Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Adaptive immune system wikipedia , lookup
DNA vaccination wikipedia , lookup
Molecular mimicry wikipedia , lookup
Complement system wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
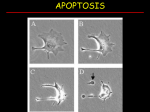
DEGRADATION OF THE EXTRACELLULAR MATRIX Tissue remodeling - CT and ECM composed of collagens/proteoglycans - tissue remodeling CT is hydrolyzes by specific proteases and resynthesized by fibroblasts; proteases include serine proteases and metalloproteinases Serine proteases - used in blood clotting - uPA urokinase-like plasminogen activator proteolitically activates the plasminogen zymogen plasmin - plasminogen is a serum protein but is also present in all tissues (especially if blood vessel endothelium is interrupted) - activated plasmin is a potent and non-specific protease that will hydrolyze any protein in its path (including self) - when not bound to substrate plasmin is inactivated by serine protease inhibitor (serpin), alpha2-anitplasmin - when bound to substrate (fibrin, collagen) serpins less active - cells in remodeling tissues express receptor for uPA, so that they carry this protease activator on cell surface can move through the lattice of the ECM easily Matrix metalloproteinases (MMPs) - used in directed remodeling of the ECM - includes specific collagenases and non-specific gelatinases and stromelysins - collagenases cleave specific collagens at specific sites once cleaved they become substrates for gelatinases and stromelysins which degrade the collagens into smaller peptides which are internalized by invading cells where they are completely degraded to lysosomal proteases called cathepsins - 5 smaller families in MMP superfamily: collagenases, gelatinases, stromelysins, membrane associated MMPs, and others; structurally distinct categories grouped according to structural domains - MMPs contain a signal sequence (for secretion), an inhibitory pro-sequence (because they are secreted as zymogens), a catalytic domain (may contain a gelatin-binding domain), a catalytically active Zn2+ ion, a hinge region, a hemopexin domain (for interaction with inhibitors), and a transmembrane domain Proteases in cancer metastasis - cancer progression includes transition from benign malignant - most advanced state of cancer prior to malignancy is a carcinoma in situ cancer still contained within the parenchyma of tissue; however carcinoma cells in situ are triggered to secrete proteases and increase their migration and invasion - proteases degrade the ECM and the basement membrane that separate tissue parenchyma from the blood supply/lymphatics this is knows as extravasation motile tumor cells shed into blood/lymph where they colonize distant sites when cells become lodged in vasculature - shedding requires matrix-degrading proteases - Urokinase-like plasminogen activator – Serine protease – Attached to invasive cell surface. – Activates tissue plasminogen to plasmin, which is not specific. - Matrix metalloproteinases – important in tissue remodeling - Cathepsins – lysosomal proteases, most contain cysteine in active site and cleave proteins through redox; 20 cathepsins A-Z; distinct from MMPs in that they are active at acidic pH and restricted to lysosomes - cathepsins may be secreted where they are active in acid pH environments of tumors - cathepsins B,D, and L associated with metastatic progression in colon, prostate, and breast cancers - Treatment of cancers with protease inhibitors, Serine protease inhibitors (Serpins) or Tissue Inhibitors of Metalloproteinases (TIMPs), is moderately effective in inhibiting metastasis - Effectiveness is limited due to redundancy in the system; multiple proteases are secreted by metastatic cancer cells and thus, specific inhibition of only one system is ineffective in arresting the whole process. INTRACELLULAR PROTEASES AND PROGRAMMED CELL DEATH (APOPTOSIS) - important in tissue remodeling is removal of cells no longer needed; occurs naturally during maturation of the immune system, morphogenesis, nervous system development, and in response to cell damage - lymphocytes coding for self-recognition are removed; mediated by specific ligands Fas-ligand (FasL) which bind to specific receptors (Fas) induce intracellular events that lead to selfdestruction - programmed cell death mediated by proteases is called apoptosis Apoptosis Necrosis • Cellular condensation • Cellular swelling • Membranes remain intact • Membranes are broken • Requires ATP • ATP is depleted (lack of cellular energy • Cell is phagocytosed, no tissue reaction induces necrosis) • Ladder-like DNA fragmentation • Cell lyses, eliciting an inflammatory • In vivo, individual cells appear affected reaction • DNA fragmentation is random, or smeared • In vivo, whole areas of the tissue are affected Death ligands and receptors - death receptors are a family of cell surface receptors that signal for apoptosis when bound by their ligands; ligand binding oligomerization of receptors initiation for signaling leading to cell death - immune privilege: immune cells express death receptors, while immune privileged tissues produce ligand; when immune cells enter these tissues signaling from bound receptors causes immune cells to die via apoptosis (harmful consequences of immune reaction are avoided) - human members of the death receptor family include CD95 (a.k.a. Fas or Apo1), tumor necrosis factor-receptor (TNF-R1), DR3 (Apo2), DR4 (TRAIL-R1), DR5 and DR6 - these are all membrane-bound proteins with an extracellular ligand-binding domain and an intracellular transduction domain, separated by a single TM domain - cytoplasmic portion of all death receptors contains a region of homology called death domain (DD) which allows for protein:protein interaction with cytoplasmic adaptor proteins - two of these adapters are the Fas-associated death domain containing protein FADD and TNF-R-associated death domain contain protein (TRADD); adapter have death domains and contain a death effector domain (DED), which mediates recruitment of proteases that execute death program (caspases) - assembly of proteins from receptor caspases is called death-induced signaling complex (DISC) - Death ligand-induced apoptosis plays major role in immune maturation (remove cells that recognize self), development of nervous system (removes neurons that don’t form functional synapses) and in morphogenesis (removes skin webs between fingers/toes) Caspases - FADD/Mort-1 binds to activates caspase-8 (extrinsic pathway) protease that initiates cell death - caspases are intracellular proteases that contain a cysteine residue in active site and cleave substrates C-terminal to specific aspartate residues in substrates Cysteine Aspartyl-specific proteases (same family as cathepsins) - specificity determined by 4 amino acid residues N-terminal to the cleavage site - highly conserved throughout evolution; 12 caspases found in humans; 8 function in apoptosis - three categories of caspases based on function 1.) Group 1 caspases process cytokines and don’t regulate cell death 2.) Group 2/3 caspases involved in apoptosis; involved in either initiating apoptosis program (caspases 8 10, group III) or involved in propagation or effecting program (caspases 2,3,7 – group II) - enzymes synthesized are in inactive, zymogen form; activated through proteolysis (selfproteolysis) - mature enzyme is a tetramer of two p20 and two p10 domains with two active sites - caspases-8 (initiator caspases involved in Fas signaling) is activated through the scaffold provided by the activated receptor and its associated FADD proteins - pro-caspase-8 proteins (inactive) self-activate through low activity of the zymogen form; similar to HIV protease and clotting cascade (high concentration in restricted proximity self-activation) - apoptotic signal activation of group III initiator caspases (8,9, 10) activation of group II initiator caspases (2,3,7) dismantling of cell into apoptotic bodies - once activated effector caspases go through cell and dismantle substrate proteins following a specific pattern - caspase-mediated protein surgery occurs at specific sites and results in inactivation of target proteins - when caspases are substrates it can lead to activation by cleaving off inhibitory domain - activated substrate caspase-activated DNAse (CAD) that pre-exists in an inactive complex with an inhibitory subunit, ICAD; caspases-3 cleaves ICAD activation of CAD cuts dsDNA every 200 nucleotides (or between histone bundles); DNA ladder that results is used as hallmark for apoptosis; multiple free ends of DNA provide increased substrates for 5’ terminal kinase - 100 different caspases substrates identified all play a role in apoptotic cascade - final result is fragmentation of DNA, disintegration of nucleus and blebbing of membranes into apoptotic bodies which are internalized by macrophages - apoptosis does not elicit immune reaction; mediated through exhibition of phophatidyl serine (PtdSer) on cell surface this phospholipid is in the inner leaflet of the plasma membrane but during apoptosis is exposed this serves as a signal through a protein called Annexin V (its binding is used for apoptosis assay), for macrophages to consume the apoptotic bodies but not to release cytokines and elicit a major immune response - necrosis associated with edema and immune involvement - caspase activation through proteolytic cleavage is mediated by activate caspases working on effector caspases 2,3,6, and 7 - caspases activation through induced proximity is used to activate the initiator caspases, caspase-8, through its associations with FADD or TRADD on the inside surface of the activated death receptors; when death receptors are activated they from stable trimers; this complex of three FADD/TRADD domain serves as a scaffold to induce pro-caspases into close proximity Caspase-9 and Apaf-1 - initiator caspase-9 only modestly activated by proteolytic processing; the key requirement for caspase-9 activation is an association with a dedicated protein cofactor, Apaf-1 - Apaf-1 is one of two proteins required for caspase-9 activation (other is cytochrome c from mitochondria) - active form of caspase-9 is an ATP-dependent oligomerized complex formed between Apaf-1, cytochrome c, and caspase-9; intrinsic pathway - entire complex in active form holoenzymes (apoptosome) - cytochrome c release from mitochondria is a universal feature of apoptosis arising from DNA damage (p53) - apoptosis can be induced either by specific death inducing ligands or through DNA damage - specific death inducing ligands are used in a regulated fashion during development and for immune privilege in adults - DNA damage not regulated and occurs in response to ionizing radiation or chemotherapy - in either case, these signals impinge upon initiator caspase (caspases-8 and -9) which are activated via induced proximity or holoenzymes complex assembly - these initiator caspases then activate effector caspases (effector caspase-3, produces active caspase-6) through proteolytic activation as the first in a cascade of proteolysis that leads to cell degradation 3 mechanisms of caspase activation • Direct proteolytic cleavage that releases p10 and p20 domains from, e.g. pro-caspase 3. Active form has two p20 and two p10 in tetramer. • Induced proximity, such as the binding of caspase 8 to receptor-associated FADD and DED domains, releasing p10/p20 domains. • Oligomerization, as in the case of caspase 9 associating with Apaf-1 and cytochrome c to form holoenzyme