are similar to those produced by cisplatin, which Received 12/21/92: accepted 3/24/93. feminizes the pattern of liver cytochrome P450 (22. 23) as well as The costs of publication of this article were defrayed in part by the payment of page glutathione 5-transferase enzyme expression (24). Thus, the effects of charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. cyclophosphamide are complex and are not simply the result of a 1 Supported in part by Grant CA-49248 from the NIH (D. J. W.). T. K. H. C. was direct inactivation of cytochromes P450 by acrolein. In principle, supported by a Canadian Association of Gastroenterology/Janssen Research Fellowship (1991-1992) and a Canadian Liver Foundation Research Fellowship (1992-1993). these major effects of cyclophosphamide on liver cytochrome P450 - To whom requests for reprints should he addressed, at Dana-Farber Cancer Institute. profiles could be mediated by either acrolein or phosphoramide mus Room JF-525. 44 Binney Street. Boston. MA 02115. 1The abbreviations used are: CYP. cytochrome P450: HPD-cyclophosphamide. tard, both of which are reactive electrophilic molecules. Acrolein has 4-hydroperoxydechlorocyclophosphamide; deCl-cyclophosphamide. bisfethyllaminocybeen identified as the primary mediator of the urotoxicity that accom clophosphamide. panies the clinical use of cyclophosphamide and ifosphamide (25, 26), 4 Individual liver CYP forms are designated according to the systematic nomenclature (48). whereas phosphoramide mustard appears to be responsible for the 2490 Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research. ACROI.KIN. PHOSPHOR AMIDI CICH2CH2x V Ml STARI). vr-C P CH,CH P XN acHfttf O CICH2CH2 P u V OH CH,CH2CI chromatographed on silica gel thin layer chromatography plates developed with dichloromethane/acetone (4/1) followed by chloroform/ethyl acetate/ absolute ethanol (4/1/0.7). Metabolites were localized by autoradiography and quantitated by liquid scintillation counting. Microsomal steroid 5«-reductase activity was determined by the reduction of |4-l4C|testoslerone to 5a-[4-'4C]dihydrolestosterone in the same assay. Microsomal cyclophosphamide 4-hydroxylase v H CH,CH,^ IFA deCI-CPA ACROLEIN (CM), il'osphamide (//vl). phosphoramide mustard (PM ), acrolein. and the acrolein precursors HPD-cyclophosphamide and deCTcyclophosphamide (di'Cl-CPA). and ifosphamide 4-hydroxy- lase activities were determined by a fluorescence assay (31 ). Each incubation mixture contained 100 nisi potassium phosphate. pH 7.4. 0.1 HIMEDTA. 5 msi semicarba/ide HC1. 0.5 msi cyclophosphamide or ifosphamide. 100 ug mi crosomal protein, and I nisi NADPH. in a total volume of 2(X)ul. The reaction was incubated for 60 min at 37°Cand then stopped by the addition of 80 ul XP. CHjCH/ Fig. 1. Structures of cyclophosphumide 1 M'KI -SSION laheled testosterone, and 1 m.wNADPH. in a total volume of 2(X)ul. Reactions were incubated for IO min at 37°C.extracted with ethyl acetate, and then HPD-CPA CPA (il.SI. activity was determined as described previously (30). Incubation mixtures contained 100 msi 4-(2-hydroxyethyl)-l-pipera/.ineethanesullbnic acid (HEPES). (pH 7.4). O.I msi EDTA. 30 ug microsomal protein. 50 UM I4C- CICH2CH2/ CH,CH,' \ J ' \M> IUPD-CPA ) ice-cold 5.5% zinc sulfate solution, followed by 80 pi saturated barium hy droxide and 40 ul 0.01 si HCI. After centrifugation. 300 ul of the supernatant were derivati/ed with a IftO-ul solution containing aminophenol (6 mg/ml) and hydroxvlamine hvdrochloride (ft mg/ml) in I si HCI. This mixture was heated at 9()°Cfor 20 min and then cooled to room temperature before the fluores ovarian toxicity of cyclophosphamide (27) and has also been impli cated as an important cardiotoxic metabolite of cyclophosphamide (28). The present study compares the effects of/« vivo ifosphamide and cyclophosphamide treatment of adult male rats on hepatic CYP 2CII and steroid 5«-reduetase enzyme activities and mRNA levels and examines the role of the metabolites acrolein and phosphoramide mustard. /'/; vivo, in the femini/.ation of these hepatic drug- and ste cence was measured (350-nm excitation wavelength and 515-nm emission wavelength). 4-Hydroperoxyifosphamide was used as a standard for acrolein (32) and was incubated as described above hut with heat-inactivated roid-metabolizing enzymes. The results obtained establish that (a) ifosphamide is less potent than cyclophosphamide in suppressing CYP 2CI1 and elevating steroid 5ct-reductase mRNA levels and enzyme activities, including CYP 2C11-catalyzed cyclophosphamide and if osphamide activation, (h) modulation of these gender-dependent he patic mRNA levels and enzyme activities by cyclophosphamide is due to the action of phosphoramide mustard, and (<•) acrolein potentiates NEN. Boston. MA) (29). The nucleotide sequences for the oligonucleotide probes ON-5 (CYP 2CII). ON-48 (steroid 5«-reductase). and ON-50 (tt-tubulin) are presented elsewhere (29. 33). microsomes and in the absence of NADPH. Oligonudeotide Probes. Gene-specific oligonucleotide otide kinase. and subsequently purified on NENSORB 20 columns (DuPont- Northern Blot Analysis. Total liver RNA samples were isolated from frozen liver tissue and then electrophoresed in \'/< agarose/O.ftft si formalde hyde gels as detailed elsewhere (29). The RNA was transferred to nylon (liters (Genescreen; DuPont-NEN) and then UV cross-linked. Prehybridization and hybridization for Northern blot analysis were carried out at a temperature (4()°Cor 45°C)and tbrmamide concentration (0-25%, v/v) suitable for each the modulating activity of the mustard. MATERIALS probes were syn- thesi/.cd on an Applied Biosystems DNA synthesizer, purified by high perfor mance liquid chromalography. ^'-labeled with [-y-12P]ATP by T4 polynucle- oligonucleotide probe (29. 33). The nylon fillers were then washed and ex posed to Kodak XAR-5 film, w ith intensifying screens, al -8(FC for 2-6 days. Each blot was probed with rat «-tuhulinoligonucleotide probe ON-50 to assess AND METHODS RNA loading consistency and integrity. Serum Testosterone Assay. Serum testosterone concentration was meas ured by solid-phase '-*! radioimmunoassay with the Coat-A-Count total tes Chemicals. Cyclophosphamide. ifosphamide. and phosphoramide mustard Iphosphoramidic acid yV.jV-bis(2-chloroethyl). cyclohexylumine salt] were ob tained from the Drug Synthesis and Chemistry Branch, National Cancer In stitute (Bethesda. MDl. HPD-cyclophosphamide and 4-hydroperoxyitospha- tosterone kit (Diagnostic Products Corp.. Los Angeles. CA). mide were kindly provided by Dr. J. Pohl (ASTA Pharma. Bielefeld. Germany). Dr. J. Hilton (Johns Hopkins Oncology Center. Baltimore. MD) provided deCl-cyclophosphamide. Acrolein and [4-l4C]testosterone were purchased RESULTS from Aldrich Chemical Co. (Milwaukee. WI) and Amershum Corp. (Arlington Heights. IL), respectively. Animal Treatments. Adult male Fischer 344 rats (190-200 g. 8-9 weeks old: Taconic Farms. Germamown. NY) were treated with single i.p. injections of cyclophosphamide. ifosphamide. phosphoramide mustard, acrolein. or HPD-cyclophosphamide. at the doses indicated in the text. deCl-cyclophosphamide was administered i.p. at a dose of 2<X)nig/kg in a previous experiment conducted by Dr. G. F. Weher of this laboratory. Control rats were given i.p. injections of the vehicle (0.9% NaCI solution). On the day of treatment, drugs were dissolved in the vehicle and administered to rats (4 ml/kg body weight) immediately thereafter. At 7 or 9 days after treatment, as specified in the text for each experiment, rats were killed by cervical dislocation following brief asphyxiation under CO2. Livers were quickly excised, washed with ice-cold 1.15% KC1 solution, cut into small pieces, frozen in liquid nitrogen, and then stored at -8()°Cuntil used for microsomal preparation or RNA isolation. Blood was collected by cardiac puncture and was allowed to clot at 4°C.Serum was prepared by centrifugaron and then stored at -20CC until use. Knzyme Assays. Microsomes were prepared from individual rat livers by a calcium precipitation method (29) and were then assayed for testosterone, cyclophosphamide. and ¡fosphamide metabolism. Testosterone 2«-hydroxylase Modulation of Liver Enzyme Patterns by Ifosphamide. Treat ment of adult male rats with ifosphamide or cyclophosphamide led to significant decreases in liver microsomal ifosphamide 4-hydroxylase (Fig. 2A ) and cyclophosphamide 4-hydroxylase activities (see Table 2. below) (9). Since CYP 2C11 is a major catalyst of both ifosphamide (15) and cyclophosphamide (2) activation in adult male rat liver microsomes, these results suggest that ifosphamide suppresses the expression of CYP 2CII, just as cyclophosphumide does (9). We examined this possibility by comparing the effects of these oxazaphosphorines on the levels of CYP 2CII, which is an adult malespecific liver cytochrome P450 enzyme that is suppressed by cyclo phosphamide. and on the cytochrome P450-independent enzyme steroid 5a-reductase. which is a female-predominant enzyme whose levels in the liverare increased by cyclophosphamide treatment. When the drugs were given at equimolar doses (120 mg/kg. single i.p. injection), ifosphamide decreased CYP 2C11-catalyzed hepatic mi crosomal testosterone 2a-hydroxylase activity by ~50%, compared to an ~80<7r decrease by cyclophosphamide. 7 days after drug treatment 2491 Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research. ACROLEIN. PHOSPHORAMIDE (A) IFOSPHAMIDE 4-HYDROXYLASE (B) TESTOSTERONE 2«-HYDROXYUSE CD E c E \ o E c (C)STEROID Sa-REDUCTASE . 5 O < (D) SERUM TESTOSTERONE o I— < I— z O) LJ C O z o Ãœ SALINE IFA 120 IFA 180 CPA 120 mg/kg mg/kg mg/kg MUSTARD. AND GENE EXPRESSION levels (Fig. 3, A and B) substantiated the effects of the drugs on the corresponding enzyme activities. Thus, ifosphamide elicits the same effects as cyclophosphamide on these two hepatic enzymes but re quires a higher dose. The feminization of the gender-dependent hepatic cytochrome P450 enzymes and steroid 5a-reductase by cyclophosphamide treatment of adult male rats is associated with depletion of serum testosterone (9). Therefore, serum testosterone concentrations were compared in ifos phamide- and cyclophosphamide-treated adult male rats. As shown in Fig. 2D. serum testosterone was largely depleted by both drugs, sug gesting that the effects of ifosphamide and cyclophosphamide on CYP 2C11 and steroid 5a-reductase occur by the same mechanism. In Vivo Acrolein Treatment. Cyclophosphamide and ifosphamide are both metabolized to yield two electrophilic metabolites, i.e., acrolein. which has been implicated in protein alkylation, and a mustard, which alkylates DNA. Either of these reactive metabolites could, in principle, mediate the major changes in liver enzyme patterns that follow in vivo treatment with the parent oxazaphosphorine. To address this issue, we first examined the effects of acrolein, since this elec trophilic aldehyde has been shown to bind to cytochrome P450 en zymes in vitro by interacting with cysteine sulfhydryl groups, result ing in protein denaturation (6-8). As shown in Table 1, at a dose of 3 mg/kg (single i.p. injection) acrolein decreased testosterone 2a-hydroxylase activity by ~50% and increased steroid 5a-reductase ac tivity by 1.5-fold in isolated liver microsomes. This corresponds to a feminization ratio (defined as the ratio of microsomal steroid 5areductase activity to microsomal testosterone 2a-hydroxylase activ ity) of only 1.7, compared to a ratio of 19 for the cyclophosphamidetreated group (Table 1). At a higher dose of acrolein (5 mg/kg) testosterone 2a-hydroxylase activity was decreased to a level similar to that observed following cyclophosphamide treatment but steroid 5«-reductase activity was not further increased (Table 1). However, in contrast to cyclophosphamide treatment, severe toxicity (body weight loss) and some lethality occurred with this treatment. Rats treated with acrolein at a higher dose ( 10 mg/kg) died within 24 h after injection. Thus, although acrolein can elicit some of the effects of cyclophos phamide. this response is observed only under conditions of severe toxicity. Effects of Acrolein Precursors. In order to better model the liver metabolism-dependent release of acrolein from 4-hydroxycyclophosphamide that occurs in vivo, we examined the effects of deCl-cyclophosphamide (Fig. 1), which is a cyclophosphamide analogue that yields acrolein enzymatically but without the formation of phosphoramide mustard. Treatment of rats with deCl-cyclophosphamide (200 mg/kg. i.p.; the animals were killed 7 days later) did not alter hepatic CYP 2C11 or steroid 5a-reductase mRNA levels (Fig. 4, A and B). Preliminary experiments showed that /'/; vitro the formation of acrolein from deCl-cyclophosphamide catalyzed by uninduced rat liver mi crosomes occurs with a lower efficiency than does the chemical de Fig. 2. Effect of in vivo ifosphamide and cyclophosphamide treatment on hepatic composition of HPD-cyclophosphamide (Fig. 1). which is another microsomal en/yme activities and serum testosterone levels. Adult male rats were admin cyclophosphamide analogue that yields acrolein but not phosphoraistered single i.p. injections of ifosphamide (IFA ) ( 120 or 180 mg/kg), cyclophosphamide mide mustard. Therefore, we examined the effects of HPD-cyclophos (CPA I ( 120 mg/kg). or saline (control) and were killed 7 days later. Microsomal enzyme activities and serum testosterone levels were determined as described in "Materials and phamide and found that at an i.p. dose of 100. 150, or 200 mg/kg it did Methods." Points, determination for each individual rat; bars, mean values for each not affect hepatic microsomal testosterone 2a-hydroxylase or steroid treatment group. 5a-reductase activity 7 days after treatment (Table 1). Together, these (Fig. 2B). At this dose ifosphamide did not significantly alter hepatic data indicate that neither acrolein nor the acrolein precursors can microsomal steroid 5a-reductase activity, whereas cyclophosphamide mimic the effects of cyclophosphamide with respect to feminization of increased it by ~7-fold (Fig. 2C). In contrast, when ifosphamide was the pattern of hepatic CYP 2CI1 and steroid 5a-reductase enzyme given at a 50% higher dose (180 mg/kg) it suppressed testosterone expression. 2a-hydroxylase activity (Fig. 2B) to the same extent as did cyclo In Vivo Phosphoramide Mustard Treatment. We next examined phosphamide, whereas it increased steroid 5a-reductase activity (Fig. whether phosphoramide mustard might account for the changes in liver enzyme levels observed following oxazaphosphorine treatment. 2C) but to a lesser extent than did cyclophosphamide. Northern blot analysis of hepatic CYP 2C11 mRNA and steroid 5a-reductase mRN A Adult male rats were given single i.p. injections of phosphoramide 2492 Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research. ACROLEIN. PHOSPHORAMIDK MUSTARD. M Fig. 3. Alteration in liver mRNA expression fol lowing treatment with ifosphamide. Total liver RNA was prepared from adult male rats (M ) treated with saline (¡tint's1. 2. and //). ifosphamide (IFA } ( 120 mg/kg. lanes } and 4: 180 mg/kg. lunes 5 and 6). or cychiphosphamide iCPA I ( 120 mg/kg, lanes 7 and K) and killed 7 days later. RNA samples from untreated adult female rat livers (Ft. which do not express the male-specific CYP 2CII hut have high levels of the female-predominant steroid 5o>reductase. are included for comparison (lanes 9 and 10 ). Shown are autoradiograms of a Northern hlot ana lyzing total liver RN'A (one individual liver RNA AND GENE EXPRESSION (120) (180) (120) IFA IFA CPA M 2C11 B. 5ccR sample per lane), probed sequentially with the CYP 2C1I (A) and steroid 5a-reductase (ßlgene-spe cific oligonucleotides (29). Rat a-iubulin mRNA levels (C) are indicative of the RNA load and in tegrity for the samples shown in A and H. Tubulin i mustard at 20-100 mg/kg and then were killed 7 days later. At the lower doses used in the experiments, phosphoramide mustard did not affect CYP 2C11-catalyzed testosterone 2a-hydroxylase activity or steroid 5a-reductase activity (Table I and data not shown), nor did it alter CYP 2C11 mRNA or steroid 5a-reductase mRNA levels (Fig. 4. A and ß).In contrast, phosphoramide mustard at 100 mg/kg decreased testosterone 2a-hydroxylase activity by 90<7cand increased steroid 5a-reductase activity by ~4.5-fold, corresponding to a feminization ratio of 28 (Table 1). Although this dose of phosphoramide mustard altered these enzyme activities in a manner similar to that effected by cyclophosphamide (120 mg/kg). it also conferred general systemic toxicity, as exemplified by major body weight loss (Table 1). In cyclophosphamide-treated rats, body weight typically decreases by ~5-10% during the first 7 days after drug administration but then Table I Effect of in VÕYO treatment with cvcloplutsphantitle, 10 8 11 stabilizes for the remainder of the experimental period (see Table 3). In contrast, a continual decline in body weight occurred during the 7-day observation period in rats treated with the 100 mg/kg dose of phosphoramide mustard (Table I and data not shown). Thus, phos phoramide mustard can modulate liver enzyme activities in a manner similar to that of cyclophosphamide. Kffects of Phosphoramide Mustard and HPD-cyclophosphamide in Combination. To test whether the modulation of the specific hepatic enzymes by cyclophosphamide is achieved through (he com bined action of acrolein and phosphoramide mustard, rats were given i.p. injections of phosphoramide mustard (50 mg/kg). HPD-cyclophosphamide (200 mg/kg). or both agents in combination, and the animals were killed 9 days later. The doses chosen were shown to have minimal gross toxicity (body weight loss) in earlier experiments (Ta- acrolein, HPD-c\cltiphcrsphainide. or phosphoratniilt1 tnusuinl un hepatic tnicnisomal c/i-vmc activities anil hntl\ weight Hepatic microsomes were prepared from adult male rats treated with single i.p. injections of saline (control), cyclophosphamide (CPA). phosphoramide mustard (PM), acrolein. or HPD-cyclophosphamide (HPD-CPA) at the doses indicated and killed 7 days later. Enzyme activities were determined as described in "Materials and Methods." Results are expressed as mean ±SD in cases where the number («)of rats was three or four per group or as mean ±half the range for groups with two rats. 5«-reduclasc activity activity (nmol/min/mg)"1 (nmol/min/mg)"0.88 TreatmentSalineCPAAcroleinHPD-CPAPMn4322"2'32222Dose (mg/kg)1203510100ISO20050100Testosterone2a-hydroxylase in ratio''0.6191.7fi.4NA0.90.51.20.528Change body(g)'+32 weight 0.400.33 .47 ± 0.306.38 ± ±4-21 ±0.130.75 2.581.30 ± 10-3± 70.23NA'1.31 ±0.1 ±0.121.47NA1.12*0.120.55 3-59NA+ ± 41.79±0.1 0.530.97 ± 0.430.82 ± I6±5+ 1+5 I0± ±3+ ±0.090.14 ±0.02Steroid 0.083.94 ± ±0.49Feminization ±3-56 15 ±4 ±0.371.18 ±0.190.83 " Activity expressed as nmol product formed/min/mg protein. '' Ratio of steroid 5a-reductase to testosterone 2a-hydroxylase activity. Typical feminization ratio for untreated adult female rats is >30. 1 Difference between body weights on the day of sacrifice and the day of drug treatment. Of the two rats treated with this dose, one died within 24 h after injection. '' Of the two rats treated with this dose, both died within 24 h after injection. ' NA, not applicable. 2493 Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research. ACROLEIN. F M PHOSPHORAMIDI: MUSTARD. HPD-cyclophosphamide. half responded to the drug combination. In the responsive rats. CYP 2C11-catalyzed microsomal cyclophosphamide hydroxylation and testosterone 2a-hydroxylation were de creased by -45% and -90%, respectively, hepatic CYP 2C11 mRNA <j u g. -a 2C11 became undetectable. and serum testosterone was depleted. This pat tern of response is similar to that observed following cyclophospha mide administration (Table 2 and Fig. 5A). Steroid 5a-reductase ac tivity and mRNA were both increased in the responsive rats but to a lesser extent than in the cyclophosphamide-treated rats (Table 2 and Fig. 5B}. Body weights of the responsive rats declined for 5 days after treatment and then stabilized for the remainder of the experimental period but were still lower than those of the cyclophosphamide-treated rats (Table 3). Overall, these findings demonstrate that, while phos phoramide mustard is responsible for modulation of liver enzyme expression by cyclophosphamide. acrolein potentiates the modulating activity of the mustard. 5aR DISCUSSION A. B. t Previous studies have shown that the decreases in total liver cytochrome P450 content and cytochrome P45()-dependent enzyme activ ities following acute cyclophosphamide treatment of adult male rats (reviewed in Ref. 19) are the result of changes in the levels of specific liver cytochrome P450 enzymes (9). While some cytochromes P450 were found to decrease, others increased substantially following cy clophosphamide administration. The overall effect is to feminize the pattern of expression of liver enzymes in a manner that is similar to. but mechanistically distinct from, that observed with the alkylating agent cisplatin (22, 23). The present study establishes that ifosphamide is also capable of feminizing the expression of these liver en zymes and that the effects occur at a pretranslational step, involving suppression of the male-specific CYP 2C11 mRNA and elevation of the female-dominant steroid 5a-reductase mRNA. The present study also provides insight into the mechanism by which these oxazaphosphorines alter the expression of these mRNAs. Phosphoramide mus tard is shown to be responsible for the modulation of liver enzyme expression by cyclophosphamide. while acrolein potentiates these effects of the mustard. While ifosphamide and cyclophosphamide were both effective in modulating liver enzyme levels, ifosphamide was found to be less potent than cyclophosphamide. Higher doses of ifosphamide are also required to achieve an equivalent plasma alkylating activity in cancer patients (3.8 g/m2 versus 1.1 g/m2 for cyclophosphamide) (34). The c. Tubulin 1 AND GENE EXPRESSION 23456 Fig. 4. Phosphoramide mustard (20 mg/kg) and deCI-cyclophosphamide (200 mg/kg) do not alter hepatic CYP 2CII and steroid 5a-reductase mRNA levels. Adult male rats (M) were given i.p, injections of single doses of saline (lime* 3 and 4), cyclophosphamide (CPA ) ( 120 mg/kg. lane 5 ), phosphoramide mustard (PM ) (20 mg/kg. lane f>). or deClcyclophosphamide ttleCI-CPA} (2(X) mg/kg. lane 7) and were killed 7 days later. Total liver RNA was prepared and Northern blot analysis was performed as in Fig. 3, RNA samples from untreated adult female rat livers (Fi are included for comparison (lanes I and 2). A. CYP 2C11 mRNA: B, steroid 5a-reductase mRNA; C. a-tubulin mRNA (control). ble 1). Neither agent, when administered alone, had major effects on hepatic microsomal enzyme activities, serum testosterone levels (Ta ble 2). or body weight profiles (Table 3). CYP 2C1I mRNA and steroid 5«-reductase mRNA levels were also unaffected (Fig. 5. A and B). Of the 12 rats treated with both phosphoramide mustard and Table 2 Effecl of etnnbineil pliosplturuinitle mustard ami HPD-cvclttphosphantitle treatment on hepatic inicnistirtwl eii'vme adivines antl serum testosterone levels Adult male rats were treated with single i.p. injections of saline (control), cyclophosphamide (CPA) ( 120 mg/kg). phosphoramide mustard (PM) (50 mg/kg). HPD-cyclophosphamide (HPD-CPA) (200 mg/kg). or both phosphoramide mustard and HPD-cyclophosphamide. and the animals were killed 9 days later. Hepatic microsomes were isolated and enzyme activities and serum testosterone concentrations were determined as described in "Materials and Methods." Results are expressed as mean ±SD for the indicated number (in of rats per treatment group. TreatmentSalineCPAPMHPD-CPAPM 4-hydroxylase activity (nmol/min/mg)"4. 2tt-hydroxy]ase activity (nmol/min/mg)"1.53 5a-reductase activity (nmol/min/mg)"0.98 testosterone ratio'10.6370.81.3190.9Serum (ng/ml)'2.6 ±0.48''2.01 12 ±0.300.23 0.488.47 ± 1.9I.I ± ±0.86''4.32 0.041.31 ± 2.081± ±0.53.4 0.663.70 ± ±0.271 0.31 .02 ± ±2.12.8 0.732.33 ± 0.250.15±O.I5 . 11 ± 0.652.79 .49 ± 1.9<0.042.5 ± HPD-CPAResponders + ±0.49 ±1.02 Nonrespondersn13g6666Cyclophosphamide 4.03 ±0.59Testosterone I.I7±O.I8Steroid 1.02 ±0.49Femini/alion " Activity expressed us nmol product formed/min/mg microsomal protein. h Fernini/ution ratio defined us in Tuble I. ' Inlerindividuul variations in serum testosterone are commonly observed in adult male rats due to the intermittent release of testosterone bv the testis. ll n = 6 rats used tor these activity measurements. 2494 Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research. ±1.0 ACROLEIN. Table 3 Effect ofphosphoramide PHOSPHORAMIDE MUSTARD. AND GENE EXPRESSION mustard und HPD-c\clopho\plmniide combination treatment un hody weight Data shown are tor the same groups of adult male rats treated on day 0 that are described in Table 2. Data are expressed as mean ±SD tor the indicated number in) of rats per treatment group. Body weight (g) TreatmentSalineCPAPMHPD-CPAPM (I"199 1201 ±5204 ±6192 ±4205 ±6201 10205 ± 10I90±ll190 ± 5218*8193 ±4205 7226 9235 ±8180 ±8188 ±3217± ±5225 ±7236 12204 13215 ± 15225 ± 16161 ± 13160 ± 13159 ± 10205 ± ±12Day 13215 ± ±11Day 17225 ± ± HPD-CPAResponders + ±6 194±9Day 6207± ±1Day 13" Nonrespondersn986666Day Day after treatment. observed differences in the potency of these two drugs can largely be explained by quantitative differences in their metabolism. Cyclophosphamide is metaboli/ed predominantly at the C4 position of the oxa/aphosphorine ring and side-chain /V-dechloroethylation is minor (<IO%) (I, 18. 35), whereas in the case of ifosphamide side-chain metabolism accounts for an estimated 5()9r of the administered dose (16). Therefore, given the same dose, ifosphamide generates less mustard and less acrolein than does cyclophosphamide. The suppression by cyclophosphamide and ifosphamide of hepatic CYP 2CII occurred at a pretranslational step. It is likely that CYP 2C1I transcription is the step that is affected in oxazaphosphorinetreated rats, because the male-specific expression of the CYP 2C1I gene is regulated at the level of transcript initiation in adult rats (36). The underlying causes for these effects on CYP 2C11 and steroid 5a-reductase levels could involve effects on one or more of the hypothalamo-pituitary and gonadal factors that regulate expression of these genes (37). rather than direct effects on the liver. Indeed, the suppression of CYP 2CI1 mRNA and the elevation of steroid 5areductase mRNA by cyclophosphamide and ifosphamide. or by the combination of HPD-cyclophosphamide and phosphoramide mustard. M is shown to be associated with a substantial decrease in serum levels of testosterone, which is required for maintenance of the sexually dimorphic expression of these enzymes (38—40).However, while en dogenous androgen secretion in cyclophosphamide-treated rats can be stimulated by the luteinizing hormone analogue chorionic gonadotropin. the resultant increase in serum testosterone does not reverse the suppression of hepatic CYP 2C11 (9). This observation is analogous to the finding that the suppression of CYP 2CII by 3.4.5,3'.4',5'hexachlorobiphenyl is also not causally related to the associated de pletion of serum testosterone (41 ). Consequently, modulation of liver enzyme expression by cyclophosphamide and ifosphamide may in volve action at the hypothalamo-pituitary axis, which establishes the sex-dependent plasma growth hormone profile that in turn determines the expression of hepatic CYP 2C11, steroid 5a-reductase, and other drug- and steroid-metaboli/.ing enzymes in adult male rats (37, 42, 43). The present study demonstrates that, while phosphoramide mustard is primarily responsible for the modulation of hepatic CYP 2C11 and steroid 5a-reductase by cyclophosphamide. acrolein potentiates the modulating activity of the mustard. When administered alone, neither F CPA PM HPD-Cm PM+HPD-CPA •¿â€¢ Fig. 5. Effect of phosphoramide mustard und HPD-cyclophosphamide combination treatment on hepatic CYP 2C11 and steroid So-rcductase mRNA levels. Adult male rats (M\ were treated with saline (lanes I and 2}, cyclophosphamide (CPA} (120 mg/kg. lanes 5 and 6). phosphoramide mustard (PM] (50 mg/kg. lanes 7-V). HPD-cyclophospha mide (HPD-CPA} (200 mg/kg. lanes 10-12). or a combination ol phosphoramide mustard (50 mg/kg) plus HPD-cyclophosphamide (2(K) mg/kg) (lanes 13-16) and were killed 9 days later. Total liver RNA was prepared and Northern blot analysis was performed as in Fig. 3. RNA samples from un treated adult female rat livers (/•") are included for - 2C11 B. 5aR comparison (lanes 3 and 4). A. CYP2C1I mRNA; B, steroid 5a-reductase mRNA; C. u-tubulin mRNA (control). The reduced signal in ¡tine5 for tubulin mRNA indicates that the RNA analyzed in this lane was underloaded, compared to the other samples. Tubulin 123456 8 9 10 11 12 13 14 15 16 2495 Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research. ACROLEIN. PHOSPHORAMIDE MUSTARD. acrolein nor acrolein precursors (HPD-cyclophosphamide and deClcyclophosphamide) feminized the expression of these enzymes to the same extent as did cyclophosphamide. In contrast, phosphoramide mustard altered the expression of CYP 2C11 and steroid So-reductase, similarly to cyclophosphamide. hut this effect was achieved at a dose of mustard ( KM)mg/kg) that produced greater body weight loss than did cyclophosphamide. This difference in toxicity might be due to differences in the pharmacokinetics and tissue distribution of phos phoramide mustard given as a bolus i.p. injection, compared to phos phoramide mustard derived from chemical decomposition of the pri mary 4-hydroxy metabolite in cyclophosphamide-treated rats. Whereas treatment with a lower dose of phosphoramide mustard (50 mg/kg) or with the acrolein precursor HPD-cyclophosphamide (200 mg/kg) alone did not affect liver enzyme profiles, the combination of these two agents suppressed CYP 2CI1 and elevated steroid 5areductase enzyme activities and mRNA levels, in addition to depleting serum testosterone, an overall pattern that is similar to that produced by cyclophosphamide. These effects of the combination treatment were observed in only half of the animals tested and suggest that a concentration threshold exists at the site(s) of action of these metab olites to elicit the modulation of liver enzyme expression. Such a threshold has been documented for acetaminophen-induced hepatic necrosis (44). The glutathione-depleting agent buthionine sulfoximine also appears to potentiate phosphoramide mustard-mediated cardiotoxicity in rodents (28). Acrolein or cyclophosphamide treatment of rats can decrease cellular glutathione content, whereas phosphora mide mustard is only somewhat effective at glutathione depletion when given at high doses (28, 45). Thus, acrolein may render target cells more sensitive to the modulating activity of phosphoramide mustard by reducing intracellular glutathione levels. As noted above, the effects of cyclophosphamide and the combination of phosphora mide mustard and HPD-cyclophosphamide on liver mRNA levels indicate that these sensitizing effects of acrolein probably occur at endocrine secretory organs, rather than the liver. Glutathione is found not only in liver but also in many other tissues, including brain (46). Cyclophosphamide and ifosphamide are typically administered to cancer patients as part of a combination chemotherapy regimen. The present study indicates that these alkylating agent prodrugs not only can alter their own biotransformation but also may affect cytochrome P450-mediated bioactivation or deactivation of concurrently admin istered drugs. Clinical studies have shown that chronic administration of oxazaphosphorines to cancer patients can increase drug clearance (14, 18), suggesting that human liver cytochrome P450 enzymes are also subject to modulation by these oxazaphosphorines, albeit by mechanisms that lead to increased rates of drug metabolism. Since our studies implicate phosphoramide mustard as the primary mediator of the effects of the parent drug on liver drug metabolism, it may not be possible to design active oxazaphosphorines that lack these potential drug interactions. Finally, the finding that acrolein potentiates the enzyme-modulating effects of phosphoramide mustard has a broader implication, insofar as environmental exposure to acrolein may trigger or exacerbate drug- and xenobiotic agent-induced systemic toxicity. Indeed, significant levels of acrolein are found in tobacco smoke (47). tication of the hepatic monooxygenase catalysts of drut! activation. Cancer Res., 4V: 2344-2350. 1989. Colvin. M., and Hilton. J. Pharmacology of cyclophosphamide and metabolites. Cancer Treat. Rep.. 65 (Suppl. 3): 89-95. 1981. Brock. N. Comparative pharmacologie study in vitro and m vivo with cyclophospha mide (NSC-26271 ), cyclophosphamide metabolites, and plain nitrogen mustard com pounds. Cancer Treat. Rep.. 60: 301-307, 1976. Wrabetz, E.. Peter. G., und Hohorst. H. J. Does acrolein contribute to the cytotoxicity of cyclophosphamide1' J Cancer Res. Clin. Oncol.. 9«.-119-126. 1980. 23. 24. 26. 27. 28. 29. ACKNOWLEDGMENTS The authors wish to thank Dr. J. Pohl (ASTA Pharma. Bielefeld. Germany) and Dr. J. Hilton (Johns Hopkins Oncology Centre. Baltimore. MD) for kindly providing the acrolein precursors used in this study. 30. 31. 32. REFERENCES 1. Sladek. N. E. Metabolism of oxazaphosphorines. Pharmacol. Then. $1: 301-355, 1988. 2. Clarke. L.. and Waxman. D. J. Oxidative metabolism of cyclophosphamide: identi- AND GENE EXPRESSION 33. Mannello. A. J., Gunoo. H. L.. Struck. R. F.. and Paul. B. Denaturation of cytochrome P450 by cyclophosphamide metabolites. Biochem. Biophys. Res. Commun.. 83: 1347-1353. 1978. Gurtoo. H. L.. Mannello. A. J.. Struck. R. F.. Paul. B.. and Dahms. R. P. Studies on the mechanism of denaturation of cytochrome P450 by cyclophosphamide and its metabolites. J. Biol. Chem.. 256: 11691-11701. 1981. Mannello. A. J.. Bansal. S. K., Paul. B.. Koser. P. L.. Love, J., Struck, R. F.. and Gurtoo. H. L. Metabolism and binding of cyclophosphamide and its metabolite acrolein to rat hepatic microsomal cytochrome P450. Cancer Res.. 44: 4615—4621. 1984. LeBlanc. G. A., and Waxman. D. J. Mechanisms of cyclophosphamide action on hepatic P-450 expression. Cancer Res., 50: 5720-5726. 1990. Mannello. A. J.. Berrigan. M. J.. Struck. R. F., Guengerich. F. P.. and Gurtoo, H. L. Inhibition of NADPH-cytochrome P450 reducÃ-aseby cyclophosphamide and its me tabolites. Biochem. Biophys. Res. Commun.. 99: 399-406. 1981. Manthey. C. L.. Landkamer. G. J.. and Sladek. N. E. Identification of the mouse aldehyde dehydrogenases important in aldophosphamide detoxification. Cancer Res.. SO: 4991-5002. 1990. Dockham. P. A.. Lee. M-O.. and Sladek. N. E. Identification of human liver aldehyde dehydrogenases that catalyze the oxidation of aldophosphamide and retinaldehyde. Biochem. Pharmacol.. 43: 2453-2469. 1992. Brock. N. Oxazaphosphorine cytostatics: past-present-future. Cancer Res.. 49: 1-7, 1989. Allen. L. M.. Creaven. P. J.. and Nelson, R. L. Studies on the human pharmacokinetics of ¡sophosphamide (NSC-109724). Cancer Treat. Rep.. 60: 451-458. 1976. Weber. G. F.. and Waxman. D. J. Activation of the anti-cancer drug ifosphamide by rat liver microsomal P450 enzymes. Biochem. Pharmacol., in press, 1993. Norpoth. K. Studies on the metabolism of isophosphamide (NSC-109724) in man. Cancer Treat. Rep., 60: 437-443, 1976. Goren. M. P.. Wright, R. K.. Pratt, C. B., and Pell, F. E. Dechloroethylation of ifosfamide and neurotoxicity. Lancet. 2: 1219-1220. 1986. Moore. M. J. Clinical pharmacokinetics of cyclophosphamide. Clin. Pharmacokinet.. 20: 194-208. 1991. LeBlanc. G. A., and Waxman. D. J. Interaction of anticancer drugs with hepatic monooxygenase enzymes. Drug Metab. Rev.. 20: 395-439. 1989. Berrigan. M. J.. Gurtoo. H. L.. Sharma, S. D.. Struck, R. F.. and Mannello. A. J. Protection by iV-acelylcysteine of cyclophosphamide metabolism-related in vivo de pression of mixed function oxygenase activity and in vitro denaturation of cytochrome P450. Biochem. Biophys. Res. Commun., 93: 797-803. 1980. McClure. M. T.. and Stupans. I. Investigation of the mechanism by which cyclophos phamide alters cytochrome P450 in male rats. Biochem. Pharmacol.. 43: 2655-2658. 1992. LeBlanc. G. A., and Waxman, D. J. Feminization of rat hepatic P-450 expression by cisplatin. Evidence for perturbations in the hormonal regulation of steroid-metabo lizing enzymes. J. Biol. Chem.. 263: 15732-15739, 1988. LeBlanc. G. A.. Sundseth, S. S.. Weber. G. F.. and Waxman. D. J. Platinum anticancer drugs modulate P-450 mRNA levels and differentially alter hepatic drug and steroid hormone metabolism in male and female rats. Cancer Res.. 52: 540-547. 1992. Waxman. D. J.. Sundseth. S. S.. Srivastava. P. K.. and Lapenson. D. R Gene-specific oligonucleotide probes for a. u. TT,and microsomal rat glutathione 5-transferases: analysis of liver transferase expression and its modulation by hepatic enzyme inducers and platinum anticancer drugs. Cancer Res., 52: 5797-5802. 1992. Brock. N.. Stekar. J.. Pohl. J.. Niemeyer, L'.. and Scheffler. G. Acrolein. the causative factor of urotoxic side-effects of cyclophosphamide. ifosfamide. trofosfamide and sufosfamide. Arzneim.-Forsch.. 29: 659-661. 1979. Cox. P. J. Cyclophosphamide cystitis: identification of acrolein as the causative agent. Biochem. Pharmacol.. 2H: 2045-2049. 1979. Plowchalk, D. R.. and Madison. D. R. Phosphoramide mustard is responsible for the ovarian toxicity of cyclophosphamide. Toxicol. Appi. Pharmacol.. 107: 472—481. 1991. Friedman. H. S.. Colvin. O. M.. Aisaka. K.. Popp. J.. Bossen, E. H.. Reimer, K. A., Powell, J. B., Hilton. J.. Gross, S. S„Levi, R., Bigner. D. D., and Griffith, O. W. Glutathione protects cardiac and skeletal muscle from cyclophosphamide-induced toxicity. Cancer Res.. 50: 2455-2462. 1990. Waxman. D. J. Rat hepatic P450IIA and P450IIC subfamily expression using cata lytic, immunochemical. and molecular probes. Methods Enzymol.. 206: 249-267. Õ99I. Waxman. D. J. P450-catalyzed steroid hydroxylation: assay and product identification by thin-layer chromatography. Methods Enzymol., 206: 462—476. 1991. Masurel. D.. Houghton. P. J.. Young. C. L.. and Wainer. I. W. Efficacy, toxicity. pharmacokinetics. and in vitro metabolism of the cnantiomers of ifosfamide in mice. Cancer Res.. 50: 252-255. 1990. Sladek. N. E.. Smith. P. C.. Bratt. P. M., Low. J. E.. Powers. J. F.. Borch. R. F.. and Coveney. J. R. Influence of diuretics on urinary general base catalytic activity and cyclophosphamide-induced bladder toxicity. Cancer Treat. Rep.. 66: 1889-1900, 1982. Ram. P. A., and Waxman. D. J. Pretranslational control by thyroid hormone of rat liver 2496 Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research. ACROl.MIN. PHOSl'HOKAMIDI-: 34. 35. 36. 37. 38. 39. 40. 41. MUSTARD. steroid 5«-reductaseand comparison to the thyroid dependence of two growth hor mone-regulated CYP2C mRNAs. J. Biol. Chem.. 265: I9223-I9229. 1990. Creaven, P.J.. Allen. L. M. Alford. D. A., and Cohen. M. H. Clinical pharmacology ot isophosphamide. Clin. Pharmacol. Ther.. 16: 77-S6. 1974. Colvin. M. The comparative pharmacology of cyclophosphamide and ifosfamide. Semin. Oncol.. 9 (Suppl. I).' 2-7. 1982. Sundseth. S. S.. Alhena. J. A., and Waxman. D. J. Sex-specific, growth hormoneregulated transcription ot the cytochrome P450 2C11 and 2CI2 genes.J. Biol. Chem., 267: 3907-3914. 1992. Waxman. D. J. Regulation of liver-specific steroid metabolizing cylochromes P450: cholesterol 7«-hydroxylase.hile acid 6ß-hydroxylase.and growth hormone-respon sive steroid hormone hydroxylases. J. Steroid Biochem. Mol. Biol.. 41: 1055-1072, 1992. Waxman. D. J.. Dannan. G. A., and Guengerich. F. P. Regulation of rat hepatic cytochrome P450: age-dependent expression, hormonal imprinting and xenohiotic inducihility ot sex-specific isoen/ymes. Bii>chemistry. 24: 4409-4417. 1985. Shimada. M.. Murayama. N.. Yama/tH.1.Y.. Kamataki. T.. and Kalo. R. Further studies on the persistenceof neonatal androgen imprinting on sex-specific cytochrome P-450. testosterone and drug oxidations. Jpn. J. Pharmacol.. 45: 467—478.19X7. Waxman. D. J. Interactions of hepatic cytochromes P-450 with steroid hormones. Regioselectivity and stcreospeciticily of steroid melaholism and hormonal regulation of rat P-450 enzyme expression. Biochem. Pharmacol.. J7: 71-84. 1988. Yeowell, H. N.. Waxman. D. J.. I.eBlanc. G. A.. Linko. P.. and Goldstein. J. A. Suppression of male-specific cytochrome P450 2c and its mRNA by 3.4.5.3'.4',5'hcxachlorohiphenyl in rat liver is not causally related to changes in serum testoster AND GENE EXPRESSION one. Arch. Biochem. Biophys.. 271: 508-514. 1989. 42. Zaphiropoulos. P. G.. Mode. A.. Norstedt. G.. and Gustal'sson. J. A. Regulation of sexual differentiation in drug and steroid melaholism. Trends Pharmacol. Sei., lü: 149-153. 1989. 43. Waxman. D. J.. Pampori. N. A.. Ram. P. A.. Agrawal. A. K.. and Shapiro. B. H. Interpulse interval in circulating growth hormone patterns regulates sexually dimor phic expression of hepatic cytochromc P450. Prix-. Nati. Aead. Sci. USA, XH: 68686872. 1991. 44. Mitchell. J. R.. Jollow. D. J.. Potter. W. Z.. Gillette. J. R.. and Brodie. B. B. Acetaminophen-induced hepatic necrosis. IV. Protective role of glulalhione. J. Pharmacol. Exp. Ther.. 1X7: 211-221. 1973. 45. Ciurtoo. H. L.. Hipkens. J. H.. and Sharma. S. I). Role of glutathione in the metab olism-dependent toxicity and chemotherapy ol cyclophosphamide. Cancer Res.. 41: 3584-3591. 1981. 46. Jain. A.. Martensson. J.. Stole. E.. Auld. P. A. M.. and Meister. A. Glutathione deficiency leads to mitochondria! damage in brain. Proc. Nail. Acad. Sci. USA. jfi: 1913-1917. 1991. 47. Beauchamp. R. O.. Jr.. Andjelkovich. D. A.. Kligerman. A. D.. Morgan. K. T., and Heck. H. D. A critical review of the literature on acrolein toxicity. Crit. Rev. Toxicol.. 14: 309-3X0. 1985. 48. Nelson. D. R.. Kamataki. T.. Waxman. D. J.. Guengerich. F. P.. Eslabrook. R. W.. Feyereisen. R.. Gonzalez. F. J.. Coon. M. J.. Gunsalus. 1. C.. Gotoh. O.. Okuda. K.. and Neben. D. W. The P450 superfamily: update on new sequences,gene mapping, accession numbers, early trivial names of enzymes, and nomenclature. DNA Cell Biol.. 12: 1-51. 1993. 2497 Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research. Cyclophosphamide Modulates Rat Hepatic Cytochrome P450 2C11 and Steroid 5 α-Reductase Activity and Messenger RNA Levels through the Combined Action of Acrolein and Phosphoramide Mustard Thomas K. H. Chang and David J. Waxman Cancer Res 1993;53:2490-2497. Updated version E-mail alerts Reprints and Subscriptions Permissions Access the most recent version of this article at: http://cancerres.aacrjournals.org/content/53/11/2490 Sign up to receive free email-alerts related to this article or journal. To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at [email protected]. To request permission to re-use all or part of this article, contact the AACR Publications Department at [email protected]. Downloaded from cancerres.aacrjournals.org on April 30, 2017. © 1993 American Association for Cancer Research.